


-
随着种质资源收集与保存的发展,种质资源库不断扩大,这为种质保存、评价和利用带来了一定难度[1]。为此,Frankel等提出了核心种质的概念[2]。其目的是以最少的种质资源数量最大程度的代表原种质的遗传多样性,从而在遗传资源保护的过程中,大幅降低保存成本,提高效率。同时便于开展资源评价研究,减少工作量,降低筛选和挖掘难度[3],促进种质资源的合理开发与利用[4-5]。目前,国内外已经构建出了多种农作物的核心种质[6-8],木本植物也得到了越来越多的重视,已成功构建核心种质的树种包括苹果[9-10](Malus pumila Mill.)、猕猴桃[11](Actinidia chinensis Planch.)、茶树[12](Camellia sinensis (L.) O. Ktze.)、刺槐[13](Robinia pseudoacacia L.)、白桦[14](Betula platyphylla Suk.)、美洲黑杨[15](Populus deltoides Marsh.)和水青树[16](Tetracentron sinense Oliv.)等。
灰楸(Catalpa fargesii Bur.)为紫葳科(Bignoniaceae)梓属(Catalpa)乔木,材质优良,木材细致,且根系发达,具有很强的固土、抗风、耐旱耐寒能力,为我国特有的珍贵用材树种[17-18];其生态分布幅度较大,主要分布于陕西省、甘肃省、河北省、河南省等地[18]。以往的研究都集中在灰楸的种子发芽特性[19]、组织培养[20]、嫁接育苗[20-21]、抗性评价[22-23]、种质多样性研究[18]及优良种质的选择[24]方面。李秀兰等[25]曾将267个灰楸种质基于生长和叶部性状,通过欧氏距离和离差平方和法,进行随机取样,初步构建了63份核心种质。然而,在核心种质的构建过程中,不同构建策略的效果不同。如在选择遗传距离时,所测性状量纲单位不一致会影响欧氏距离,但不影响马氏距离[26];在选择取样方法时,随机取样法能更好的保留原始种质的群体结构,但不利于核心种质的代表性和多样性;偏离度取样法能保留原种质的变异程度和遗传多样性;优先取样法具有保留特殊或有价值种质的优点[27-28]。因此,选择最适的核心种质构建策略尤为重要。
本研究对11~13年生灰楸的生长、叶部和材性性状的遗传变异与多样性进行分析,基于不同遗传距离、不同系统聚类方法、不同取样方法和取样比例构建灰楸初级核心种质资源,并对不同方法构建的核心种质进行评价,最终选择最适合灰楸的核心种质构建策略,得出较为可靠的灰楸初级核心种质群体,对解决其种质资源保存成本较高问题,促进灰楸种质资源的鉴定和有效利用奠定基础。
-
2007年,在甘肃省、陕西省、山西省和河南省开展灰楸种质资源全面调查收集,收集材料为1年生和2年生小枝,共收集267株优良单株。2008年,在甘肃省天水市麦积区嫁接扩繁(无性系化),每株嫁接扩繁36株。2009年,在甘肃省小陇山林业实验局林业科学研究所沙坝实验基地营建种质资源对比试验林,采用随机完全区组设计,4次重复,共200份种质,其中,甘肃省163份、陕西省4份、山西省22份、河南省11份。
-
试验地位于甘肃省天水市麦积区(105°53′24.468″ E,4°34′14.484″ N),隶属于甘肃省小陇山林业实验局林业科学研究所。该地地处秦岭西段,土壤为黄绵土,年平均气温10.7℃, ≥ 10℃ 积温3 359℃,极端高温39℃,极端低温−19.2℃,无霜期约190 d,年降水量600 mm以上。
-
分别于2018年(11年生)测定灰楸种质的叶部性状,2020年(13年生)测定灰楸种质的生长和材性性状,每份种质选择生长正常的3个分生株,测定指标包括树高、胸径、冠幅、Pilodyn值、弹性模量、叶长、叶宽、叶长宽比、叶柄长和SPAD值,共10项指标。树高和胸径的测定精确度分别为0.01 m和0.1 cm;测定东西冠幅和南北冠幅,取平均值为单株冠幅(0.01 m)[29];在树高1.3 m处,使用Pilodyn (6J,PROCEQ,Switzerland)分别测定南北向的Pilodyn值[30-31],取平均值作为单株测定值(0.01 mm);使用Director ST300活立木力学性质测定仪进行弹性模量的测定(0.01 GPa)[32-33];每株选择倒数第5~6轮完全展开、无病虫害的3片成熟南向叶,用直尺测量叶长度(0.1 cm)、叶宽度(0.1 cm)和叶柄长度(0.1 cm),计算叶长宽比(0.1),用SPAD-502测定叶绿素含量(0.01)[25]。
-
参照Hu等[28]的逐步聚类方法进行聚类,遗传距离的计算分别采用欧氏距离、切比雪夫距离、绝对值距离、兰氏距离和明科夫斯基距离。
-
参照魏志刚等[14]的方法,分别采用离差平方和法、最短距离法、类平均法、最长距离法、相似法和中间距离法6种系统聚类方法进行系统聚类。
-
根据刘遵春等[27]提出的随机取样法、优先取样法和偏离度取样法进行取样。
-
按照10%、15%、20%、25%和30%的比例进行取样。
-
按照Hu等[28]的方法,计算均值差异百分率(MD)、方差差异百分率(VD)、极差符合率(CR)和变异系数变化率(VR)来评价核心种质的代表性。
-
用Excel 2013整理和分析数据,用SAS 9.4进行巢式设计方差分析,用SPSS 16.0软件进行单因素方差分析,用RStudio进行数据标准化、聚类分析和Shannon Weaver遗传多样性指数(H')的计算。
变异系数(CV) = 标准差(SD)/无性系平均值(
$\overline {{X}} $ )×100%。 -
参试的200个灰楸无性系的生长、叶部和材性性状分析结果(表1)表明:10个性状在无性系间均达极显著差异,灰楸的树高平均值为6.37 m,变幅为3.60~14.90 m;胸径平均值为9.4 cm,变幅为4.3~21.5 cm;冠幅平均值为2.64 m,变幅为1.13~6.05 m。树高、胸径和冠幅的变异系数(CV)分别为18.49%、22.57%和30.39%,遗传多样性指数(H')分别为1.778、1.908和1.915。Pilodyn值和弹性模量的平均值分别为14.01 mm和9.22 GPa,CV分别为12.09%和15.55%,H'分别为1.910和2.003。灰楸的叶部性状中,CV均介于9.66%~18.09%之间,H'均大于1.988。因此,灰楸种质各性状均存在丰富的遗传变异,遗传多样性分布范围较广,具有核心种质构建的必要性和可能性。
表 1 灰楸无性系遗传变异和遗传多样性
Table 1. Genetic variation and genetic diversity of C. fargesii clones
性状
Traits平均值
Mean标准差
Standard deviation变异系数
CV/%变异幅度
Variation rangeF值
F value遗传多样性指数
H'生长性状(13年生)
Growth traits树高 Height/m 6.37 1.18 18.49 3.60~14.90 14.10** 1.778 胸径 Diameter/cm 9.4 2.13 22.57 4.3~21.5 6.48** 1.908 冠幅 Crown width/m 2.64 0.82 30.39 1.13~6.05 2.65** 1.915 木材性状(13年生)
Wood traitsPilodyn值 Pilodyn value/mm 14.01 1.69 12.09 8.92~25.00 8.27** 1.910 弹性模量 MOE/GPa 9.22 1.43 15.55 4.76~15.45 3.92** 2.003 叶片性状(11年生)
Leaf traits叶长 Leaf length/cm 14.1 1.81 12.77 9.6~18.9 3.13** 2.074 叶宽 Leaf width/cm 11.0 1.68 14.00 7.3~16.1 4.02** 2.085 叶长宽比 L/W 1.3 0.13 9.66 1.0~1.9 4.30** 1.988 叶柄长 Petiole length/cm 8.2 1.55 18.09 4.1~14.0 3.24** 2.031 SPAD值 SPAD value 44.66 4.59 10.27 27.41~55.68 4.39** 2.056 注:**代表差异极显著(p< 0.01)
Notes: ** represents p < 0.01, the difference is very significant. -
基于欧氏距离、切比雪夫距离、绝对值距离、兰氏距离和明科夫斯基距离5种遗传距离,采用离差平方和法进行聚类,以30%的抽样比例随机取样,构建出的核心种质遗传差异见表2。仅基于兰氏距离构建出的核心种质方差差异百分数(VD)大于0.00%,然而其均值差异百分率(MD)为30.00%,不符合核心种质(MD)低于20.00%的要求,因此,兰氏距离不适于灰楸核心种质的构建。基于切比雪夫距离和明科夫斯基距离构建出的核心种质MD为20.00%;运用欧式距离和绝对值距离构建出的核心种质MD为0.00%,各性状较原始种质均无显著差异。将欧氏距离和绝对值距离进行比较发现,欧氏距离构建的核心种质的极差符合率(CR)(89.14%)和变异系数变化率(VR)(114.09%)均大于绝对值距离的CR(87.52%)和VR(113.07%)。综合分析表明,基于欧式距离构建出的核心种质在还原原种质均值的前提下,极大的保留了原种质的极差,提高了变异系数,更加适用于构建灰楸的核心种质。
表 2 基于不同遗传距离构建的核心种质与原种质差异百分率
Table 2. Percentage difference between core collection and original collection based on different genetic distance
遗传距离
Genetic distance欧氏距离
Euclidean切比雪夫距离
Maximum绝对值距离
Manhattan兰氏距离
Canberra明科夫斯基距离
Minkowski均值差异百分率 MD/% 0.00 20.00 0.00 30.00 20.00 方差差异百分数 VD/% 0.00 0.00 0.00 10.00 0.00 极差符合率 CR/% 89.14 89.39 87.52 79.01 92.40 变异系数变化率 VR/% 114.09 116.92 113.07 105.95 117.48 -
在采用欧氏距离的前提下,基于离差平方和法、最短距离法、类平均法、最长距离法、相似法和中间距离法6种系统聚类方法,以30%的抽样比例进行随机取样,构建出的核心种质遗传差异见表3。基于离差平方和法、最短距离法、类平均法和最长距离法构建的核心种质MD均为0.00%;相似法和中间距离法的MD为20.00%。基于最短距离法构建出的核心种质CR最大,为93.51%,其次为类平均法(89.53%)和离差平方和法(89.14%)。运用最短距离法构建的核心种质VR最大,为120.35%,其次为类平均法(113.10%)和最长距离法(108.95%)。因此,最短距离法更适用于构建灰楸核心种质。
表 3 基于不同系统聚类方法构建的核心种质与原种质差异百分率
Table 3. Percentage of difference between core collection and original collection based on different clustering methods
系统聚类方法
Clustering methods离差平方和法
Ward最短距离法
Single类平均法
Average最长距离法
Compelet相似法
Mcquitty中间距离法
Median均值差异百分率 MD/% 0.00 0.00 0.00 0.00 20.00 20.00 方差差异百分数 VD/% 0.00 0.00 0.00 0.00 0.00 0.00 极差符合率 CR/% 89.14 93.51 89.53 87.61 85.42 84.61 变异系数变化率 VR/% 106.92 120.35 113.10 108.95 108.49 106.92 -
采用欧氏距离,最短距离法聚类后,通过随机取样法、优先取样法和偏离度取样法3种方法进行取样,取样比例分别为10%、15%、20%、25%、30%,构建的核心种质遗传差异见图1。结果表明:采用随机取样法时(图1A),仅在取样比例为10%时,VD > 0.00%,此时VR达到最大值151.46%,MD为10.00%。因此,随机取样法应选择10%的取样比例。在采用优先取样法时(图1B),5种取样比例下,VD均为0.00%,CR均为100.00%;在抽样比例为15%、25%、30%时,MD为0.00%;抽样比例越大,VR值越小。因此,进行优先取样法时应选择15%的取样比例。采用偏离度取样法时(图1C),在抽样比例为15%时,MD最小,为10.00%;VD最大,为40.00%;此时CR和VR分别为91.95%和136.96%。因此,偏离度取样法构建核心种质时应选择15%的取样比例。综合分析可知,采用偏离度取样法,取样比例为15%时,不仅能够较好的还原原种质的均值和极差,提高变异系数,还显著增加了核心种质的方差。
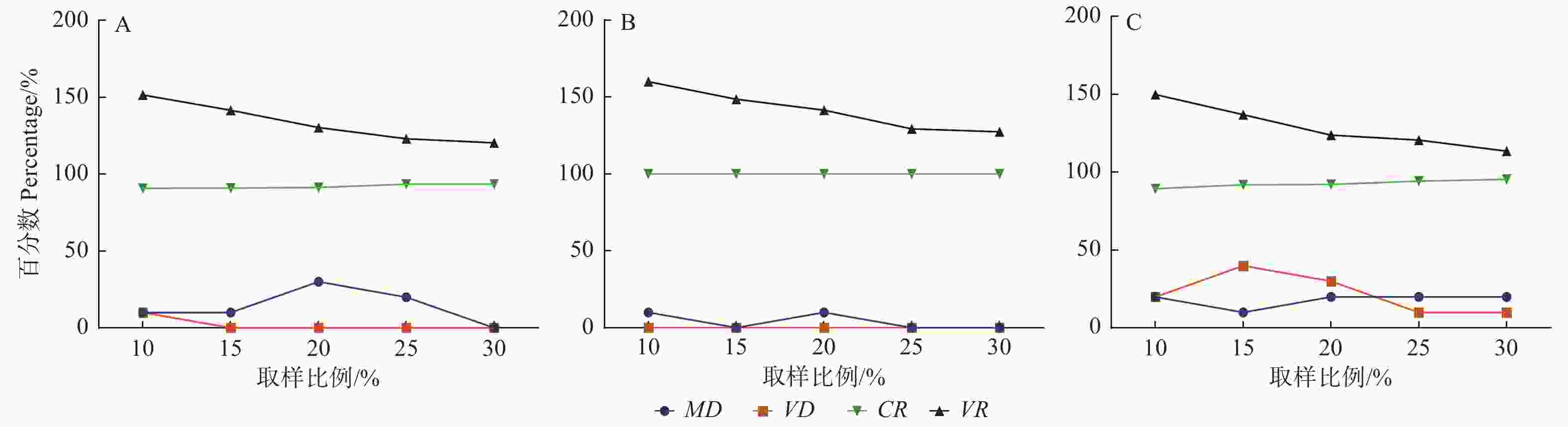
图 1 不同取样比例构建的核心种质与原种质差异百分率
Figure 1. The difference percentage between core collection and original collection constructed by different sampling proportion
-
采用欧式距离和最短距离系统聚类法,以15%的取样比例进行偏离度取样法构建的包括30个无性系的灰楸初级核心种质,MD为10.00%、VD为40.00%、CR为91.95%、VR为136.96%,最能代表原有的种质群体。
将各性状的均值进行分析(表4)表明:各性状核心种质的均值均略大于原种质的均值。将原始种质和核心种质在各省中的分布情况进行分析,结果(图3)显示:原始种质中,甘肃省的种质份数占比最大,为81.50%;陕西省、山西省和河南省的种质份数占比分别为2.00%、11.00%和5.50%。在笔者初步构建的核心种质中,甘肃省的种质份数依旧最大,但低于原始种质的占比,为66.67%;陕西省、山西省和河南省的核心种质份数占比均高于原始种质,分别为3.33%、13.33%和16.67%,表明陕西省、山西省和河南省的种质具有较大的变异程度。
表 4 原始种质和核心种质均值
Table 4. The mean value of original germplasm and core germplasm
性状
Traits原种质均值
Mean value of original germplasm核心种质均值
Mean value of core germplasm生长性状(13年生)
Growth traits树高 Height/m 6.39 6.59 胸径 Diameter/cm 9.4 9.6 冠幅 Crown width/m 2.64 3.05 木材性状(13年生)
Wood traitsPilodyn值 Pilodyn value/mm 14.01 14.19 弹性模量 MOE/GPa 9.22 9.44 叶片性状(11年生)
Leaf traits叶长 Leaf length/cm 14.1 15.0 叶宽 Leaf width/cm 11.0 11.3 叶长宽比 L/W 1.3 1.3 叶柄长 Petiole length/cm 8.2 9.0 SPAD值 SPAD value 44.66 44.83 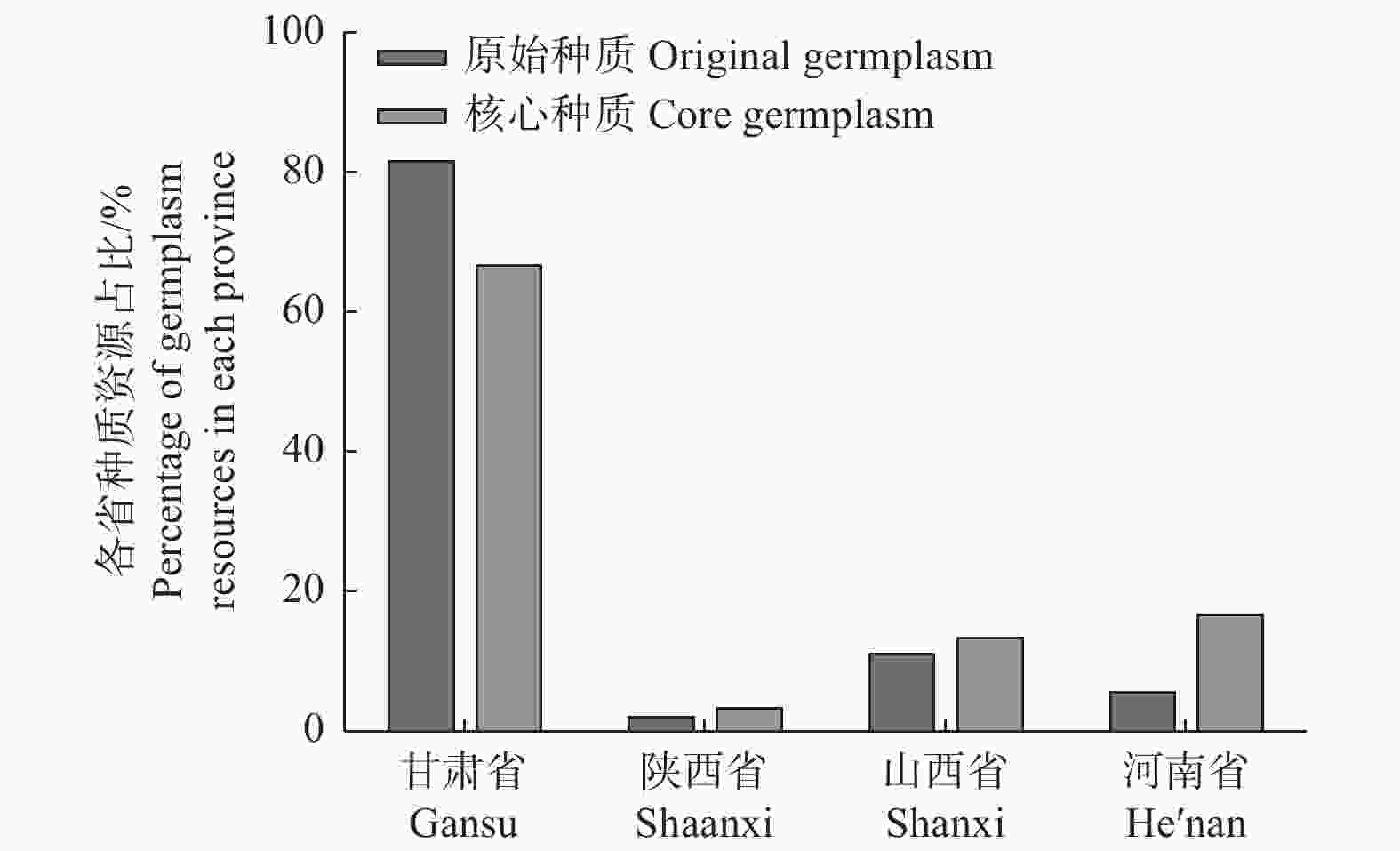
图 3 原始种质和核心种质在4省分布比例
Figure 3. Distribution proportion of original germplasm and core germplasm in four provinces
-
通过聚类法构建核心种质时,首先要计算种质间的遗传距离,再进行聚类分析,然后通过一定的取样比例和取样策略,进行核心种质的抽取[26-27]。遗传距离是度量样品间遗传相似性的综合数量指标,不同遗传距离的计算方法直接影响了聚类结果和核心种质的构建[28]。目前,先通过原始数据标准化以排除各性状量纲的不同对种质材料的影响再采用欧式距离构建核心种质的方法被广泛应用。如刘遵春等将数据标准化后,对不同距离构建的新疆野苹果(Malus sieversii(Ledeb.)Roem.)核心种质进行比较,发现欧式距离要优于马氏距离[27]。在枣树(Ziziphus jujuba Mill.)[34]和棉花(Gossypium hirsutum Linn.)[35]核心种质构建的研究中,同样都是采用欧式距离的效果更好。本研究将原始数据进行标准化后,发现基于欧式距离构建的核心种质具有最小的均值差异百分率,较大的极差符合率和变异系数变化率。因此,欧氏距离更适于灰楸核心种质的构建。然而,基于欧式距离、切比雪夫距离、绝对值距离和明科夫斯基距构建的核心种质方差差异百分数均为0.00%,这可能是由于在比较不同遗传距离时,本研究统一采用离差平方和法进行聚类,以30%的抽样比例进行随机取样,而随机取样法不利于保持原种质的变异程度和多样性[28]。
核心种质受聚类方法的影响[14],通过对不同聚类方法构建的核心种质进行差异分析能达到最优结果。魏志刚等对白桦核心种质进行初步构建,比较了8种不同聚类方法,认为最短距离法构建的核心种质最具代表性[14]。Zhang等发现,离差平方和法最适合芝麻(Sesamum indicum Linn.)核心种质的构建[36]。刘遵春等研究证明,最短距离法是构建新疆野苹果核心种质最好的系统聚类方法[27]。本研究发现,最短距离法的均值差异百分率为0.00%,极差符合率和变异系数变化率均最大,因此,欧氏距离结合最短距离法进行聚类,最适合灰楸核心种质的构建。
取样方法和取样比例是进一步保持核心种质遗传多样性的重要环节[8]。目前常用的取样策略包括随机取样法、优先取样法和偏离度取样法,取样比例一般为原始种质的5%~30%[36]。Hu等研究发现,随机取样法能够保留原始种质的遗传多样性;优先取样法既能获得各性状的极值,又能保持原始种质的遗传变异结构;偏离度取样法选择的种质具有较大的偏离程度,能够保留原始种质的遗传变异丰富度[28]。在对新疆野苹果的核心种质进行构建时,发现优先取样法和偏离度取样法均具有最高的方差差异百分数、极差符合率和变异系数变化率,保留了原种质的遗传变异。但优先取样法具有保留特殊或有价值特性种质的优点,略优于偏离度取样法[27]。这与在棉花[26]、水稻[36](Oryza sativa L.)、白桦[14]和水青树[16]上的研究结果一致。而在对苦瓜[37](Momordica charantia L.)、辣椒[38](Capsicum annuum L.)和樱桃番茄[39](Lycopersivon esculentum Mill.)等植物的研究中发现,采用偏离度取样法构建的核心种质具有较大的遗传差异,更适用于核心种质的构建。在本研究中,以15%取样比例进行偏离度取样法,不仅能较好的还原原种质的均值和极差,还能提高变异系数,不改变原种质的变异水平。因此,采用欧式距离和最短距离法进行聚类,以15%的取样比例通过偏离度取样法构建的包括30个无性系的灰楸初级核心种质最能代表原有的种质群体。
-
本研究基于灰楸无性系11年生的叶部性状和13年生的生长和材性性状,通过欧氏距离、最短距离系统聚类法,以15%的取样比例进行偏离度取样构建的30个核心种质,MD为10.00%、VD为40.00%、CR为91.95%、VR为136.96%,不仅还原了原种质的均值和极差,提高原种质的变异系数,保留变异程度,还略增大了核心种质各性状的均值。在各省的种质分布比例上,核心种质与原种质相差不大,陕西省、山西省和河南省略有增加,表明上述3省的种质变异程度较大,甘肃省较小。这可能是由于在甘肃省取样较为密集,种质间差异较小。另一个原因可能是,4省采集的种质均栽植于甘肃省天水市麦积区,甘肃省的种质更能适应当地的气候环境。而经过10 a以上的生长,来自于陕西省、山西省和河南省的种质受到现栽植地的环境条件影响,发生了适应性变异。
李秀兰曾将267 份1年生灰楸无性系基于生长性状和叶部性状,通过欧氏距离和离差平方和法,进行随机取样,初步构建了63份核心种质[25]。本研究对灰楸核心种质的构建进行了优化。在种质材料方面,以灰楸中龄林为材料,相较于幼龄林,中龄林对立地环境更加适应,具有更强的稳定性[40]。在测量性状方面,灰楸为我国珍贵的用材树种,木材材性是衡量其优良性的一个重要指标,因此,本研究加入Pilodyn值和弹性模量两种材性性状。基于生长、叶片大小和形状以及木材质量3个方面构建的核心种质结果更可靠。在构建方法上面,本研究采用了5种距离计算方法、6种系统聚类方法、5种取样比例和3种取样方法构建灰楸初级核心种质资源,并利用均值差异百分率、方差差异百分率、极差符合率和变异系数变化率对不同方法构建的核心种质进行评价,最终挑选的核心种质更具代表性。所获核心种质中胸径、冠幅、叶长、叶宽、叶长宽比、叶柄长和弹性模量高于原均值的种质份数占比提高,Pilodyn值高于原均值的种质份数占比则降低。因此,本研究构建出的灰楸初级核心种质也在一定程度上提高了核心种质的生长和材性水平。
-
灰楸种质表型性状存在丰富的遗传变异,遗传多样性分布范围较广,运用欧氏距离、最短距离系统聚类法、以15%的取样比例进行偏离度取样构建出30个核心种质,不仅还原了原种质的均值、极差和变异程度,还提高了种质的变异系数,略提高了核心种质的生长和材性水平,为降低种质资源保存成本,促进灰楸种质资源的鉴定和有效利用奠定了基础。
基于表型的灰楸核心种质构建
Construction of Core Collection of Catalpa fargesii Bur. Based on Phenotype
-
摘要:
目的 为得出较为可靠的灰楸初级核心种质群体,以加强灰楸种质选育、开发利用和分子遗传学研究,解决其种质资源保存成本较高问题,促进灰楸种质资源的鉴定和有效利用。 方法 本研究以来自甘肃省、陕西省、山西省和河南省的200个11年生灰楸无性系的叶部性状,13年生的生长性状和材性性状为依据,采用5种距离计算方法、6种系统聚类方法、3种取样方法和5种取样比例构建灰楸初级核心种质资源,并进行评价。 结果 采用欧式距离和最短距离系统聚类法,以15%的取样比例进行偏离度取样构建的包括30个无性系的灰楸初级核心种质,均值差异百分率为10.00%、方差差异百分数为40.00%、极差符合率为91.95%、变异系数变化率为136.96%,最能代表原有的种质群体;核心种质的构建提高了叶长、叶宽、叶长宽比、叶柄长、胸径、冠幅和弹性模量高于均值的种质份数占比,降低了Pilodyn值高于均值的种质份数占比;陕西省、山西省和河南省的核心种质份数占比均高于原始种质。 结论 灰楸种质各性状均存在丰富的遗传变异,30份核心种质不仅还原了原种质的均值、极差和变异程度,还提高了种质的变异系数,略提高了核心种质的生长和材性水平,为促进种质资源的鉴定和有效利用奠定了基础。 Abstract:Objective To obtain a more reliable primary core germplasm group of Catalpa fargesii, to strengthen the breeding, development and utilization and molecular genetics research, to reduce the conservation cost of germplasm resources, and to promote the identification and effective utilization of C. fargesii germplasm resources. Method Based on the leaf traits, the growth traits and the wood traits of 200 C. fargesii clones from Gansu, Shaanxi, Shanxi and He'nan provinces, five genetic distance calculation methods, six clustering methods, three sampling methods and five sampling ratios were used to construct the primary core germplasm resources, and then evaluated them. Result Using the Euclidean distance and the Single system clustering method, the deviation sampling with 15% sampling proportion was adopt to construct the primary core collection of 30 clones, the mean difference percentage was 10%, the variance difference percentage was 40%, the coincidence rate of range was 91.95%, and the changeable rate of coefficient of variation was 136.96%, which could best represent the original germplasm group. The construction of core collection increased the percentage of germplasm with leaf length, leaf width, leaf length width ratio, petiole length, diameter at breast height, crown width and elastic modulus higher than the average, and reduced the percentage of germplasm with Pilodyn value higher than the average. The percentage of core collection in Shaanxi, Shanxi and He'nan provinces were higher than that of the original collection. Conclusion This research showed that there are abundant genetic variation in each trait of C. fargesii clones. The 30 core germplasms not only retain the mean, range and degree of variation of the original germplasms, but also increase the variationcoefficient of the germplasms, and slightly increase the growth and the material quality level of the germplasms. This research laid a foundation for promoting the identification and effective utilization of germplasm resources. -
Key words:
- Catalpa fargesii Bur.
- / phenotypic traits
- / genetic variation
- / genetic diversity
- / core germplasm
-
图 1 不同取样比例构建的核心种质与原种质差异百分率
Figure 1. The difference percentage between core collection and original collection constructed by different sampling proportion
图 3 原始种质和核心种质在4省分布比例
Figure 3. Distribution proportion of original germplasm and core germplasm in four provinces
表 1 灰楸无性系遗传变异和遗传多样性
Table 1. Genetic variation and genetic diversity of C. fargesii clones
性状
Traits平均值
Mean标准差
Standard deviation变异系数
CV/%变异幅度
Variation rangeF值
F value遗传多样性指数
H'生长性状(13年生)
Growth traits树高 Height/m 6.37 1.18 18.49 3.60~14.90 14.10** 1.778 胸径 Diameter/cm 9.4 2.13 22.57 4.3~21.5 6.48** 1.908 冠幅 Crown width/m 2.64 0.82 30.39 1.13~6.05 2.65** 1.915 木材性状(13年生)
Wood traitsPilodyn值 Pilodyn value/mm 14.01 1.69 12.09 8.92~25.00 8.27** 1.910 弹性模量 MOE/GPa 9.22 1.43 15.55 4.76~15.45 3.92** 2.003 叶片性状(11年生)
Leaf traits叶长 Leaf length/cm 14.1 1.81 12.77 9.6~18.9 3.13** 2.074 叶宽 Leaf width/cm 11.0 1.68 14.00 7.3~16.1 4.02** 2.085 叶长宽比 L/W 1.3 0.13 9.66 1.0~1.9 4.30** 1.988 叶柄长 Petiole length/cm 8.2 1.55 18.09 4.1~14.0 3.24** 2.031 SPAD值 SPAD value 44.66 4.59 10.27 27.41~55.68 4.39** 2.056 注:**代表差异极显著(p< 0.01)
Notes: ** represents p < 0.01, the difference is very significant. 下载: 导出CSV
下载: 导出CSV
表 2 基于不同遗传距离构建的核心种质与原种质差异百分率
Table 2. Percentage difference between core collection and original collection based on different genetic distance
遗传距离
Genetic distance欧氏距离
Euclidean切比雪夫距离
Maximum绝对值距离
Manhattan兰氏距离
Canberra明科夫斯基距离
Minkowski均值差异百分率 MD/% 0.00 20.00 0.00 30.00 20.00 方差差异百分数 VD/% 0.00 0.00 0.00 10.00 0.00 极差符合率 CR/% 89.14 89.39 87.52 79.01 92.40 变异系数变化率 VR/% 114.09 116.92 113.07 105.95 117.48
下载: 导出CSV
表 3 基于不同系统聚类方法构建的核心种质与原种质差异百分率
Table 3. Percentage of difference between core collection and original collection based on different clustering methods
系统聚类方法
Clustering methods离差平方和法
Ward最短距离法
Single类平均法
Average最长距离法
Compelet相似法
Mcquitty中间距离法
Median均值差异百分率 MD/% 0.00 0.00 0.00 0.00 20.00 20.00 方差差异百分数 VD/% 0.00 0.00 0.00 0.00 0.00 0.00 极差符合率 CR/% 89.14 93.51 89.53 87.61 85.42 84.61 变异系数变化率 VR/% 106.92 120.35 113.10 108.95 108.49 106.92
下载: 导出CSV
表 4 原始种质和核心种质均值
Table 4. The mean value of original germplasm and core germplasm
性状
Traits原种质均值
Mean value of original germplasm核心种质均值
Mean value of core germplasm生长性状(13年生)
Growth traits树高 Height/m 6.39 6.59 胸径 Diameter/cm 9.4 9.6 冠幅 Crown width/m 2.64 3.05 木材性状(13年生)
Wood traitsPilodyn值 Pilodyn value/mm 14.01 14.19 弹性模量 MOE/GPa 9.22 9.44 叶片性状(11年生)
Leaf traits叶长 Leaf length/cm 14.1 15.0 叶宽 Leaf width/cm 11.0 11.3 叶长宽比 L/W 1.3 1.3 叶柄长 Petiole length/cm 8.2 9.0 SPAD值 SPAD value 44.66 44.83
下载: 导出CSV
-
[1] Brown A H D. Core collections: a practical approach to genetic resources management[J]. Genome, 1989, 31(2): 818-824. doi: 10.1139/g89-144 [2] Frankel O H, Brown A H D. Current plant genetic resources—A critical appraisal[C]//Chopra V L. Genetics: New Frontier. New Delhi: Oxford and IBH Publishing, 1984:1-11. [3] Gepts P. Plant genetic resources conservation and utilization: The accomplishments and future of a societal insurance policy[J]. Crop Science, 2006, 46(5): 2278-2292. doi: 10.2135/cropsci2006.03.0169gas [4] Tana W, Hitomi A, Xu J, et al. Population structure of and conservation strategies for wild Pyrus ussuriensis Maxim. in China[J]. PLoS ONE, 2015, 10(8): e0133686. doi: 10.1371/journal.pone.0133686 [5] Lv J, Li C, Zhou C, et al. Genetic diversity analysis of a breeding population of Eucalyptus cloeziana F. Muell. (Myrtaceae) and extraction of a core germplasm collection using microsatellite markers[J]. Industrial Crops and Products, 2020, 145(3): 112157. [6] 崔艳华, 邱丽娟, 常汝镇, 等. 植物核心种质研究进展[J]. 植物遗传资源学报, 2003,4(3):279-284. [7] 陈学军, 雷 刚, 周坤华, 等. 蔬菜核心种质研究进展[J]. 江西农业大学学报, 2015, 37(1):60-66. doi: 10.3969/j.issn.1000-2286.2015.01.010 [8] 李保印, 张启翔. 我国园艺作物核心种质研究进展[J]. 果树学报, 2007, 24(2):204-209. [9] Hokanson S C, Szewc-Mcfadden A K, Lamboy W F, et al. Microsatellite (SSR) markers reveal genetic identities, genetic diversity and relationships in a Malus domestica Borkh core subset collection[J]. Theoretical & Applied Genetics, 1998, 97(5): 671-683. [10] Hokanson S C, Lamboy W F, Szewc-Mcfadden A K, et al. Microsatellite (SSR) variation in a collection of Malus (apple) species and hybrids[J]. Euphytica, 2001, 118(3): 281-294. doi: 10.1023/A:1017591202215 [11] 邹梁峰. 毛花猕猴桃雄株核心种质构建及遗传多样性分析[D]. 江西农业大学, 2019. [12] 李 娟, 江昌俊. 中国茶树核心种质的初步构建[J]. 安徽农业大学学报, 2004,31(3):282-287. doi: 10.3969/j.issn.1672-352X.2004.03.007 [13] 郭 琪. 刺槐种质资源的遗传多样性评价及核心种质构建[D]. 北京: 北京林业大学, 2019. [14] 魏志刚, 高玉池, 刘桂丰, 等. 白桦核心种质初步构建[J]. 林业科学, 2009, 45(10):74-80. doi: 10.11707/j.1001-7488.20091013 [15] 倪茂磊. 美洲黑杨遗传多样性分析与核心种质库构建[D]. 南京: 南京林业大学, 2011. [16] 张 欢, 王 东, 段 帆, 等. 基于水青树叶表型性状的核心种质资源库构建策略[J]. 林业科学研究, 2019, 32(2):166-173. [17] 中国科学院中国植物志委员会. 中国植物志[M]. 北京: 科学出版社, 1978. [18] 赵秋玲, 马建伟, 王军辉, 等. 灰楸不同流域种质变异与多样性研究[J]. 植物遗传资源学报, 2012, 13(5):803-809. doi: 10.3969/j.issn.1672-1810.2012.05.017 [19] 李平英, 董菊兰, 王军辉, 等. 不同灰楸家系种子生活力试验初报[J]. 甘肃林业科技, 2008,33(2):15-17. doi: 10.3969/j.issn.1006-0960.2008.02.005 [20] 翟晓巧, 聂 琳, 张晓申. 灰楸体外植株再生体系建立[J]. 江西农业学报, 2011, 23(3):17-19. doi: 10.3969/j.issn.1001-8581.2011.03.006 [21] 戴秀芳, 赵秋玲, 冯小芹, 等. 影响灰楸嫁接成活因素研究[J]. 甘肃林业科技, 2013, 38(2):17-19,46. doi: 10.3969/j.issn.1006-0960.2013.02.005 [22] 肖 遥, 麻文俊, 杨桂娟, 等. 6个灰楸无性系叶片表皮微形态特征比较[J]. 电子显微学报, 2017, 36(4):414-421. doi: 10.3969/j.issn.1000-6281.2017.04.015 [23] 于晓池, 杨桂娟, 董菊兰, 等. 梓属5个种对干旱胁迫的生理响应[J]. 植物研究, 2021, 41(1):44-52. doi: 10.7525/j.issn.1673-5102.2021.01.006 [24] 凌娟娟, 肖 遥, 杨桂娟, 等. 灰楸无性系生长和形质性状变异与选择[J]. 林业科学研究, 2019, 32(5):149-156. [25] 李秀兰, 贾继文, 王军辉, 等. 灰楸形态多样性分析及核心种质初步构建[J]. 植物遗传资源学报, 2013, 14(2):243-248. doi: 10.3969/j.issn.1672-1810.2013.02.009 [26] 徐海明, 邱英雄, 胡 晋, 等. 不同遗传距离聚类和抽样方法构建作物核心种质的比较[J]. 作物学报, 2004, 30(9):932-936. doi: 10.3321/j.issn:0496-3490.2004.09.016 [27] 刘遵春, 张春雨, 张艳敏, 等. 利用数量性状构建新疆野苹果核心种质的方法[J]. 中国农业科学, 2010, 43(2):358-370. doi: 10.3864/j.issn.0578-1752.2010.02.017 [28] Hu J, Zhu J, Xu H M. Methods of constructing core collections by stepwise clustering with three sampling strategies based on the genotypic values of crops[J]. Theoretical & Applied Genetics, 2000, 101(1-2): 264-268. [29] 董 晨, 吴保国, 张 瀚. 基于冠幅的杉木人工林胸径和树高参数化预估模型[J]. 北京林业大学学报, 2016, 38(3):55-63. [30] Chen Z Q, Karlsson B, Lundqvist S O, et al. Estimating solid wood properties using pilodyn and acoustic velocity on standing trees of Norway spruce[J]. Annals of Forest Science, 2015, 72(4): 499-508. doi: 10.1007/s13595-015-0458-9 [31] 成俊卿. 主编. 木材学[M]. 北京: 中国林业出版社, 1985. [32] 边黎明, 叶代全, 陈 松, 等. 杉木嫁接植株胸径和木材弹性模量变异分析[J]. 森林与环境学报, 2019, 39(1):104-108. [33] 张帅楠, 姜景民, 徐永勤, 等. 湿地松活立木弹性模量无损评估技术研究[J]. 林业科学研究, 2017, 30(1):75-80. [34] 董玉慧. 枣树农艺性状遗传多样性评价与核心种质构建[D]. 保定: 河北农业大学, 2008. [35] Wang J, Hu J, Huang X, et al. Assessment of different genetic distances in constructing cotton core subset by genotypic values[J]. Journal of Zhejiang University. Science. B, 2008, 9(5): 356-362. doi: 10.1631/jzus.B0710615 [36] Zhang X, Zhao Y, Cheng Y, et al. Toby Hodgkin. Establishment of sesame germplasm core collection in China[J]. Genetic Resources and Crop Evolution, 2000, 47(3): 273-279. doi: 10.1023/A:1008767307675 [37] 刘子记, 牛 玉, 朱 婕, 刘昭华, 杨 衍. 苦瓜核心种质资源构建方法的比较[J]. 华南农业大学学报, 2017, 38(1):31-37. doi: 10.7671/j.issn.1001-411X.2017.01.006 [38] 刘子记, 曹振木, 朱 婕, 等. 甜椒核心种质资源比较构建研究[J]. 东北农业大学学报, 2016, 47(1):21-29. doi: 10.3969/j.issn.1005-9369.2016.01.004 [39] 牛 玉, 刘维侠, 杨 衍, 等. 樱桃番茄核心种质资源构建策略[J]. 热带作物学报, 2019, 40(12):2356-2363. [40] 沈国舫. 森林培育学[M]. 北京: 中国林业出版社, 2001: 21. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4176
- HTML全文浏览量: 2308
- PDF下载量: 69
- 被引次数: 0

