


-
三花槭(Acer triflorum Komarov)属槭树科(Aceraceae)槭属(Acer Linn.)植物,落叶乔木,是我国的珍稀濒危树种[1-4];其主要分布于日本、朝鲜、俄罗斯、蒙古和中国,在中国主要分布于黑龙江、吉林和辽宁地区[5]。三花槭在秋季颜色艳红,引人瞩目,是优良的绿化、美化工程的点缀树种,也是营造彩叶景观特别是廊道彩叶林景观的优选树种[6-7]。因其较高的应用价值,研究者对三花槭的生理生化[8]、播种繁殖[9]、种子内含物[10]、叶色变化和呈色机制等[1,11]进行了研究,并取得了一定的进展。
种子发育是林木生长过程中重要的阶段,由于生长环境迥异、植物长期自然选择和遗传因素的综合影响,使其具有极其丰富的变异,在林木育种中具有重要意义[12-13]。长期以来,在槭属树种研究中,国外对于三花槭的表型变异研究较少,只对三花槭的叶绿体全基因组进行分析[14],利用全基因组超矩阵对槭树周期性迁移、大规模灭绝和迅速的谱系分化及槭树的历史生物地理学等进行研究[15-16]。在国内主要研究三花槭的叶片生理特性、山地移植技术、播种繁殖技术和种实多样性等方面。贾庆斌等[8]对3种叶色型三花槭叶片生理特性比较,不同叶色型三花槭的叶片生理存在显著性差异;在其它树种中,姬志峰等[17]以五角枫为实验材料,探究不同海拔种群的表型多样性,表明五角枫不同种群表型性状存在丰富的遗传变异,为五角枫种质资源的保护、评价和利用提供重要的理论依据;王丹等[18]探究茶条槭不同海拔种群的表型多样性,结果表明5个不同海拔种群表型具有丰富的变异,茶条槭种群内、种群间变异的利用对其遗传改良具有重要的意义;赵阳等[19]探究文冠果种子及苗期生长性状地理种源变异,发现种子性状变异主要受海拔的影响,不同种源间文冠果表型性状均存在极显著差异,且这些差异能够稳定遗传,为文冠果种质基因资源的遗传改良和引种推广提供了遗传背景和实验依据。一系列研究发现,植物繁殖体表型性状的研究将有助于提供丰富的基础数据,也为其资源保护及合理利用提供宝贵资料。
由于槭树是我国的濒危树种,了解其表型变异及生态地理变异规律,为树种的良种选育、资源保护利用和制定保护对策等提供参考依据。梁坤楠等[20]以种源/家系柚木生长、形质性状和适应性为指标,筛选出最适宜生长的优良种源/家系,结果表明,广州龙洞种源、云南芒市种源和2个云南畹町家系是最适宜生长的优良种源/家系,为我国柚木人工林的发展提供优良的种植材料。三花槭作为优良的园林绿化树种之一,虽然用途广泛,但有关其繁殖体表型性状差异及选择研究的报道较少。因此,对其群体种质资源收集、评价和良种选育显得愈发重要[21]。本研究以东北地区不同产地三花槭繁殖体为材料,对其表型与种子活力进行测定,分析表型性状与环境因子的相关性,探讨三花槭繁殖体的表型变异程度与变异规律,最终筛选出优良家系和优良产地,可为三花槭保存、引种和种质资源开发利用提供理论依据。
-
试验材料来自东北三省的4个地点,采样地地理环境、气候因子及各地点采样数量见表1。对种子表型和种子生活力(SV)进行测定,具体测定性状包括翅果长(KFL)、翅果宽(KFW)、翅果长宽比(KFLW)、着生痕长(BM)、果实长(FL)、果实宽(FW)、果实长宽比(FLW)、果实厚度(FT)、翅长(W)、带翅千粒质量(WTKW)、去翅千粒质量(TKW)、空壳率(SR)。
表 1 三花槭4个产地的地理环境因子
Table 1. Geographical environmental factors of four producing areas of A. triflorum
产地
Origin数量
Number家系号
Number经度(E)
Longitude纬度(N)
Latitude海拔
Altitude/m年均气温
Annual temperature/℃年降水量
Annual recipitation/mm林型
Forest stype吉林红石林业局P1 10 1~10 127°06′ 42°67′ 350 3.9 650~750 针阔混交林 黑龙江江山娇林场P2 6 11~16 128°13′ 43°44′ 195 4.3 450~550 针阔混交林 辽宁本溪草河口P3 4 17~20 123°34′ 40°49′ 100 7.5 800~900 针阔混交林 黑龙江五常宝龙店P4 10 21~30 126°33′ 44°04′ 204 3.4 500~800 阔叶林 每个家系随机选择30颗繁殖体进行表型测定,翅果长、翅果宽、着生痕长、果实长、果实宽、果实厚度和翅长(图1)[22]用游标卡尺直接测量,测量精度为0.01 mm;翅果长宽比和果实长宽比直接计算。每株选取400颗带翅繁殖体,利用天平测定,每100颗繁殖体的质量作为百粒质量,对获得数据乘以10作为带翅千粒质量。测量后手动去翅,同样方法测量去翅百粒质量并计算去翅千粒质量;每个单株随机选取400颗繁殖体,用钳子夹碎,计算空壳率;选取200颗繁殖体,分成4份,去壳,温水浸泡24 h,放置30 ℃恒温箱中保存。配置1%的TTC溶液,进行染色,放置恒温箱中避光4~6 h后,放在显微镜下观察胚的染色情况(图2)。
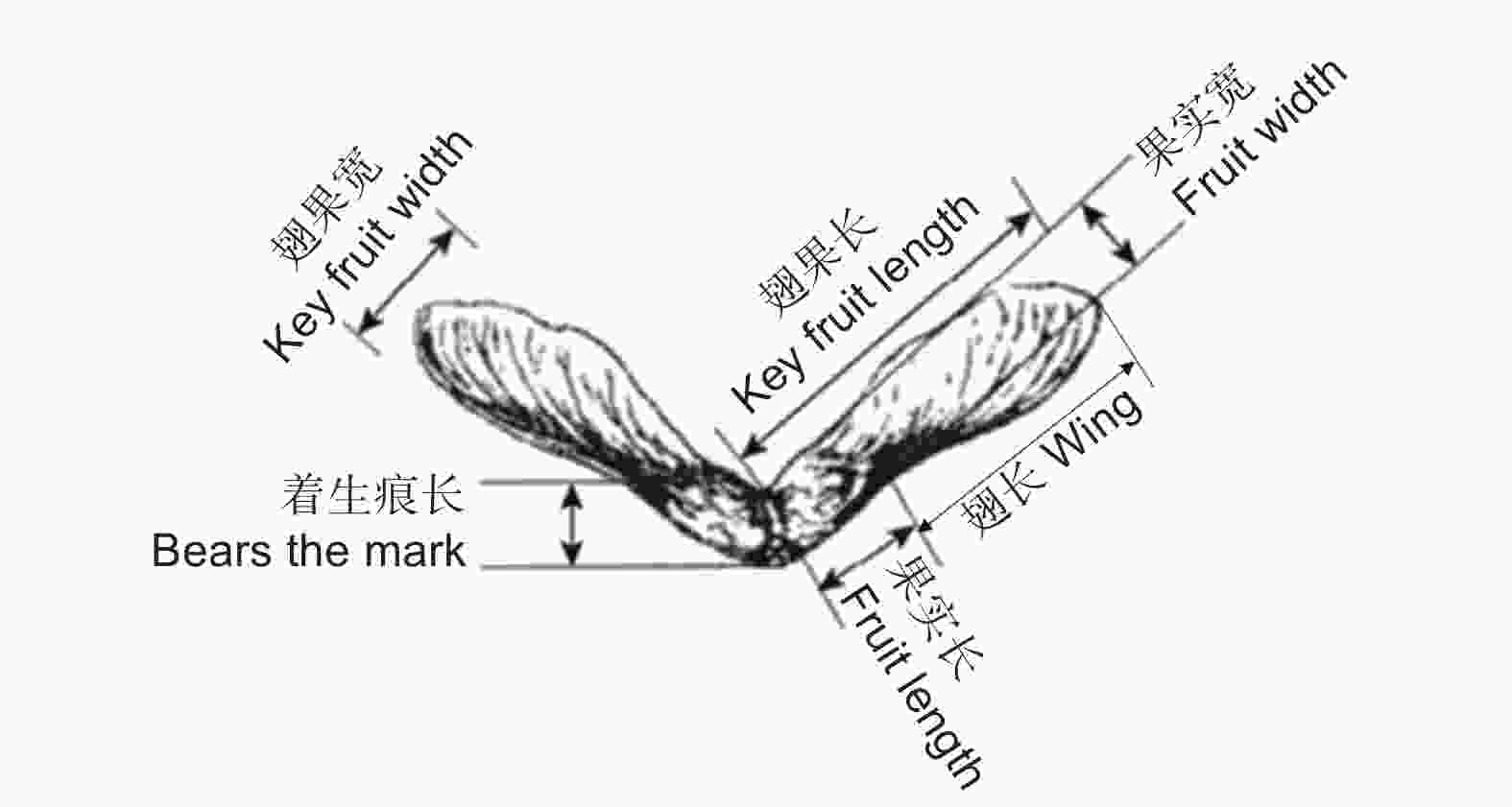
图 1 三花槭果实各指标测定示意图
Figure 1. Diagrammatic drawing of different traits of A. triflorum fruits

图 2 三花槭胚染色与未染色对比图
Figure 2. Contrast picture stained and unstand of A. triflorum embryo
-
所有数据利用SPSS 19.0软件和EXCEL 2010进行计算,空壳率和生活力在方差分析过程中进行反正弦数据转换。
种源间/种源内家系间方差分析线性模型为[23]:
$ X_{ij}=\mu+F_j+e_{ij} $
式中:µ为总体平均值,Fj为种源效应,eij为机误。
表型变异系数(PCV)计算公式[24]:
$ P C V=\frac{S}{\bar{X}} \times 100 {\text{%}} $
式中:S为表型标准差,
$\bar X $ 为某一性状群体平均值。表型相关系数(r)计算公式[25]:
$ r_{A(x y)}=\frac{\sigma_{a(x y)}}{\sqrt{\sigma_{a(x)}^{2} \cdot \sigma_{a(y)}^{2}}} $
式中:σα(xy)为2个性状的表型协方差,σ2a(x)、σ2a(y)分别为2性状的表型方差。
多性状综合评价计算公式[26]:
$ {Q_i}{_ = {\sqrt {\displaystyle{\sum\limits_{j = 1}^n} {{a_i}} } }} $
式中:ai=Xij /Xjmax,Xij为性状的平均值,Xjmax为性状的最优值。
现实遗传增益估算公式[27]:
$ \Delta G=(S / \bar{X}) \times 100 {\text{%}} $
式中:S为选择差,
$\bar X $ 为总体平均值 -
表2表明:P2产地的果实长、果实宽、翅果宽、着生痕长、翅长最大,分别为11.50、10.25、15.70、7.69、33.66 mm;P1产地的果实长、果实宽、翅果长和翅果宽平均值最小,分别为9.85、9.23、38.11、12.82 mm;P3产地的空壳率最小,种子生活力最大,分别为23.56%和87.00%;P4产地的空壳率最大,种子生活力最小,分别为83.00%和16.00%。表明P2产地果实生长状况较好,P3产地种子生活力最好。
表 2 三花槭繁殖体表型性状平均值及标准差
Table 2. Mean average and standard deviation of phenotypic traits of A. triflorum propagules
性状
Trait产地
OriginP1 P2 P3 P4 果实长 FL/mm 9.85 ± 0.60 11.50 ± 0.97 10.20 ± 0.62 10.26 ± 0.66 果实宽 FW/mm 9.23 ± 0.80 10.25 ± 0.81 9.34 ± 0.67 9.72 ± 0.71 果实长宽比 FLW/mm 1.07 ± 0.06 1.13 ± 0.11 1.09 ± 0.06 1.06 ± 0.06 翅果长 KFL/mm 38.11 ± 3.88 43.24 ± 4.78 41.82 ± 2.91 44.75 ± 4.13 翅果宽 KFW/mm 12.82 ± 1.68 15.70 ± 2.12 14.43 ± 1.42 15.00 ± 1.85 翅果长宽比 KFLW/mm 3.00 ± 0.29 2.78 ± 0.33 2.91 ± 0.22 3.01 ± 0.29 果实厚度 FT/mm 6.36 ± 0.58 7.04 ± 0.50 7.07 ± 0.50 6.70 ± 0.45 着生痕长 BM/mm 7.18 ± 0.79 7.69 ± 0.77 7.41 ± 0.66 7.69 ± 0.85 翅长 W/mm 27.78 ± 3.26 33.66 ± 3.78 30.83 ± 2.99 33.25 ± 3.23 带翅千粒质量 WTKW/g 185.93 ± 35.88 220.62 ± 26.78 238.88 ± 9.83 190.48 ± 33.75 去翅千粒质量 TKW/g 161.98 ± 37.79 199.96 ± 27.26 208.04 ± 11.21 165.70 ± 34.91 空壳率 SR/% 36.10 ± 13.49 56.42 ± 18.78 23.56 ± 11.75 83.00 ± 12.00 种子生活力 SV/% 85.00 ± 6.00 73.00 ± 11.00 87.00 ± 4.00 16.00 ± 3.00 -
表3表明:在家系内,不同产地三花槭各性状表型的平均变异系数变幅为6.74%~36.25%,总平均变异系数为12.17%;各表型性状中,空壳率变异系数最大(36.25%),其次是去翅千粒质量,变异系数为15.86%,果实长的变异系数最小(6.74%),种子生活力在P4产地中变异系数最大(20.34%)。各表型性状在不同产地变异程度不同,平均变异系数的变幅为10.44%~13.28%,其中,P1产地的变异较大。
表 3 三花槭繁殖体各表型变异系数
Table 3. Coefficients of phenotypic variation of A. triflorum propagules
% 性状
Trait产地
Origin平均值
MeanP1 P2 P3 P4 果实长 FL 6.07 8.43 6.08 6.39 6.74 果实宽 FW 8.71 7.89 7.13 7.34 7.77 果实长宽比 FLW 6.30 9.36 5.72 5.70 6.77 翅果长 KFL 10.19 11.05 6.96 9.22 9.36 翅果宽 KFW 13.14 13.49 9.83 12.35 12.20 翅果长宽比 KFLW 9.80 11.74 7.71 9.73 9.75 果实厚度 FT 9.19 7.15 7.11 6.66 7.53 着生痕长 BM 10.97 9.97 8.93 11.10 10.24 翅长 W 11.75 11.24 9.69 9.73 10.60 带翅千粒质量 WTKW 19.30 12.14 4.11 17.72 13.32 去翅千粒质量 TKW 23.33 13.63 5.39 21.07 15.86 空壳率 SR 37.37 33.28 52.06 22.30 36.25 种子生活力 SV 7.09 15.20 5.05 20.34 11.92 平均值 Mean 13.28 12.66 10.44 12.28 12.17 -
对4个不同产地三花槭的13个繁殖体表型进行方差分析,结果(表4)表明:除翅果长宽比在不同产地间性状差异不显著(P = 0.787)外,其它性状均差异极显著(P < 0.01),该结果说明不同产地间表型性状存在较大差异。
表 4 三花槭繁殖体表型性状产地间方差分析
Table 4. Variance analysis of phenotypic traits of A. triflorum propagules between origins
性状
Trait变异来源
Variance source自由度
df均方
MSF P 果实长 FL 产地间
Producing areas3 19.297 40.176 0.000 果实宽 FW 产地间
Producing areas3 9.484 16.720 0.000 果实长宽比 FLW 产地间
Producing areas3 0.029 5.714 0.001 翅果长 KFL 产地间
Producing areas3 450.894 26.470 0.000 翅果宽 KFW 产地间
Producing areas3 57.918 17.814 0.000 翅果长宽比 KFLW 产地间
Producing areas3 0.051 0.353 0.787 果实厚度 FT 产地间
Producing areas3 2.493 10.137 0.000 着生痕长 BM 产地间
Producing areas3 2.608 4.420 0.006 翅长 W 产地间
Producing areas3 271.282 25.018 0.000 带翅千粒质量 WTKW 产地间
Producing areas3 11 565.028 11.786 0.000 去翅千粒质量 TKW 产地间
Producing areas3 10 797.737 9.739 0.000 空壳率 SR 产地间
Producing areas3 3.028 109.547 0.000 种子生活力 SV 产地间
Producing areas3 2.940 685.948 0.000 -
表5表明:P1产地的果实长、果实宽、果实厚度、翅长、带翅千粒质量、去翅千粒质量和空壳率在家系间均差异极显著(P < 0.01);P2产地的果实宽、翅果长、翅长、空壳率在家系间均差异显著(P < 0.05);P3产地的翅果长、果实厚度、翅长和空壳率在家系间均差异极显著(P < 0.01);P4产地的果实长、翅长、带翅千粒质量、去翅千粒质量和空壳率在家系间均差异极显著(P < 0.01)。结果说明果实长、果实宽、千粒质量和空壳率等指标在家系间差异是极显著的。
表 5 三花槭繁殖体表型性状家系间方差分析
Table 5. Variance analyis among families of phenotypic traits A. triflorum propagules
性状
Trait产地
Origin变异来源
Variance source自由度
df均方
MSF P 果实长
FLP1 家系间
Families9 0.543 4.156 0.004 P2 5 1.501 1.935 0.162 P3 3 0.466 3.009 0.095 P4 9 1.192 6.584 0.000 果实宽
FWP1 家系间
Families9 1.001 5.120 0.001 P2 5 1.602 4.371 0.017 P3 3 0.510 0.771 0.542 P4 9 1.123 3.444 0.010 果实长宽比
FLWP1 家系间
Families9 0.004 1.448 0.234 P2 9 0.012 0.968 0.475 P3 3 0.002 0.388 0.765 P4 9 0.002 0.593 0.787 翅果长
KFLP1 家系间
Families9 17.935 0.549 0.821 P2 5 16.334 3.901 0.025 P3 3 29.499 8.620 0.007 P4 9 34.198 7.219 0.000 翅果宽
KFWP1 家系间
Families9 3.946 2.897 0.023 P2 5 12.219 2.397 0.100 P3 3 4.164 3.018 0.094 P4 9 4.604 3.110 0.017 翅果长宽比
KFLWP1 家系间
Families9 0.254 1.284 0.304 P2 5 0.360 2.123 0.132 P3 3 0.028 0.572 0.649 P4 9 0.114 2.503 0.042 果实厚度
FTP1 家系间
Families9 0.545 5.327 0.001 P2 5 0.284 1.425 0.284 P3 3 1.073 8.149 0.008 P4 9 0.365 2.583 0.037 着生痕长
BMP1 家系间
Families9 0.977 3.348 0.012 P2 5 0.545 2.001 0.151 P3 3 1.717 2.372 0.146 P4 9 1.292 3.410 0.011 翅长
WP1 家系间
Families9 19.553 5.135 0.001 P2 5 37.740 3.992 0.023 P3 3 27.834 8.747 0.007 P4 9 20.103 4.549 0.002 带翅千粒质量
WTKWP1 家系间
Families9 4 145.949 125.167 0.000 P2 5 1 872.427 13.586 0.000 P3 3 257.472 5.056 0.030 P4 9 3 704.825 85.131 0.000 去翅千粒质量
TKWP1 家系间
Families9 4 510.696 83.982 0.000 P2 5 2 336.348 14.481 0.000 P3 3 214.040 1.921 0.205 P4 9 4 121.409 57.383 0.000 空壳率
SRP1 家系间
Families9 0.038 13.716 0.000 P2 5 0.053 3.475 0.036 P3 3 0.050 20.817 0.000 P4 9 0.127 11.956 0.000 种子生活力
SVP1 家系间
Families9 0.004 1.080 0.418 P2 5 0.037 17.755 0.000 P3 3 0.002 0.927 0.471 P4 9 0.001 1.141 0.381 -
表6表明:果实长与翅果宽呈极显著正相关(r = 0.477),与果实宽、翅果长、着生痕长、翅长和空壳率均呈显著正相关(0.368 ≤ r ≤ 0.421);果实宽与翅果长、翅果宽、果实厚度、着生痕长、翅长和千粒质量均呈极显著正相关(0.512 ≤ r ≤ 0.795);翅果长与翅果宽、果实厚度、着生痕长和翅长均呈极显著正相关(0.545≤ r ≤0.902);翅果宽与果实厚度、着生痕长、翅长、带翅千粒质量和去翅千粒质量均呈极显著正相关(0.523 ≤ r ≤ 0.841);带翅千粒质量与去翅千粒质量呈极显著正相关(r = 0.993);种子生活力与空壳率和翅果长呈极显著负相关(r = −0.897,r = −0.507)。说明三花槭繁殖体大部分性状间存在较大的相关性。
表 6 三花槭繁殖体表型性状间的相关系数
Table 6. Correlation coefficient between phenotypic characters of Acer triflorum propagules
性状
Trait果实长
FL果实宽
FW果实长宽比
FLW翅果长
KFL翅果宽
KFW翅果长宽比
KFLW果实厚度
FT着生痕长
BM翅长
TL带翅千粒质量
WTSW去翅千粒质量
TSW空壳率
SR果实宽
FW0.411* 果实长宽比
FLW−0.310 −0.066 翅果长
KFL0.368* 0.512** −0.124 翅果宽
KFW0.477** 0.605** −0.003 0.808** 翅果长宽比
KFLW−0.277 −0.335 −0.168 −0.004 −0.588** 果实厚度
FT0.333 0.753** 0.133 0.545** 0.710** −0.479** 着生痕长
BM0.371* 0.769** −0.207 0.616** 0.698** −0.344 0.699** 翅长
W0.421* 0.535** 0.062 0.902** 0.841** −0.188 0.674** 0.655** 带翅千粒质量
WTKW0.267 0.764** −0.055 0.354 0.535** −0.447* 0.763** 0.576** 0.351 去翅千粒质量
TKW0.278 0.795** −0.068 0.333 0.523** −0.451* 0.759** 0.573** 0.340 0.993** 空壳率
SR0.376* 0.315 −0.301 0.460* 0.270 0.159 0.090 0.331 0.423* −0.077 −0.053 种子生活力
SV−0.251 −0.175 0.295 −0.507** −0.257 −0.249 −0.077 −0.351 −0.441* 0.193 0.194 −0.897** 注:** 极显著相关水平(P < 0.01);* 显著相关水平(P < 0.05)。下同。
Notes:** means correlation is significant at the 0.01 level;* means correlation is significant at the 0.05 level.The same below. -
表7表明:果实长、果实宽、千粒质量与海拔呈显著负相关(−0.456 ≤ r ≤−0.380);翅果长、翅果宽、果实厚度、翅长与海拔呈极显著负相关(−0.556≤ r ≤−0.529);空壳率与纬度呈极显著正相关(r = 0.764),与年均气温呈极显著负相关(r = −0.549);种子生活力与年均气温呈极显著正相关(r = 0.516),与纬度呈极显著负相关(r = −0.736)。说明果实的长宽随着海拔的升高而减小,相应的千粒质量也减轻;空壳率随着年均气温的升高而减少,相反种子生活力随着年均气温的升高而增强。
表 7 三花槭繁殖体表型性状与地理环境因子间的相关系数
Table 7. Correlation between phenotypic traits of Acer triflorum propagules and geographical environmental factors
性状
Trait经度
Longitude纬度
Latitude海拔
Altitude年均气温
Annual temperature果实长 FL 0.233 0.130 −0.456* 0.141 果实宽 FW 0.164 0.230 −0.396* 0.012 果实长宽比 FLW 0.162 −0.140 −0.195 0.245 翅果长 KFL −0.070 0.370* −0.535** −0.075 翅果宽 KFW 0.008 0.255 −0.556** 0.050 翅果长宽比 KFLW −0.069 0.101 0.230 −0.216 果实厚度 FT −0.050 0.044 −0.556** 0.234 着生痕长 BM 0.093 0.344 −0.322 −0.138 翅长 W 0.072 0.418* −0.529** −0.103 带翅千粒质量 WTSW −0.207 −0.280 −0.408* 0.433* 去翅千粒质量 TSW −0.146 −0.240 −0.380* 0.390* 空壳率 SR 0.167 0.764** −0.242 −0.549** 种子生活力 SV 0.003 −0.736** 0.300 0.516** -
表8表明:主成分I特征值为5.087,贡献率为39.133%,累计贡献率为38.133%,果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长和翅长等性状特征值较高,分别为0.727、0.840、0.699、0.748、0.752、0.747、0.753;主成分Ⅱ特征值为2.544,贡献率为19.571%,累计贡献率为58.704%,带翅千粒质量、去翅千粒质量和种子生活力等性状特征值较高,分别为0.633、0.615、0.838;主成分Ⅲ特征值为1.367,贡献率为10.513%,累计贡献率为69.217%,其中,果实长宽比特征值较高,为0.777;主成分Ⅳ特征值为1.332,贡献率为10.243%,累计贡献率为79.460%,其中,翅果长宽比特征值较高为0.795。包含了供试产地和性状的大部分信息。
表 8 三花槭繁殖体各表型性状主成分分析
Table 8. Principal component analysis of phenotypic traits of Acer triflorum propagules
主要成分因子
Pricipal Component Factor主成分I
component I主成分Ⅱ
component Ⅱ主成分Ⅲ
component Ⅲ主成分Ⅳ
component Ⅳ特征值
Eigenvalue5.087 2.544 1.367 1.332 贡献率
contribution/%39.133 19.571 10.513 10.243 累计贡献率
Cumulative Component/%38.133 58.704 69.217 79.460 果实长 FL 0.727 0.237 0.321 0.287 果实宽 FW 0.840 0.043 −0.243 −0.089 果实长宽比 FLW −0.096 0.287 0.777 0.515 翅果长 KFL 0.699 −0.401 −0.039 0.366 翅果宽 KFW 0.748 −0.094 0.395 −0.352 翅果长宽比 KFLW −0.175 −0.274 −0.506 0.795 果实厚度 FT 0.752 0.227 −0.081 0.072 着生痕长 BM 0.747 0.011 −0.322 −0.199 翅长 W 0.753 −0.290 0.095 0.160 带翅千粒质量 WTSW 0.611 0.633 −0.133 0.064 去翅千粒质量 TSW 0.626 0.615 −0.148 0.061 空壳率 SR 0.423 −0.736 0.145 −0.092 种子生活力 SV −0.358 0.838 −0.089 0.007 -
依据主成分分析和相关性结果表明,以果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长、翅长和生活力为选择指标对4个产地进行综合评价,P1、P2、P3、P4产地的Qi值分别为2.657、2.793、2.744、 2.631。由于P2产地具有较高的综合评价值(2.793),入选为优良产地,该产地果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长、翅长均值分别为11.50、10.25、43.24、15.70、7.04、7.69、33.66 mm,带翅千粒质量、去翅千粒质量均值分别为220.62、199.96 g,种子生活力均值为73.00(表2),各性状的现实增益分别为10.97%、6.63%、3.34%、9.48%、5.03%、2.83%、7.92%、9.52%、13.00%、22.08% 。
-
依据主成分分析和相关性分析结果对30个家系进行综合评价,各家系Qi值见表9。按果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长、翅长和生活力现实增益在2%以上,兼顾其他性状对家系进行评价选择,入选率为20%,家系19、12、6、22、27、28家系入选,入选家系果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长、翅长均值分别为10.56、9.66、44.29、14.48、6.76、7.43、33.81 mm,带翅千粒质量、去翅千粒质量均值分别为200.56、174.35 g,生活力均值为49%。入选家系的现实增益分别为11.43%~52.81%、7.03%~78.65%、4.08%~47.19%、3.36%~67.50%、2.68%~53.80%、1.13%~26.37%。
表 9 不同家系Qi值
Table 9. Qi value of different families
家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue19 2.692 28 2.668 1 2.663 23 2.653 14 2.633 16 2.605 12 2.681 13 2.667 17 2.662 10 2.652 15 2.633 26 2.568 6 2.680 21 2.665 18 2.662 3 2.649 2 2.620 8 2.561 22 2.680 7 2.664 30 2.657 25 2.649 29 2.616 20 2.510 27 2.673 24 2.664 5 2.656 11 2.636 4 2.608 9 2.490 -
植物表型性状不仅受遗传因素影响,还与外部环境相关,因存在生态适应性,致使不同产地存在较大的表型变异,且这种变异可能遗传给下一代[28-29]。本研究中,根据方差分析结果表明,4个产地的三花槭各性状间差异极显著(除翅果长宽比外),这可能是三花槭对不同生长环境所采取的适应性策略,这现象与Gouwakinnou等[30]研究的伯尔硬胡桃 (Sclerocarya birrea Rich)、龙凤等[31]研究的雷公藤 (Triptery giumwilfor-dill Hook)、Munthali等[32]研究的猴面包树 (Adansonia digitata Linn) 和杨志玲等[33]研究的石蒜 (Lycoris radiate Herb) 等变异情况相似。变异系数是衡量群体变异程度大小的指标,变异系数越大其性状变异程度越高[34],本研究三花槭繁殖体各表型变异系数变幅为6.74%~36.25%,平均变异系数为12.17%,说明产地间有丰富的表型变异,可为下一步选择优良产地奠定基础。
林木改良过程中,常要求综合性改良,因此,各性状的相关性也十分重要[35]。本研究的相关性结果表明,种子生活力与翅果长和空壳率呈极显著负相关,这与王宁等[36]对野生元宝枫相关性分析研究结果相似,表明种子生活力与空壳率不是独立遗传的2个性状,在对三花槭优良产地筛选时应考虑在内。由于三花槭来源于4个不同产地,可能会产生与其生长环境相适应的遗传变异,根据繁殖体表型性状与环境因子间的相关性分析发现,不同产地三花槭繁殖体表型性状受海拔影响较大,果实长、果实宽、翅果长、翅果宽、果实厚度、翅长、千粒质量均受产地海拔影响,表现为海拔越高,果实长宽越小,千粒质量越轻。这与林玮等[34]发现任豆种子厚度随纬度的增大而变薄,种子长宽比随海拔的升高而变小的研究结论相似。千粒质量和种子生活力均与产地年均气温显著相关,表现为气温越高,千粒质量越重,种子生活力越好。这与何霞等[12]研究发现苦楝在气温较高、日照丰富地区的种源苗生长快和生物量大的结论相似。其次,种子上的果实大小和空壳率与纬度也具有相关性,这与高张莹等[37]对核桃楸的研究相似,果核形态变异主要受纬度、温度和湿度的影响。因此,在考虑三花槭选取优良产地和引种时,不能只考虑经纬度,还要将海拔和温度等因素考虑在内,为后续联合选择提供依据。
本研究对不同产地的三花槭繁殖体进行初选,利用主成分分析可以把多个测定指标转化为少数指标对待评价植物进行综合评价,选用果实长、果实宽、翅果长、翅果宽、果实厚度、着生痕长、翅长和生活力等性状生长特征值较高的为筛选指标,筛选出P2产地为最优产地,确定的最优家系号为19、12、6、22、27、28。
-
槭树是我国濒危树种之一,因地理环境和人为活动等因素使其表型存在显著变异,了解产地的地理变异规律,可为后续优良三花槭的收集、保存、利用提供思路。本研究发现三花槭4个产地的果实表型性状之间存在丰富的遗传变异,其主要受海拔和年均气温2个环境因子的影响,果实大小和种子生活力对环境的反映最敏感。初选优良产地和家系,为下一步进行三花槭的遗传多样性研究、种质资源保护和良种选育提供材料与理论基础。
不同产地三花槭繁殖体表型变异及选择研究
Variation and Selection of Phenotypic Traits of Acer triforum Propagules from Different Producing Areas
-
摘要:
目的 阐明东北地区不同产地三花槭繁殖体表型性状变异规律,为三花槭种质资源保护和良种选育提供基础。 方法 以黑龙江、辽宁和吉林三省的4个产地的三花槭繁殖体为材料,对其表型和活力等性状进行测定分析。 结果 果实方差分析结果表明:除翅果长宽比(P = 0.787)外,其它性状在产地间均差异极显著(P < 0.01),不同产地间三花槭表型性状存在显著差异。各性状间相关性分析结果表明:果实长与果实宽、翅果长、着生痕长、翅长和空壳率呈显著正相关(0.368≤r≤0.421),种子生活力与空壳率和翅果长呈极显著负相关(r = −0.897,r = −0.507)。三花槭繁殖体与环境因子的相关性分析表明:翅果长、翅果宽、果实厚度和翅长均与海拔呈极显著负相关(−0.556≤r≤−0.529),果实长、果实宽和千粒质量与海拔呈显著负相关(−0.456≤r≤−0.380),空壳率与年均气温呈极显著负相关(r = −0.549),种子生活力与年均气温呈极显著正相关(r = 0.516)。利用多性状综合评价法对产地和家系进行综合评价,P2产地综合评价值较高,果实长等现实增益为2.83%~22.08%。以20%的入选率,初选出6个优良家系,果实长等现实增益为1.13%~78.65%。 结论 三花槭繁殖体表型性状存在丰富的差异,主要受海拔和年均气温2个环境因子的影响。根据多性状综合评价初步选定P2产地和家系19、12、6、22、27、28为优良材料。 Abstract:Objective To elucidate the variation pattern of the propagules phenotypic characters of Acer triflorum from different origins in Northeast China, and provide the basis for genetic resource conservation and selection breeding of Acer triflorum. Method Acer triflorum propagules from four producing areas of Heilongjiang, Liaoning and Jilin were used to determine the phenotype and vigor of the propagules. Result The results of variance analysis showed that, except the ratio of length to width of key fruit (P = 0.787), the differences of other traits among origins were extremely significant (P < 0.01). There were significant differences in phenotypic characters of Acer triflorum among different producing areas. Correlation analysis of each character showed that fruit length was positively correlated with fruit width, samara length, bears mark, wing length and shell percentage (P < 0.05, 0.368≤r≤0.421), and seed viability wasnegativly correlated withsamara length and shell rate (r = −0.897, r = −0.507, P < 0.01). Correlation analysis between the propagules of Acer triflorum and environmental factors showed that the samara length, samara width, fruit thickness and wing length were negatively correlated with altitude (−0.556≤r≤−0.529, P < 0.05). Fruit length, fruit width and 1000-seed weight were negatively correlated with altitude (−0.456≤r≤−0.380, P < 0.05). Shell rate was negatively correlated with annual mean temperature (r = −0.549, P < 0.05), whereas seed viability was positively correlated with annual mean temperature (r = 0.516, P < 0.05). The comprehensive evaluation value of P2 origin was higher than the others, with realistic gain 2.83%-22.08%. The realistic gain of 6 excellent families was 1.13%-78.65% based on the 20% selection rate. Conclusion There are abundant differences in phenotypic traits of Acer trifolium, which is mainly affected by two environmental factors: altitude and average annual temperature. According to the comprehensive evaluation of multiple characters, preliminarily selected P2 origin and 6 families including 19, 12, 6, 22, 27, 28 are excellent materials. -
Key words:
- Acer triforum
- / fruit character
- / genetic variation
- / family selection
- / producing areas selection
- / realistic gain
-
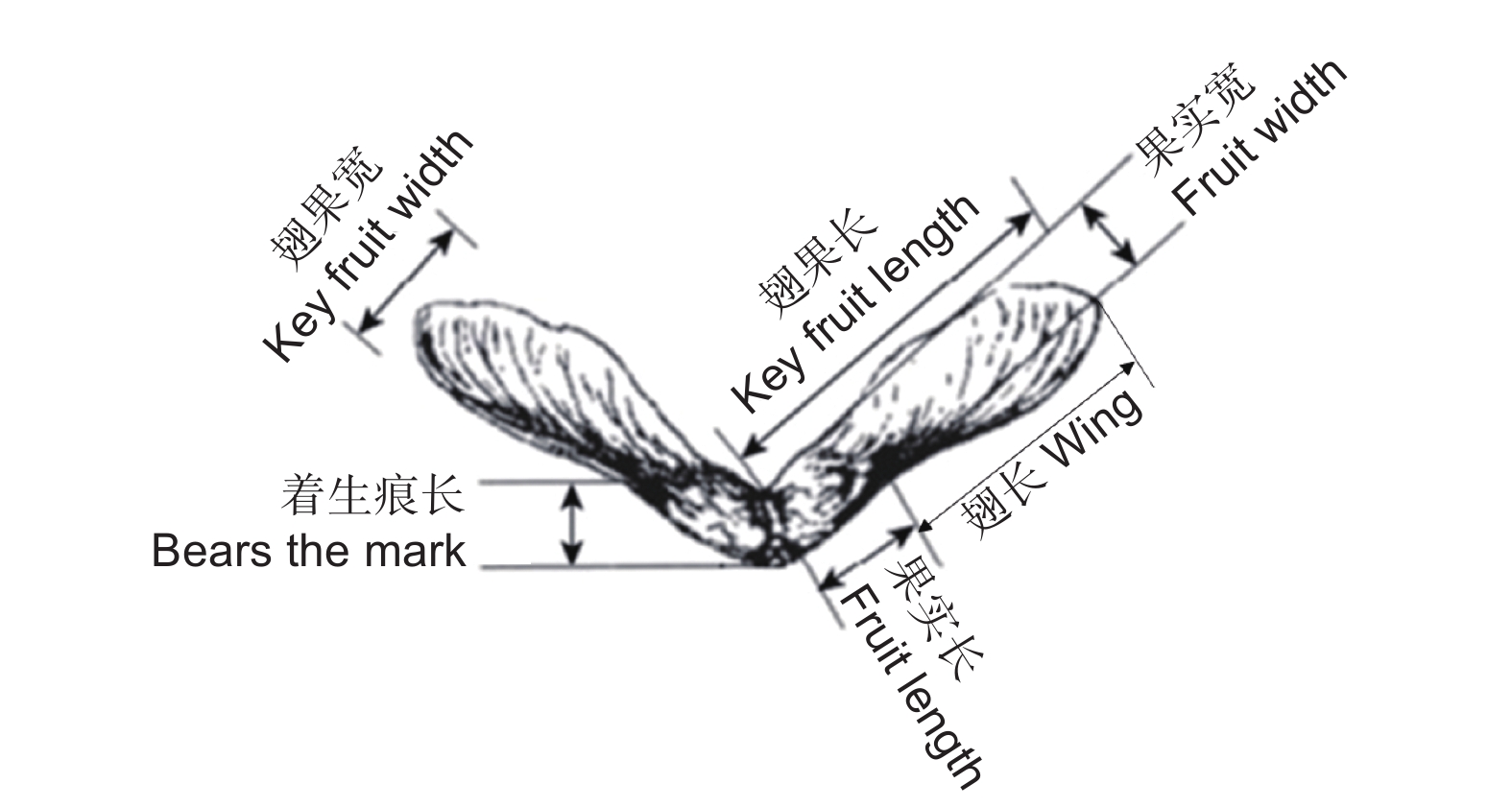
图 1 三花槭果实各指标测定示意图
Figure 1. Diagrammatic drawing of different traits of A. triflorum fruits
图 2 三花槭胚染色与未染色对比图
Figure 2. Contrast picture stained and unstand of A. triflorum embryo
表 1 三花槭4个产地的地理环境因子
Table 1. Geographical environmental factors of four producing areas of A. triflorum
产地
Origin数量
Number家系号
Number经度(E)
Longitude纬度(N)
Latitude海拔
Altitude/m年均气温
Annual temperature/℃年降水量
Annual recipitation/mm林型
Forest stype吉林红石林业局P1 10 1~10 127°06′ 42°67′ 350 3.9 650~750 针阔混交林 黑龙江江山娇林场P2 6 11~16 128°13′ 43°44′ 195 4.3 450~550 针阔混交林 辽宁本溪草河口P3 4 17~20 123°34′ 40°49′ 100 7.5 800~900 针阔混交林 黑龙江五常宝龙店P4 10 21~30 126°33′ 44°04′ 204 3.4 500~800 阔叶林  下载: 导出CSV
下载: 导出CSV
表 2 三花槭繁殖体表型性状平均值及标准差
Table 2. Mean average and standard deviation of phenotypic traits of A. triflorum propagules
性状
Trait产地
OriginP1 P2 P3 P4 果实长 FL/mm 9.85 ± 0.60 11.50 ± 0.97 10.20 ± 0.62 10.26 ± 0.66 果实宽 FW/mm 9.23 ± 0.80 10.25 ± 0.81 9.34 ± 0.67 9.72 ± 0.71 果实长宽比 FLW/mm 1.07 ± 0.06 1.13 ± 0.11 1.09 ± 0.06 1.06 ± 0.06 翅果长 KFL/mm 38.11 ± 3.88 43.24 ± 4.78 41.82 ± 2.91 44.75 ± 4.13 翅果宽 KFW/mm 12.82 ± 1.68 15.70 ± 2.12 14.43 ± 1.42 15.00 ± 1.85 翅果长宽比 KFLW/mm 3.00 ± 0.29 2.78 ± 0.33 2.91 ± 0.22 3.01 ± 0.29 果实厚度 FT/mm 6.36 ± 0.58 7.04 ± 0.50 7.07 ± 0.50 6.70 ± 0.45 着生痕长 BM/mm 7.18 ± 0.79 7.69 ± 0.77 7.41 ± 0.66 7.69 ± 0.85 翅长 W/mm 27.78 ± 3.26 33.66 ± 3.78 30.83 ± 2.99 33.25 ± 3.23 带翅千粒质量 WTKW/g 185.93 ± 35.88 220.62 ± 26.78 238.88 ± 9.83 190.48 ± 33.75 去翅千粒质量 TKW/g 161.98 ± 37.79 199.96 ± 27.26 208.04 ± 11.21 165.70 ± 34.91 空壳率 SR/% 36.10 ± 13.49 56.42 ± 18.78 23.56 ± 11.75 83.00 ± 12.00 种子生活力 SV/% 85.00 ± 6.00 73.00 ± 11.00 87.00 ± 4.00 16.00 ± 3.00
下载: 导出CSV
表 3 三花槭繁殖体各表型变异系数
Table 3. Coefficients of phenotypic variation of A. triflorum propagules
% 性状
Trait产地
Origin平均值
MeanP1 P2 P3 P4 果实长 FL 6.07 8.43 6.08 6.39 6.74 果实宽 FW 8.71 7.89 7.13 7.34 7.77 果实长宽比 FLW 6.30 9.36 5.72 5.70 6.77 翅果长 KFL 10.19 11.05 6.96 9.22 9.36 翅果宽 KFW 13.14 13.49 9.83 12.35 12.20 翅果长宽比 KFLW 9.80 11.74 7.71 9.73 9.75 果实厚度 FT 9.19 7.15 7.11 6.66 7.53 着生痕长 BM 10.97 9.97 8.93 11.10 10.24 翅长 W 11.75 11.24 9.69 9.73 10.60 带翅千粒质量 WTKW 19.30 12.14 4.11 17.72 13.32 去翅千粒质量 TKW 23.33 13.63 5.39 21.07 15.86 空壳率 SR 37.37 33.28 52.06 22.30 36.25 种子生活力 SV 7.09 15.20 5.05 20.34 11.92 平均值 Mean 13.28 12.66 10.44 12.28 12.17
下载: 导出CSV
表 4 三花槭繁殖体表型性状产地间方差分析
Table 4. Variance analysis of phenotypic traits of A. triflorum propagules between origins
性状
Trait变异来源
Variance source自由度
df均方
MSF P 果实长 FL 产地间
Producing areas3 19.297 40.176 0.000 果实宽 FW 产地间
Producing areas3 9.484 16.720 0.000 果实长宽比 FLW 产地间
Producing areas3 0.029 5.714 0.001 翅果长 KFL 产地间
Producing areas3 450.894 26.470 0.000 翅果宽 KFW 产地间
Producing areas3 57.918 17.814 0.000 翅果长宽比 KFLW 产地间
Producing areas3 0.051 0.353 0.787 果实厚度 FT 产地间
Producing areas3 2.493 10.137 0.000 着生痕长 BM 产地间
Producing areas3 2.608 4.420 0.006 翅长 W 产地间
Producing areas3 271.282 25.018 0.000 带翅千粒质量 WTKW 产地间
Producing areas3 11 565.028 11.786 0.000 去翅千粒质量 TKW 产地间
Producing areas3 10 797.737 9.739 0.000 空壳率 SR 产地间
Producing areas3 3.028 109.547 0.000 种子生活力 SV 产地间
Producing areas3 2.940 685.948 0.000
下载: 导出CSV
表 5 三花槭繁殖体表型性状家系间方差分析
Table 5. Variance analyis among families of phenotypic traits A. triflorum propagules
性状
Trait产地
Origin变异来源
Variance source自由度
df均方
MSF P 果实长
FLP1 家系间
Families9 0.543 4.156 0.004 P2 5 1.501 1.935 0.162 P3 3 0.466 3.009 0.095 P4 9 1.192 6.584 0.000 果实宽
FWP1 家系间
Families9 1.001 5.120 0.001 P2 5 1.602 4.371 0.017 P3 3 0.510 0.771 0.542 P4 9 1.123 3.444 0.010 果实长宽比
FLWP1 家系间
Families9 0.004 1.448 0.234 P2 9 0.012 0.968 0.475 P3 3 0.002 0.388 0.765 P4 9 0.002 0.593 0.787 翅果长
KFLP1 家系间
Families9 17.935 0.549 0.821 P2 5 16.334 3.901 0.025 P3 3 29.499 8.620 0.007 P4 9 34.198 7.219 0.000 翅果宽
KFWP1 家系间
Families9 3.946 2.897 0.023 P2 5 12.219 2.397 0.100 P3 3 4.164 3.018 0.094 P4 9 4.604 3.110 0.017 翅果长宽比
KFLWP1 家系间
Families9 0.254 1.284 0.304 P2 5 0.360 2.123 0.132 P3 3 0.028 0.572 0.649 P4 9 0.114 2.503 0.042 果实厚度
FTP1 家系间
Families9 0.545 5.327 0.001 P2 5 0.284 1.425 0.284 P3 3 1.073 8.149 0.008 P4 9 0.365 2.583 0.037 着生痕长
BMP1 家系间
Families9 0.977 3.348 0.012 P2 5 0.545 2.001 0.151 P3 3 1.717 2.372 0.146 P4 9 1.292 3.410 0.011 翅长
WP1 家系间
Families9 19.553 5.135 0.001 P2 5 37.740 3.992 0.023 P3 3 27.834 8.747 0.007 P4 9 20.103 4.549 0.002 带翅千粒质量
WTKWP1 家系间
Families9 4 145.949 125.167 0.000 P2 5 1 872.427 13.586 0.000 P3 3 257.472 5.056 0.030 P4 9 3 704.825 85.131 0.000 去翅千粒质量
TKWP1 家系间
Families9 4 510.696 83.982 0.000 P2 5 2 336.348 14.481 0.000 P3 3 214.040 1.921 0.205 P4 9 4 121.409 57.383 0.000 空壳率
SRP1 家系间
Families9 0.038 13.716 0.000 P2 5 0.053 3.475 0.036 P3 3 0.050 20.817 0.000 P4 9 0.127 11.956 0.000 种子生活力
SVP1 家系间
Families9 0.004 1.080 0.418 P2 5 0.037 17.755 0.000 P3 3 0.002 0.927 0.471 P4 9 0.001 1.141 0.381
下载: 导出CSV
表 6 三花槭繁殖体表型性状间的相关系数
Table 6. Correlation coefficient between phenotypic characters of Acer triflorum propagules
性状
Trait果实长
FL果实宽
FW果实长宽比
FLW翅果长
KFL翅果宽
KFW翅果长宽比
KFLW果实厚度
FT着生痕长
BM翅长
TL带翅千粒质量
WTSW去翅千粒质量
TSW空壳率
SR果实宽
FW0.411* 果实长宽比
FLW−0.310 −0.066 翅果长
KFL0.368* 0.512** −0.124 翅果宽
KFW0.477** 0.605** −0.003 0.808** 翅果长宽比
KFLW−0.277 −0.335 −0.168 −0.004 −0.588** 果实厚度
FT0.333 0.753** 0.133 0.545** 0.710** −0.479** 着生痕长
BM0.371* 0.769** −0.207 0.616** 0.698** −0.344 0.699** 翅长
W0.421* 0.535** 0.062 0.902** 0.841** −0.188 0.674** 0.655** 带翅千粒质量
WTKW0.267 0.764** −0.055 0.354 0.535** −0.447* 0.763** 0.576** 0.351 去翅千粒质量
TKW0.278 0.795** −0.068 0.333 0.523** −0.451* 0.759** 0.573** 0.340 0.993** 空壳率
SR0.376* 0.315 −0.301 0.460* 0.270 0.159 0.090 0.331 0.423* −0.077 −0.053 种子生活力
SV−0.251 −0.175 0.295 −0.507** −0.257 −0.249 −0.077 −0.351 −0.441* 0.193 0.194 −0.897** 注:** 极显著相关水平(P < 0.01);* 显著相关水平(P < 0.05)。下同。
Notes:** means correlation is significant at the 0.01 level;* means correlation is significant at the 0.05 level.The same below.
下载: 导出CSV
表 7 三花槭繁殖体表型性状与地理环境因子间的相关系数
Table 7. Correlation between phenotypic traits of Acer triflorum propagules and geographical environmental factors
性状
Trait经度
Longitude纬度
Latitude海拔
Altitude年均气温
Annual temperature果实长 FL 0.233 0.130 −0.456* 0.141 果实宽 FW 0.164 0.230 −0.396* 0.012 果实长宽比 FLW 0.162 −0.140 −0.195 0.245 翅果长 KFL −0.070 0.370* −0.535** −0.075 翅果宽 KFW 0.008 0.255 −0.556** 0.050 翅果长宽比 KFLW −0.069 0.101 0.230 −0.216 果实厚度 FT −0.050 0.044 −0.556** 0.234 着生痕长 BM 0.093 0.344 −0.322 −0.138 翅长 W 0.072 0.418* −0.529** −0.103 带翅千粒质量 WTSW −0.207 −0.280 −0.408* 0.433* 去翅千粒质量 TSW −0.146 −0.240 −0.380* 0.390* 空壳率 SR 0.167 0.764** −0.242 −0.549** 种子生活力 SV 0.003 −0.736** 0.300 0.516**
下载: 导出CSV
表 8 三花槭繁殖体各表型性状主成分分析
Table 8. Principal component analysis of phenotypic traits of Acer triflorum propagules
主要成分因子
Pricipal Component Factor主成分I
component I主成分Ⅱ
component Ⅱ主成分Ⅲ
component Ⅲ主成分Ⅳ
component Ⅳ特征值
Eigenvalue5.087 2.544 1.367 1.332 贡献率
contribution/%39.133 19.571 10.513 10.243 累计贡献率
Cumulative Component/%38.133 58.704 69.217 79.460 果实长 FL 0.727 0.237 0.321 0.287 果实宽 FW 0.840 0.043 −0.243 −0.089 果实长宽比 FLW −0.096 0.287 0.777 0.515 翅果长 KFL 0.699 −0.401 −0.039 0.366 翅果宽 KFW 0.748 −0.094 0.395 −0.352 翅果长宽比 KFLW −0.175 −0.274 −0.506 0.795 果实厚度 FT 0.752 0.227 −0.081 0.072 着生痕长 BM 0.747 0.011 −0.322 −0.199 翅长 W 0.753 −0.290 0.095 0.160 带翅千粒质量 WTSW 0.611 0.633 −0.133 0.064 去翅千粒质量 TSW 0.626 0.615 −0.148 0.061 空壳率 SR 0.423 −0.736 0.145 −0.092 种子生活力 SV −0.358 0.838 −0.089 0.007
下载: 导出CSV
表 9 不同家系Qi值
Table 9. Qi value of different families
家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue家系
FamiliesQi值
Qivalue19 2.692 28 2.668 1 2.663 23 2.653 14 2.633 16 2.605 12 2.681 13 2.667 17 2.662 10 2.652 15 2.633 26 2.568 6 2.680 21 2.665 18 2.662 3 2.649 2 2.620 8 2.561 22 2.680 7 2.664 30 2.657 25 2.649 29 2.616 20 2.510 27 2.673 24 2.664 5 2.656 11 2.636 4 2.608 9 2.490
下载: 导出CSV
-
[1] 庞秋颖, 卓丽环. 槭树树冠秋季变色过程中叶片生理指标的空间差异[J]. 东北林业大学学报, 2007, 35(6):16-17,20. doi: 10.3969/j.issn.1000-5382.2007.06.006 [2] 张振全. 色木槭和拧筋槭种子发育过程中物质变化与脱水耐性的关系[D]. 哈尔滨: 东北林业大学, 2008. [3] 郭 冰, 姜卫兵, 翁忙玲. 论槭属树种及其在园林绿化中的应用[J]. 江西农业学报, 2010, 22(9):49-52. doi: 10.3969/j.issn.1001-8581.2010.09.014 [4] 国 政, 臧润国. 中国极小种群野生植物濒危程度评价指标体系[J]. 林业科学, 2013, 49(6):10-17. doi: 10.11707/j.1001-7488.20130602 [5] 徐廷志. 槭树科的地理分布[J]. 云南植物研究, 1996, 18(1):43-50. [6] 陈继卫, 沈朝栋, 贾玉芳, 等. 鸡爪槭转色期叶色变化生理研究[J]. 核农学报, 2010, 24(1):171-175. [7] 吴培培, 孙 冰, 罗水兴, 等. 岭南槭不同种源家系种子形态和幼苗生长变异[J]. 林业科学研究, 2017, 30(6):1015-1021. [8] 贾庆彬, 孙文生, 刘 庚, 等. 3种叶色型拧筋槭叶片生理特性比较[J]. 北华大学学报:自然科学版, 2020, 21(2):157-162. [9] 史 晔, 刘文华. 彩叶树种三花槭的苗木培育与应用[J]. 特种经济动植物, 2017, 20(4):31-32. doi: 10.3969/j.issn.1001-4713.2017.04.011 [10] 林士杰, 徐 丹, 崔 珍, 等. 拧筋槭种子甲醇浸提液生物活性测定[J]. 中国农学通报, 2014, 30(10):19-22. doi: 10.11924/j.issn.1000-6850.2013-3036 [11] Valentina Schmitzer, Franci Stampar, Robert Veberic, et al. Phase change modifies anthocyanin synthesis in Acer palmatum Thunb. (Japanese maple) cultivars[J]. Acta Physiologiae Plantarum, 2009, 31(2): 415-418. doi: 10.1007/s11738-008-0237-4 [12] 何 霞, 廖柏勇, 王 芳, 等. 苦楝种源幼林期生长性状地理变异的研究[J]. 华南农业大学学报, 2016, 37(4):75-81. doi: 10.7671/j.issn.1001-411X.2016.04.013 [13] 陆云峰, 王 豪, 徐沁怡, 等. 樟科楠属4种植物种实表型性状及其发芽特性研究[J]. 种子, 2020, 39(12):107-112,119. [14] Xia Xinhe, Yu Xuedan, Fu Qidi, et al. Complete chloroplast genome sequence of the three-flowered maple, Acer triflorum (Sapindaceae)[J]. Mitochondrial DNA Part B, 2020, 5(2): 1859-1860. doi: 10.1080/23802359.2020.1751000 [15] Fabiola Areces-Berazain, Damien D Hinsinger, Joeri S Strijk. Genome-wide supermatrix analyses of maples (Acer, Sapindaceae) reveal recurring inter-continental migration, mass extinction, and rapid lineage divergence[J]. Genomics, 2021, 113(2): 681-692. doi: 10.1016/j.ygeno.2021.01.014 [16] Gao J, Liao P C, Huang B H, et al. Historical biogeography of Acer L. (Sapindaceae): genetic evidence for Out-of-Asia hypothesis with multiple dispersals to North America and Europe[J]. Scientific Reports, 2020, 10(1): 21178. doi: 10.1038/s41598-020-78145-0 [17] 姬志峰, 高亚卉, 李 乐, 等. 山西霍山五角枫不同海拔种群的表型多样性研究[J]. 园艺学报, 2012, 39(11):2217-2228. [18] 王 丹, 庞春华, 高亚卉, 等. 茶条槭不同海拔种群的表型多样性[J]. 云南植物研究, 2010, 32(2):117-125. [19] 赵 阳, 毕泉鑫, 句 娇, 等. 文冠果种子及苗期生长性状地理种源变异[J]. 林业科学研究, 2019, 32(1):160-168. [20] 梁坤南, 黄桂华, 林明平, 等. 琼西南柚木次生种源/家系多性状综合选择[J]. 林业科学研究, 2020, 33(6):13-22. [21] Loveless M D. , Hamrick J L. Ecological Determinants of Genetic Structure in Plant Populations[J]. Annual Review of Ecology and Systematics, 2003, 15(1): 65-95. [22] 冯秋红, 李登峰, 于 涛, 等. 极小种群野生植物梓叶槭的种实表型变异特征[J]. 生物多样性, 2020, 28(3):314-322. doi: 10.17520/biods.2019091 [23] Liang D Y, Ding C J, Zhao G H, et al. Variation and selection analysis of Pinus koraiensis clones in northeast China[J]. Journal of Forestry Research, 2018, 29(3): 611-622. doi: 10.1007/s11676-017-0471-y [24] Zhao X Y, Li Y, Zheng M, et al. Comparative analysis of growth and photosynthetic characteristics of (Populus simonii × P. nigra) × (P. nigra × P. simonii) hybrid clones of different ploidides[J]. PLos One, 2015, 10(4): e0119259. doi: 10.1371/journal.pone.0119259 [25] Zhao X Y, Hou W, Zhang H Q, et al. Analyses of genotypic variation in white poplar clones at four sites in China[J]. Silvae Genetica, 2013, 62(4-5): 187-195. [26] Yin S P, Xiao Z H, ZHAO G H, et al. Variation analyses of growth and wood properties of Larix olgensis clones in China[J]. Journal of Forestry Research, 2017, 28(4): 687-697. doi: 10.1007/s11676-016-0359-2 [27] Wang F, Zhang Q H, Tian Y G, et al. Comprehensive assessment of growth traits and wood properties in half-sib Pinus koraiensis families[J]. Euphytica, 2018, 214(11): 1-15. [28] 孟 超, 郑 昕, 姬志峰, 等. 山西葛萝槭天然种群表型多样性研究[J]. 西北植物学报, 2013, 33(11):2232-2240. doi: 10.7606/j.issn.1000-4025.2013.11.2232 [29] 魏伊楚. 不同产地元宝枫种实表型性状及化学成分研究[D]. 陕西杨凌: 西北农林科技大学, 2018. [30] Gouwakinnou G N, Assogbadjo A E, Lykke A M, et al. Phenotyp-ic variations in fruits and selection potential in Sclerocarya birrea subsp. birrea. Birrea[J]. Scientia Horticulturae, 2011, 129(4): 777-783. doi: 10.1016/j.scienta.2011.05.041 [31] 龙 凤, 余传琼, 吴承祯, 等. 不同种源雷公藤叶物候特征比较[J]. 应用与环境生物学报, 2016, 22(4):660-666. [32] Munthali C R Y, Chirwa P W, Akinnifesi F K. Phenotypic varia-tion in fruit and seed morphology of Adansonia digitata L. (bao-bab) in five selected wild populations in Malawi[J]. Agroforestry Systems, 2012, 85(2): 279-290. doi: 10.1007/s10457-012-9500-1 [33] 杨志玲, 杨 旭, 谭梓峰, 等. 不同野生居群石蒜表型变异及物候期差异[J]. 应用与环境生物学报, 2010, 16(3):369-375. [34] 林 玮, 周 鹏, 周祥斌, 等. 任豆种源种子性状地理变异研究[J]. 华南农业大学学报, 2016, 37(4):69-74. doi: 10.7671/j.issn.1001-411X.2016.04.012 [35] 蒋路平, 王景源, 张 鹏, 等. 170个红松无性系生长及结实性状变异及选择[J]. 林业科学研究, 2019, 32(1):58-64. [36] 王 宁. 野生元宝枫资源调查及种子脂肪酸评价研究[D]. 陕西杨凌: 西北农林科技大学, 2019. [37] 高张莹, 张海峰, 陈国平, 等. 核桃楸种群果核形态及地理变异[J]. 应用与环境生物学报, 2017, 23(4):609-615. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3508
- HTML全文浏览量: 2193
- PDF下载量: 49
- 被引次数: 0







