


-
水分是干旱半干旱地区植物生长和存活的关键因子,在很大程度上决定了植物的生长和存活[1-2]。当土壤水分亏缺时,不同植物表现出不同的忍耐能力,从而表现出不同的生理响应。长期营林实践表明,由于对所选造林树种的水分生理生态特征认识不够,没有很好地做到适地适树原则,从而形成了大量的“小老树”或“早衰”林分,严重影响森林生态效益的发挥[3-5]。因而,如何根据生境选择合适的造林树种是当前植被恢复和重建工作中一个十分重要的科学问题[6-8]。而解决这一问题的关键是要了解不同造林树种的水分利用特性[6,9]。植物水分利用效率是反应植物水分利用状况和抗旱性的一个重要指标,它可为不同生境合适造林树种的选择提供理论依据[10-11]。
叶片稳定性碳同位素组成(δ13C)是表征植物长期水分利用效率的可靠指标[12-13],已经被广泛用于研究陆地各种生态系统植物、植被的水分利用效率[14-16]。因为叶片δ13C整合记录了气候因子和影响植物碳同化以及气孔导度的生理因子的相关信息[17]。Farquhar等研究表明,C3植物叶片δ13C值与植物水分利用效率间存在显著的正相关关系[17]。同一生境下,植物叶片δ13C值高,通常表明植物具有高的水分利用效率,也意味着植物具有较强的抗旱能力[18-19]。在干旱半干旱地区,随着植物水分可利用性降低,植物叶片δ13C值呈增加趋势,水分利用方式更加保守[20-21]。
科尔沁沙地位于我国北方半干旱地区,是我国典型的农牧交错区,也是我国沙漠化最为严重的地区之一[22-23]。为控制沙漠化,改善当地脆弱的生态环境,该地区从1952年后就开始了防护林体系建设[24]。在防护林建设过程中,以杨树(Populus L.)、樟子松(Pinus sylvestris var. mongolica Litv.)、油松(Pinus tabuliformis Carr.)和白榆(Ulmus pumila L.)等树种为主要造林树种,营造了大面积的人工林。由于对这些树种的水分生理特性认识并不十分清楚,且栽植过程中并没有严格遵从适地适树原则,导致樟子松和杨树人工林在干旱年份出现衰退、死亡现象;而相同水分条件下的油松和白榆并没有出现衰退现象[22,25-26]。前期研究结果表明,水分亏缺是引起科尔沁沙地防护林树种衰退的主要原因[22,27]。因此,有必要明确不同树种水分利用特性(例如,水分利用效率)。关于科尔沁沙地造林树种水分利用效率已进行了大量研究,但多是利用气体交换法测定的瞬时水分利用效率[28-29],这些数值并不能很好解释植物长期的生理变化以及水分适应策略。因此,本研究以科尔沁沙地南缘主要造林树种樟子松、彰武小钻杨(Populus × xiaozhuanica cv. Zhangwu)、油松和白榆为研究对象,测定了生长季(6—10月)不同树种叶片δ13C值,同时监测不同树种土壤含水量(0~60 cm),比较了不同树种的水分利用效率(δ13C指示),探讨了水热因子对树木水分利用效率的影响,以期为干旱半干旱沙区植被恢复和重建过程中造林树种的选择提供理论依据。
-
研究区位于科尔沁沙地南缘,辽宁省阜新市彰武县章古台镇辽宁省沙地治理与利用研究所试验基地(42°42′ N, 122°22′ E; 平均海拔226 m)。该区属于半干旱气候区。年均气温7.2 ℃,年降水量479 mm(1954—2019),年蒸发量1 300~1 800 mm[4]。冬春两季风大而持续时间长,风速为4.15~5.10 m·s−1 ,无霜期150 d。土壤为风沙土,pH 值6.7左右,养分元素N、P 等含量低[22]。按地貌特征、地形起伏状况、植物覆盖和土壤性质等因素将章古台沙地划分为4种类型:流动与半流动沙丘、固定沙丘、平缓沙地和风蚀洼地[24]。土壤盐分含量、土壤质地和土壤结构在研究区呈均匀分布。该地区主要树种有樟子松、杨树、油松和白榆等。林下植被以1年生草本植物为主,主要为禾本科(Graminaea)、藜科(Chenopodiaceae)、菊科(Compositae)、莎草科(Cyperaceae)[22]。研究区平均地下水位为5.0 m 左右。2011年,研究期间(6—10月)降水量为361.1 mm,占常年同期降水量(384.7 mm)的94%;监测期间气温平均值为19.1 ℃,略高于同期平均气温(18.7 ℃),平均相对湿度为69%,平均饱和水汽压差为0.66 kPa,光合有效辐射为400.5 μmol·m−2 ·s−1,平均风速为2.0 m·s−1(图1)
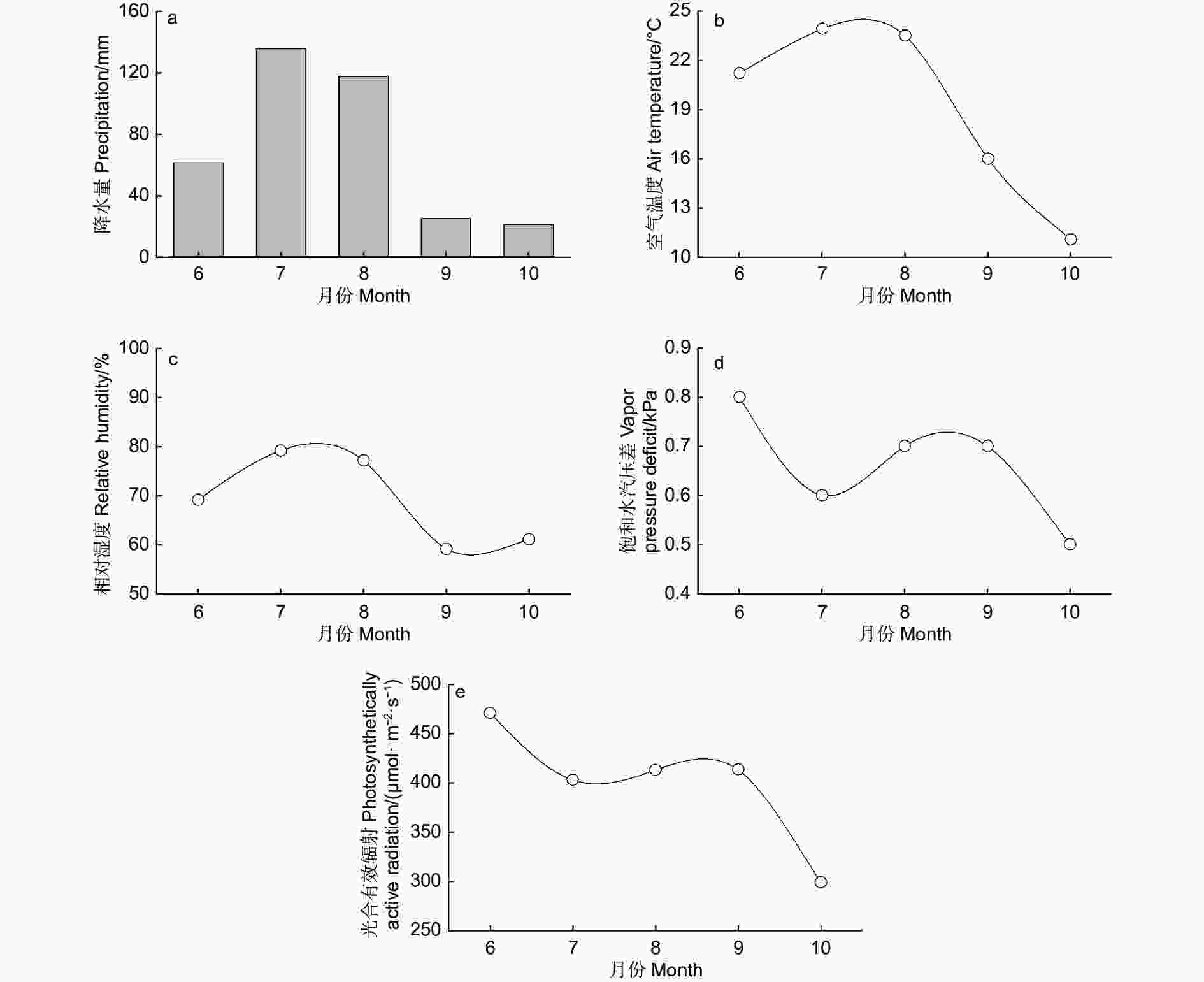
图 1 2011年研究区气象因子季节性变化
Figure 1. Seasonal variation in the climatic variables for the study region in 2011
-
在生长季初期的5月,在对试验区进行全面踏查的基础上,选择31年生片状樟子松林分、12年生彰武小钻杨林带、40年生片状油松林分和42年生散生白榆(表1)。为了减小林分密度对叶片δ13C值的影响,在樟子松、油松和杨树林分内随机选择5株优势木,同时随机选取5株散生白榆,共计20株,挂牌作为研究样树。白榆、油松和樟子松林分属于中龄林,杨树林带接近中龄林(中华人民共和国林业行业标准:主要树种龄级与龄组划分)。于2011年生长季的6 — 10月每月中旬取1次叶片和土壤样品,整个生长季共计5次。
表 1 不同树种样地特征
Table 1. Characteristics of different tree species plots
树种
Species树龄/a
Tree age平均树高/m
Mean height平均胸径/cm
Mean DBH密度/(株·hm−2)
Density取样高度/m
Sampling height樟子松 P. mongolica 31 6.7 11.2 132 8 5.0 彰武小钻杨 P. xiaozhuanica 12 12.0 13.0 — 7.0 油松 P. tabuliformis 40 5.6 18.9 300 4.5 白榆 U. pumila 42 10.2 23.2 — 7.0 -
为了避免取样高度对叶片δ13C影响,不同树种每次取样高度保持基本一致(表1)。每次取样集中于上午8:00—10:00,用高枝剪采集每株样树树冠中部、向阳部位的叶片。樟子松和油松每株样树取当年生叶各100束;杨树和白榆每株样树取30片叶。将取得的叶立即装进信封内,迅速带回实验室。在实验室内用蒸馏水清洗叶片样品,在105 ℃烘箱中杀青20 min,然后在70 ℃烘箱中烘48 h,使样品完全干燥。利用粉碎机(FW100, 天津泰斯特)将叶样品粉碎,过100目筛使样品均一化,以备测定δ13C值。
-
叶片δ13C值在中国科学院沈阳应用生态研究所农产品安全与环境质量检测中心用稳定性同位素比率质谱仪(DELTA plus XP , America)进行测定,测试误差小于0.15 ‰,以PDB(Pee Dee Belemnite)为标准,根据下面公式(1)进行计算:
$ \delta {}^{{\text{13}}}{{\text{C}}_{{\text{Sample}}}}{\text{ = }}\left[ {\frac{{{{{\text{(}}{}^{{\text{13}}}{\text{C/}}{}^{{\text{12}}}{\text{C)}}}_{{\text{Sample}}}}}}{{{{{\text{(}}{}^{{\text{13}}}{\text{C/}}{}^{{\text{12}}}{\text{C)}}}_{{\text{PDB}}}}}}{{ - 1}}} \right]{{ \times 1\;000}} $
(1) 其中, δ13C表示样品13C/12C与标准样品偏离的千分率,(13C/12C)PDB表示南卡罗来纳州白碚石(Pee DeeBelemnite)中的13C/12C。
-
每次叶片样品取样的当天,在每株样树树冠下选择1个样点,用土钻取土。取土深度为0~20,20~40 和40~60 cm。用烘干法测定土壤含水量。
-
用SPSS19.0软件进行双因素方差分析,新复极差法(Duncan)比较树种和取样日期对土壤含水量和当年生叶片δ13C值影响。采用单因素方差分析比较同一月份不同土壤深度土壤含水量和同一月份不同树种叶片δ13C值的差异。利用Pearson相关性分析方法分析不同树种叶δ13C逐月变化量(将当月与前月叶片δ13C值相减)与水热因子关系。显著性水平设定为α=0.05。
-
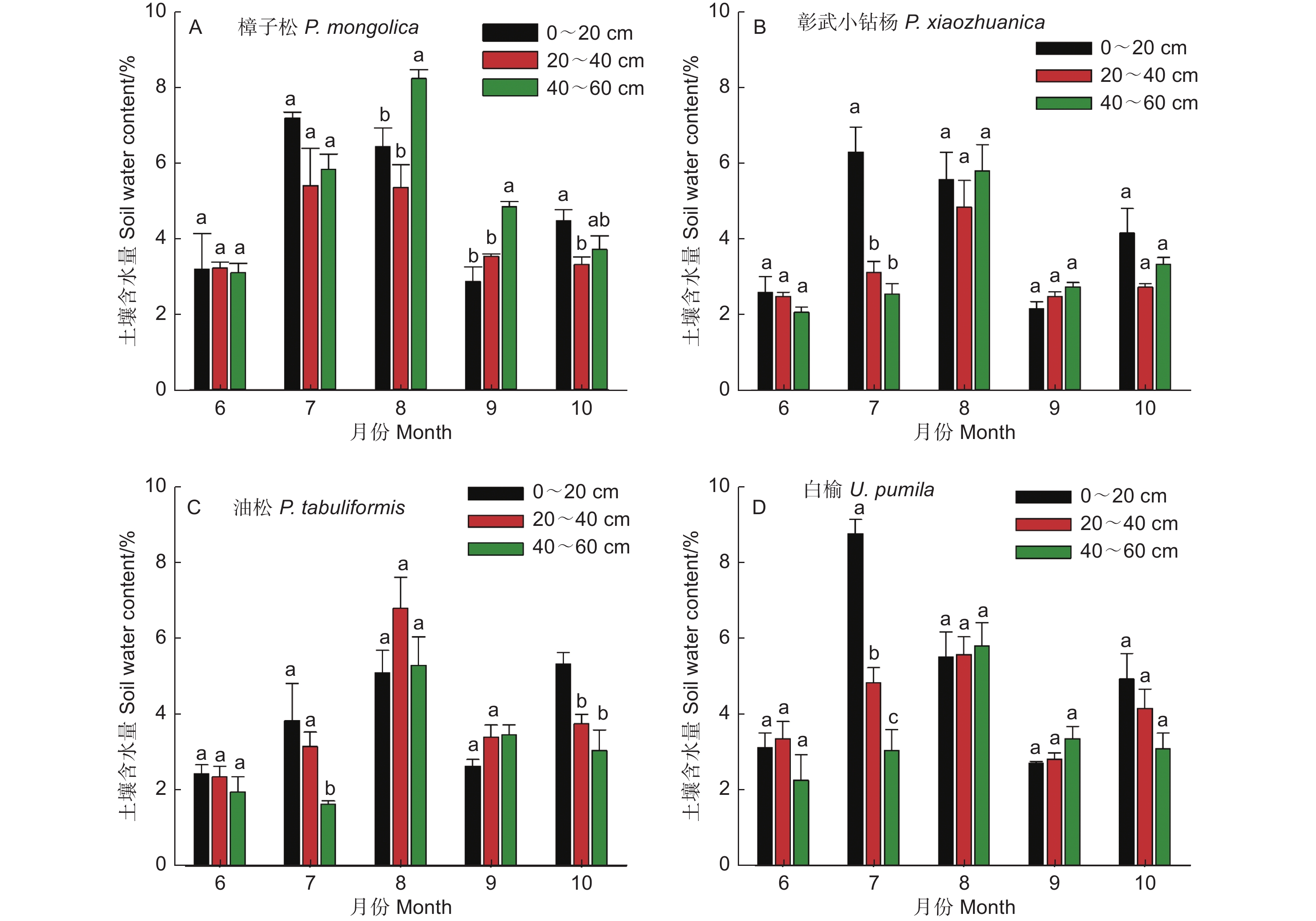
不同树种土壤含水量(0~60 cm)随着季节变化呈先增加后降低趋势(图2)。樟子松和彰武小钻杨土壤含水量按月份排序均为:8月和7月>10月、9月和6月(图2A 和2B, P<0.05);油松土壤含水量按月份排序为:8月>10月和9月>7月和6月(图2C, P<0.05);白榆土壤含水量按月份排序为:8月和7月>10月>6月和9月(图2D, P<0.05)。双因素方差分析结果表明(表2),不同树种和月份间土壤含水量存在显著性差异。多重比较结果表明(表3),樟子松和白榆的土壤含水量显著高于彰武小钻杨和油松;土壤含水量按月份排序为:8月>7月> 10月>6月和9月。
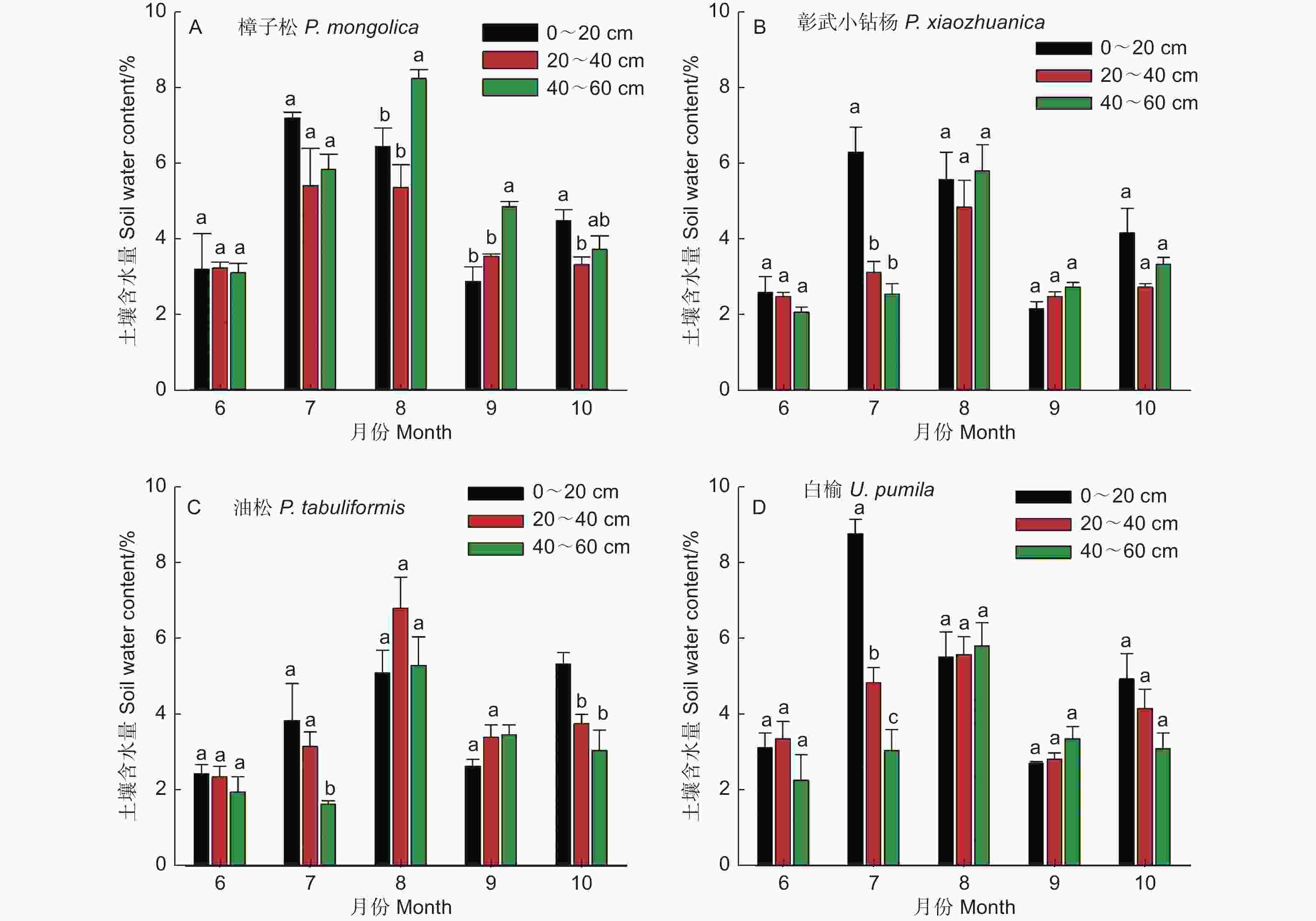
图 2 不同树种土壤含水量(0~60 cm)季节动态
Figure 2. Seasonal variations in soil water content at 0-60 cm depth for different tree species
表 2 树种和月份对土壤含水量(0~60 cm)和叶片δ13C影响的双因素方差分析
Table 2. Two-way ANOVA analysis for the effect of species and month on soil water content at the depth of 0~60 cm and leaves δ13C
差异来源
Sources of differences土壤含水量/%
Soil water content叶δ13C/‰
Leaf δ13C自由度 df F P 自由度 df F P 树种 Species 3 9.76 <0.001 3 103.53 <0.001 月份 Month 4 62.54 <0.001 4 12.33 <0.001 树种 × 月份 Species × Month 12 3.98 <0.001 12 3.39 <0.001 误差 Error 80 80 -
不同月份樟子松叶片δ13C值介于−27.44‰~−26.36‰,平均值为−26.92‰;彰武小钻杨叶片δ13C值介于−27.87‰~−26.97‰,平均值为−27.40‰;油松叶片δ13C值介于−28.00‰~−26.72‰,平均值为−27.57‰;白榆叶片δ13C值介于−30.02‰~−28.45‰,平均值为−29.23‰(图3A)。樟子松、油松和白榆叶δ13C值月变化量(将当月与前月叶片δ13C值相减)生长季几乎都为负值,而彰武小钻杨叶δ13C值月变化7月和8月为正值,9月和10月为负值(图3B)。樟子松、油松和白榆叶δ13C值不同月份间差异都显著(P<0.05),排序都为:6月和7月> 8月、9月和10月;彰武小钻杨叶δ13C值不同月份间差异显著(P<0.05),排序为:8、7和9月>6月和10月。同一月份不同树种间叶δ13C值存在显著性显著(图3A, P<0.05),但是这种差异随月份变化而不同。双因素方差分析结果表明(表2),不同树种和月份间叶δ13C值(整个生长季平均值)存在显著性差异(P<0.05)。多重比较结果表明(表3),樟子松叶δ13C值显著高于彰武小钻杨和油松,而彰武小钻杨和油松显著高于白榆;6月和7月叶δ13C显著高于8月和9月,显著高于10月。
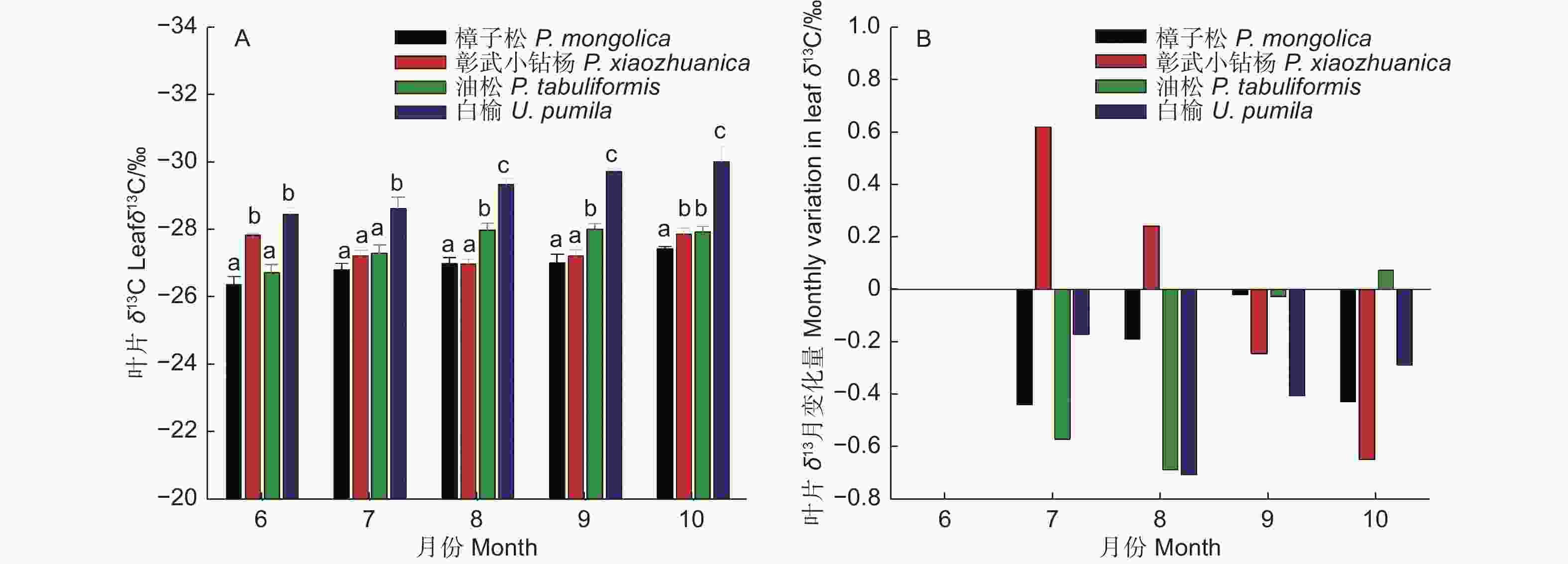
图 3 不同树种叶片δ13C值季节动态和月变化量
Figure 3. Seasonal changes in the leaf δ13C values and leaf δ13C monthly change values for different tree species
表 3 不同树种逐月δ13C变化量与各月气象要素的相关系数及P 值
Table 3. Correlation coefficients and p values between leaf δ13C monthly change values and environmental variables
树种
Tree species降水量
Precipitation气温
Temperature相对湿度
RH饱和水汽压差
VPD光合有效辐射
PAR土壤水分
SM樟子松 P. mongolica r −0.295 0.010 −0.359 0.841 0.591 −0.201 P 0.705 0.990 0.641 0.159 0.409 0.799 彰武小钻杨 P. xiaozhuanica r 0.948 0.968 0.907 0.410 0.719 0.531 P 0.052 0.032 0.093 0.590 0.281 0.469 油松 P. tabuliformis r −0.966 −0.964 −0.962 −0.443 −0.629 −0.392 P 0.034 0.036 0.038 0.557 0.508 0.608 白榆 U. pumila r −0.115 −0.258 −0.136 −0.653 −0.368 −0.165 P 0.885 0.742 0.864 0.347 0.632 0.835 RH: relative humidity, VPD: vapor pressure deficit, PAR: photosynthetically active radiation; SM: soil moisture -
将各月δ13C变化量与各月气象要素和土壤含水量进行相关分析。樟子松叶片δ13C变化量与降水量、相对湿度和土壤含水量呈负相关关系,与温度、饱和水汽压差和光合有效辐射呈正相关关系,相关关系都不显著(表3)。彰武小钻杨叶片δ13C变化量与气温呈显著正相关关系(P<0.05),与其他水热因子也呈正相关关系,相关关系不显著(表3)。油松叶片δ13C变化量与降水量、气温和相对湿度呈显著负相关关系(P<0.05),与其他水热因子也呈负相关关系,相关关系不显著(表3)。白榆叶片δ13C变化量与水热因子呈负相关关系,相关关系都不显著(表3)。
-
本研究发现,樟子松、油松和白榆叶片δ13C值随着季节性变化呈下降趋势,而彰武小钻杨叶片δ13C值随着季节变化呈先升高后降低趋势,表明樟子松、油松和白榆水分利用效率随着季节变化逐渐下降,而彰武小钻杨水分利用效率随着季节变化呈先增加后降低(图3)。这与其他研究结果基本一致。例如,郭树江等[30]研究发现,民勤3种主要灌木植物水分利用效率随着季节变化呈降低趋势,主要与平均气温、相对湿度、蒸发量等环境因子季节性变化有关。曹生奎等[31]研究表明,极端干旱区胡杨(Populus euphratica Oliv.)生长季水分利用效率逐渐下降,主要是因为气温升高、土壤含水量减小和地下水位埋深下降导致的气孔导度的变化引起的。方晓娟等[32]研究发现随着季节变化,毛白杨(Populus tomentosa)杂种无性系水分利用效率呈先增加后降低趋势。本研究中不同树种水分利用效率随季节变化有所不同,这可能与不同树种对外界水热因子季节性变化响应程度不同有关[33]。樟子松、油松和白榆叶片δ13C变化量与降水量、相对湿度和土壤含水量呈负相关关系,尤其是油松(表3),表明当土壤含水量降低, 空气湿度降低以及降水量不足时, 树木为了减少水分的蒸发, 降低气孔导度,提高水分利用效率[33]。另外,油松叶片δ13C变化量与温度呈显著负相关关系,表明随着温度升高,油松水分利用效率降低。这与路伟伟等[34]研究结果一致,即随着气温升高,北京市密云东部山区油松水分利用效率降低。一般情况下,干旱半干旱地区降水对树木水分利用效率的影响程度高于气温[10]。6月份降水量和土壤含水量相对较低(图1),但是温度相对较高,土壤蒸发也强烈,因此,6月份,樟子松、油松和白榆为减少水分丧失,维持较低的气孔导度,从而具有较高的水分利用效率。6月份以后,由于降水量的增加,土壤含水量显著增加,能被树木利用的水分增加,树木气孔导度增加,蒸腾强度增加,导致树木在生长旺盛期的水分利用效率降低。8月之后,随着降水量减少和温度降低(图1),尽管土壤含水量降低,但是树木蒸腾需求量逐渐降低。因此,树木水分利用效率仍呈降低趋势[35]。另外,彰武小钻杨叶片δ13C变化量与温度呈显著正相关关系,表明随着温度升高,促使彰武小钻杨气孔导度降低,水分利用效率增加。这与Sun 等[36]研究结果一致,随着温度升高,张北地区小叶杨(Populus simonii Carr.)水分利用效率显著升高。6—8月,随着温度和光照强度增加,彰武小钻杨光合速率和蒸腾能力逐渐增加[32],尽管降水量增加,但是土壤含水量远远不能满足其蒸腾耗水的需要,彰武小钻杨通过降低气孔导度,从而提高水分利用效率,维持树木生长。8月份后,随着光照强度和温度逐渐降低,光合速率和蒸腾能力逐渐降低,但是光合速率的降低程度大于蒸腾速率[37],从而导致彰武小钻杨水分利用效率呈降低趋势。
另外,彰武小钻杨叶片δ13C变化量与相对湿度和饱和水汽压差呈正相关关系,而油松和白榆叶片δ13C变化量与相对湿度和饱和水汽压差呈负相关关系(表3),这表明不同树种叶片δ13C对空气相对湿度和饱和水汽压差变化具有不同的响应。这可能是因为随着空气相对湿度降低,彰武小钻杨叶片羧化效率降低,从而导致叶片δ13C降低;伴随着饱和水汽压差增加,气孔导度降低,引起彰武小钻杨叶片δ13C升高。然而,随着空气相对湿度降低,油松和白榆气孔导度和胞间CO2浓度降低,导致其对13C分辨率下降,从而引起叶片δ13C值增加;同时,饱和水气压差升高,引起气孔导度下降,但是饱和水汽压差升高对光合速率限制作用可能高于气孔导度,从而导致油松和白榆叶片δ13C值降低。由于叶片δ13C是环境因子与生物因子综合作用的结果,有关树木叶片δ13C与水热因子关系有待于进一步研究。
-
本研究结果表明,樟子松叶δ13C值显著高于彰武小钻杨和油松,高于白榆,表明樟子松具有较高的水分利用效率,彰武小钻杨和油松次之,散生白榆最低。这与其他人的研究结果基本一致,例如,丁晓纲等[38]比较了毛乌素沙地腹地樟子松和油松人工林生长季水分利用效率,发现生长季樟子松水分利用效率始终高于油松。高素华等[39]研究表明,北方多粗沙区杨树水分利用效率显著高于白榆。本研究中,尽管樟子松土壤含水量较高,但是樟子松具有较高的水分利用效率,主要是因为樟子松叶片具有发达的角质层、气孔小而多、比叶面积小等特征[40],导致其扩散效应能力低,从而具有较高的水分利用效率。另外,与其他树种相比,樟子松根系分布较浅[22,41],容易受到干旱胁迫的威胁;因而采取更加保守的水分利用方式,从而具有较高的水分利用效率。尽管彰武小钻杨是落叶阔叶树种,但是其水分利用效率与针叶树种油松没有明显差异(图3),主要是因为彰武小钻杨的根系分布比油松深[22],可以利用深层土壤水和地下水;但是彰武小钻杨蒸腾耗水量高于油松,且油松和杨树0~60 cm土壤含水量并没有显著差异,因而彰武小钻杨和油松具有相似的水分利用效率。油松的气孔开度大于樟子松,而气孔密度小于樟子松[40],从而导致其扩散能力高于樟子松。因此,油松的水分利用效率低于樟子松。与其他树种相比,白榆具有最低的水分利用效率,可能是因为白榆叶片的气孔导度和比叶面积较大,其扩散能力高,导致白榆具有较低的水分利用效率。例如,陈仁升等[42]研究表明,西北干旱区榆树的气孔导度显著高于樟子松和二白杨(Populus gansuensis C. Wang et H. L. Yang)。另外,白榆的土壤含水量高于杨树和油松,也可能是其具有较低水分利用效率原因之一。
-
樟子松的水分利用效率显著高于彰武小钻杨和油松,而彰武小钻杨和油松水分利用效率显著高于白榆。随着季节变化,樟子松、油松和白榆水分利用效率呈降低趋势,而彰武小钻杨水分利用效率呈先增加后降低趋势。叶片δ13C逐月变化量与水热因子的响应程度因树种而异,樟子松和白榆叶δ13C逐月变化量与水热因子相关关系不显著,而油松叶δ13C逐月变化量与降水量、气温和湿度呈显著负相关关系,而彰武小钻杨叶δ13C逐月变化量与气温呈显著正相关关系。不同树种水分利用效率差异不仅与树种生物学特性有关,而且也与受外界水热因子影响程度有关。因此,造林树种选择过程中,不仅要考虑树种生物学特性,还应该考虑树种对外界水热因子响应程度。
科尔沁沙地主要造林树种叶片δ13C比较研究
Leaf δ13C of the Main Afforestation Tree Species in Horqin Sandy Land
-
摘要:
目的 明确不同造林树种水分利用特征差异,为干旱半干旱沙区植被恢复和重建过程中造林树种的选择提供理论依据。 方法 通过测定科尔沁沙地南缘主要造林树种樟子松(Pinus sylvestris var. mongolica)、彰武小钻杨(Populus × xiaozhuanica)、油松(Pinus tabuliformis)和白榆(Ulmus pumila)生长季(6—10月)叶片δ13C值和不同深度土壤含水量,比较了不同树种的水分利用效率(δ13C指示),并探讨了水热因子对树木水分利用效率的影响。 结果 樟子松、油松和白榆叶片δ13C值随季节变化逐渐降低,而彰武小钻杨叶片δ13C值随季节变化呈先增加后降低趋势;从整个生长季δ13C平均值来看,樟子松叶片δ13C值(−26.92‰ ± 0.11‰)显著高于彰武小钻杨(−27.40 ‰± 0.10‰)和油松(−27.57 ‰± 0.14‰),而彰武小钻杨和油松显著高于白榆(−29.23‰ ± 0.17‰)。叶片δ13C逐月变化量与水热因子的响应程度因树种而异,樟子松和白榆叶δ13C逐月变化量与水热因子相关关系不显著,而油松叶δ13C逐月变化量与降水量、气温和湿度呈显著负相关关系,彰武小钻杨叶δ13C逐月变化量与气温呈显著正相关关系。 结论 樟子松具有较高的水分利用效率,彰武小钻杨和油松次之,白榆最低。不同树种水分利用效率差异不仅与树种生物学特性有关,而且与受外界水热因子响应程度有关。 Abstract:Objective Determining the water use characteristics of different afforestation tree species would supply a theoretical basis for the selection of tree species in vegetation restoration and reconstruction in semiarid and arid sandy lands. Method Leaf δ13C values of Pinus sylvestris var. mongolica, Populus × xiaozhuanica, Pinus tabuliformis, and Ulmus pumila trees in the south edge of the Horqin Sandy Land were measured together with monitoring soil water content at different depths to compare their water use efficiency (indicative of leaf δ13C) during the growing season, and the effect of hydrothermal factors on the water use efficiency was discussed. Result The leaf δ13C values of P. sylvestris var. mongolica, P. tabuliformis, and U. pumila trees decreased with seasons, whereas the leaf δ13C values of P. × xiaozhuanica firstly increased and then declined during the measurement period. In addition, the mean leaf δ13C value was higher in P. sylvestris var. mongolica trees (−26.92‰ ± 0.11‰) than in P. × xiaozhuanica (−27.40 ‰± 0.10‰) and P. tabuliformis (−27.57 ‰± 0.14‰) trees, whereas the mean leaf δ13C value was higher in P. × xiaozhuanica and P. tabuliformis trees than in U. pumila trees (−29.23 ‰± 0.17‰). The responses of the leaf δ13C monthly values to hydrothermal factors varied with tree species. There were no significant relationships between leaf δ13C monthly values and hydrothermal factors for P. sylvestris var. mongolica and U. pumila trees, whereas a significant negative relationship between monthly change in leaf δ13C and precipitation, air temperature, or relative humidity was observed for P. tabuliformis trees. However, a significant positive relationship between leaf δ13C monthly change values and air temperature was found for P. × xiaozhuanica trees. Conclusion The water use efficiency is the highest in P. sylvestris var. mongolica trees, followed by P. × xiaozhuanica and P. tabuliformis trees and the lowest value in U. pumila trees. In addition, water use efficiency for different tree species is not only related to biological characteristics but also related to the degree of influence by hydrothermal factors. -
图 1 2011年研究区气象因子季节性变化
Figure 1. Seasonal variation in the climatic variables for the study region in 2011
图 2 不同树种土壤含水量(0~60 cm)季节动态
Figure 2. Seasonal variations in soil water content at 0-60 cm depth for different tree species
图 3 不同树种叶片δ13C值季节动态和月变化量
Figure 3. Seasonal changes in the leaf δ13C values and leaf δ13C monthly change values for different tree species
表 1 不同树种样地特征
Table 1. Characteristics of different tree species plots
树种
Species树龄/a
Tree age平均树高/m
Mean height平均胸径/cm
Mean DBH密度/(株·hm−2)
Density取样高度/m
Sampling height樟子松 P. mongolica 31 6.7 11.2 132 8 5.0 彰武小钻杨 P. xiaozhuanica 12 12.0 13.0 — 7.0 油松 P. tabuliformis 40 5.6 18.9 300 4.5 白榆 U. pumila 42 10.2 23.2 — 7.0  下载: 导出CSV
下载: 导出CSV
表 2 树种和月份对土壤含水量(0~60 cm)和叶片δ13C影响的双因素方差分析
Table 2. Two-way ANOVA analysis for the effect of species and month on soil water content at the depth of 0~60 cm and leaves δ13C
差异来源
Sources of differences土壤含水量/%
Soil water content叶δ13C/‰
Leaf δ13C自由度 df F P 自由度 df F P 树种 Species 3 9.76 <0.001 3 103.53 <0.001 月份 Month 4 62.54 <0.001 4 12.33 <0.001 树种 × 月份 Species × Month 12 3.98 <0.001 12 3.39 <0.001 误差 Error 80 80
下载: 导出CSV
表 3 不同树种逐月δ13C变化量与各月气象要素的相关系数及P 值
Table 3. Correlation coefficients and p values between leaf δ13C monthly change values and environmental variables
树种
Tree species降水量
Precipitation气温
Temperature相对湿度
RH饱和水汽压差
VPD光合有效辐射
PAR土壤水分
SM樟子松 P. mongolica r −0.295 0.010 −0.359 0.841 0.591 −0.201 P 0.705 0.990 0.641 0.159 0.409 0.799 彰武小钻杨 P. xiaozhuanica r 0.948 0.968 0.907 0.410 0.719 0.531 P 0.052 0.032 0.093 0.590 0.281 0.469 油松 P. tabuliformis r −0.966 −0.964 −0.962 −0.443 −0.629 −0.392 P 0.034 0.036 0.038 0.557 0.508 0.608 白榆 U. pumila r −0.115 −0.258 −0.136 −0.653 −0.368 −0.165 P 0.885 0.742 0.864 0.347 0.632 0.835 RH: relative humidity, VPD: vapor pressure deficit, PAR: photosynthetically active radiation; SM: soil moisture
下载: 导出CSV
-
[1] 朱教君. 防护林学研究现状与展望[J]. 植物生态学报, 2013, 37(9):872-888. [2] 严昌荣, 韩兴国, 陈灵芝. 六种木本植物水分利用效率和其小生境关系研究[J]. 生态学报, 2001, 21(11):1952-1956. doi: 10.3321/j.issn:1000-0933.2001.11.028 [3] Zhu J J, Song L N. A review of ecological mechanisms for management practices of protective forests[J]. Journal of Forestry Research, 2021, 32(2): 435-448. doi: 10.1007/s11676-020-01233-4 [4] 曹 宇, 巢 林, 安宇宁, 等. 科尔沁沙地刺榆水力结构特征对土壤水分环境的响应[J]. 林业科学, 2021, 57(7):32-42. doi: 10.11707/j.1001-7488.20210704 [5] 朱教君, 郑 晓. 关于三北防护林体系建设的思考与展望——基于40年建设综合评估结果[J]. 生态学杂志, 2019, 38(5):1600-1610. [6] 车力木格, 刘新平, 何玉惠, 等. 半干旱沙地草本植物群落特征对短期降水变化的响应[J]. 草业学报, 2020, 29(4):19-28. doi: 10.11686/cyxb2019155 [7] 王宇祥, 刘廷玺, 段利民, 等. 基于Hydrus-1D模型的科尔沁沙地沙丘-草甸相间区土壤水分动态模拟[J]. 中国沙漠, 2020, 40(2):195-205. [8] 刘小英, 段爱国, 张建国, 等. 不同种源杉木树轮α 纤维素δ13C 对年气候因子的响应[J]. 林业科学研究, 2020, 33(2):9-18. [9] 赵 娜, 李少宁, 徐晓天, 等. 北京地区典型绿化树种水分利用效率及其影响因素[J]. 北京林业大学学报, 2021, 43(3):44-54. doi: 10.12171/j.1000-1522.20200293 [10] 王云霓, 熊 伟, 王彦辉, 等. 干旱半干旱地区主要树种叶片水分利用效率研究综述[J]. 世界林业研究, 2012, 25(2):17-23. [11] 何春霞, 张劲松, 孟 平, 等. 太行山南麓3种常见灌木的水分利用特性[J]. 林业科学, 2018, 54(9):137-145. doi: 10.11707/j.1001-7488.20180916 [12] 沈芳芳, 樊后保, 吴建平, 等. 植物叶片水平δ13C与水分利用效率的研究进展[J]. 北京林业大学学报, 2017, 39(11):114-124. [13] 刘 飞, 刘 攀, 曹 铭, 等. 稳定同位素技术在植物水分关系研究中的应用综述[J]. 生态科学, 2021, 39(6):224-232. [14] 吕春平, 张秋芳, 郝亚群, 等. 短期增温对亚热带杉木幼林碳氮同位素组成的影响[J]. 林业科学研究, 2018, 31(5):27-32. [15] 周 佳, 孟 平, 张劲松, 等. 河南民权与陕西白水刺槐径向生长与水分利用效率对气候响应的差异[J]. 林业科学研究, 2021, 34(6):1-8. [16] 何春霞, 陈 平, 张劲松, 等. 太行山南麓核桃-菘蓝/决明复合系统种间水分关系[J]. 林业科学研究, 2021, 34(4):22-31. [17] Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis[J]. Annual Review of Plant Biology, 1989, 40: 503-537. doi: 10.1146/annurev.pp.40.060189.002443 [18] 赵 丹, 程军回, 刘耘华, 等. 荒漠植物梭梭稳定碳同位素组成与环境因子的关系[J]. 生态学报, 2017, 37(8):2743-2752. [19] Brienen R J W, Gloor E, Clerici S, et al. Tree height strongly affects estimates of water-use efficiency responses to climate and CO2 using isotopes[J]. Nature Communications, 2017, 8: 288. doi: 10.1038/s41467-017-00225-z [20] Wang J, Fu B J, Wang L X, et al. Water use characteristics of the common tree species in different plantation types in the Loess Plateau of China[J]. Agricultural and Forest Meteorology, 2020, 288–289:108020 [21] Imin B, Dai Y, Shi Q D, et al. Responses of two dominant desert plant species to the changes in groundwater depth in hinterland natural oasis, Tarim Basin[J]. Ecology and Evolution, 2021, 11(14): 9460-9471. doi: 10.1002/ece3.7766 [22] Song L N, Zhu J J, Li M C, et al. Comparison of water-use patterns for non-native and native woody species in a semiarid sandy region of Northeast China based on stable isotopes[J]. Environmental and Experimental Botany, 2020, 174: 103923. doi: 10.1016/j.envexpbot.2019.103923 [23] 陈 晨, 王 寅, 王健铭, 等. 科尔沁沙地植物群落物种多样性及其主要影响因素[J]. 北京林业大学学报, 2020, 42(5):106-114. doi: 10.12171/j.1000-1522.20190284 [24] 焦树仁. 章古台固沙林生态系统的结构与功能[M]. 沈阳: 辽宁科学技术出版社, 1989. [25] 焦树仁. 科尔沁沙地极端气候条件对外来树种影响的研究[J]. 防护林科技, 2006,(6):15-17. doi: 10.3969/j.issn.1005-5215.2006.06.005 [26] 王 凯, 逄迎迎, 吕林有, 等. 杨树幼苗自然干旱过程中非结构性碳水化合物变化[J]. 生态学杂志, 2021, 40(7):1969-1978. [27] Zheng X, Zhu J J, Yan Q L, et al. Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land, Northeast China[J]. Agricultural Water Management, 2012, 109: 94-106. doi: 10.1016/j.agwat.2012.02.010 [28] 魏雅芬, 方 杰, 赵学勇, 等. 科尔沁沙地樟子松人工林不同年龄针叶生理生态性状[J]. 植物生态学报, 2011, 35(12):1271-1280. [29] 刘建华. 干旱胁迫对杨树幼苗生长的影响[J]. 防护林科技, 2016,(6):8-11. [30] 郭树江, 王 飞, 张卫星, 等. 民勤4种主要灌木植物δ13C季节变化及与气象因子的关系[J]. 西北林学院学报, 2020, 35(5):41-46. doi: 10.3969/j.issn.1001-7461.2020.05.07 [31] 曹生奎, 冯 起, 司建华, 等. 不同立地条件下胡杨叶片稳定碳同位素组成及水分利用效率的变化[J]. 冰川冻土, 2012, 34(1):155-160. [32] 方晓娟, 李吉跃, 聂立水, 等. 毛白杨杂种无性系稳定碳同位素值的特征及其水分利用效率[J]. 生态环境学报, 2009, 18(6):2267-2271. doi: 10.3969/j.issn.1674-5906.2009.06.049 [33] 赵良菊, 肖洪浪, 刘晓宏, 等. 沙坡头不同微生境下油蒿和柠条叶片δ13C的季节变化及其对气候因子的响应[J]. 冰川冻土, 2005, 27(5):747-754. [34] 路伟伟, 余新晓, 贾国栋, 等. 基于树轮 δ13C 值的北京山区油松水分利用效率[J]. 生态学报, 2017, 37(6):2093-2100. [35] 张日升, 刘 敏. 半干旱地区主要造林树种蒸腾耗水特点的研究进展[J]. 防护林科技, 2008 ,(1):69-71. doi: 10.3969/j.issn.1005-5215.2008.01.027 [36] Sun S J, Qiu, L F, He C X, et al. Drought-affected Populus simonii Carr. show lower growth and long-term increases in intrinsic water-use efficiency prior to tree mortality[J]. Forests, 2018, 9: 564. doi: 10.3390/f9090564 [37] Song L N, Zhu J J, Zhang T, et al. Higher canopy transpiration rates induced dieback in poplar (Populus × xiaozhuanica) plantations in a semiarid sandy region of Northeast China[J]. Agricultural Water Management, 2021, 243: 106414. doi: 10.1016/j.agwat.2020.106414 [38] 丁晓纲, 何 茜, 李吉跃, 等. 毛乌素沙地樟子松和油松人工林光合生理特性[J]. 水土保持研究, 2011, 18(1):215-219. [39] 高素华, 郭建平, 康玲玲, 等. 我国北方多沙粗沙区常见树种水分利用效率的研究[J]. 中国生态农业学报, 2006, 14(1):90-92. [40] 吴祥云, 王晓娇, 李宏昌, 等. 科尔沁沙地主要造林树种抗旱生理生态特性[J]. 东北林业大学学报, 2008, 36(3):3-4. doi: 10.3969/j.issn.1000-5382.2008.03.002 [41] 刘新平, 张铜会, 何玉惠, 等. 科尔沁沙地三种常见乔木根-土界面水分再分配初探[J]. 生态环境学报, 2009, 18(6):2360-2365. doi: 10.3969/j.issn.1674-5906.2009.06.065 [42] 陈仁升, 康尔泗, 赵文智, 等. 中国西北干旱区树木蒸腾对气象因子的响应[J]. 生态学报, 2004, 24(3):477-485. doi: 10.3321/j.issn:1000-0933.2004.03.013 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3711
- HTML全文浏览量: 2180
- PDF下载量: 53
- 被引次数: 0

