



-
植物作为固着生物,需要应对干旱、盐碱、低温、高温以及病虫害侵袭等多种逆境胁迫,而对胁迫信号的感知和转导是植物应对环境胁迫的前提和基础[1]。研究表明,植物体内胁迫信号的转导需要多种信号转导物质参与,如ABA、乙烯、H2O2和NO等激素和小分子化合物[2]以及信号转导蛋白G蛋白[3]、磷脂酶A[4]、磷脂酶C[5]、磷脂酶D(PLD)[6]等激酶,其中,PLD能够水解磷酸二脂键使细胞膜磷脂产生磷脂酸(PA)和可溶性头基,而PA是多种激素信号和胁迫信号转导过程的第二信使,在信号转导、膜迁移、细胞程序性死亡和细胞骨架重排等多种细胞活动中发挥着重要作用[7-10]。研究表明,PLD的典型特征是具有2个间隔250~400个氨基酸的HKD(HxKxxxxD)结构域,虽然2个HKD在蛋白初级结构上相距较远,但它们在三级结构上能形成互作,且这种互作也是其酶活性发挥的关键[11-15]。根据氨基酸序列特征,PLD可分为α、β、γ、δ、ε、ζ、φ 7种类型,其中,PLDα、PLDγ、PLDδ和PLDζ数量较多,PLDβ和PLDε数量较少,而PLDφ发现较晚且较少见[13, 16]。根据蛋白的N端结构特征和其酶活性对Ca2+的依赖性,PLD也可分为C2-PLD、PX/PH-PLD和SP-PLD 3个亚族,其中,C2-PLD蛋白的N端含有一个高度保守Ca2+依赖型磷脂结合C2结构域,需要结合Ca2+以维持酶活性[12];PX/PH-PLD的N端特征是一个串连的PX(phox homology)和PH(pleckstrin homology)结构域,活性不依赖于Ca2+[13];SP-PLD亚家族规模较小,于2007年在水稻(Oryza sativa L.)中被首次发现,其N端含有信号肽,而不是C2、PX或PH结构域[13]。其中,PLDα、PLDβ、PLDγ、PLDδ和PLDε属于C2-PLD亚家族,PLDζ属于PX/PH-PLD亚家族,PLDφ属于SP-PLD亚家族[11]。植物中,首个PLD于1994年从蓖麻(Ricinus Communis L.)[17]中克隆出来后,随后水稻[18]、拟南芥(Arabidopsis thaliana L.)[19]、玉米(Zea mays L.)以及烟草(Nicotiana tabacum L.)[20]等多个物种中相继被克隆出PLD。随着多个植物全基因组序列的发布[21-27],人们发现PLD在植物中多以基因家族形式存在,如PLD家族成员拟南芥有12个[12]、水稻有17个[13]、白菜(Brassica rapa Lour.)有18个[14]、亚洲棉(Gossypium arboreum L.)有19个[11]、葡萄(Vitis vinifera L.)有11个[28]、大豆(Glycine max L.)有18个[15]等。
研究发现,PLD在植物响应逆境胁迫反应中有着重要作用,如在拟南芥中,AtPLDα1的沉默会严重减弱植株由ABA诱导的耐旱反应[29];Atpldα1缺陷型株系中由创伤诱导的 JA合成和相关基因的表达显著降低[30];Atpldδ缺陷型株系对H2O2胁迫的敏感性显著上升[31];AtPLDδ在响应真菌(Blumeria graminis DC.)胁迫的防御信号转导过程中有着关键作用, Atpldδ突变体株系中响应微生物的早期应激响应基因的上调表达被显著延迟[32]; C2-PLD突变体株系在响应缺氧胁迫过程中关键信号转导物质PA和胞质钙的含量上升显著受阻[33];在其他植物中,如在毛白杨(Populus tomentosa Carr.in Rev.Hort.)中过表达AtPLDα1能够显著提高其耐盐性与耐旱性[34];过表达黄瓜(Cucumis sativus L.)CsPLDα的烟草抗旱性增强[35];小麦(Triticum aestivum L.)TaPLDα能够响应多种胁迫下表达上调,过表达TaPLDα的拟南芥耐旱能力显著增强[36]。由此可见,PLD家族在植物应对逆境胁迫时具有重要生物学功能。
毛果杨(Populus trichocarpa Torr. & Gray)是首个全基因组测序的木本植物[37],也是研究木本植物生长发育、材质材性、逆境胁迫响应和其他重要性状的模式植物。然而,目前为止,尚无毛果杨PLD家族鉴定及其在逆境胁迫响应特性的研究报道。本项研究在全基因组水平鉴定出毛果杨PLD家族全部成员的基础上,对其系统进化、同源关系、选择压力在其家族进化中的作用、基因结构、家族各基因的顺式作用元件和蛋白保守基序进行分析,从而全面阐述该基因家族的基本特征。此外,利用实时荧光定量PCR和已报道的毛果杨转录组数据,对其PLD家族基因的组织表达特性以及盐胁迫的响应特性进行分析。本研究为进一步阐明毛果杨PLD家族基因的生物学功能和抗逆基因的挖掘奠定了基础。
-
利用报道的拟南芥PLD的氨基酸序列比对Phytozome网站(
https://phytozome.jgi.doe.gov )中的毛果杨基因组数据库[12],同时在毛果杨数据库检索已注释的PLD,将获得的序列结果汇总并剔除重复序列后作为候选基因。为了验证初始结果的可靠性,将候选基因的氨基酸序列上传至HMMER网站 (https://www.ebi.ac.uk/Tools/hmmer )以及NCBI保守结构域数据库(https://www.ncbi.nlm.nih.gov/cdd/ )鉴定保守结构域,以含有2个间隔250~400氨基酸的PLD典型HKD结构域(HMM模型登录号为PF00614)为标准进行筛选,并根据保守结构域鉴定蛋白类型,最终鉴定出毛果杨PLD家族全部成员[11, 14, 38-40]。从Phytozome网站中获得PtrPLD家族成员的染色体位置、基因序列和开放阅读框长度等信息,并根据基因编码蛋白类型和进化关系进行命名,染色体位置信息经Tbtools软件进行可视化处理[41]。通过ExPasy(
https://web.expasy.org )在线预测其等电点与分子质量[42],通过Plant-mPLoc(http://www.csbio.sjtu.edu.cn/ )在线预测其亚细胞定位位置[43]。 -
将鉴定出的毛果杨全部PtrPLDs以及拟南芥AtPLDs编码的氨基酸序列在MEGA X软件的ClustaW程序中进行多重序列比对[44]。采用邻近法(Neighbor-joining)构建系统进化树,其中,bootstrap设置1000次重复,再经最大似然法(Maximum-likeihood)验证,得到系统发育进化树数据,经Evoview(
https://www.evolgenius.info/evolview/ )网站可视化处理[12, 44-45]。 -
Blast相互比对PtrPLDs的CDS序列,超过300个bp且同源性超过80%为标准确定为同源基因对[46-47],同源关系经Tbtools软件可视化处理[41]。利用Tbtools软件计算同源基因对之间的同义替换率(Ks)、非同义替换率(Ka)以及Ka/Ks比率,并以此分析PtrPLD家族基因进化过程中的选择压力[41, 46, 48]。
-
从毛果杨数据库(
https://genome.jgi.doe.gov/portal/pages/dynamicOrganismDownload.jsf?organism=Ptrichocarpa )下载基因组数据后,通过TBtools软件提取各PtrPLD外显子、内含子长度及位置信息并进行可视化处理[41]。将各PtrPLD氨基酸序列提交到Motif Elicitation 网站的MEME程序(
http://meme-suite.org/tools/meme )中分析保守基序并进行可视化处理[41, 49]。 -
在Phytozome网站(
https://phytozome.jgi.doe.gov )中将各PtrPLD起始密码子前2 000 bp的序列作为启动子区域汇总,再上传至Plantcare网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html )进行顺式作用元件在线分析[46]。 -
组织表达特异性:在Phytozome网站(
https://phytozome.jgi.doe.gov )中下载各PtrPLD在各组织中的表达量数据,再采用qRT-PCR进行验证。野生型毛果杨来自中国科学院上海生命科学研究院,用组织培养的方法扩繁后,将1月大小的组培苗移植到土壤中,在25℃、长日照(光照16 h黑暗8 h)温室中培养3周。分别采集根、茎和叶组织,提取总RNA后反转录获得 cDNA用于qRT-PCR。每组处理重复3次,采用2 −∆∆CT法计算相对表达量并利用Tbtools可视化[41]。盐胁迫下的响应特性:将1月大小的组培苗移植到土壤中,在25℃、长日照(光照16 h黑暗8 h)温室中培养3周时,将长势一致的幼苗随机分成7组,每组包含5棵毛果杨幼苗。用100 mmol·L−1NaCl处理3、6、12、24、36、72 h,同时用水处理作为对照组。分别采集上述处理组内各植株材料的根、茎和叶组织,提取总RNA后反转录获得 cDNA进行qRT-PCR分析。每组处理重复3次,采用2 −∆∆CT法计算相对表达量并利用Tbtools可视化[41]。
RNA提取、反转录及qRT-PCR:利用植物总RNA提取试剂盒(MiniBEST,TaKaRa)提取总RNA,然后采用 PrimeScript TM RT reagent Kit(Perfect Real Time,TaKaRa)试剂盒反转录 RNA获得 cDNA 用于 qRT-PCR。根据荧光定量引物设计原则,利用NCBI Primer-Blast(
www.ncbi.nlm.nih.gov/tools/primer )工具设计特异性的PtrPLD家族基因定量引物,以PtrACTIN为内参基因(表1),并通过PCR、电泳验证引物特异性[50]。在赛默飞ABI 7500实时荧光定量PCR仪上进行试验,体系如下: 2×TransStart TOP/Tip Green qPCR Supermix 10 μL、上下游混合引物(10 μmol·L−1) 0.4 μL、cDNA 1.5 μL,Passive Reference Dye (50×) 0.4 μL,加ddH2O至20 μL。反应程序:94℃ 30 s; 94℃ 5 s,60℃ 15 s,72℃ 35 s,循环40次; 95℃ 15 s,60℃ 1 min,95℃ 30 s。表 1 毛果杨PtrPLD定量引物序列
Table 1. PtrPLD quantitative primer sequence
定量引物名称
qRT-PCR
primer nameF端引物序列(5′–3′)
F-terminal primer sequence
(5′–3′)R端引物序列(5′–3′)
R-terminal primer sequence
(5′–3′)PtrPLDα1 TTGATAAAGATGCAGAGATTCAGGG AATATCAACCTTGTGCTTCTCCA PtrPLDα2 GAATGGAAGCCCCACTTACG AGTTGAGGATTCGGCTTTCC PtrPLDα3 GCATGGCTTCCTCCACAAG AAGGATTCTGGTCCTCCCAA PtrPLDα4 TAGGCCAGCTTGATGACACT CCTGGTAAATCGTCGTCCAA PtrPLDα5 TCATTTGCTCCGTTACCCCA TAAAATCGGAGGAGCAGGGA PtrPLDα6 TGGGAGGATACCAACCACAT ATCCTGTTCGAGAGTTTCGC PtrPLDβ1 CCTCACTCTGTTTTGTGGGC ACAGCAATCACAGCCTATCCA PtrPLDβ2 AGCGTCCTCGTAACCTGAAA TGGACAATCCCCTGAACAAGAA PtrPLDδ1 TGTGACCAGGTGAGATGTGG AAAACCCACTGGTTGCTCCT PtrPLDδ2 CACACAAAGGGTCACGAGGT CAGCCATTTTATTGGGCGGG PtrPLDδ3 CTCAGCTTTTTCAAGCAGCA CATAGCGGCCATCACAAAGA PtrPLDδ4 CTTTGTGATGGCCGCTATGATACACC GAAACTTCGTCCAAACTCTGACCATT PtrPLDδ5 ATACCAAAAGCCCAAGGCAA ATGCCAACGAGTGACCTTTT PtrPLDξ1 CTAGCTCCCGGCTTGATTCC ATGGCGCAGAACCCTCAAAT PtrPLDξ2 TTGACCACGTTGGCTGGTT TTAATTGCGCTGACGATGCC PtrPLDξ3 CCACCTTTTGAACGCTTCCAG GAGAAAGGGAGAGACGGTGG PtrActin AGGCAGGTTTCGCAGGAGATGA TCCATCACCAGAATCCAGCACA -
根据已报道的AtPLD家族各基因氨基酸序列比对毛果杨的基因组数据库得到18个候选基因,再经过HKD结构域且二者间隔250~400个氨基酸为标准筛选,剔除Potri.001G112100.1和Potri.T180000.1(2个基因所编码的蛋白中HKD序列不符合HxKxxxxD特征),共得到16个PtrPLD(表2)。按编码PLD蛋白类型和进化关系命名为PtrPLDα1到PtrPLDξ3(表2);同时,对PtrPLD家族成员编码蛋白特性进行分析,结果表明,PtrPLD家族基因编码蛋白的氨基酸残基数为 645~1141、分子量为73.79~129.02 kDa、理论等电点为5.41~6.90、单个HKD结构域跨度在35个氨基酸左右、2个HKD结构域间隔均在300个氨基酸左右(表2)。亚细胞定位预测发现,PtrPLDα1和PtrPLDα2定位在细胞质中,PtrPLDα3、PtrPLDα4、PtrPLDα5和PtrPLDα6定位在内质网和液泡中,PtrPLDβ1定位在叶绿体中,PtrPLDβ2定位在叶绿体和细胞质中,PtrPLDδ1~5和PtrPLDξ1~3均定位在细胞质中(表2)。
表 2 毛果杨PLD基因及所编码蛋白的特征
Table 2. Characteristics of PLD genes and encoded proteins in Populus trichocarpa
基因名称
Gene name登录号
ID基因位置
Gene location(5′–3′)蛋白 Protein 氨基酸数/
aa等电点
PI分子量/
kDa亚细胞定位
Subcellular localizationHKD位置
Location CoordinatesPtrPLDα1 Potri.003G035300.1 4 527 875–4 531 587 645 6.16 73.79 C 185-221 493-520 PtrPLDα2 Potri.003G037700.1 4 909 260–4 912 047 759 6.73 86.87 C 300-336 608-635 PtrPLDα3 Potri.003G032800.1 4 092 810–4 086 017 808 5.41 91.75 E,V 326-364 654-681 PtrPLDα4 Potri.018G131200.1 15 250 553–15 246 358 808 6.57 91.83 E,V 327-365 655-682 PtrPLDα5 Potri.001G193000.1 17 591 592–17 585 937 808 5.63 91.82 E,V 326-364 654-681 PtrPLDα6 Potri.006G253900.1 25 888 555–25 896 527 791 5.85 89.89 E,V 309-347 637-664 PtrPLDβ1 Potri.002G152100.1 11 493 281–11 501 595 1 141 6.54 126.41 Chl 655-690 992-1 019 PtrPLDβ2 Potri.014G074700.1 6 036 483–6 044 469 1 122 6.4 124.16 Chl,C 639-674 974-1 001 PtrPLDδ1 Potri.007G060300.1 6 495 119–6 501 652 867 6.90 98.19 C 367-402 715-742 PtrPLDδ2 Potri.005G105600.1 8 078 955–8 085 766 882 6.65 100.29 C 382-417 730-757 PtrPLDδ3 Potri.005G246000.1 25 011 569–25 017 349 836 6.77 95.29 C 345-380 684-711 PtrPLDδ4 Potri.002G016100.1 930 355–925 137 794 6.77 90.27 C 303-338 643-670 PtrPLDδ5 Potri.003G015000.1 1 911 667–1 901 766 853 6.87 96.57 C 362-397 701-728 PtrPLDξ1 Potri.013G012300.1 813 485–807 432 1 111 6.26 126.35 C 485-512 907-933 PtrPLDξ2 Potri.008G211100.1 16 452 650–16 466 873 1 140 6.4 129.02 C 513-540 938-964 PtrPLDξ3 Potri.010G006300.1 579 514–567 938 1 123 6.26 126.77 C 496-523 922-948 注:C:细胞质;E:内质网;V:液泡;Chl:叶绿体。
Notes:C, cytoplasm; E, Endoplasmic reticulum; V, Vacuole; Chl, Chloroplast. -
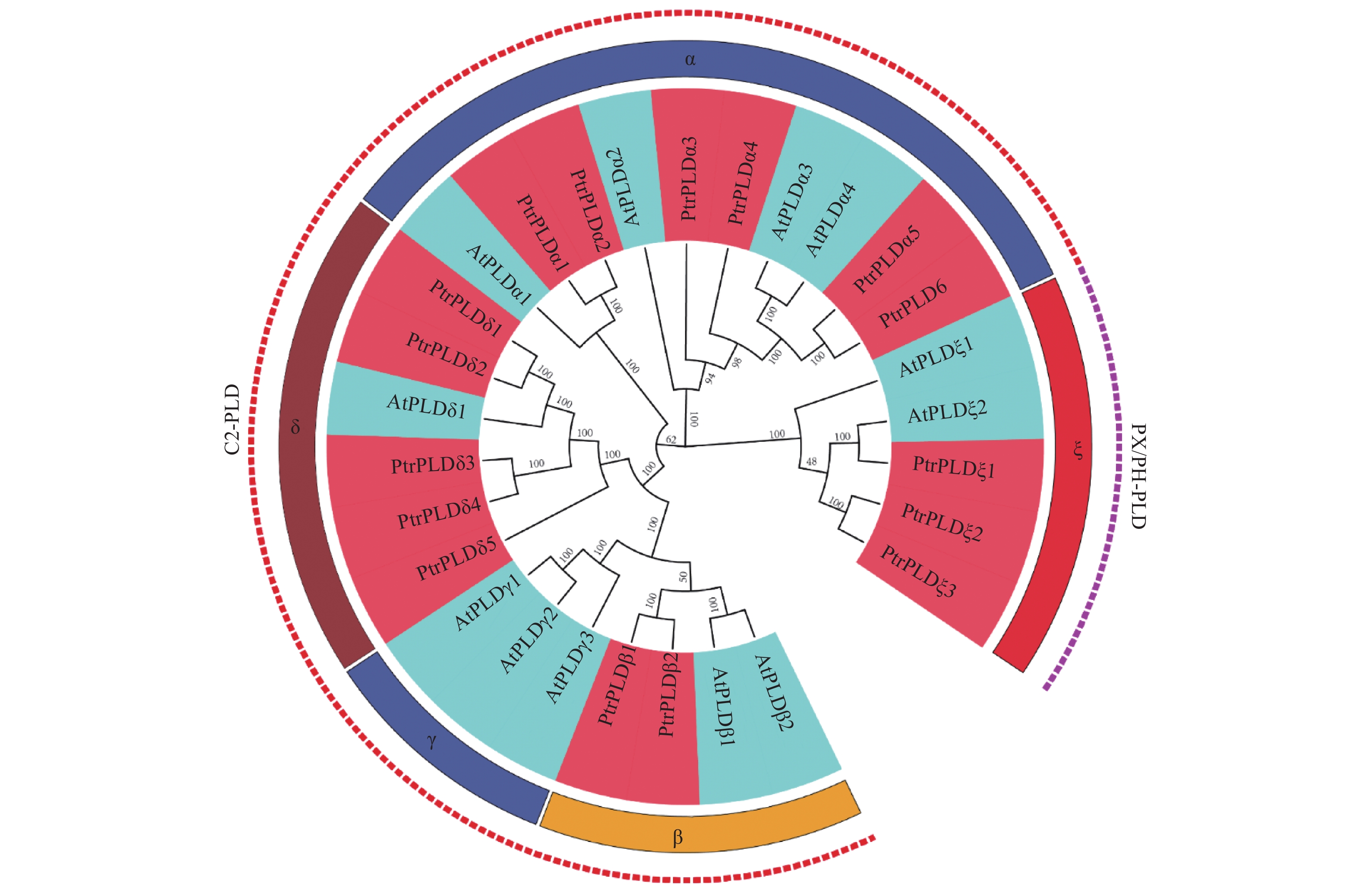
为了解PtrPLD家族成员的进化关系,将16个PtrPLDs与12个AtPLDs编码的蛋白构建进化树。根据拟南芥PLD基因家族进化关系,将PtrPLD家族分为α、β、δ及ζ 4个类型,其中α类型有6个,β类型有2个,δ类型有5个,ζ类型有3个,未鉴定出γ、ε和φ类型的PtrPLD(图1)。结果表明:毛果杨基因组并不存在SP-PLD亚族,PX/PH-PLD亚家族规模较小仅有3个成员,C2-PLD亚家族共有13个成员。
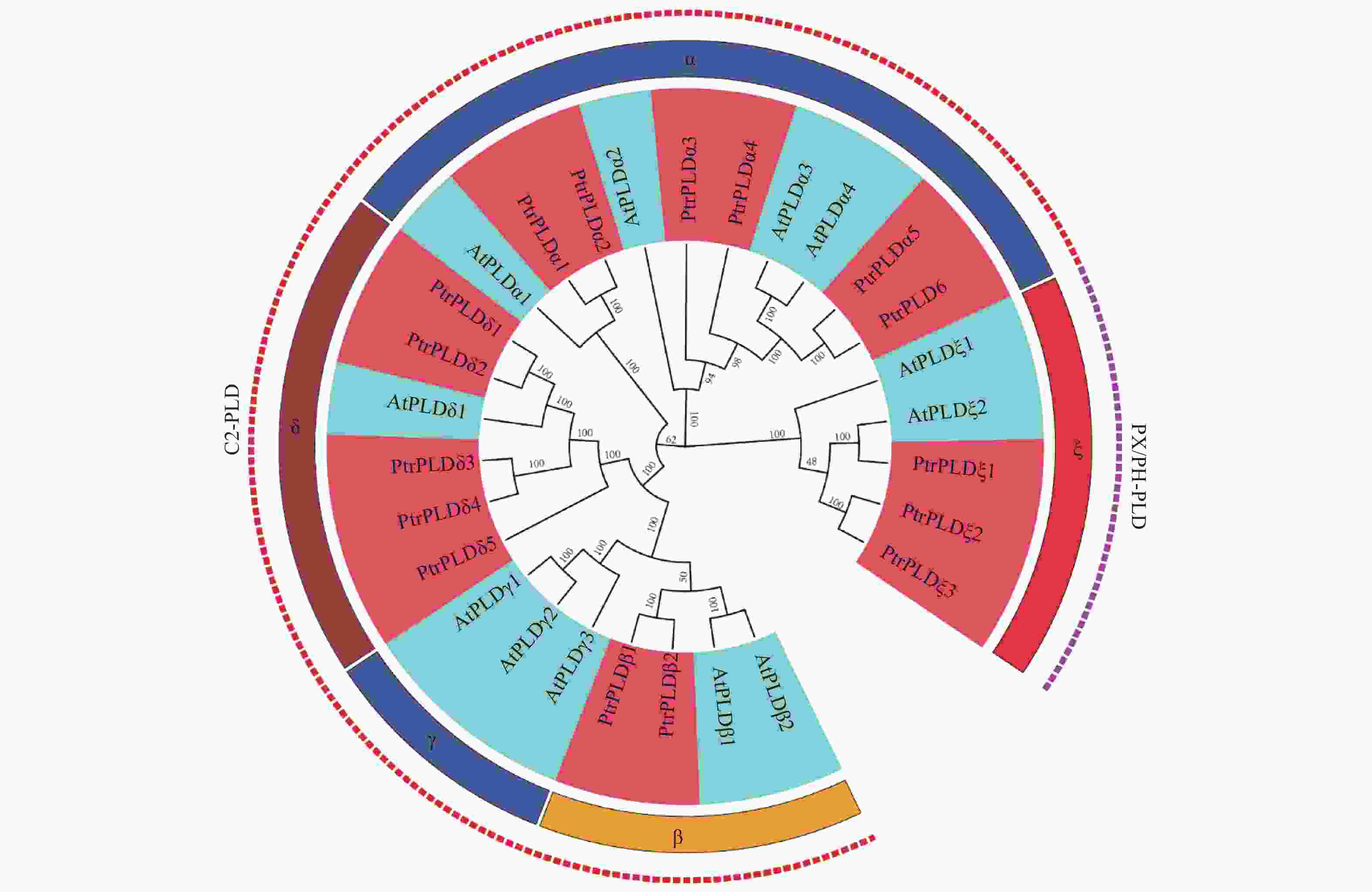
图 1 毛果杨与拟南芥PLD基因家族的系统发育进化分析。
Figure 1. Phylogenetic analysis of PLD gene family in Populus trichocarpa and Arabidopsis thaliana.
-
为了分析PtrPLD家族扩张原因,利用Tbtools构建了家族基因染色体定位图以及同源关系图[41]。结果表明:毛果杨16个PtrPLDs不均匀地分布在1、2、3、5、6、7、8、10、13、14和18号染色体上,其中,3号染色体上有4个基因、2号和5号染色体上有2个,其余的染色体上均仅有1个PtrPLD家族基因(图2)。

图 2 PtrPLDs基因染色体分布
Figure 2. Chromosome distribution of PtrPLDs
毛果杨PtrPLD家族共线性分析以及序列Blastn分析结果表明:PtrPLDα1和PtrPLDα2、PtrPLDα5和PtrPLDα3、PtrPLDβ1和PtrPLDβ2、PtrPLDδ1和PtrPLDδ4以及PtrPLDδ2、PtrPLDδ3和PtrPLDδ4、PtrPLDξ2和PtrPLDξ3有共线性关系且同源片段长度远大于300 bp同源性超过80%(表3),因此上述基因对是由基因复制事件的进化形成的旁系同源基因[46- 47]。其中,3号染色体上出现由PtrPLDα1、PtrPLDα2和PtrPLDα3形成的基因簇(图2),PtrPLDα1和PtrPLDα2同源性极高(表3),因此,PtrPLDα1和PtrPLDα2是由基因串联复制事件演化而来。综上所述,多数PtrPLDs具有基因复制现象,表明基因复制是PtrPLDs家族扩张的主要原因。
表 3 同源基因的Ka/Ks比值及同源性
Table 3. Ka/Ks values and homologous status of homologous genes
同源基因 Paralogous gene 非同义替换率
Ka同义替换率
KsKa/Ks 同源片段长度
The length of homologous
fragment/bp同源性
Homology基因1
Gene1基因2
Gene2PtrPLDα1 PtrPLDα2 0.0356523 0.0647533 0.5505866 1777 0.99 PtrPLDα5 PtrPLDα3 0.0360217 0.2439526 0.1476586 2426 0.93 PtrPLDβ1 PtrPLDβ2 0.0716041 0.2495736 0.2869057 2986 0.92 PtrPLDδ1 PtrPLDδ4 0.0599965 0.2616273 0.2293205 1520 0.91 PtrPLDδ2 0.2140527 1.7074231 0.1253659 1487 0.92 PtrPLDδ3 PtrPLDδ4 0.0503757 0.3069272 0.1641291 1487 0.92 PtrPLDξ2 PtrPLDξ3 0.0461424 0.2774425 0.1663134 2982 0.91 为了阐明选择压力在PtrPLD家族进化中的作用,利用Tbtools分析了PtrPLD家族同源基因的Ks值、Ka值和Ka/Ks值。结果发现:7对同源基因Ka/Ks均远小于1(表3),表明PtrPLD家族在进化过程中经历了较强的纯化选择。
-
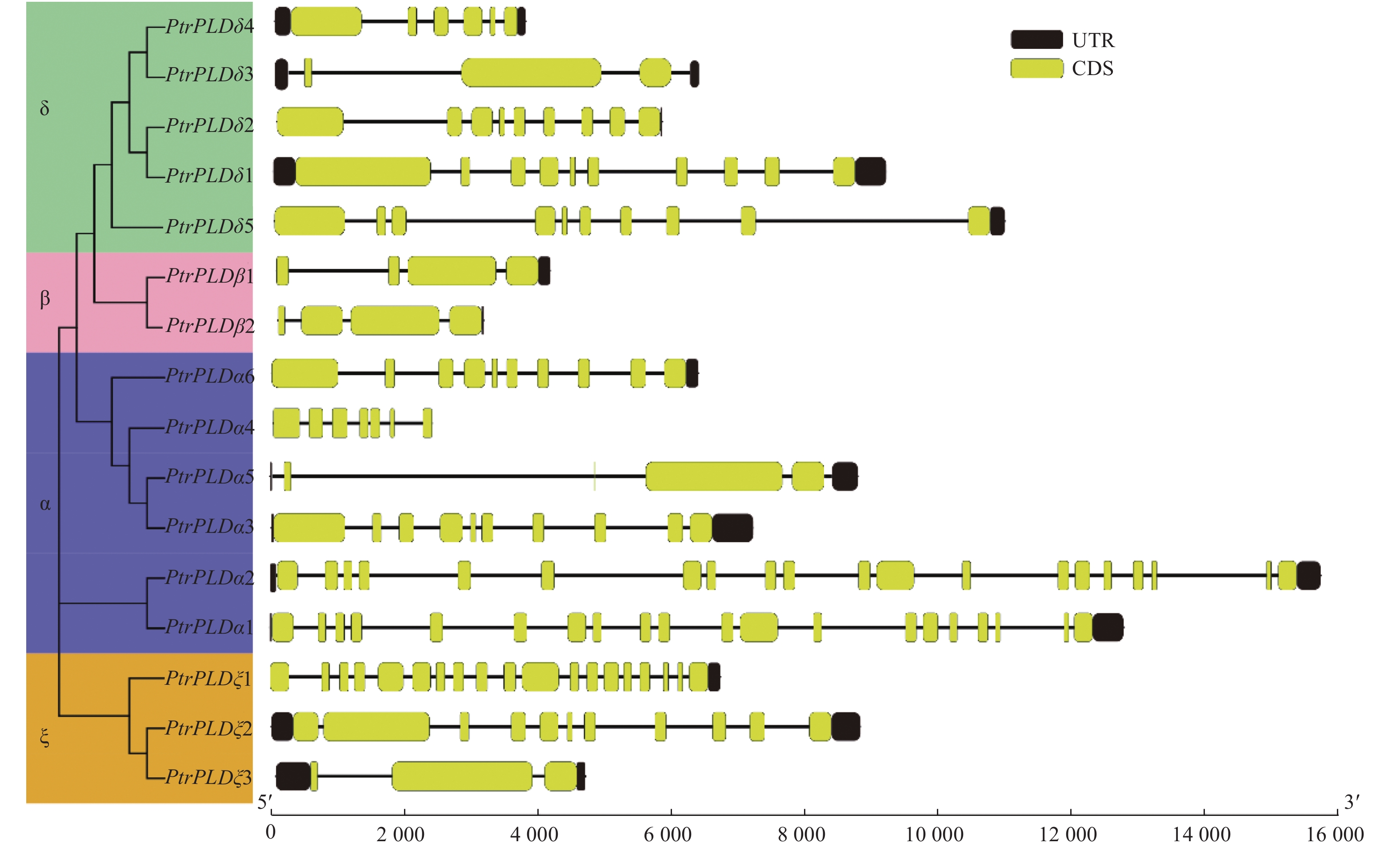
为了解PtrPLD家族各基因系统进化与其基因结构及其编码蛋白中保守基序之间的关系,对PtrPLD家族基因外显子与内含子的结构以及编码蛋白保守结构域进行了分析。结果表明:PtrPLD家族各基因的内含子和外显子分布方式在保持家族保守性的同时,在进化过程中也产生了明显的分化。除PtrPLDα5和PtrPLDδ3外,其他的家族基因最后一个外显子均紧靠3′端UTR(图3)。同时,多数处于同一进化分支上的基因结构较为相似,如PtrPLDβ1和PtrPLDβ2均仅含有4个外显子;值得注意的是,PtrPLDα1和 PtrPLDα2均含有20个外显子,但内含子与外显子在基因中的分布松散,同时,上述2个基因序列长度虽然超过12 000 bp,且远大于家族内其他基因序列长度,然而二者编码氨基酸长度却是家族中最短的;此外,PtrPLDβs基因长度均在5 000 bp以下,但其氨基酸数量最多,长度最长(图3)。
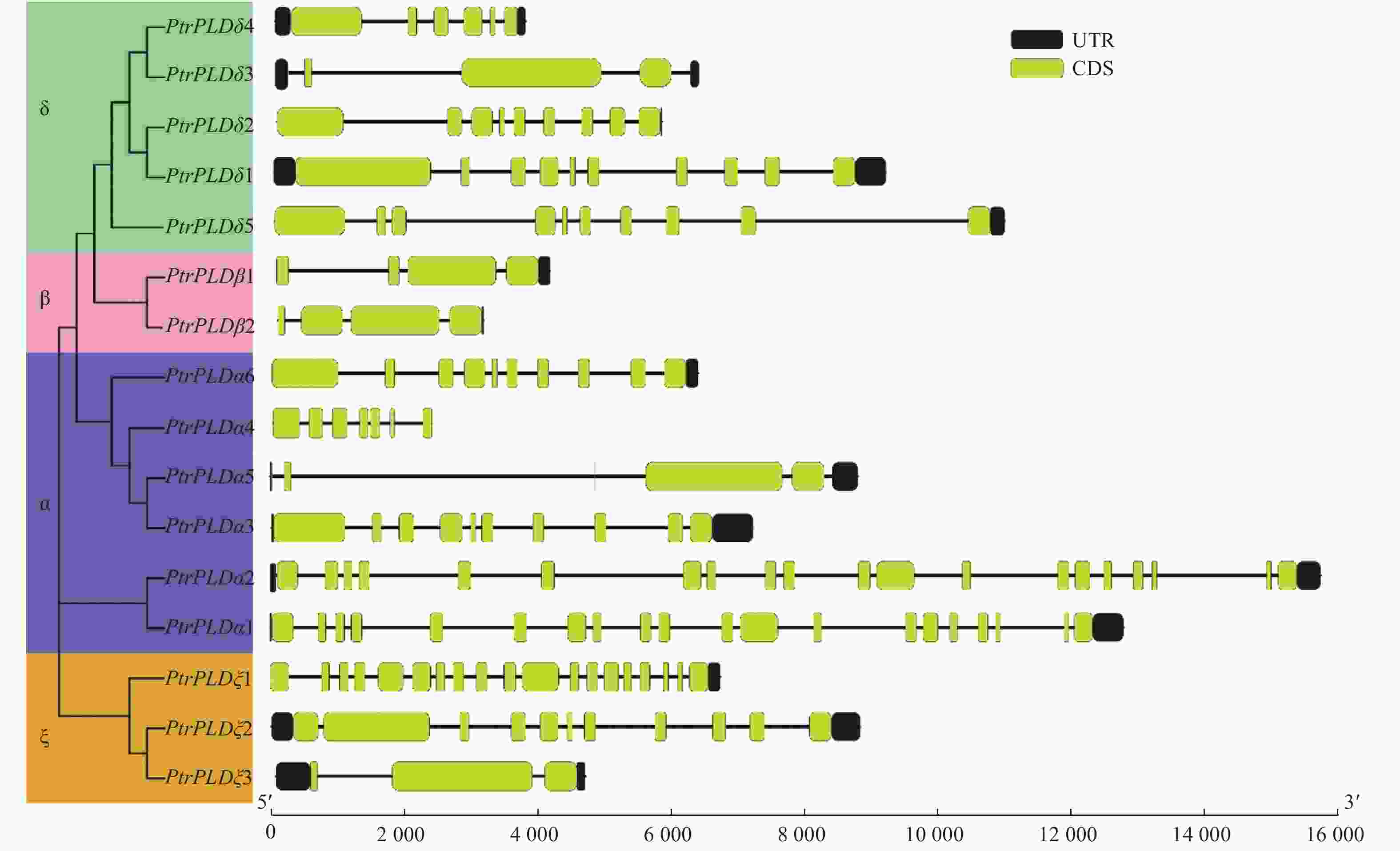
图 3 PtrPLD家族基因的外显子-内含子结构分析
Figure 3. Exon-intron organization analysis of PtrPLD genes
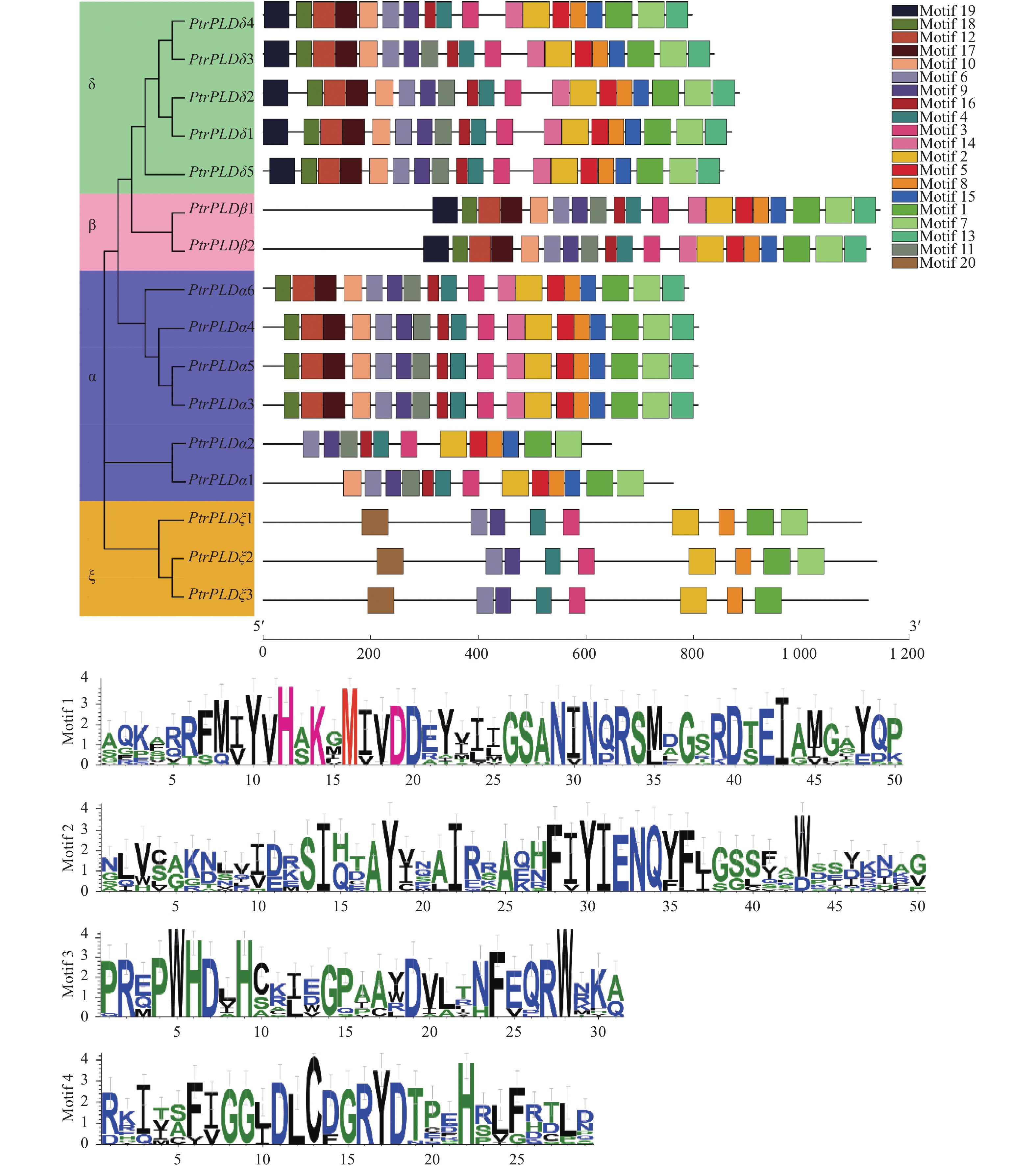
PtrPLD家族基因编码蛋白具有20个基序,其中Motif 1、Motif 2、Motif 3 和Motif 4为家族共有基序。虽然不同基因编码蛋白所含基序数量与种类存在一定的差异,但相同分支的基因编码蛋白具有相似的基序组合,而不同亚族、类型间的基序有明显的区别(图4)。上述结果表明:PtrPLD家族基因编码蛋白结构在具有保守性的同时也产生了明显分化。
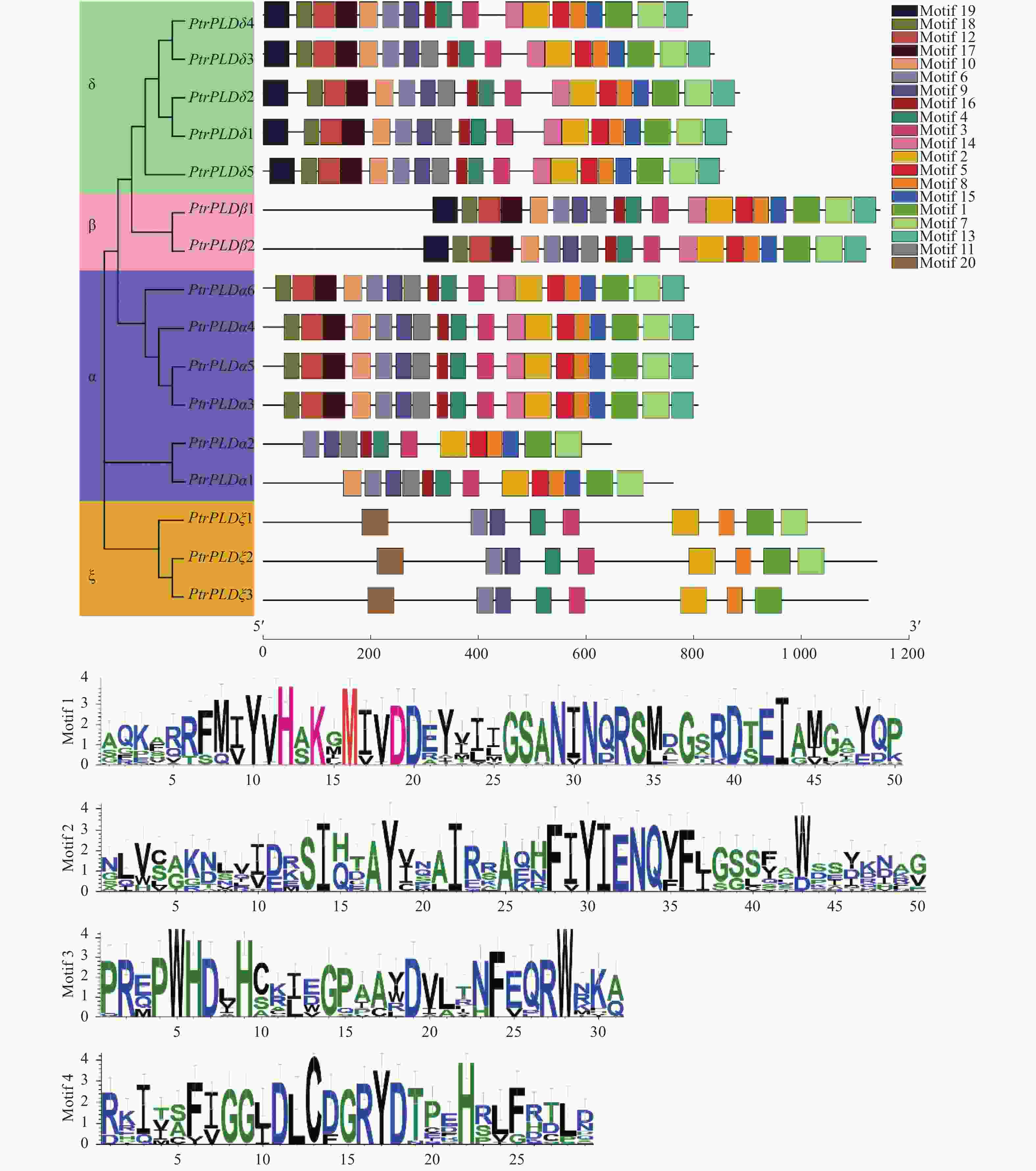
图 4 PtrPLD蛋白的保守基序分析
Figure 4. Conserved motifs analysis of PtrPLD proteins
-
为了解PtrPLD家族基因可能的生物学功能和调控特性,在利用PlantCARE分析家族成员启动子的同时,结合PtrPLD家族基因系统发育树分析各基因启动子元件组成与基因的进化之间是否具有相关性。结果表明:PtrPLD家族16个基因启动子中共包含2类、12种、154个顺式作用元件(表4)。一大类是非生物胁迫响应元件,如厌氧诱导元件(ARE)、损伤反应元件(WUN-motif)、胁迫响应元件(STRE)、MYB干旱诱导性结合位点(MBS)、DREB/CBF转录因子识别位点(DRE)、抗病和胁迫诱导元件(TC-rich repeats)以及低温响应元件(LTR);另一大类是植物激素响应元件,如生长素响应元件(TGA-element)、水杨酸诱导元件(TCA-element)、茉莉酸甲酯响应元件(CGTCA-motif 和TGACG-motif)、ABA应答元件(ABRE)。
表 4 PtrPLD家族基因启动子区域的各顺式作用元件分析
Table 4. Analysis of cis acting elements in PtrPLD genes promoter region
基因名称
Gene name植物激素响应元件
Phytohormone response element非生物胁迫响应元件
Abiotic stress response element共计
AmountTGA-element TCA-element CGTCA-motif ABRE TGACG-motif ARE WUN-motif STRE MBS DRE TC-richrepeats LTR PtrPLDδ4 1 0 2 8 2 2 1 2 0 1 1 0 20 PtrPLDδ3 0 0 1 2 0 0 0 0 3 1 0 0 7 PtrPLDδ2 2 0 0 8 0 0 1 3 0 0 0 0 14 PtrPLDδ1 0 0 0 8 4 1 2 0 0 1 1 1 18 PtrPLDδ5 0 2 0 2 0 2 0 0 0 0 1 2 9 PtrPLDβ1 1 0 0 7 0 3 1 0 0 0 0 0 12 PtrPLDβ2 0 0 0 0 0 0 1 3 1 0 1 0 6 PtrPLDα6 0 0 0 0 0 1 0 1 0 0 0 0 2 PtrPLDα4 1 1 2 2 0 1 0 1 2 0 0 1 11 PtrPLDα5 1 3 0 0 0 2 3 3 1 0 0 0 13 PtrPLDα3 0 0 0 1 0 2 0 0 0 0 0 0 3 PtrPLDα2 1 0 0 4 1 1 2 0 0 0 0 1 10 PtrPLDα1 0 1 1 4 1 0 0 0 0 0 1 0 8 PtrPLDξ1 0 0 1 0 1 3 2 2 1 0 1 0 11 PtrPLDξ2 1 0 2 2 2 1 0 0 0 0 0 0 8 PtrPLDξ3 0 0 0 0 0 0 1 0 0 1 0 0 2 共计Amount 8 7 9 48 11 19 14 15 8 4 6 5 154 PtrPLD家族各基因启动子含有元件的种类和数量存在差异,其中,PtrPLDδ启动子中含有的顺式作用元件数量最多,如PtrPLDδ4启动子中具有的元件数量最多(20个),而PtrPLDα6和PtrPLDξ3启动子中仅含有2个元件。上述结果表明,基因在进化的同时,其启动子中元件也会产生相应的分化。
此外,在所有顺式作用元件中ABA应答元件ABRE数量最多,在11个PtrPLD启动子中有48个ABRE,其中,PtrPLDδ1、PtrPLDδ2以及PtrPLDδ4含有的ABRE元件达8个;厌氧诱导元件ARE含量次之,共有19个,分布在PtrPLDα6、PtrPLDδ4、PtrPLDδ2等11个基因启动子中;而DREB/CBF转录因子识别位点DRE数量最少,仅4个,分布在PtrPLDδ1、PtrPLDδ3、PtrPLDδ4和PtrPLDξ3基因启动子中。上述结果表明:PtrPLD家族不同的亚族基因响应植物激素和非生物胁迫的能力存在差异。
-
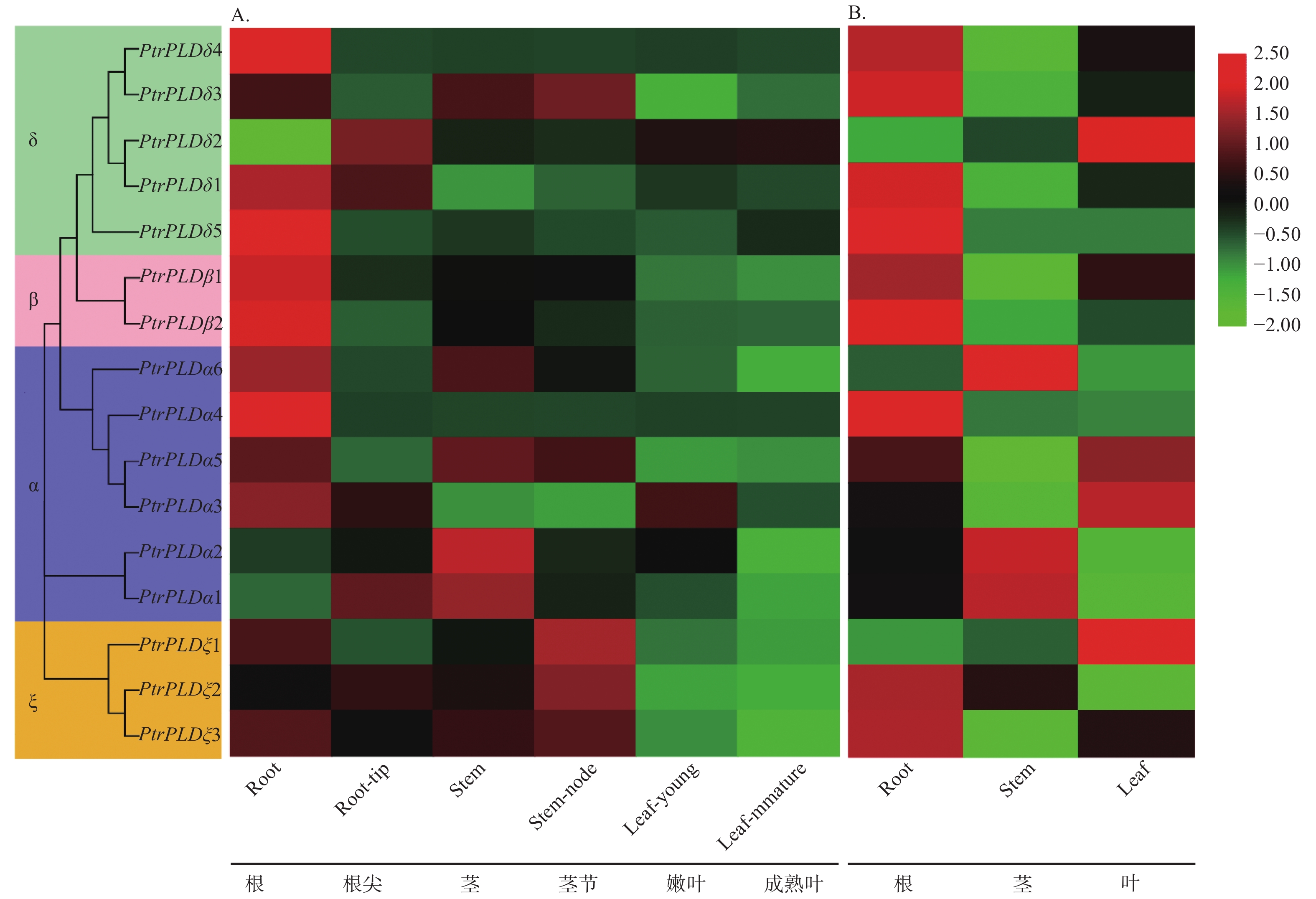
为了解PtrPLD家族基因的表达模式,对phytozome网站中PtrPLD家族各基因在不同组织的表达量进行分析。结果表明:6个PtrPLDα型蛋白基因中PtrPLDα1、PtrPLDα2和 PtrPLDα6在茎中的表达水平较高,PtrPLDα4和PtrPLDα5在根部的表达水平较高,PtrPLDα3在嫩叶和根部的表达水平较高,并未有明显的表达部位偏好;2个PtrPLDβ型蛋白基因在根部的表达水平均较高;5个PtrPLDδ型蛋白基因中只有PtrPLDδ2在根、嫩叶和成熟叶的表达水平较高,其他4个均在根的表达量较高;3个PtrPLDξ型蛋白基因均在茎节的表达水平较高,根部次之,叶中表达量最低(图5A)。同时,利用qRT-PCR对PtrPLD家族各基因在根、茎和叶中的表达量做进一步分析,结果表明:16个PtrPLD家族基因中有9个在根部的表达水平较高,3个在茎部表达水平较高,4个在叶部表达水平较高。qRT-PCR与phytozome网站中的基本相符,大部分的基因在根部的表达水平较高,但不同的是qRT-PCR结果中的PtrPLDα3和PtrPLDα5在叶中表达水平较高;3个PtrPLDξ型蛋白基因中PtrPLDξ1在叶部表达水平较高,PtrPLDξ2和PtrPLDξ3均在根部的表达量较高(图5B);PtrPLDδ2在叶中的表达水平高于根部的。

图 5 PtrPLD家族基因组织表达特异性
Figure 5. Tissue expression specificity of PtrPLD family genes
为了阐明PtrPLD家族基因的盐胁迫响应特性,利用qRT-PCR对100 mmol·L−1NaCl胁迫处理3、6、12、24、48、72 h的毛果杨幼苗的根、茎、叶中PtrPLD家族各基因的表达进行了研究。结果(图6)表明:NaCl胁迫对16个PtrPLD基因在各组织中表达均产生了影响;在根、茎、叶3种组织中全部PtrPLD家族基因在盐胁迫后12 h内表现出显著的表达上调趋势,且大部分是在6 h或12 h时达到表达高峰,之后快速下调,在48 h到72 h之间又略有上升。较为独特的是,PtrPLDδ4在茎部的表达高峰处于胁迫处理72 h时;PtrPLDα1在根、茎、叶中的48~72 h之间的上调表达趋势相较于其他PtrPLD的均较明显。从响应速度来看,叶和茎部中大多数PtrPLD家族基因响应盐胁迫的快速上升期发生在3 h或6 h,而在根部中的快速上升期发生在6 h或12 h。
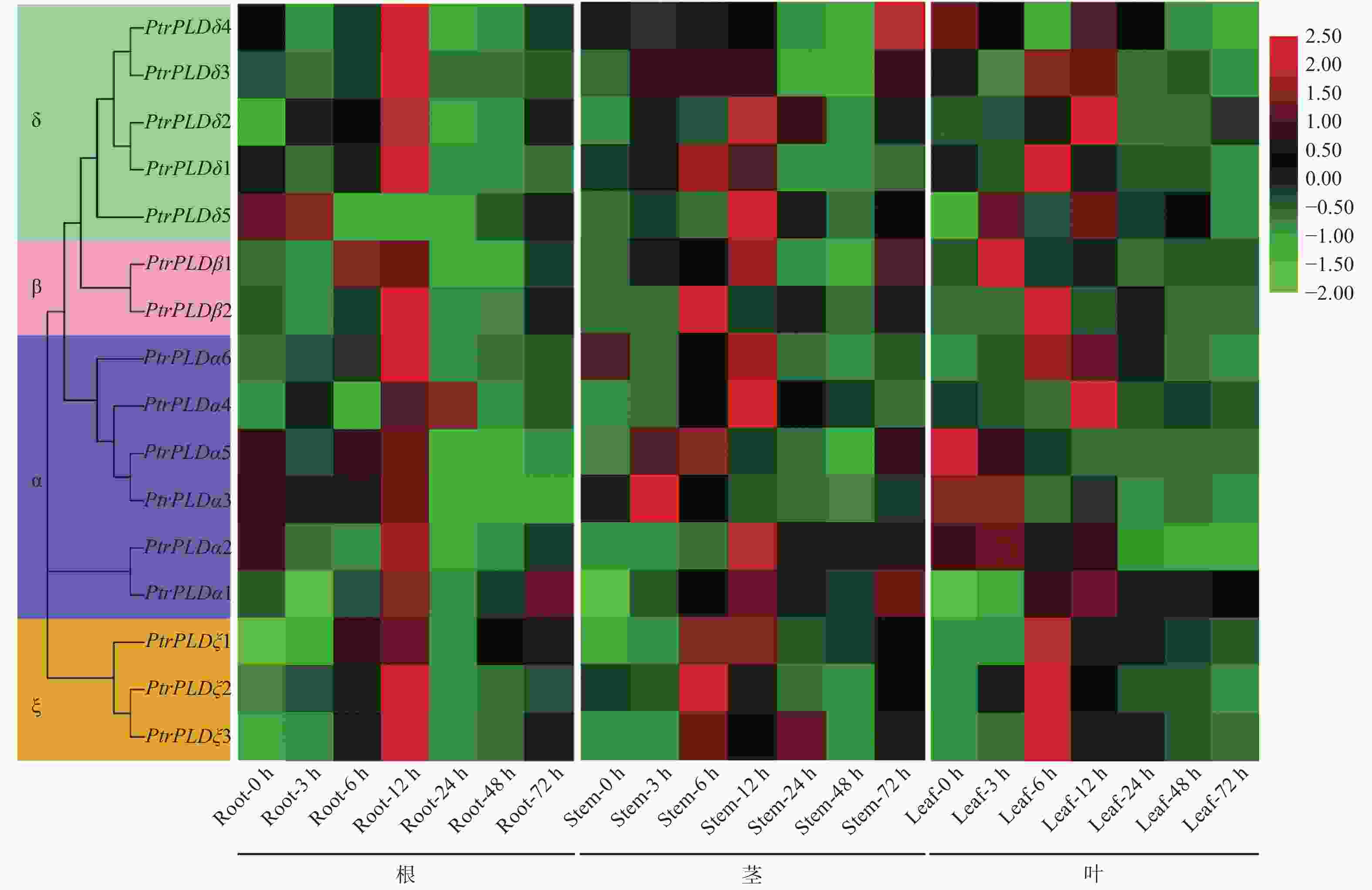
图 6 不同组织中PtrPLD基因家族在盐胁迫下的表达特性
Figure 6. Analysis of salt stress expression of PtrPLD family genes in different tissues
此外,各部位中均有部分PtrPLD家族基因在12 h内的表达高峰前出现下调表达的情况,而不是在胁迫处理后一直上调表达到峰值,如根部中的全部PtrPLDα、PtrPLDδ1、PtrPLDδ3和PtrPLDδ4,茎部的PtrPLDδ2、PtrPLDδ4和PtrPLDδ5,叶部的PtrPLDα2、PtrPLDδ1和PtrPLDδ4。
-
研究表明,PLD以多基因家族的形式存在于拟南芥[12]、水稻[13]、白菜[14]、亚洲棉[11]等植物中,并在胁迫响应中起到重要作用[10, 17, 29, 34]。然而,目前为止,尚无模式植物毛果杨PLD家族基因基本特征及其在逆境胁迫响应特性的研究报导。本研究通过对毛果杨全基因组分析,共鉴定出16个PtrPLD,其编码蛋白包括3个PX/PH-PLD和13个C2-PLD亚家族成员(图1),这与其它植物中C2-PLD成员数量相对较少的情况相同[12]。PtrPLD家族中共有7对具有旁系同源关系,其中PtrPLDα1和PtrPLDα2是由基因串联复制事件演化而来,其余的6对是由基因组复制事件演化而来,这表明基因复制是PtrPLD基因家族扩张的主要原因(图2和表3)。PtrPLD家族中的7对同源基因的Ka/Ks比值远低于1(表3),表明它们在进化过程中经过了较强的纯化选择,有害的非同义替换在进化过程消失,极少数的无害或有益替换得以保留[48]。上述现象间接证明了PtrPLD家族基因在毛果杨的生命活动中发挥着重要作用。
PtrPLD家族基因编码蛋白含有多个基序,表明该基因家族生物学功能的多样性。同时,所有的PtrPLD家族基因编码蛋白均含有Motif 1~4基序(图4),表明这4个基序是该基因家族的特征基序,且是毛果杨生命活动所必须的。此外,序列比对表明,Motif 1中的HKD基序中的甲硫氨酸高度保守,这种现象也存在于其他植物的PLD蛋白中[11, 51-52],因此,推测Motif 1中的甲硫氨酸是HKD基序功能的关键位点。
PtrPLD家族成员的表达特性分析表明,其中9个家族成员在根部表达水平较高,3个在茎部表达水平较高,4个在叶部表达水平较高(图5)。前期研究认为,PLD基因多涉及环境胁迫响应[10, 17, 29, 34],而根部是植物感知渗透胁迫的首要部位。据此推测,在毛果杨根部表达水平较高的9个PtrPLD基因可能在根部渗透胁迫信号转导过程中发挥重要作用,而其余7个在茎、叶中表达水平较高的可能在茎部或叶部的胁迫信号转导过程中发挥功能。此外,phytozome网站中数据与qRT-PCR结果中的组织表达特性不同的原因可能是所用材料的树龄不同,同时它们在不同发育阶段中的表达模式存在差异所致。
上游转录因子结合下游基因启动子中顺式作用元件,从而激活逆境响应基因的表达是植物响应环境胁迫的重要环节[53]。因此,对基因启动子区域顺式元件的分析,是鉴定分析基因生物学功能的一项重要内容。本研究发现,PtrPLD家族基因启动子中含有多种与植物激素和逆境胁迫响应相关的两大类响应元件(表4),表明该基因家族广泛地参与毛果杨多种逆境胁迫的响应。PtrPLD家族基因在盐胁迫下的响应特性也证实了上述观点。
-
本研究通过氨基酸序列比对及保守结构域检验,在毛果杨中鉴定了16个PtrPLDs;通过系统发育进化分析将其分为2个亚族,4种类型;利用同源性分析及Ka、Ks分析PtrPLD基因家族说明基因复制是其家族扩张的主要动力,且在进化过程中受到较强地纯化选择;系统发育进化树与基因结构和保守结构域分析表明,同一亚族、同一类型的PtrPLD具有相似的基因结构和蛋白保守结构域;启动子中顺式作用元件分析表明,16个PtrPLDs均能够响应多种激素和胁迫信号;组织表达特异性以及盐胁迫下的表达特性分析表明,同一进化分支上基因仍具有相同表达模式,而不同进化分支的基因存在功能分化。本研究对于PtrPLD家族基因生物学功能的鉴定和抗逆基因资源挖掘具有积极意义。
毛果杨PLD基因家族全基因组水平鉴定及其盐胁迫下的表达分析
Genome-wide Identification of PLD Gene Family of Populus trichocarpa and Its Responses to Salt Stress
-
摘要:
目的 对木本模式植物毛果杨PLD基因家族的进化中的选择压力、启动子中顺式作用元件、组织表达特性以及盐胁迫下表达模式进行分析,为挖掘PtrPLD在非生物胁迫中作用提供参考。 方法 利用拟南芥PLD基因家族蛋白序列比对得到毛果杨基因组同源基因, 再经过保守结构域鉴定后确定PtrPLD基因;利用软件ClustalW和MEGA对PtrPLD和AtPLD基因的氨基酸序列进行比对和系统进化分析;利用MEME、Plant-mPLoc、ExPasy等软件工具分析PtrPLD基因及编码蛋白的特征;利用Tbtools软件分析同源基因的Ka/Ks值;利用Plantcare在线工具分析PtrPLD启动子中顺式作用元件;利用Phytozome转录组数据库以及qRT-PCR分析PtrPLD组织表达特性;利用qRT-PCR分析各组织中PtrPLD对盐胁迫响应情况。 结果 PtrPLD家族的16个基因可分为C2-PLD和PX/PH-PLD 2个亚家族,分别包含13个和3个基因;有7对旁系同源基因且它们之间的Ka/Ks均远小于1;PtrPLD家族基因启动子区含有大量非生物胁迫和激素响应元件,其中,PtrPLDδ4启动子共含有9种、20个元件;PtrPLD家族编码的蛋白均含有Motif 1~4,且同一进化分支上的基因编码蛋白序列高度保守。PtrPLD基因家族表达特性分析表明,PtrPLD家族基因在根、茎和叶中具有表达特异性,并且多数成员主要在根部表达;NaCl胁迫下,PtrPLD家族基因在根、茎和叶中表达量在0~72 h内均表现为先上升后下降再上升的变化趋势。 结论 PtrPLD家族基因在毛果杨响应盐胁迫过程中有着重要作用,本项研究对于今后PtrPLD家族基因生物学功能的鉴定与非生物逆境胁迫响应基因资源的挖掘具有推动作用。 Abstract:Objective To analyze the pressure of selection in the evolution of PLD gene family in the woody model plant Populus trichocarpa, the cis-acting elements in the promoter, the tissue expression characteristics, and the expression pattern under salt stress in order to provide references for mining the role of PtrPLD in abiotic stress. Method Using Arabidopsis PLD gene family protein sequence alignment to obtain the P. trichocarpa genome homologous gene, and then identify the PtrPLD gene after the conserved domain identification; using the software ClustalW and MEGA to perform the amino acid sequence alignment and systematic evolution analysis of the PtrPLD and AtPLD genes; using MEME, Plant-mPLoc, ExPasy and other software tools to analyze the characteristics of PtrPLD genes and encoded proteins; using Tbtools software to analyze homologous genes and Ka/Ks values; using Plantcare online tools to analyze the cis-acting elements in the promoter of PtrPLD; using Phytozome transcriptome database and qRT-PCR to analyze the PtrPLD tissue expression characteristics; and using qRT-PCR to analyze the PtrPLD response to salt stress in each tissue. Result The results show that the PtrPLD family can be divided into 2 subfamilies, C2-PLD and PX/PH-PLD, which containing 13 and 3 genes respectively. There are 7 pairs of paralogous genes and the Ka/Ks between them is far less than 1. The promoter region of PtrPLD family genes contains a large number of abiotic stress and hormone response elements, among which the PtrPLDδ4 promoter contains 20 elements. The PtrPLD family of encoded proteins all contain Motif 1-4, and the sequences on the same evolutionary branch of the evolutionary tree are highly conserved. Analysis of the expression characteristics showed that the PtrPLD family genes have specific expression in roots, stems and leaves, and most members are mainly expressed in the roots. Under NaCl stress, the expression level of PtrPLD family genes in the roots, stems and leaves showed a trend of up-down-up within 72 hours. Conclusion The results show that the PtrPLD family genes play an important role in the response of P. trichocarpa to salt stress. This study will promote the identification of the biological functions of PtrPLD family genes and the mining of genetic resources in response to abiotic stress. -
Key words:
- Populus trichocarpa
- / PLD gene family
- / bioinformatics analysis
- / salt stress
-
图 1 毛果杨与拟南芥PLD基因家族的系统发育进化分析。
Figure 1. Phylogenetic analysis of PLD gene family in Populus trichocarpa and Arabidopsis thaliana.
图 3 PtrPLD家族基因的外显子-内含子结构分析
Figure 3. Exon-intron organization analysis of PtrPLD genes
图 6 不同组织中PtrPLD基因家族在盐胁迫下的表达特性
Figure 6. Analysis of salt stress expression of PtrPLD family genes in different tissues
表 1 毛果杨PtrPLD定量引物序列
Table 1. PtrPLD quantitative primer sequence
定量引物名称
qRT-PCR
primer nameF端引物序列(5′–3′)
F-terminal primer sequence
(5′–3′)R端引物序列(5′–3′)
R-terminal primer sequence
(5′–3′)PtrPLDα1 TTGATAAAGATGCAGAGATTCAGGG AATATCAACCTTGTGCTTCTCCA PtrPLDα2 GAATGGAAGCCCCACTTACG AGTTGAGGATTCGGCTTTCC PtrPLDα3 GCATGGCTTCCTCCACAAG AAGGATTCTGGTCCTCCCAA PtrPLDα4 TAGGCCAGCTTGATGACACT CCTGGTAAATCGTCGTCCAA PtrPLDα5 TCATTTGCTCCGTTACCCCA TAAAATCGGAGGAGCAGGGA PtrPLDα6 TGGGAGGATACCAACCACAT ATCCTGTTCGAGAGTTTCGC PtrPLDβ1 CCTCACTCTGTTTTGTGGGC ACAGCAATCACAGCCTATCCA PtrPLDβ2 AGCGTCCTCGTAACCTGAAA TGGACAATCCCCTGAACAAGAA PtrPLDδ1 TGTGACCAGGTGAGATGTGG AAAACCCACTGGTTGCTCCT PtrPLDδ2 CACACAAAGGGTCACGAGGT CAGCCATTTTATTGGGCGGG PtrPLDδ3 CTCAGCTTTTTCAAGCAGCA CATAGCGGCCATCACAAAGA PtrPLDδ4 CTTTGTGATGGCCGCTATGATACACC GAAACTTCGTCCAAACTCTGACCATT PtrPLDδ5 ATACCAAAAGCCCAAGGCAA ATGCCAACGAGTGACCTTTT PtrPLDξ1 CTAGCTCCCGGCTTGATTCC ATGGCGCAGAACCCTCAAAT PtrPLDξ2 TTGACCACGTTGGCTGGTT TTAATTGCGCTGACGATGCC PtrPLDξ3 CCACCTTTTGAACGCTTCCAG GAGAAAGGGAGAGACGGTGG PtrActin AGGCAGGTTTCGCAGGAGATGA TCCATCACCAGAATCCAGCACA  下载: 导出CSV
下载: 导出CSV
表 2 毛果杨PLD基因及所编码蛋白的特征
Table 2. Characteristics of PLD genes and encoded proteins in Populus trichocarpa
基因名称
Gene name登录号
ID基因位置
Gene location(5′–3′)蛋白 Protein 氨基酸数/
aa等电点
PI分子量/
kDa亚细胞定位
Subcellular localizationHKD位置
Location CoordinatesPtrPLDα1 Potri.003G035300.1 4 527 875–4 531 587 645 6.16 73.79 C 185-221 493-520 PtrPLDα2 Potri.003G037700.1 4 909 260–4 912 047 759 6.73 86.87 C 300-336 608-635 PtrPLDα3 Potri.003G032800.1 4 092 810–4 086 017 808 5.41 91.75 E,V 326-364 654-681 PtrPLDα4 Potri.018G131200.1 15 250 553–15 246 358 808 6.57 91.83 E,V 327-365 655-682 PtrPLDα5 Potri.001G193000.1 17 591 592–17 585 937 808 5.63 91.82 E,V 326-364 654-681 PtrPLDα6 Potri.006G253900.1 25 888 555–25 896 527 791 5.85 89.89 E,V 309-347 637-664 PtrPLDβ1 Potri.002G152100.1 11 493 281–11 501 595 1 141 6.54 126.41 Chl 655-690 992-1 019 PtrPLDβ2 Potri.014G074700.1 6 036 483–6 044 469 1 122 6.4 124.16 Chl,C 639-674 974-1 001 PtrPLDδ1 Potri.007G060300.1 6 495 119–6 501 652 867 6.90 98.19 C 367-402 715-742 PtrPLDδ2 Potri.005G105600.1 8 078 955–8 085 766 882 6.65 100.29 C 382-417 730-757 PtrPLDδ3 Potri.005G246000.1 25 011 569–25 017 349 836 6.77 95.29 C 345-380 684-711 PtrPLDδ4 Potri.002G016100.1 930 355–925 137 794 6.77 90.27 C 303-338 643-670 PtrPLDδ5 Potri.003G015000.1 1 911 667–1 901 766 853 6.87 96.57 C 362-397 701-728 PtrPLDξ1 Potri.013G012300.1 813 485–807 432 1 111 6.26 126.35 C 485-512 907-933 PtrPLDξ2 Potri.008G211100.1 16 452 650–16 466 873 1 140 6.4 129.02 C 513-540 938-964 PtrPLDξ3 Potri.010G006300.1 579 514–567 938 1 123 6.26 126.77 C 496-523 922-948 注:C:细胞质;E:内质网;V:液泡;Chl:叶绿体。
Notes:C, cytoplasm; E, Endoplasmic reticulum; V, Vacuole; Chl, Chloroplast.
下载: 导出CSV
表 3 同源基因的Ka/Ks比值及同源性
Table 3. Ka/Ks values and homologous status of homologous genes
同源基因 Paralogous gene 非同义替换率
Ka同义替换率
KsKa/Ks 同源片段长度
The length of homologous
fragment/bp同源性
Homology基因1
Gene1基因2
Gene2PtrPLDα1 PtrPLDα2 0.0356523 0.0647533 0.5505866 1777 0.99 PtrPLDα5 PtrPLDα3 0.0360217 0.2439526 0.1476586 2426 0.93 PtrPLDβ1 PtrPLDβ2 0.0716041 0.2495736 0.2869057 2986 0.92 PtrPLDδ1 PtrPLDδ4 0.0599965 0.2616273 0.2293205 1520 0.91 PtrPLDδ2 0.2140527 1.7074231 0.1253659 1487 0.92 PtrPLDδ3 PtrPLDδ4 0.0503757 0.3069272 0.1641291 1487 0.92 PtrPLDξ2 PtrPLDξ3 0.0461424 0.2774425 0.1663134 2982 0.91
下载: 导出CSV
表 4 PtrPLD家族基因启动子区域的各顺式作用元件分析
Table 4. Analysis of cis acting elements in PtrPLD genes promoter region
基因名称
Gene name植物激素响应元件
Phytohormone response element非生物胁迫响应元件
Abiotic stress response element共计
AmountTGA-element TCA-element CGTCA-motif ABRE TGACG-motif ARE WUN-motif STRE MBS DRE TC-richrepeats LTR PtrPLDδ4 1 0 2 8 2 2 1 2 0 1 1 0 20 PtrPLDδ3 0 0 1 2 0 0 0 0 3 1 0 0 7 PtrPLDδ2 2 0 0 8 0 0 1 3 0 0 0 0 14 PtrPLDδ1 0 0 0 8 4 1 2 0 0 1 1 1 18 PtrPLDδ5 0 2 0 2 0 2 0 0 0 0 1 2 9 PtrPLDβ1 1 0 0 7 0 3 1 0 0 0 0 0 12 PtrPLDβ2 0 0 0 0 0 0 1 3 1 0 1 0 6 PtrPLDα6 0 0 0 0 0 1 0 1 0 0 0 0 2 PtrPLDα4 1 1 2 2 0 1 0 1 2 0 0 1 11 PtrPLDα5 1 3 0 0 0 2 3 3 1 0 0 0 13 PtrPLDα3 0 0 0 1 0 2 0 0 0 0 0 0 3 PtrPLDα2 1 0 0 4 1 1 2 0 0 0 0 1 10 PtrPLDα1 0 1 1 4 1 0 0 0 0 0 1 0 8 PtrPLDξ1 0 0 1 0 1 3 2 2 1 0 1 0 11 PtrPLDξ2 1 0 2 2 2 1 0 0 0 0 0 0 8 PtrPLDξ3 0 0 0 0 0 0 1 0 0 1 0 0 2 共计Amount 8 7 9 48 11 19 14 15 8 4 6 5 154
下载: 导出CSV
-
[1] Chinnusamy V, Schumaker K, Zhu J. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants[J]. Journal of Experimental Botany, 2004, 55(395): 225-236. [2] 朱健康, 倪建平. 植物非生物胁迫信号转导及应答[J]. 中国稻米, 2016, 22(6):52-60. doi: 10.3969/j.issn.1006-8082.2016.06.012 [3] Pandey S, Assmann S. M. The Arabidopsis putative Gprotein-coupled receptor GCR1 interacts with the G protein α subunit GPA1 and regulates abscisic acid signaling[J]. The Plant Cell, 2004, 16(6): 1616-1632. doi: 10.1105/tpc.020321 [4] Roos W, Dordschbal B, Steighard J, et al. A redox-dependent, G-protein-coupled phospholipase A of the plasma membrane is involved in the elicitation of alkaloid biosynthesis in Eschscholtzia californica[J]. BBA- Molecular Cell Research, 1999, 1448(3): 390-402. [5] Gao K, Liu Y L, Li B, et al. Arabidopsis thaliana phosphoinositide-specific phospholipase C isoform 3 (AtPLC3) and AtPLC9 have an additive effect on thermotolerance[J]. Plant & Cell Physiology, 2014, 55(11): 1873-1883. [6] Wan S, Tian L, Tian R, et al. Involvement of phospholipase D in the low temperature acclimation-induced thermotolerance in grape berry[J]. Plant Physiology & Biochemistry, 2009, 47(6): 504-510. [7] Hong Y, Zhao J, Guo L, et al. Plant phospholipases D and C and their diverse functions in stress responses[J]. Progress in Lipid Research, 2016, 6: 55-74. [8] 王雅静, 张欣莹, 黄桂荣, 等. 植物磷脂酸的特性及其在ABA诱导气孔运动中的作用[J]. 植物学报, 2019, 54(2):245-254. doi: 10.11983/CBB18115 [9] Welti R, Li W, Li M, et al. Profiling membrane lipids in plant stress responses. Role of phospholipase D alpha in freezing-induced lipid changes in Arabidopsis[J]. Journal of Biological Chemistry, 2002, 277(35): 31994-32002. doi: 10.1074/jbc.M205375200 [10] Hong Y, Zhang W, Wang X. Phospholipase D and phosphatidic acid signalling in plant response to drought and salinity[J]. Plant Cell & Environment, 2010, 33(4): 627-635. [11] Tank K, Dong C, Liu J. Genome-wide analysis and expression profiling of the phospholipase D gene family in Gossypium arboreum[J]. Science China-Life Sciences, 2016, 59(2): 130-141. doi: 10.1007/s11427-015-4916-2 [12] Qin C, Wang X. The Arabidopsis phospholipase D family. Characterization of a calcium-independent and phosphatidylcholine-delective PLD zeta 1 with distinct regulatory domains[J]. Plant Physiology, 2002, 128(3): 1057-1068. doi: 10.1104/pp.010928 [13] Li G, Lin F, Xue H-W. Genome-wide analysis of the phospholipase D family in Oryza sativa and functional characterization of PLDβ1 in seed germination[J]. CELL RESEARCH., 2007, 17(10): 881-894. doi: 10.1038/cr.2007.77 [14] 张艺思, 刘高峰, 刘路平,等. 白菜PLD基因家族全基因组鉴定及对高温胁迫的响应[J]. 西北植物学报, 2019, 39(8):1361-1370. [15] Zhao J, Zhou D, Zhang Q, et al. Genomic analysis of phospholipase D family and characterization of GmPLDαs in soybean (Glycine max)[J]. Journal of Plant Research, 2012, 125(4): 569-578. doi: 10.1007/s10265-011-0468-0 [16] Marek E, Martin P, Fatima C, et al. Molecular diversity of phospholipase D in angiosperms[J]. BMC genomics, 2002, 3:2.DOI: 10.1186/1471-2164-3-2. [17] Wang X, XU L, Zheng L. Cloning and expression of phosphatidylcholine-hydrolyzing phospholipase D from Ricinus communis L [J]. Journalof Biological Chemistry, 1994, 269(32): 20312-20317. [18] Ueki J, Morioka S, Komari T, et al. Purification and characterization of phospholipase D (PLD) from rice (Oryza sativa L.) and cloning of cDNA for PLD from rice and maize (Zea mays L.)[J]. Plant and Cell Physiology, 1995, 36(5): 903-914. doi: 10.1093/oxfordjournals.pcp.a078837 [19] Pappan K, Qin W, Dyer J H, et al. Molecular cloning and functional analysis of polyphosphoinositide-dependent phospholipase D, PLDbeta, from Arabidopsis[J]. Journal of Biological Chemistry, 1997, 272(11): 7055-7061. doi: 10.1074/jbc.272.11.7055 [20] Lein W, Saalbach G. Cloning and direct G-protein regulation of phospholipase D from tobacco[J]. BBA- Molecular and Cell Biology of Lipids, 2001, 1530(2-3): 172-183. doi: 10.1016/S1388-1981(00)00182-7 [21] The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana[J]. Nature, 2000, 408(6814): 796-815. doi: 10.1038/35048692 [22] Yu J, Hu S, Wang J, et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica)[J]. Science, 2002, 296(5565): 79-92. doi: 10.1126/science.1068037 [23] Zhang L, Cai X, Wu J, et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies[J]. Horticulture research, 2018,5: 50. DOI: 10.1038/s41438-018-0071-9. [24] Li F, Fan G, Wang K, et al. Genome sequence of the cultivated cotton Gossypium arboreum[J]. Nature Genetics, 2014, 46(6): 567-572. doi: 10.1038/ng.2987 [25] Ma J, Wan D, Duan B, et al. Genome sequence and genetic transformation of a widely distributed and cultivated poplar[J]. Plant biotechnology journal, 2019, 17(2): 451-460. doi: 10.1111/pbi.12989 [26] Laucou V, Launay A, Bacilieri R, et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs[J]. PLoS One, 2018, 13(2): e0192540. doi: 10.1371/journal.pone.0192540 [27] Schumutz J, Cannon S B, Schlueter J, et al. Genome sequence of the palaeopolyploid soybean[J]. Nature, 2010, 465(7294): 120. doi: 10.1038/nature08957 [28] Liu Q, Zhang C, Yang Y, et al. Genome-wide and molecular evolution analyses of the phospholipase D gene family in Poplar and Grape[J]. BMC Plant Biology, 2010, 10: 117. DOI: 10.1186/1471-2229-10-117. [29] Sang Y, Zheng S, Li W, et al. Regulation of plant water loss by manipulating the expression of phospholipase D alpha[J]. Plant Journal, 2001, 28(2): 135-144. doi: 10.1046/j.1365-313X.2001.01138.x [30] Wang C, Zien C A, Afitlhile M, et al. Involvement of phospholipase D in wound-induced accumulation of jasmonic acid in Arabidopsis[J]. Plant cell, 2000, 12(11): 2237-2246. [31] Zhang W, Wang C, Qin C, et al. The oleate-stimulated phospholipase D, PLDdelta, and phosphatidic acid decrease H2O2-induced cell death in Arabidopsis[J]. Plant cell, 2003, 15(10): 2285-2295. doi: 10.1105/tpc.013961 [32] Pinosa F, Buhot N, Keaaihaal M, et al. Arabidopsis phospholipase dδ is involved in basal defense and nonhost resistance to powdery mildew fungi[J]. Plant physiology, 2013, 163(2): 896-906. doi: 10.1104/pp.113.223503 [33] Premkumar A, Lindberg S, Lager I, et al. Arabidopsis PLDs with C2-domain function distinctively in hypoxia[J]. Physiologia Plantarum, 2019, 167(1): 90-110. doi: 10.1111/ppl.12874 [34] Zhang T, Song Y, Liu Y, et al. Overexpression of phospholipase Dα gene enhances drought and salt tolerance of Populus tomentosa[J]. Chinese Science Bulletin, 2008, 53(23): 3656-3665. doi: 10.1007/s11434-008-0476-1 [35] Ji T, Li S, Li L, et al. Cucumber Phospholipase D alpha gene overexpression in tobacco enhanced drought stress tolerance by regulating stomatal closure and lipid peroxidation[J]. BMC plant biology, 2018,18: 355. DOI: 10.1186/s12870-018-1592-y. [36] Wang J, Ding B, Guo Y, et al. Overexpression of a wheat phospholipase D gene, TaPLDα, enhances tolerance to drought and osmotic stress in Arabidopsis thaliana[J]. Planta, 2014, 240(1): 103-115. doi: 10.1007/s00425-014-2066-6 [37] Tuskan G A, Difazio S, Jansson S, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & amp; Gray)[J]. Science, 2006, 313(5793): 1596-1604. doi: 10.1126/science.1128691 [38] Finn R D, Penelope C, Eberhardt R Y, et al. The Pfam protein families database: towards a more sustainable future[J]. Nucleic Acids Research, 2015, 44(D1): D279-D285. [39] ron M B, Lu S, Andreson J B, et al. CDD: a conserved domaindatabase for the functional annotation of proteins[J]. Nucleic Acids Research, 2011, 39(1): D225-D229. doi: 10.1093/nar/gkq769 [40] unta M, Coggill P, Eberhardt R Y, et al. The Pfam protein families database[J]. Nucleic Acids Research, 2012, 40(D1): D290-D301. doi: 10.1093/nar/gkr1065 [41] Chen C, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13(8): 1194-1202. doi: 10.1016/j.molp.2020.06.009 [42] Widershain G Y. The proteomics protocols handbook[J]. Biochemistry, 2006, 71(6): 861-861. [43] Chou K, Shen H. Cell-PLoc: a package of Web servers for predicting subcellular localization of proteins in various organisms[J]. Nature protocols, 2008, 3(2): 153-162. doi: 10.1038/nprot.2007.494 [44] Kumar S, Stecher G, Li M, et al. MEGA X: molecular evolutionary genetics analysis across computing platforms[J]. Molecular Biology and Evolution, 2018, 35(6): 1547-1549. doi: 10.1093/molbev/msy096 [45] Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 2013, 30(12): 2725-2729. doi: 10.1093/molbev/mst197 [46] Wang L, Hu W, Sun J, et al. Genome-wide analysis of SnRK gene family in Brachypodium distachyon and functional characterization of BdSnRK2.9[J]. Plant Science, 2015, 237: 33-45. doi: 10.1016/j.plantsci.2015.05.008 [47] Blang G, Wolfe K H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes[J]. Plant Cell, 2004, 16(7): 1667-1678. doi: 10.1105/tpc.021345 [48] Hurst L D. The Ka/Ks ratio: diagnosing the form of sequence evolution[J]. Trends in genetics, 2002, 18(9): 486-487. doi: 10.1016/S0168-9525(02)02722-1 [49] Bailey T L, Nadya W, Chris M, et al. MEME: discovering and analyzing DNA and protein sequence motifs[J]. Nucleic Acids Research, 2006, 34(S1): W369-W373. [50] Zhang Y, Liu C, Cheng H, et al. DNA methylation and its effects on gene expression during primary to secondary growth in poplar stems[J]. BMC Genomics, 2020, 21:498. DOI: 10.1186/s12864-020-06902-6. [51] Luz Irina A. Calderón Villalobos, Sarah Lee, Cesar De Oliveira, et al. A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin[J]. Nature Chemical Biology, 2012, 8(5): 477-485. doi: 10.1038/nchembio.926 [52] Yasushi Shimizu Mitao, Tatsuo Kakimoto. Auxin sensitivities of all Arabidopsis Aux/IAAs for degradation in the presence of every TIR1/AFB[J]. Plant and Cell Physiology, 2014, 55(8): 1450-1459. doi: 10.1093/pcp/pcu077 [53] Ghulam Muhammad Ali, Setsuko Komatsu. Proteomic analysis of rice leaf sheath during drought stress[J]. Journal of proteome research, 2006, 5(2): 396-403. doi: 10.1021/pr050291g -

 点击查看大图
点击查看大图
计量
- 文章访问数: 11643
- HTML全文浏览量: 5316
- PDF下载量: 126
- 被引次数: 0
