

-
近几十年来,生物多样性与生产力之间的关系是生态学领域的研究热点[1],Huang等[2]的研究证实了增加生物多样性不仅能提高生态系统的生产力,而且可以增加土壤的碳储量。Liang等[3]对森林生物多样性与生产力的关系研究发现,森林生物多样性与生产力关系呈正相关性的下凹式曲线关系,这也意味着持续的生物多样性丧失将加速全球森林生产力的下降。如全球森林树种丰富度每下降10%将会引起当前森林生产力下降2%~3%,而当物种丰富度变为单一时,森林生产力可能会下降26%~66%。原生植物生物多样性的丧失和外来物种入侵正在威胁着许多生态系统维持关键功能和过程的能力[4-7]。生物多样性保护是森林可持续经营的关键目标[8-9],维护和保持森林结构多样性或生境复杂性常被认为是保护生物多样性的最佳途径[10]。生物多样性测度是定量评价生物多样性现状和为其保护实践活动提供理论依据的重要工具[11-12]。寻求有效表达或度量生物多样性的方法与途径已成为生态学研究的挑战[11, 13]。生物多样性是指一个生物群落中的树种数目和各物种的个体数目分配的均匀度[14]。不同学者由于研究的目的和对象不同,对它的理解并不完全相同。有的指的是不同种群内的个体数目[15],有的指的是群落中种的数目[16-18],Poole[18]认为群落内物种的数目或丰富度这个指标才是唯一真正客观的多样性指标。可见,物种丰富度在某种程度上等同于物种多样性,进而就出现了许多直接把调查样地中的树种数作为物种丰富度来进行生物多样性分析的报道[2, 19-21],忽视了调查样地大小的问题[22]。确定一个植物群落的种类组成的古老的办法就是在这个群落地段上进行种类统计,但是由于一个群落地段所占的面积常常很大,种在群落内分布也很少是均匀的,我们既不可能对整个群落地段的物种进行全面统计,也不可能仅用只在一块很小的面积上进行的物种调查用以代表整个群落的种类组成,这就产生了一个研究群落种类组成时统计面积适当大小的问题。由此诞生了最为经典的种-面积曲线法[23-25]。所谓群落最小面积就是能够充分体现群落物种组成的面积,是在一个最小地段内,对一个特定群落类型能提供足够的空间环境,或者能够确保展现出该群落类型的种类组成和结构的真实特征,即在该面积内群落的种类组成和特征能得以充分的表现。该方法通常采用在群落地段的中央,逐步成倍扩大样方面积,统计随着面积扩大增加的种数,用种的数目随样方面积增加的关系,绘制出种-面积曲线。这种曲线的特征是,起初陡峭上升,而后慢慢趋于平缓,曲线开始平缓时所对应的面积即群落的最小面积。因此,调查的样地面积起码要大于等于群落最小面积,理论上样地面积越大(面积
$ \to +\mathrm{\infty } $ )越能真实体现群落的树种组成(期望值),但实践中,不可能直接通过调查手段解决无穷大样地上的树种数,同样,也绝无把握确信所拥有的调查样地都等于群落最小面积。本研究试图给出确定森林群落树种丰富度的方法,旨在完善森林生物多样研究的方法,以提高森林生物多样性研究的科学性。
-
为了验证本研究所提出的方法,收集了不同气候带典型森林群落每木定位大样地数据(表1):寒温带天山云杉天然林(XTS)、温带长白山针阔混交林(DDP)、亚热带黑石顶常绿阔叶林(HSD)和巴拿马热带雨林(BCI)。
表 1 大样地基本概况
Table 1. Large-scale plots profile
样地
Plots森林类型
Forest types样地大小
Plot size/ hm2断面积
Basal area/(m2·hm−2)密度
Density/(trees·hm−2)树种数
Number of species平均胸径
DBH/cmBCI 巴拿马热带雨林
Tropical rainforest in Barro Colorado Island4 30.22 4 004 226 9.8 HSD 亚热带黑石顶常绿阔叶林
Subtropical evergreen Broad-leaf forest in Heishiding4 37.69 3 852 169 11.2 DDP 温带长白山针阔混交林
Temperate coniferous and broad-leaved mixed forest in Changbai Mountain1 24.64 936 18 18.3 XTS 寒温带天山云杉天然林
Temperate natural forest of Picea schrenkiana in Tianshan Mountain4 48.78 200 3 55.8 寒温带天山云杉天然林(XTS)位于新疆维吾尔自治区伊犁地区的西天山国家级自然保护区内(43°03′~43°15′ N,82°51′~83°06′ E),海拔2 000 m,属温带大陆性气候,年平均气温5~7℃,年降水量800~1 000 mm,土壤类型以山地灰褐色森林土壤为主[26]。
温带长白山针阔混交林(DDP)位于吉林省蛟河东大坡自然保护区内(43º51′~44º05′ N,127º35′~127º51′ E),海拔在800 m以下,属温带大陆性季风山地气候,年均气温0.9~3.9℃,年均降水量700~800 mm,土壤为潜育化暗棕壤,主要是以红松(Pinus koraiensis Sieb. et Zucc.)、沙松(Abies holophylla Maxim.)、臭冷杉(Abies nephrolepis Maxim.)和鱼鳞云杉(Picea jezoensis Carr.)等为主要顶级植被的针阔混交林,植被类型属长白山植物区系[27]。
亚热带黑石顶常绿阔叶林(HSD)位于广东省肇庆市黑石顶自然保护区内(23°25′~23°29′ N,111°49′~111°55′ E),海拔多在150~698 m,属南亚热带湿润季风气候,年平均气温10.6~28.4℃,年降水量1 743. 8 mm,降水集中在4— 9月,土壤类型为红壤。植被类型为南亚热带低山常绿阔叶林[28]。
巴拿马热带雨林(BCI)位于巴拿马Barro Colorado Island(9°9.075′~9°9.345′ N,79°51.310′~79°51.318′ W),海拔为121.2~159.2 m,年均降水量2 551 mm,雨季主要集中在5—12月,年日均最高温为31.3℃,年日均最低温为23.2℃。该样地的气候信息详见
http://www.stri.org/tesp 。样地的地质及物种组成等详细信息参见Croat[29]、Leigh等[30]、Hubbell和Foster[31]等文献,植被类型为热带雨林,全部物种名录详见:https://repository.si.edu/handle/10088/20925 。 -
按Gadow and Hui[25]的研究方法,树种数随调查面积的增加而逐渐增加,起初陡峭上升,而后慢慢趋于平缓,这种种-面积关系可采用Monod模型表征[32]:
$ S=\frac{aA}{1+bA} $
(1) 式中,a、b-参数;S-种数;A-面积(m2);
对于带有林木位置坐标的大样地数据而言,可以从大样地中任意随机点开始,画10 m × 10 m的样方,计数该面积下的树种数。依此类推,获得20 m × 20 m、30 m × 30 m、40 m × 40 m……的样方,若所取样方不能完全包含于大样地内,则重新设定起始点进行取样。重复进行若干次(如随机进行100次),把相同样方面积上调查的树种数均值作为该调查面积下的统计值。基于此数据对模型(1)进行非线性拟合,即可获得该群落的种-面积曲线。
建立了群落种-面积曲线,就可以获得该群落的两个重要信息:群落最小面积(能够充分体现群落物种组成的面积)(Amin)和群落最大树种数(即
$ A\to +\mathrm{\infty } $ 时,通过Monod模型计算的树种数)(Smax),这个Smax是该群落树种数的期望值,可视为该群落的树种丰富度(R),即R = Smax。为得到群落最小面积,对模型(1)求二阶导数,于是有:
$ {S}^{\text{′}\text{′}}=\frac{-2ab}{{(1+bA)}^{3}} $
(2) 当
$S''$ 小于等于设定的阈值,即基本接近0(本研究采用$S''$ = ± 1 × 10−6)时,此时树种数增速很小,所对应的面积Amin即为该群落最小面积。为获得群落最大树种数,令模型(1)中
$ A\to +\mathrm{\infty } $ ,即当调查面积趋于无穷大时,则群落中的树种数达最大值,即$ {S}_{max}=a/b $
(3) -
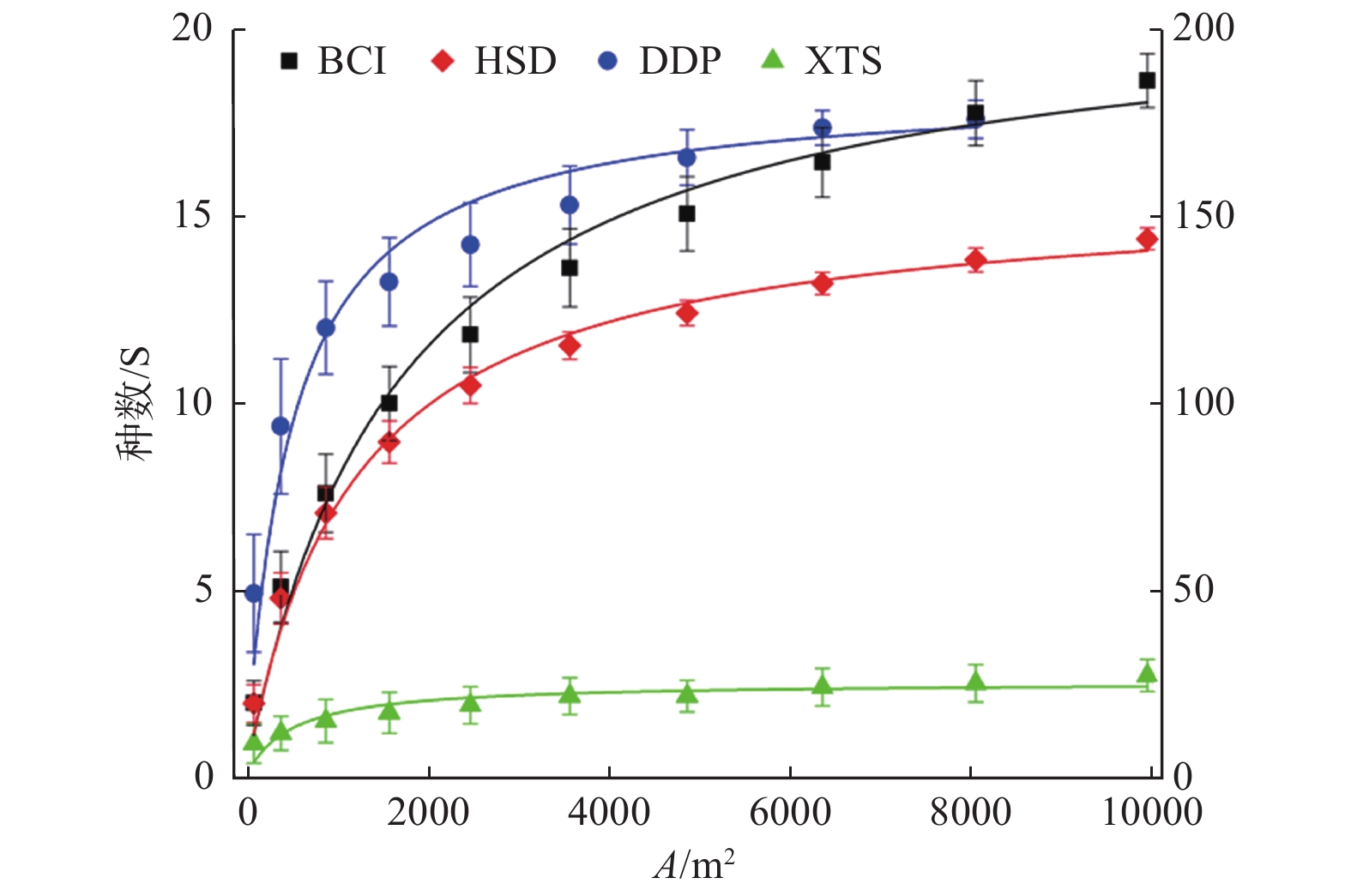
根据群落不同样地面积及所对应的树种数便可得到该群落的种-面积曲线(图1)。由图1可知,物种丰富度随取样面积的增大而增大,该曲线起初陡峭上升,而后慢慢趋于平缓。
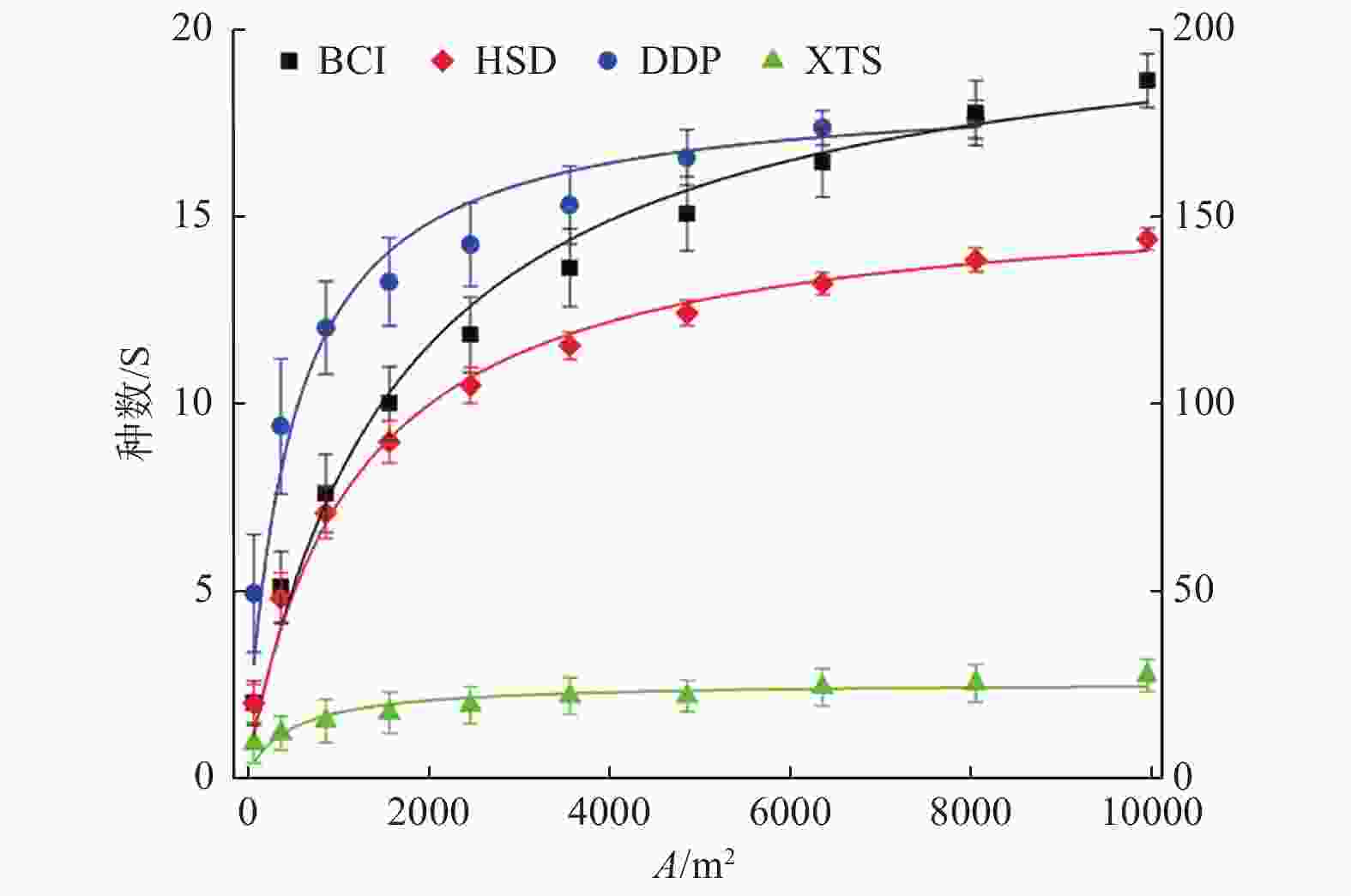
图 1 不同群落种-面积曲线,Y轴表示物种数(S/种数),其中,DDP和XTS用左Y轴,BCI和HSD用右Y轴,公用X轴为样地面积大小(A/m2)。
Figure 1. Species-area curves in different communities. Y axis indicates number of species and X axis indicates area of plot size. DDP and XTS are on the left Y axis, while BCI and HSD are on the right Y axis, with the same scale on the X axis.
采用Monod模型对不同群落的种-面积曲线进行非线性拟合得出相应的模型参数a、b值(表2),采用t检验得出回归参数的显著性均表现为Prob > |t|,说明模型单个变量系数对模型影响显著。
表 2 Monod模型参数统计量
Table 2. Monod model parameter statistics
森林类型
Forest type模型参数
Parameters参数值
Value标准误
Standard errort值
t−valueP值
P−valueBCI a 0.125 5 0.013 4 9.339 8 0.000 0 b 0.000 6 0.000 1 7.150 9 0.000 0 HSD a 0.133 6 0.011 8 11.308 8 0.000 0 b 0.000 9 0.000 1 9.337 3 0.000 0 DDP a 0.036 9 0.006 3 5.888 7 0.000 6 b 0.002 0 0.000 4 5.321 4 0.001 1 XTS a 0.005 5 0.001 5 3.589 9 0.007 1 b 0.002 1 0.000 7 3.151 7 0.013 5 不同群落种-面积曲线用Monod模型拟合的校正判定系数分别为0.987、0.989、0.947和0.825,F检验表明,Monod模型的拟合优度均表现为Prob > F,说明模型拟合效果符合要求。
这个拟合结果再次证实,种-面积关系可采用Monod模型很好描述。同时也表明,研究所采用的处理带有林木位置坐标的大样地数据的方法是可行的,即可以从大样地中任意随机点开始,画10 m × 10 m的样方,计数该面积下的树种数。依此类推,获得20 m × 20 m、30 m × 30 m、40 m × 40 m……的样方,若所取样方不能完全包含于大样地内,则重新设定起始点进行取样。重复进行若干次(如随机进行100次),把相同样方面积上调查的树种数均值作为该调查面积下的统计值。对于大样地经此处理就可获得该群落的种-面积曲线。
-
利用上述研究方法,将模型参数a、b代入公式(2)和(3)可得到不同群落最小调查面积(Amin)和最大树种数,再将参数a、b和Amin代入模型(1)便可求得最小调查面积所对应的树种数(表3)。
表 3 不同气候带森林群落树种丰富度
Table 3. Species richness of communities in different climate zones
森林类型
Forest type纬度(N)
Latitude海拔/m
Altitude气候带
Climatic zone群落最大树种数
Smax群落最小面积/m2
Amin最小调查面积所对应的树种数
SA=AminBCI 9°9′ 140 热带 209.77 8031.86 174 HSD 23°2′ 414 亚热带 156.81 6895.99 134 DDP 43º5′ 800 温带 18.35 2240.83 15 XTS 43°1′ 2000 寒温带 2.60 802.63 2 由表3可见,用最大树种数作为森林群落树种丰富度的测度,可以得出如下结果:
热带BCI最大,近210种;亚热带(HSD)次之,达157种;温带(DDP)仅为几十种,约19种;寒温带(XTS)最低,约为3种。随着纬度变化而变化,符合物种数随气候带由南(热带)到北(寒温带)逐渐减少的普遍规律。群落最小表现面积,热带BCI最大,高达8 000 m2以上,对应树种数为174;亚热带(HSD)次之,近6 900 m2,对应树种数为134;温带(DDP)约2 300 m2,对应树种数为15;寒温带(XTS)最小,约800 m2,对应
树种数为2;符合物种数越多需要调查的森林面积越大的普遍规律。 -
物种丰富度通常被定义为群落中物种数目的多少。这个定义,对于一定孤立系统,如分布范围和地理区域并与其它群落有明显区别的群落而言,非常明确。而对于一个面积非常庞大的森林群落而言,只能通过典型调查或抽样调查获得物种数量。这就存在一个测度群落物种丰富度的关键问题:调查面积大小的问题。一个森林生态系统必然占据一定的面积,在此面积上所有的植被并不是杂乱无章的堆积,而是有一定的组成结构,并与其环境构成统一的整体。样地面积大小是森林调查中的一项重要内容,调查面积的大小对于恰当表征一定森林群落特征具有重要意义,而且它直接影响到调查精度和成本,因此受到众多学者的重视。
群落物种丰富度主要是测定群落内物种数目,用以表达群落中物种的丰富程度。森林群落树种丰富度通常采用一定样地面积中的树种数目表示,这是最简单、最古老的树种多样性测度方法,但测度的关键是合适的样地大小,群落最小面积就是针对这一关键技术问题而被提出。采用不同气候带固定样地数据既可以研究物种多样性的梯度变化特征,也是对所采用方法合理与否的检验。研究结果充分说明,如果用最小体现面积上的树种数表达树种丰富度,虽然其结果不影响不同地区不同类型的森林多样性的相对比较大小,但从绝对量大小来看,其值确实远远低估了树种多样性,而且对于树种丰富地区如热带地区更是如此。当然,对于树种比较少的地区如寒温带影响则相对较少;另一方面,如果对于不同地区不同类型的森林采用任意调查面积上的树种数进行丰富度或多样性比较研究,那将会导致完全不同的结论,譬如,本研究中,如果人们试图比较巴拿马热带雨林(BCI)和我国热带常绿阔叶林(HSD)的树种多样性,均采用面积为400 m2的调查结果,将会得出二者的树种多样性几乎没有差异的错误结论,同样,如果采用不同调查面积上的数据进行比较,也会得出不同的结论。
采用本研究提出的最大树种数来测度森林群落树种丰富度具有科学合理性,因为一个森林群落只有一个种-面积曲线,在面积无穷时,也只能有一个最大值,所以比较树种丰富度或树种多样性才具有可信性,同样,基于此进行树种多样性与生产力的关系研究才具有科学性。这个研究也另一方面阐明了为什么许多在生物多样性与生产力研究结论相矛盾的问题。
-
本研究首次将最大树种数作为测度森林群落树种丰富度,克服了现有研究中直接把不同大小调查面积上的树种数都视为树种丰富度的弊端,从而为森林生物多样性研究提供了共同的比较基础。Monod模型能有效表达森林群落树种数与调查面积之间的非线性关系。基于Monod模型表达种-面积曲线的优点在于,该模型既能确定出森林群落最小面积又能计算出最大树种数。本研究用最大树种数测算出的不同气候带森林群落树种丰富度符合物种丰富度随气候带由南(热带)到北(寒温带)逐渐减少的普遍规律,从而证实了用最大树种数,通过数学手段获得的期望值(在群落面积
$ A\to +\mathrm{\infty } $ 时的树种数),测度森林树种丰富度的可行性。虽然传统森林最小调查面积所对应的树种数也符合不同气候带丰富度变化规律,但还是没有解决不同群落在相同面积上进行树种数的比较问题。
用最大树种数测度森林群落树种丰富度
Measurement of Species Richness of Forest Community by Maximum Tree Species Number
-
摘要:
目的 试图给出确定森林群落树种丰富度的方法,解决物种丰富度测度问题,完善森林生物多样性研究的方法。 方法 采用Monod模型表达种-面积曲线,通过数学手段确定最小森林群落面积和最大树种数。 结果 用本研究提出的方法对不同气候带森林物种丰富度进行测算,结果表明:热带森林群落具有更高的最大树种数和更大的群落最小面积,物种丰富度随气候带由热带到寒温带逐渐减少。 结论 用最大树种数能够确切表达不同气候带森林群落树种丰富度,克服了现有研究中直接把不同大小调查面积上的树种数量视为树种丰富度的弊端。 Abstract:Objective This paper attempts to address the forest species richness measurement by presenting a new measurement method to contribute to the forest biodiversity research. Method Monod model was adopted to express the species-area curve. The minimum forest community area and the maximum number of tree species were determined by mathematical means. Result The method applied in this study have been used to measure forest species richness in different climatic zones and the results further prove the fact that there are higher maximum species number and larger minimum community area in tropical forest communities and confirm the general rule that species richness decreased gradually with climatic zones from tropical to cold temperate zones. Conclusion The maximum number of tree species can accurately express the tree species richness of forest communities in different climatic zones, which overcomes the disadvantages of directly taking the number of tree species in survey areas of different sizes as the tree species richness in current studies. -
图 1 不同群落种-面积曲线,Y轴表示物种数(S/种数),其中,DDP和XTS用左Y轴,BCI和HSD用右Y轴,公用X轴为样地面积大小(A/m2)。
Figure 1. Species-area curves in different communities. Y axis indicates number of species and X axis indicates area of plot size. DDP and XTS are on the left Y axis, while BCI and HSD are on the right Y axis, with the same scale on the X axis.
表 1 大样地基本概况
Table 1. Large-scale plots profile
样地
Plots森林类型
Forest types样地大小
Plot size/ hm2断面积
Basal area/(m2·hm−2)密度
Density/(trees·hm−2)树种数
Number of species平均胸径
DBH/cmBCI 巴拿马热带雨林
Tropical rainforest in Barro Colorado Island4 30.22 4 004 226 9.8 HSD 亚热带黑石顶常绿阔叶林
Subtropical evergreen Broad-leaf forest in Heishiding4 37.69 3 852 169 11.2 DDP 温带长白山针阔混交林
Temperate coniferous and broad-leaved mixed forest in Changbai Mountain1 24.64 936 18 18.3 XTS 寒温带天山云杉天然林
Temperate natural forest of Picea schrenkiana in Tianshan Mountain4 48.78 200 3 55.8  下载: 导出CSV
下载: 导出CSV
表 2 Monod模型参数统计量
Table 2. Monod model parameter statistics
森林类型
Forest type模型参数
Parameters参数值
Value标准误
Standard errort值
t−valueP值
P−valueBCI a 0.125 5 0.013 4 9.339 8 0.000 0 b 0.000 6 0.000 1 7.150 9 0.000 0 HSD a 0.133 6 0.011 8 11.308 8 0.000 0 b 0.000 9 0.000 1 9.337 3 0.000 0 DDP a 0.036 9 0.006 3 5.888 7 0.000 6 b 0.002 0 0.000 4 5.321 4 0.001 1 XTS a 0.005 5 0.001 5 3.589 9 0.007 1 b 0.002 1 0.000 7 3.151 7 0.013 5
下载: 导出CSV
表 3 不同气候带森林群落树种丰富度
Table 3. Species richness of communities in different climate zones
森林类型
Forest type纬度(N)
Latitude海拔/m
Altitude气候带
Climatic zone群落最大树种数
Smax群落最小面积/m2
Amin最小调查面积所对应的树种数
SA=AminBCI 9°9′ 140 热带 209.77 8031.86 174 HSD 23°2′ 414 亚热带 156.81 6895.99 134 DDP 43º5′ 800 温带 18.35 2240.83 15 XTS 43°1′ 2000 寒温带 2.60 802.63 2
下载: 导出CSV
-
[1] 马克平. 当前我国自然保护区管理中存在的问题与对策思考[J]. 生物多样性, 2016, 24(3):249-251. doi: 10.17520/biods.2016082 [2] Huang Y Y, Chen Y X, Castro-Izaguirre N, et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment[J]. Science, 2018, 362: 80-85. doi: 10.1126/science.aat6405 [3] Liang J J, Crowther T W, Picard N, et al. Positive biodiversity-productivity relationship predominant in global forests[J]. Science, 2016, 354: 196-209. [4] Hooper D U, Adair E C, Cardinale B J, et al. A global synthesis reveals biodiversity loss as a major driver of ecosystem change[J]. Nature, 2012, 486: 105-108. doi: 10.1038/nature11118 [5] 盛炜彤. 关于我国人工林长期生产力的保持[J]. 林业科学研究, 2018, 31(1):1-14. [6] Toledo D, Sanderson M, Johnson H, et al. Evaluating plant biodiversity measurements and exotic species detection in national resources inventory sampling protocols using examples from the Northern Great Plains of the USA[J]. Ecological Indicators, 2014, 46: 149-155. doi: 10.1016/j.ecolind.2014.06.020 [7] 惠刚盈, 张弓乔, 赵中华, 等. 林木分布格局多样性测度方法: 以阔叶红松林为例[J]. 生物多样性, 2016, 24(3):280-286. doi: 10.17520/biods.2015237 [8] Hurlbert S H. The nonconcept of species diversity: A critique and alternative parameters[J]. Ecology, 1971, 52: 577-586. doi: 10.2307/1934145 [9] 姜春前, 余树全, 张守攻, 等. 社区森林经营的生态可持续性度量[J]. 林业科学研究, 2005, 18(3):305-309. doi: 10.3321/j.issn:1001-1498.2005.03.015 [10] 刘 斌, 张参参, 汪金松, 等. 江西九连山不同恢复模式林分的物种多样性特征[J]. 林业科学研究, 2020, 33(4):42-52. [11] Purvis A, Hector A. Getting the measure of biodiversity[J]. Nature, 2000, 405: 212-219. doi: 10.1038/35012221 [12] Ricotta C. A recipe for unconventional evenness measures[J]. Acta Biotheoretica, 2004, 52: 95-104. doi: 10.1023/B:ACBI.0000043438.41888.ac [13] 马克平. 生物多样性科学的若干前沿问题[J]. 生物多样性, 2017, 25(4):343-344. doi: 10.17520/biods.2017137 [14] Fisher R A, Steven-Corbet A, Williams C B. The relation between the number of species and the number of individuals in a random sample of an animal population[J]. The Journal of Animal Population, 1943, 12: 42-58. [15] Williams C B. Patterns in the Balance of Nature and Related Problems in Quantitative Ecology[M]. Academic Press, London. 1964. [16] MacArthur R H. Patterns of spices diversity[J]. Biological Reviews, 1965, 40: 510-533. doi: 10.1111/j.1469-185X.1965.tb00815.x [17] Mcintosh R P. An index of diversity and the relation of certain concepts to diversity[J]. Ecology, 1967, 48: 392-404. doi: 10.2307/1932674 [18] Poole R W. Measuring the structural similarity of two communities composed of the same species[J]. Researches on Population Ecology, 1974, 16: 138-151. doi: 10.1007/BF02514086 [19] 孙鹏飞, 沈雅飞, 王丽君, 等. 三峡库区秭归段水位消落带草本植物多样性分析[J]. 林业科学研究, 2020, 33(6):96-104. [20] Gotelli N J, Anderson M J, Arita H T, et al. Patterns and causes of species richness: a general simulation model for macroecology[J]. Ecology Letters, 2009, 12: 873-886. doi: 10.1111/j.1461-0248.2009.01353.x [21] 罗应华, 孙冬婧, 林建勇, 等. 马尾松人工林近自然化改造对植物自然更新及物种多样性的影响[J]. 生态学报, 2013, 33(19):6154−6162. [22] 涂丹丹, 刘蔚漪, 李 媚, 等. 哀牢山西坡6种典型森林群落及物种多样性研究[J]. 西部林业科学, 2020, 49(6):82-90. [23] Barkman J J. Biologisch minimumarealen en de eilandtheorie[J]. Vakblad voor Biologen, 1984, 64: 162-167. [24] Mcguinness K A. Species–area curves[J]. Biological Reviews, 1984, 59(3): 423-440. doi: 10.1111/j.1469-185X.1984.tb00711.x [25] Gadow K V, Hui G Y. Can the tree species-area relationship be derived from prior knowledge of the tree species richness[J]. Forestry Studies, 2007, 46: 13-22. [26] Zang R G, Jing X H, Ding Y, et al. Quantitative classification, ordination and environmental analysis of woody plant communities in Xiaodonggou forest area of the Altai Mountain, Xinjiang[J]. Scientia Silvae Sinicae, 2010, 31: 1055-1057. [27] Li Y, Hui G, Zhao Z, et al. The bivariate distribution characteristics of spatial structure in natural Korean pine broad‐leaved forest[J]. Journal of Vegetation Science, 2012, 23(6): 1180-1190. doi: 10.1111/j.1654-1103.2012.01431.x [28] 昝启杰, 李鸣光, 王伯荪, 等. 黑石顶针阔叶混交林演替过程中群落结构动态[J]. 应用生态学报, 2000, 11(1):1-4. doi: 10.3321/j.issn:1001-9332.2000.01.001 [29] Croat T R. Flora of Barro Colorado Island[M]. Stanford, Stanford University Press, 1978. [30] Leigh E G, Rand S A. Windsor D M. The Ecology of ATropical Forest: Seasonal Rhythms and Long-term Changes[M]. Washington DC, Smithsonian Institution Press, 1982. [31] Hubbell S P, Foster R B. Diversity of canopy trees in a neotropical forest and implications for conservation[M]//Sutton S L, Whitmore T C, Chadwick A C. Tropical Rain Forest: Ecology and Management. Oxford, Blackwell Scientific Publications, 1983. [32] Monod J. La technique de culture continue, the´orie et applications[J]. Annales de l’Institut Pasteur, 1950, 79: 390-410. -

 点击查看大图
点击查看大图
图(1) / 表(3)
计量
- 文章访问数: 4451
- HTML全文浏览量: 2780
- PDF下载量: 67
- 被引次数: 0









