

-
种群空间分布格局是指种群个体在某一地理分布区水平空间上的分布和配置状况,反映出种群个体在时空上彼此间的相互关系[1-2]。许多研究表明,自然界中植物种群空间分布格局的形成过程较为复杂,同时受环境因子、人为干扰、动物传播能力及种间竞争关系等多种非生物和生物因子综合影响[3-4]。研究植物种群空间分布格局不仅可以了解植物种群的生物学特性、种群间的相互作用及种群与环境间的关系,还可以根据空间分布格局的变化推断出植物种群的更新策略,对探索植物群落生物多样性的形成和维持机制具有重要意义[5]。
喀斯特生境是一种具有植物多样性高但整体资源量低的脆弱生境,同时由于高强度的人为活动如伐木、放牧、农业开垦和采矿等,严重破坏了当地森林植被的完整性,致使许多植物种群呈斑块化分布[6]。虽然喀斯特生境中的植物每年都结实大量的种子为后期植物种群的更新提供充足的种子资源[7],但受喀斯特生境独特地貌形成的地理屏障限制,植物种子很难通过水流、风力和重力等非生物因子进行长距离传播而到达适宜其萌发的微生境中;加之喀斯特生境中土壤浅薄且呈斑块化分布、土壤营养贫瘠和持水性能较差等因素均不利于后期种子的萌发和幼苗的生长,从而对后期植物种群的空间分布产生重要影响[8]。
单性木兰(Kmeria septentrionalis Dandy)隶属于木兰科、单性木兰属,是我国特有的珍稀濒危植物和国家I级重点保护野生植物。目前仅分布于广西罗城县和环江县、贵州荔波县和云南麻栗坡县和马关县等喀斯特石灰岩山地中,且大多数呈零散分布,成片分布区仅见于广西木论国家级自然保护区内[9]。虽然目前相关学者已对单性木兰种群结构进行了一些研究[9-11],但都未涉及到空间分布格局。同时野外调查时发现母树林附近的杂木林和杉木林内分布有大量的更新幼苗,但其周边却没有结实母树分布,而前期的研究也发现成熟的单性木兰种子能吸引27种鸟类对其进行取食和传播[12],因此推测这些幼苗可能是鸟类取食单性木兰种子后将其传播到这两种生境中所形成的。但这3种生境(母树林、杂木林和杉木林)中的幼苗结构和空间分布格局是否存在差异,以及喀斯特生境中哪些环境因子会影响其种群空间分布格局还未可知。本研究以分布于广西木论国家级自然保护区内的单性木兰更新种群为研究对象,通过对上述3种生境中的单性木兰种群结构、静态生命表、存活曲线、空间分布格局及影响其种群空间分布的环境因子进行分析,旨在阐明单性木兰种群更新现状及其对喀斯特生境的适应机制,以期为该物种的科学保护及后期进一步理解喀斯特生境中的气候和栖息地变化对其种群更新的影响等生态学问题提供理论依据。
-
木论国家级自然保护区(107°54′01″~108°05′51″ E; 25°07′01″~25°12′22″ N)位于广西北部的环江县境内,保护区东-西长20.6 km,南-北宽11.6 km,总面积10 829.7 hm2,北部与贵州茂兰国家级自然保护区相连。该区域属于中亚热带季风气候区,年平均温度19.3℃,极端最低温度−5℃,极端最高温度26.7℃。年均降水量1 529.2 mm,降水具有明显的旱雨季之分(雨季:4至8月;旱季:9月至翌年3月),≥ 10℃的年积温4 700~6 300℃,年均无霜期235~290 d,林间相对湿度为79%。保护区属于典型的喀斯特地貌,地表景观主要以峰丛洼地和谷地为主,区内岩石高度裸露,裸露面积高达80%~90%,土壤覆盖面积不足20%且土壤仅分布于岩石缝隙间,只有洼地或谷地才有成片土壤分布[9]。植被类型主要为分布不均匀的常绿落叶阔叶混交林,常见植物有柞木(Xylosma racemosum (Sieb. et Zucc.) Miq.)、栓叶安息香(Styrax suberifolius Hook. et Arn.)和小叶青冈(Cyclobalanopsis myrsinifolia (Blume) Oersted)等[13]。
-
2019年7—8月,分别在母树林、杂木林和杉木林3种生境中各设置1个50 m × 50 m的固定样方,然后按照相邻格子法将其划分为300个5 m × 5 m 的小样方,对样方内所有单性木兰更新幼苗的高度、微生境类型及经纬度进行测量,同时以样方西南角为原点,东-西方向为横轴(X),南-北方向为纵轴(Y),采用水平坐标(X, Y)的方式记录各幼苗在样方中的相对坐标[14]。对各幼苗所在位置的郁闭度(目测)、岩石裸露率(网格法测定)、海拔(手持 GPS)、坡向和坡度(罗盘)、环境温度和土壤湿度(温湿度计,型号:GM1363,深圳聚茂源科技有限公司)及光照(光照度计,型号:GM1040,深圳聚茂源科技有限公司)进行测量。为降低人为因素对实验结果的影响,所有的野外工作都是在晴天进行,同时林内光照、温度和土壤湿度等指标都是选择在中午12:00—14:00 阳光最旺盛的时候进行测量。
-
(1)年龄结构
由于单性木兰幼苗个体较小且喀斯特生境不利于其生长,很难通过胸径的大小来推断其年龄结构,因此本研究中采用幼苗高度来反映单性木兰种群的年龄结构[15]。根据实际调查情况及单性木兰幼苗的生长特点将其年龄结构分为6个龄级:0 < H ≤ 10 cm(I);10 cm < H ≤ 20 cm(II);20 cm< H ≤ 30 cm(III);30 cm< H ≤ 40 cm(IV);40 cm< H ≤ 50 cm(V);H > 50 cm(VI)。
(2)静态生命表和存活曲线
参照胡刚等[16]的方法编制单性木兰更新种群静态生命表,各参数具体含义如下:(1)a表示幼苗的龄级;(2)Na表示a在某类龄级中出现的个体数;(3)La表示为标准化的幼苗存活数即La = (Na+Na+1)/2;(4)lgLa表示La取以10为底的对数;(5)Da表示从a到a+1龄级幼苗的死亡数即Da = Na−Na+1;(6)Qa表示从a到a+1龄级幼苗的死亡率即Qa = Da/Na+1;(7)Ta表示从a龄级至超过a龄级的存活个体总数即Ta = ∑La;(8)Ea(生命期望)表示第a龄级的个体在未来所能存活的平均年数即Ea = Ta/Na。后以α为横坐标、LαlgLα为纵坐标,绘制单性木兰更新种群存活曲线。采用指数函数和幂函数对幼苗存活数和龄级间的关系进行拟合,后根据回归系数(R2)的大小来判定模型拟合效果。若指数函数Nα = N0e−bx拟合效果好,则说明更新种群存活曲线更接近Deevy-II型;若幂函数Nα = N0x−b拟合效果好,则说明更新种群存活曲线更接近Deevy-III型[17]。
(3)点格局分析
采用Programita软件对不同生境中的单性木兰更新种群的点格局进行分析。首先将幼苗的相对坐标数据导入Programita软件中计算出不同尺度下的函数值,再以距离尺度作为横坐标,上下包迹线作为纵坐标绘制函数图像。若函数值在上下包迹线间,则表明更新种群呈随机分布;若函数值大于上包迹线,则表明更新种群呈集群分布;若函数值处于下包迹线下方,则表明更新种群呈均匀分布[14,18]。
-
由于坡向数据都是0°~360°罗盘测量值,为使数据在模型中表现出较好的拟合性,本研究参照刘瑞雪等[19]的方法将坡向划分为5个等级并分别赋值,1表示北坡(0°~22.5°和337.5°~360°);2表示东北坡(22.5°~67.5°)和西北坡(292.5°~337.5°);3表示东坡(67.5°~112.5°)和西坡(247.5°~292.5°);4表示东南坡(112.5°~157.5°)和西南坡(202.5°~247.5°);5表示南坡(157.5°~202.5°)。
-
采用Kolmogorov-Smirnov对所有数据变量进行正态性检验,对不符合正态分布的数据先进行转换后再检验。采用One Sample T-test对不同龄级间的幼苗数量差异进行分析;采用Mann-Whitney U检验分别对两种生境中不同龄级间的幼苗数量差异进行分析;采用Kruskal-Wallis检验对不同生境中、不同龄级幼苗数量的差异进行分析。由于环境因子间的数量级差异较大,为更好的对数据进行拟合将所有的数值进行以10为底的对数转换,再以各样方内幼苗的平均密度作为因变量,各环境因子的平均值作为自变量,后再以前两者为固定因子,生境为随机因子,采用广义线性混合模型(Generalized Linear Mixed Model, R package lme4)对影响单性木兰种群空间分布的主要环境因子进行分析。所有的数据都是在SPSS 20.0上进行处理,差异显著水平设定为P < 0.05。采用R语言(3.6.1, R Core Team, 2016)进行作图。
-
本次共调查到747株幼苗,不同龄级的幼苗数量存在极显著差异(t = 5.049, df = 5, P = 0.004),其中II级幼苗的数量最多(201株),占总数量的26.91%(图1)。不同生境中,母树林内幼苗的数量最多462株,其次为杂木林215株、杉木林70株,分别占总数量的61.85%、28.78%和9.37%。不同龄级的幼苗数量在所有生境中都存在显著差异(母树林:t = 2.784, df = 5, P = 0.039; 杂木林:t = 2.793, df = 5, P = 0.038; 杉木林:t = 2.809, df = 5, P = 0.038)。不同龄级的幼苗数量在母树林和杂木林间不存在显著差异(Z = −1.043, P = 0.297),但在母树林和杉木林间(Z = −2.567, P = 0.01)及杂木林和杉木林间都存在显著差异(Z = −2.330, P = 0.02)。在整体上,不同生境中、不同龄级的幼苗数量存在显著差异(χ2 = 8.802, df = 2, P = 0.012),个体数量随着幼苗龄级的增加而逐渐降低,种群结构呈倒“J”型分布(图1)。调查过程中还发现,97.19%的幼苗都是生长在土面微生境中,而仅有2.00%和0.80%的幼苗分别生长在石槽和石缝微生境中。
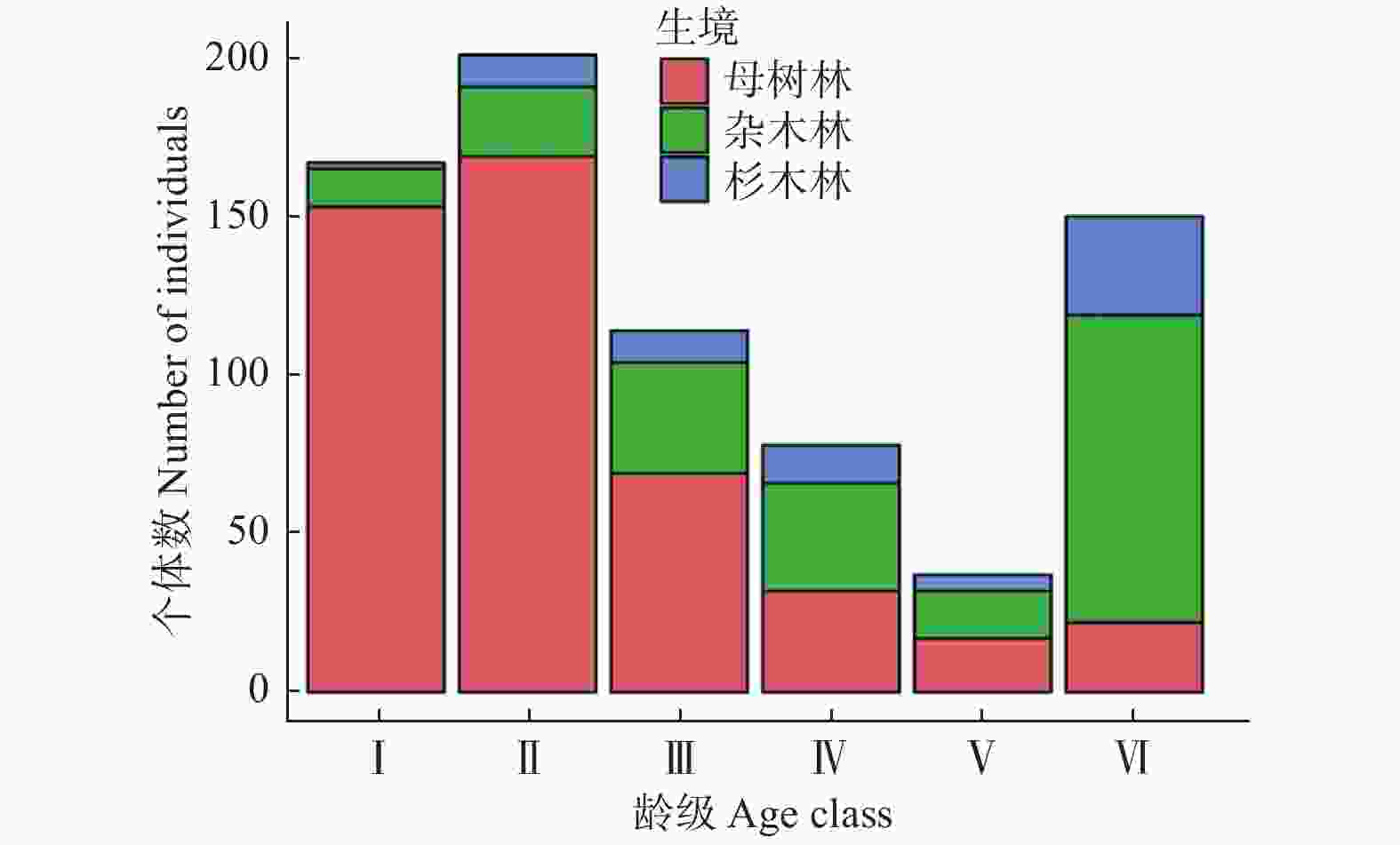
图 1 不同生境单性木兰更新种群年龄结构
Figure 1. Age structure of regeneration population of Kmeria septentrionalis in different habitats
-
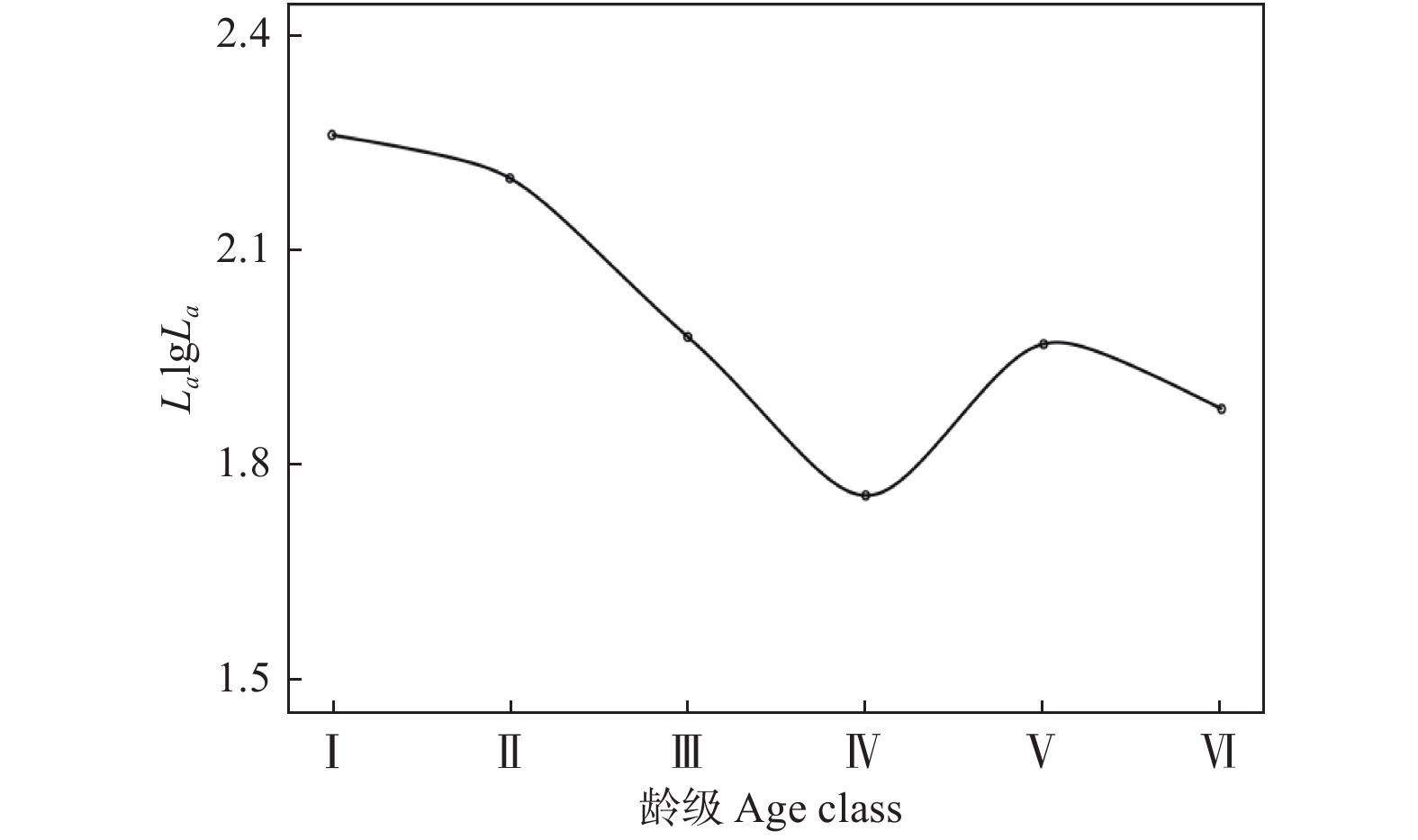
不同龄级的幼苗个体存活数量变化波动较大,其中Ⅱ-Ⅲ级的幼苗存活数量下降最为严重,说明该阶段中的幼苗对环境适应能力极弱,大量幼苗经环境筛选作用后逐步消失。Ⅳ-Ⅴ级的幼苗存活数量下降趋势减缓,说明该阶段中的幼苗生长及生存能力逐渐增强,种内和种间对有限环境资源的竞争可能是导致该阶段幼苗存活数量下降的主要原因。不同龄级的幼苗生命期望值波动幅度较大,生命期望值在V级时达到最高值(5.05)、死亡率降到最低(−0.75),说明该阶段中存活的幼苗个体应对外界不良环境的抵抗能力及对有限环境资源的竞争能力更强,幼苗的质量最优(表1)。
表 1 单性木兰更新种群静态生命表
Table 1. Static life tables of regeneration population of Kmeria septentrionalis
龄级 存活数 Na 标准化存活数 La 标准化存活数对数 LalgLa 死亡数 Da 死亡率 Qa 存活总数 Ta 生命期望 Ea Ⅰ 167 184.00 2.26 −34 −0.17 747 4.47 Ⅱ 201 157.50 2.20 87 0.76 580 2.89 Ⅲ 114 96.00 1.98 36 0.46 379 3.32 Ⅳ 78 57.50 1.76 41 1.11 265 3.40 Ⅴ 37 93.50 1.97 −113 −0.75 187 5.05 Ⅵ 150 75.00 1.88 150 1.00 150 1.00 存活曲线拟合结果表明,幂函数(y = 2.2787x−0.119, R2 = 0.693)的拟合效果优于指数函数(y = 2.294 6e−0.039x, R2 = 0.602),表明单性木兰种群存活曲线更接近Deevy-Ⅲ型。存活曲线从Ⅰ到Ⅳ段、Ⅴ到Ⅳ段存活曲线出现下降,说明该阶段的幼苗更新受到阻碍(图2)。
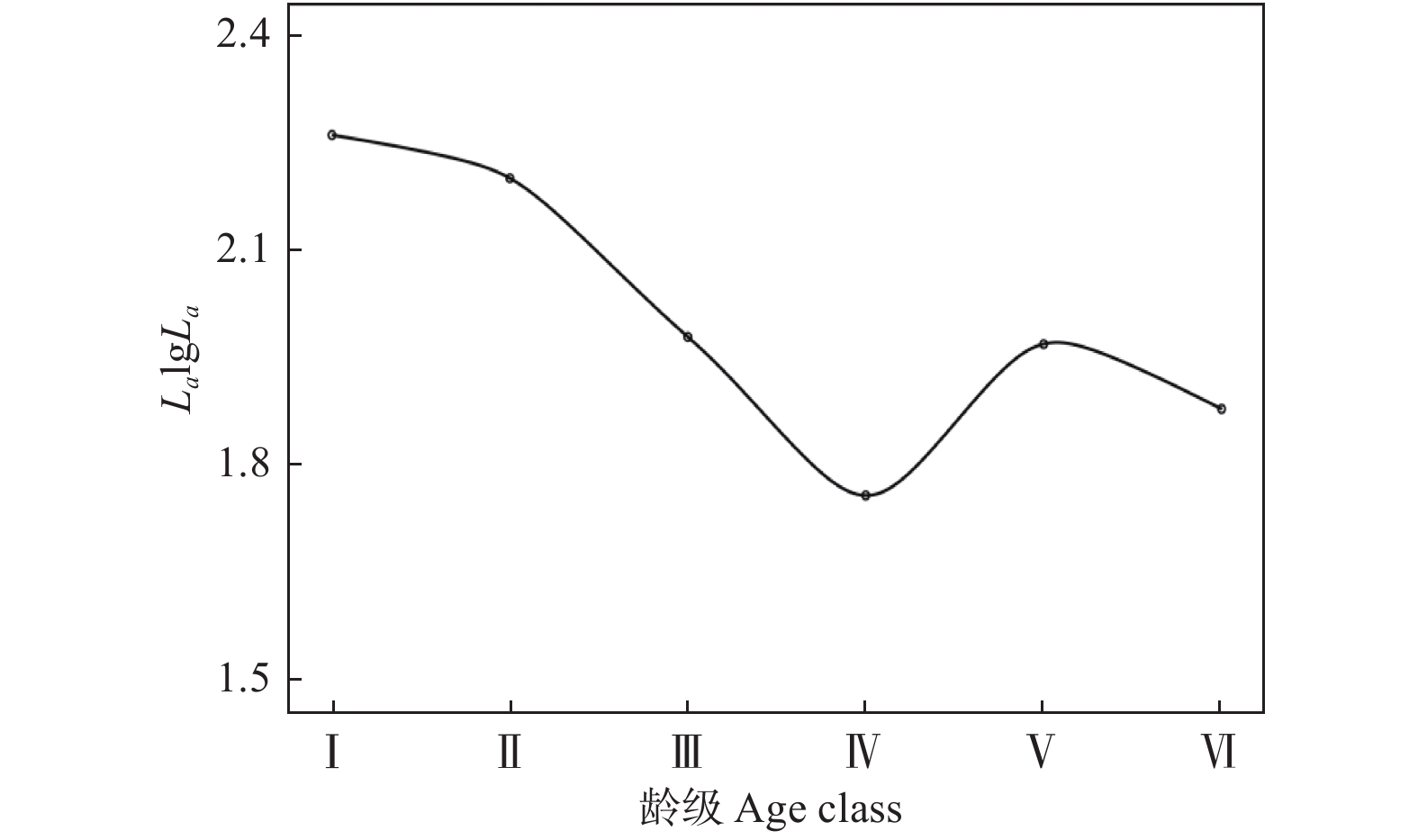
图 2 单性木兰更新种群存活曲线
Figure 2. Survival curves of regeneration population of Kmeria septentrionalis
-
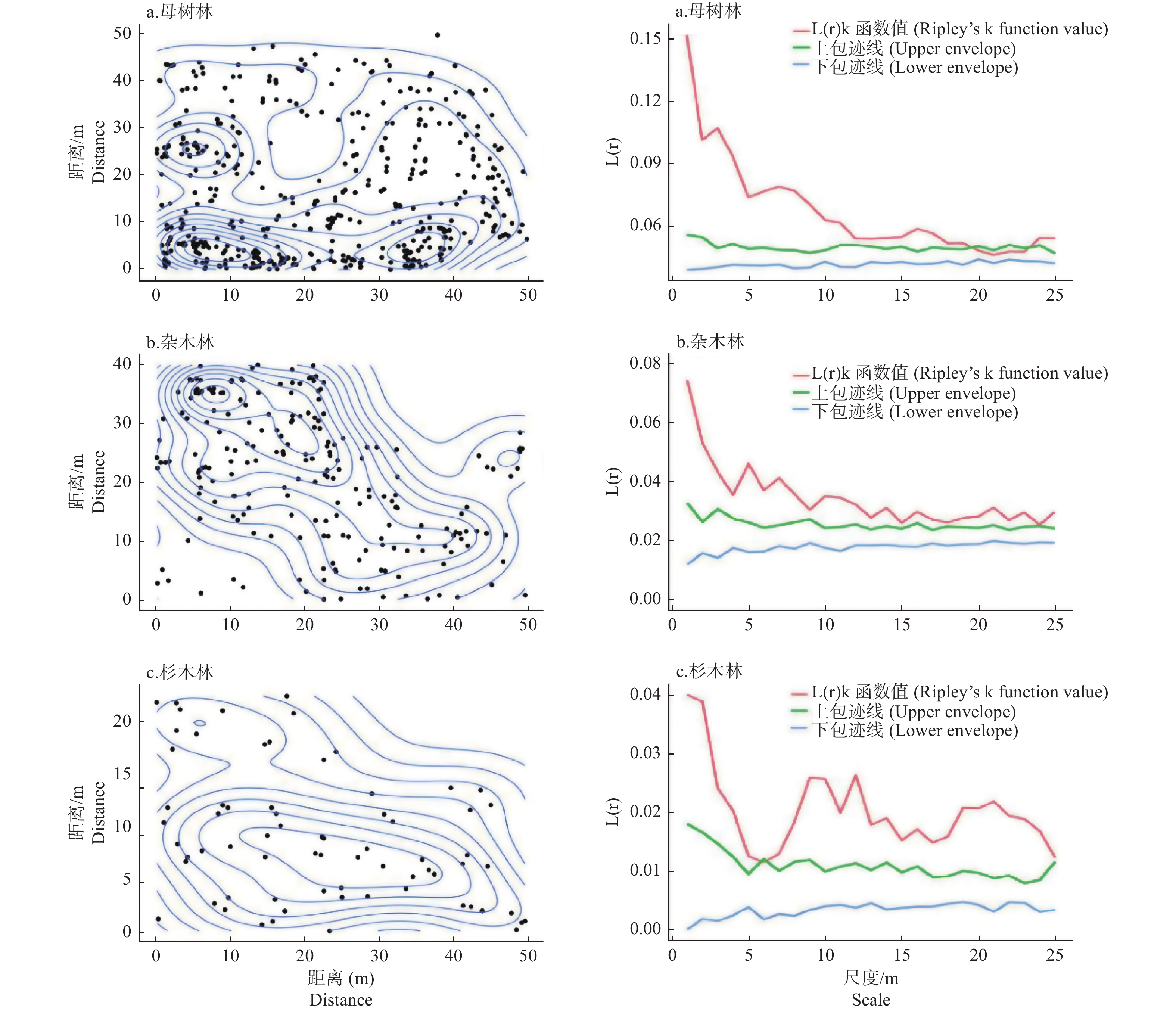
各生境中单性木兰更新种群的空间分布并不均匀,但整体上都呈聚集分布。在母树林内,除20~23 m间有微弱的随机分布特征外,其余所有尺度上都呈聚集分布;在杂木林内,所有尺度上幼苗都呈聚集分布;在杉木林内,除6 m有微弱的随机分布特征外,其余所有尺度上都呈聚集分布(图3)。
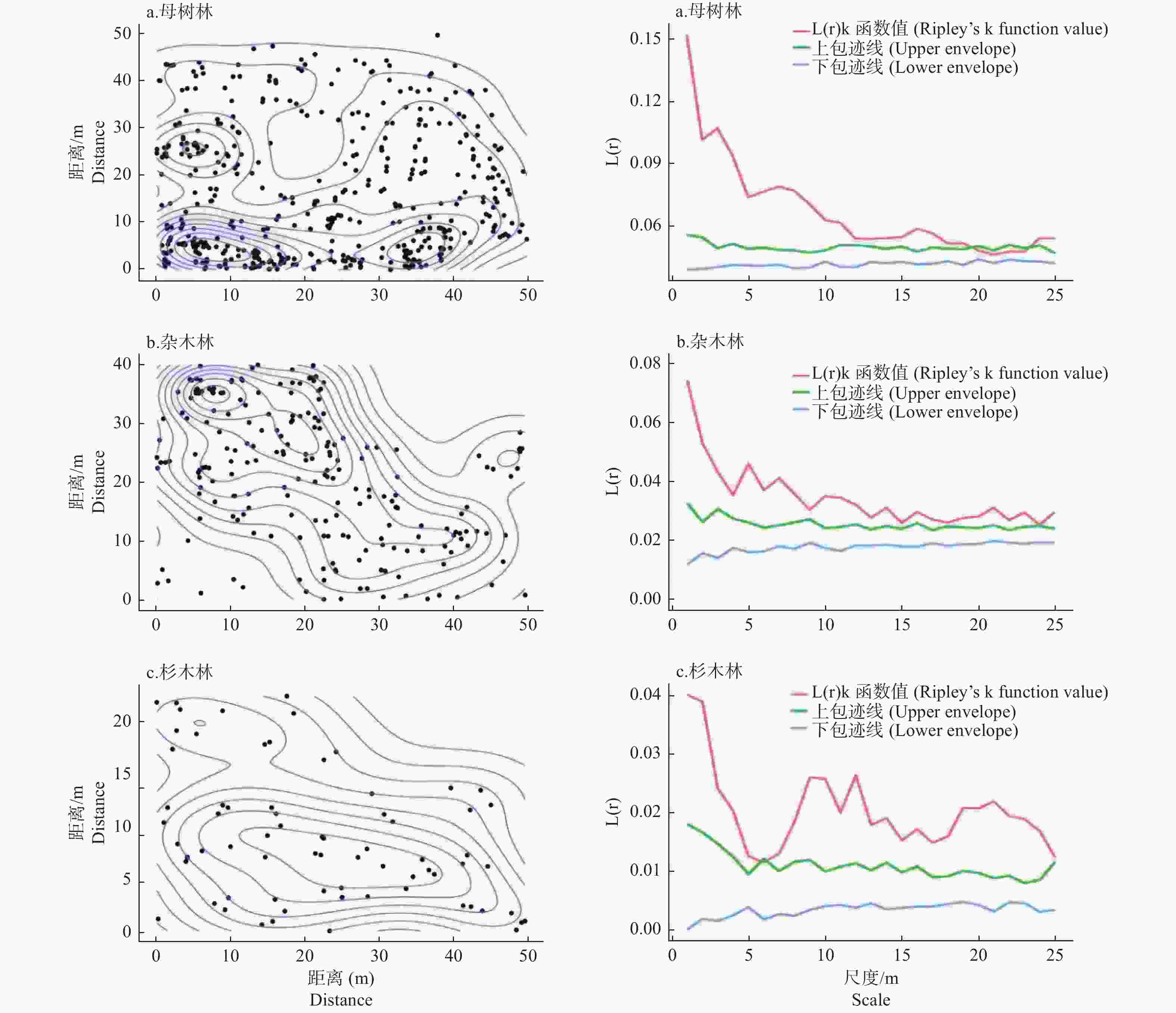
图 3 不同生境中单性木兰更新种群空间分布格局
Figure 3. Spatial distribution of regeneration population of Kmeria septentrionalis indifferent habitat
-
采用广义混合线性模型对各样方内更新幼苗的密度与环境因子间的关系进行拟合,发现土壤湿度、海拔、光照与更新幼苗的密度呈正相关,而坡向、坡度、郁闭度、温度及岩石裸露率与更新幼苗的密度呈负相关;但仅土壤湿度、郁闭度和岩石裸露率与更新幼苗的密度达到了显著差异水平,说明这3种环境因子是影响单性木兰更新种群空间分布的主要因素(表2)。
表 2 基于广义线性混合模型分析单性木兰更新种群空间分布与环境因子的关系
Table 2. Analysis of the relationship between population spatial distribution andenvironmental factors of Kmeria septentrionalis based on generalized linear mixed model
变量 估计值 标准误差 t p 截距 Intercept −4.070 4.990 −0.816 0.416 海拔/m 0.216 1.563 0.138 0.890 坡度/℃ −0.084 0.088 −0.955 0.341 坡向/° −0.043 0.071 −0.609 0.544 郁闭度/% −2.543 0.138 −18.407 P < 0.001*** 光照/Lax 0.052 0.030 1.739 0.084 湿度/℃ 1.201 0.421 2.849 0.005*** 温度/℃ −0.096 1.697 −0.057 0.955 岩石裸露率/% −0.162 0.073 −2.213 0.028** -
研究植物种群结构对摸清植物种群与环境因子间的相互关系及不同龄级个体在植物种群更新中的作用具有重要意义[20]。本研究中,单性木兰更新种群中有大量低龄级的幼苗个体储备,说明该种群属于缓慢增长型种群,具有一定维持自我更新的能力(图1)。大量低龄级幼苗的分布可能与单性木兰种群为应对恶劣的喀斯特生境所采取的更新策略有关。由于喀斯特生境岩石裸露率高、土壤营养贫瘠且呈斑块化分布[21],严重制约了单性木兰种子的萌发和幼苗的更新,大量低龄级个体的分布能使其种群最大程度的占据有利的空间环境资源,以便为后期向高龄级幼苗的转化积累更多的生物能量。但随着幼苗高度的增加,具有相同资源利用方式的不同个体间对有限环境资源(光照、水分和营养等)的竞争不断增大,加之喀斯特生境不利于幼苗的生长,从而在一定程度上造成低龄级幼苗个体生长发育迟滞或死亡(自疏效应),最终形成低龄级幼苗个体数量充足而高龄级幼苗个体数量缺乏的格局[22]。潘春柳等[9]、田淑娟[10]及张启伟等[11]的研究也得出相似的结果。不同生境中,母树林内更新幼苗数量最多(图1),说明每年都有大量的种子没有被动物传播到其它生境中而是直接沉积在母树树冠层内进行萌发;而杂木林和杉木林中更新幼苗数量较少,可能与鸟类传播有效性和取食后的生境选择偏好导致这两种生境获得的种源不足有关。但从种群可持续发展的角度来看,杂木林和杉木林内成片的土壤分布及较高的含水率能为早期种子的萌发和幼苗的生长提供有利的条件,随着时间的延长可能更有利于单性木兰种群的持续更新。
本研究中,单性木兰幼苗的存活数量随着幼苗龄级的增加而呈下降趋势(表1),说明研究区域中的单性木兰种群更新与生长受到一定程度的种内、种间竞争和强烈的环境胁迫效应。这种变化可能是由于随着龄级的增加幼苗个体对营养物质和生存空间的需求不断增大,加之喀斯特生境岩石裸露率高,土壤浅薄而空间分布不均匀[21],高密度的幼苗分布会加剧种内不同个体间对土壤生长空间、矿质物、水分和光照等环境资源的竞争致使幼苗产生自疏效应,最终导致仅有少量竞争能力较强的幼苗进入下一生长发育阶段。Ⅴ级幼苗的生命期望值最大,说明经过激烈的种内、种间竞争及环境筛选作用后幼苗个体的生理生态特征发育逐渐成熟,抵御外界不良环境的胁迫能力逐步提高。但整体上单性木兰更新种群生命期望值随着幼苗龄级的增加而降低,表明随着龄级的增加其更新种群的生存能力越来越弱。单性木兰更新种群的存活曲线属于Deevey-Ⅲ型(图2),说明其种群更新状态稳定,这与张启伟等[11]对其它片区的研究结果相似。
种群空间分布格局是对植物种群水平空间分布的定量描述,对其特征进行研究有助于揭示种群空间分布格局的形成机制及其与环境因子间的相互关系[23-24]。本研究中,3种生境中的单性木兰更新种群除在小尺度上有微弱的随机分布特征外,其余所有尺度上都呈聚集分布(图3),这种高强度的聚集分布可能与喀斯特生境地形特征和鸟类的传播行为有关。母树林内高度裸露的岩石形成的地理屏障会限制植物种子的长距离传播[22-23],致使大部分成熟的单性木兰种子会直接沉积在母树树冠层内;加之喀斯特生境地形起伏变化大、土壤浅薄且呈斑块化分布,光照、营养和水分等生态因子的再分配呈现出较高的空间异质性以致在空间上形成更多的微生境隔离[23,25],降低了单性木兰种子到达适宜萌发微生境中的概率,从而易使后期萌发的幼苗呈聚集分布。同一地区的其它研究结果也认为喀斯特地形(海拔、坡度和坡向)是影响当地植物种群空间分布的主要因素[26-27]。虽然杂木林和杉木林内成片的土壤分布有利于单性木兰幼苗的更新,但受鸟类传播者自身肠道容量的限制,其常会将种子快速的排出体外,以减轻飞翔时的体重负担和能量消耗,从而导致后期萌发的幼苗也易在小尺度空间上形成聚集分布格局[28]。此外,聚集分布也可能是单性木兰种群为应对恶劣的喀斯特环境所采取的一种适应策略。虽然聚集分布会导致幼苗种群在空间尺度上呈斑块性和镶嵌性分布[10],会加剧种内不同个体间对有限环境资源的竞争,但另一方面这种聚集分布也有利于发挥群体效应,能加快区域贫瘠生境中的营养累积及死后的营养元素反馈,使周围的环境条件更有利于更新幼苗的生长,从而提高植物种间的竞争力及其抵抗外界不良环境条件的能力[22]。
环境因子(地形、土壤及气候等)对植物种群空间分布、物种多样性的形成和维持具有重要影响[3]。本研究中,郁闭度、土壤湿度和岩石裸露率是影响单性木兰更新种群空间分布格局的主要环境因子(表2)。郁闭度主要通过控制光照的渗透率和紫外线强度来改变林下植被的光合速率及周围土壤、空气中的温度和湿度来影响植物的生长发育和空间分布[29]。野外调查过程中也发现大量的单性木兰更新幼苗的叶片因缺乏光照而黄化死亡。喀斯特生境中常年缺乏地表径流,降雨是该地区植物生长所需水分的主要来源。但由于喀斯特生境中降雨时空差异大,地下溶洞发达且渗漏性强,降雨时大部分地表水流会直接沿着溶蚀裂隙、溶洞等向地下快速的渗透,导致存留在地表土壤和植物根系中的水分含量非常低[30],喀斯特高度异质性的地形会再次调控土壤中水分的分配过程导致土壤含水量存在明显的空间差异[27],从而影响单性木兰种群的空间分布。单性木兰种子中含有大量的挥发性芳香物质[31],高度裸露的岩石会降低种子进入土壤中的速度,延长其暴露在外界环境中的时间从而提高其被啮齿动物大量捕食的风险;岩石高度裸露的区域中土壤面积稀缺、土壤营养和含水率频繁不利于单性木兰种群的更新。野外调查中发现几乎所有的幼苗都是生长在土面微生境中,土面微生境较裸岩生境更能提供充足的营养物质和水分含量,为种子的萌发和幼苗的生长提供良好的条件,从而导致幼苗的密度与岩石裸露率呈显著负相关。
喀斯特生境中濒危植物单性木兰种群结构及空间分布格局研究
Population Structure and Spatial Distribution Pattern of Kmeria septentrionalis an Endangered Species, in Karst Habitat
-
摘要:
目的 以分布在喀斯特生境中的濒危植物单性木兰自然更新种群为研究对象,探究其种群年龄结构、空间分布格局及环境影响因素。 方法 采用点格局对单性木兰更新种群的空间分布格局进行分析,利用广义混合线性模型对幼苗的密度与环境因子间的关系进行线性拟合。 结果 (1)研究区域中的单性木兰种群结构呈倒“J”型,种群存活曲线接近Deevey-III型,大量小龄级幼苗的分布预示其种群更新状况良好;(2)单性木兰种群除在小尺度上有微弱的随机分布外,其余所有尺度上都呈聚集分布;(3)郁闭度、土壤湿度和岩石裸露率是影响单性木兰种群空间分布的主要因素。 结论 研究区域中的单性木兰更新种群状态良好,高强度的空间聚集分布主要是种子生物学特性和喀斯特生境特征共同作用的结果。 Abstract:Objective Taking the natural regeneration population of endangered plant Kmeria septentrionalis in karst habitat as object to study the population regeneration structure, spatial distribution pattern and environmental impact factors. Method Point pattern and generalized mixed linear model were used to complete a linear fitting of the spatial distribution pattern and environmental impact factors of the regeneration population of K. septentrionalis. Result (1) The population structure of K. septentrionalis was in an inverted “J” type, the survival curve was close to Deevey-Ⅲ; the existence of a large number of young-aged seedlings indicated that the regeneration condition of K. septentrionalis population was good. (2) Except for the weak random distribution on a small scale, the other scales were in a clustered distribution. (3) The canopy density, soil moisture and rock exposure rate were the main factors affecting the spatial distribution of K. septentrionalis. Conclusion The condition of the regeneration population of K. septentrionalis in the study area is good, and the high-intensity spatial aggregation distribution is mainly the combined action of the seed biological characteristics and karst habitat. -
图 1 不同生境单性木兰更新种群年龄结构
Figure 1. Age structure of regeneration population of Kmeria septentrionalis in different habitats
图 2 单性木兰更新种群存活曲线
Figure 2. Survival curves of regeneration population of Kmeria septentrionalis
图 3 不同生境中单性木兰更新种群空间分布格局
Figure 3. Spatial distribution of regeneration population of Kmeria septentrionalis indifferent habitat
表 1 单性木兰更新种群静态生命表
Table 1. Static life tables of regeneration population of Kmeria septentrionalis
龄级 存活数 Na 标准化存活数 La 标准化存活数对数 LalgLa 死亡数 Da 死亡率 Qa 存活总数 Ta 生命期望 Ea Ⅰ 167 184.00 2.26 −34 −0.17 747 4.47 Ⅱ 201 157.50 2.20 87 0.76 580 2.89 Ⅲ 114 96.00 1.98 36 0.46 379 3.32 Ⅳ 78 57.50 1.76 41 1.11 265 3.40 Ⅴ 37 93.50 1.97 −113 −0.75 187 5.05 Ⅵ 150 75.00 1.88 150 1.00 150 1.00  下载: 导出CSV
下载: 导出CSV
表 2 基于广义线性混合模型分析单性木兰更新种群空间分布与环境因子的关系
Table 2. Analysis of the relationship between population spatial distribution andenvironmental factors of Kmeria septentrionalis based on generalized linear mixed model
变量 估计值 标准误差 t p 截距 Intercept −4.070 4.990 −0.816 0.416 海拔/m 0.216 1.563 0.138 0.890 坡度/℃ −0.084 0.088 −0.955 0.341 坡向/° −0.043 0.071 −0.609 0.544 郁闭度/% −2.543 0.138 −18.407 P < 0.001*** 光照/Lax 0.052 0.030 1.739 0.084 湿度/℃ 1.201 0.421 2.849 0.005*** 温度/℃ −0.096 1.697 −0.057 0.955 岩石裸露率/% −0.162 0.073 −2.213 0.028**
下载: 导出CSV
-
[1] 袁春明, 孟广涛, 方向京, 等. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布[J]. 生态学报, 2012, 32(12):3866-3872. [2] 舒 兰, 刘兆刚, 董灵波. 帽儿山天然次生林内主要木本植物空间格局及更新特点[J]. 应用生态学报, 2019, 30(6):1945-1955. [3] Tingstad L, Olsen S L, Klanderud K, et al. Temperature, precipitation and bioticinteractions as determinants of tree seedling recruitment across the tree lineecotone[J]. Oecologia, 2016, 180(3): 917-918. doi: 10.1007/s00442-015-3535-8 [4] 刘梦婷, 王振锡, 王雅佩, 等. 新疆天山云杉林群落分布格局及环境解释[J]. 林业科学研究, 2019, 32(6):90-98. [5] 李文英, 李 欣, 甘小洪. 濒危植物水青树的种群结构与数量动态[J]. 亚热带植物科学, 2018, 47(3):26-32. [6] Zhang Z H, Hu G, Zhu J D, et al. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China[J]. Journal of Plant Ecology, 2013, 6(2): 131-140. doi: 10.1093/jpe/rts027 [7] 唐樱殷, 谢永贵, 余刚国, 等. 黔西北喀斯特土壤种子库季节动态及种子库对策[J]. 生态学杂志, 2011, 30(7):1454-1460. [8] Du H, Hu F, Zeng F P, et al. Spatial distribution of tree species inevergreen-deciduous broadleaf karst forests in southwest China[J]. Scientific Reports, 2017, 7(1): 15664. [9] 潘春柳, 赖家业, 黎向东, 等. 单性木兰种子雨与天然更新的初步调查[J]. 生态学杂志, 2008, 27(12):2235-2239. [10] 田淑娟. 珍稀濒危植物单性木兰生理生态特征及种群更新研究[D]. 贵阳, 贵州大学, 2010. [11] 张启伟, 林燕芳, 覃文更, 等. 濒危植物单性木兰种群径级结构和生命表分析[J]. 广西植物, 2013, 33(3):291-294. doi: 10.3969/j.issn.1000-3142.2013.03.002 [12] Wang G H, Yang Z X, Chen P, et al. Seed dispersal of an endangered Kmeria septentrionalis by frugivorous birds in a karst habitat[J]. Pakistan Journal of Zoology, 2019, 51(3): 1195-1198. [13] 彭玉华, 何琴飞, 谭长强, 等. 广西极小种群单性木兰群落空间结构量化[J]. 生态学杂志, 2016, 35(2):362-369. [14] 张晓晨, 赵 洋, 熊中人, 等. 宝华山青冈种群年龄结构及点格局分析[J]. 南京林业大学学报: 自然科学版, 2018, 42(6):81-87. [15] 欧芷阳, 苏志尧, 彭玉华, 等. 桂西南喀斯特山地蚬木幼龄植株的天然更新[J]. 应用生态学报, 2013, 24(9):2440-2446. [16] 胡 刚, 黎 洁, 覃盈盈, 等. 广西北仑河口红树植物种群结构与动态特征[J]. 生态学报, 2018, 38(9):3022-3034. [17] 陈国鹏, 鲜骏仁, 曹秀文, 等. 林窗对岷江冷杉幼苗生存过程的影响[J]. 生态学报, 2016, 36(20):6475-6486. [18] 李小东, 丁 晖, 吴 俊, 等. 黄山常绿阔叶林优势种甜槠的种群动态与点格局分析[J]. 西北植物学报, 2018, 38(11):2110-2119. [19] 刘瑞雪, 陈龙清, 史志华. 丹江口水库水滨带植物群落空间分布及环境解释[J]. 生态学报, 2015, 35(4):1208-1216. [20] Paul A, Khan M L, Das A K. Population structure and regeneration status of rhododendrons in temperate mixed broad-leaved forests of western Arunachal Pradesh, India[J]. Geology, Ecology, and Landscapes, 2019, 3(3): 168-186. doi: 10.1080/24749508.2018.1525671 [21] 郑生猛, 盖爽爽, 谢 强, 等. 桂西北喀斯特峰丛洼地典型灌丛植物种间关系及环境解释[J]. 农业现代化研究, 2016, 37(6):1198-1205. [22] 涂洪润, 李娇凤, 刘润红, 等. 桂林岩溶石山檵木种群空间格局及其关联性[J]. 应用生态学报, 2019, 30(8):2621-2630. [23] Guo Y L, Wang B, Li D X, et al. Effects of topography and spatial processes on structuring tree species composition in a diverse heterogeneous tropical karst seasonal rainforest[J]. Flora, 2017, 231: 21-28. doi: 10.1016/j.flora.2017.04.002 [24] Omelko A, Ukhvatkina O, Zhmerenetsky A, et al. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pine-broadleaved forest[J]. Forest Ecologyand Management, 2018, 411: 46-66. doi: 10.1016/j.foreco.2018.01.023 [25] 张忠华, 胡 刚, 倪 健. 茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征[J]. 生态学报, 2015, 35(24):8221-8230. [26] 金俊彦, 覃文更, 罗柳娟, 等. 濒危植物单性木兰群落优势种群生态位研究[J]. 广西植物, 2013, 33(3):300-305. doi: 10.3969/j.issn.1000-3142.2013.03.004 [27] 兰斯安, 宋 敏, 曾馥平, 等. 木论喀斯特森林木本植物多样性垂直格局[J]. 生态学报, 2016, 36(22):7374-7383. [28] Masaki T, Nakashizuka T, Niiyama K, et al. Impact of the spatial uncertainty of seed dispersal on tree colonization dynamics in a temperate forest[J]. Oikos, 2019, 128(12): 1816-1828. doi: 10.1111/oik.06236 [29] Wagner S, Fischer H, Huth F. Canopy effects on vegetation caused by harvesting and regeneration treatments[J]. European Journal of Forest Research, 2011, 130(1): 17-40. doi: 10.1007/s10342-010-0378-z [30] Liu C C, Liu Y G, Guo K, et al. Exploitation of patchy soil water resources by the clonal vine Ficustikoua in karst habitats of southwestern China[J]. Acta Physiologiae Plantarum, 2011, 33(1): 93-102. [31] 黄品鲜, 周永红, 赖家业, 等. 珍稀濒危植物单性木兰种皮的挥发性成分分析[J]. 广西植物, 2010, 30(5):691-695. doi: 10.3969/j.issn.1000-3142.2010.05.022 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4693
- HTML全文浏览量: 3122
- PDF下载量: 71
- 被引次数: 0
