

-
自然条件下,林下光环境是林下更新的重要驱动因子之一,对林下幼苗建成、生长及生物量分配具有重要影响[1-4]。同时,温室育苗过程中容易形成弱光环境,影响叶片的光合同化过程,不利于植株的形态建成[5]。因此,研究人工补光措施对林木培育有重要意义。近年来,许多学者已针对光环境对树木生长的影响做了大量的研究[6-10]。光周期作为季节性休眠的诱导因子[9],影响叶片对光合有效辐射(PAR)吸收积累的时间长度,会对物质生产产生显著影响[11];而光质以触发植物体内的光信号受体的方式,影响植物体内相关激素含量和相关酶活性[12-13],进而影响植物的物质合成、新陈代谢和生长发育等[8]。在自然光环境下,在高密度环境下生长的植物可感知入射光红光远红光比率(R∶FR)降低,这是由于冠层叶片吸收红光和来自邻近植物的远红光反射所致,而低R∶FR会诱导植物产生避阴反应(SAS),即在冠层下方遮阴处或有较高邻近植物的环境中生长的植物在感应到低R∶FR后,表现出如茎和叶柄伸长、叶面积增加等现象[1,14-16],促其叶片投射到日光未衰减的区域。此外,植物对不同光周期或光质条件的生理响应也存在种间和种内差别[17-21]。然而,目前针对植物幼苗对光周期和光质及其交互作用机制的研究还较少,需要进一步研究来确定其对植物光合及生长的影响机理。
栎属(Quercus L.)是北半球天然分布最广的木本植物类群之一。我国拥有丰富的栎属植物资源,据第九次森林资源清查[22],我国栎类林在森林面积和蓄积量均列10大树种组的首位;栎类树种作为我国暖温带和温带森林的重要建群种[23],在涵养水源、保持水土和维持生物多样性等方面发挥了重要功能。同时,由于栎属植物普遍具有较高的抗逆性[24],在困难立地造林和园林绿化中应用广泛[25]。然而,大量研究[26]表明,在栎类天然林分中,由于种子及幼苗有较大比例被动物取食[27],且幼苗早期生长受到林分密度[28]、光照[7]等微环境的限制,导致天然更新困难;而在人工育苗实践中,不同栎类树种幼苗的适宜受光条件尚缺乏深入研究。在栎类树种中,栓皮栎(Q. variabilis)和短柄枹栎(Q. glandulifera)分布较广,在我国大部分地区均有分布[29];蒙古栎(Q. mongolica)则主要分布在东北及华北。因此,本研究以3个落叶栎属植物为例,通过LED人工光源的诱导,来探讨不同光周期与光质比对栎属植物幼苗的生长及叶绿素荧光参数的影响及种间和种内的响应差异,为栎属植物育苗壮苗及人工辅助更新提供科学参考。
-
2018年9—10月,选择5个不同纬度的栎属植物天然林分收集种子,包括3个栓皮栎种源(Qv):辽宁大连(LN,北方种源)、陕西太白(SX,中部种源)、云南安宁(YN,南方种源);1个蒙古栎种源(Qm,黑龙江加格达奇)和1个短柄枹栎种源(Qg,河南内乡)(表1)。各种源地地理气候信息见表2。将采集到的种子用60℃温水浸泡2 h杀虫处理后阴干至表面干燥,然后将种子储存至4℃冰箱。2019年春,选择健康且大小均一的种子进行催芽以缩短发芽时间。发芽后于同一天播种于定制育苗容器(内径7 cm,深度36 cm)内预培养2个月,每容器播1粒,培养基质为珍珠岩。预培养期间,每3日浇1次1/2倍浓度的Hoagland标准营养液来提供幼苗生长所需养分,同时观测生长动态。试验在中国林科院科研温室中进行(40°0′10′′ N,116°14′38′′ E),温室昼夜温度约为25/20℃,湿度约为70%/85%,透光度为50%~60%。
表 1 主要缩略词中英对照
Table 1. List of main abbreviations
缩略词
Abbreviations英文全称/含义
English name/meaning中文全称/含义
Chinese name/meaningPAR Photosynthetically active radiation 光合有效辐射 R:FR Red / far red light ratios 红光/远红光的光质比 SAS Shade avoidance syndrome 避阴反应 PP Photoperiod 光周期 LQ Light quality ratio 光质比 IS Interspecies 种间 IP Inter provenances 种源间 Qv Q. Variabilis 栓皮栎 Qm Q.Mongolica 蒙古栎 Qg Q. Glandulifera 短柄枹栎 LN Liaoning 辽宁大连,北方栓皮栎种源 SX Shaanxi 陕西太白,中部栓皮栎种源 YN Yunnan 云南安宁,南方栓皮栎种源 S Short photoperiod 短光周期,日夜时长分别为10 h/14 h M Medium photoperiod 中光周期,日夜时长分别为14 h/10 h L Long photoperiod 长光周期,日夜时长分别为8 h/6 h D Low light quality ratio 低光质比,R/FR为0.83 ± 0.04 N Normal light quality ratio 正常光质比(即日光下光质比),R/FR为1.37 ± 0.04 H High light quality ratio 高光质比,R/FR为4.69 ± 0.04 RGR Relative growth rate 相对生长速率 Fo、Fj、Fi、Fm / O、J、I、P四个拐点的荧光强度 Vj Relative variable fluorescence at the J-step J 相的相对可变荧光 Vi Relative variable fluorescence at 30 ms I 相的相对可变荧光 Fv/Fo Conformation term for primary photochemistry PS II潜在活性 Mo Approximated initial slope (in m·s−1) of the fluorescence
transient normalized on the maximal variable fluorescence FvOJIP 荧光诱导曲线的初始斜率 Ψo Openness of active reaction center 有活性反应中心的开放程度。 φPo Maximum photochemical efficiency 最大光化学效率 φEo Quantum yield 电子传递链的量子产额 φDo Maximum quantum yield for non-photochemical quenching 非光化学淬灭最大量子产额 ABS/RC Light energy absorbed by unit reaction center; 单位反应中心吸收的光能 TRo/RC Light energy captured by unit reaction center; 单位反应中心捕获的光能 ETo/RC Unit reaction center for electron transfer energy 单位反应中心用于电子传递的能量 DIo/RC The energy dissipated per unit reaction center 单位反应中心耗散掉的能量 PIAbs Photosynthetic performance index 以吸收光能为基础的光合性能指数 -
将5个种源幼苗分别进行不同光周期处理[日夜时长分别为10 h/14 h(短光周期,S),14 h/10 h(中光周期,M),18 h/6 h(长光周期,L)],并照射不同红光/远红光比(R : FR)的光照[分别为0.83 ± 0.04(低光质比,D)、1.37 ± 0.04(正常光质比,N)、4.69 ± 0.04(高光质比,H)],不同光质比(R : FR)的值是基于Ballaré[30]等和Smith [31]等研究确定。每个处理每个种(源)随机分配4株幼苗,3次重复。通过智能定时开关(GND-1,Bull,China)来控制不同照光时长。不同的光质比由220 W的定制LED补光灯(APO,深圳市联邦重科电子科技有限公司)来实现,每种补光灯由不同比例的白光灯珠、红光灯珠、远红光灯珠组合而成。白光灯珠为全光谱,红光(R)灯珠波长为660 nm,远红光(FR)灯珠波长为730 nm。用照度计(LI-250A,LI‐COR,USA)测得低光质比(D)和正常光质比(N)的PAR均为513 ± 3 μmol·m−2·s−1(在植株高度);高光质比(H)下,PAR为495 ± 3 μmol·m−2·s−1。在高光质比(H)下,由于补光灯光强略低,灯的位置略微降低以达到相同PAR。不同处理之间用四面包围的遮光布隔开,顶部设置补光灯,留有通风口,底部为育苗容器,容器放置在透气的铁丝网苗床上。
-
利用游标卡尺(0.01 mm)和直尺(0.1 cm)对每株幼苗进行2次基径(cm)和株高(cm)的测量,每处理测3个重复,两次测量间隔15天。进而计算相对生长速率(RGR),计算公式如下[32]:
$ {RGR}_{{d}^{2}h}=\dfrac{\mathit{ln}\left({d}_{2}^{2}{h}_{2}\right)-\mathit{ln}\left({d}_{1}^{2}{h}_{1}\right)}{\left({t}_{2}-{t}_{1}\right)} $
其中,d1、h1、t1分别为第1次测量时幼苗的基径、株高和测量时间,d2、h2、t2分别为第2次测量时幼苗的基径、株高和测量时间,t2-t1为2次测量的时间间隔(月)。
-
选择长势良好的幼苗暗适应20 min后,用FluorPen-FP100手持式荧光仪(Photon System Instruments,Drasov,Czech Republic)测定叶片的OJIP曲线,OJIP曲线由3000 μmol·m−2·s−1的脉冲红光诱导,每处理测3个重复。按照每次重复OJIP曲线各时间点的荧光强度求其平均值,按照平均值绘制OJIP曲线,曲线横坐标以对数形式表示。其中20 μs测量的荧光强度为O相荧光强度(Fo),2 ms测量的荧光强度J相荧光强度(Fj),30 ms测量的荧光强度I相荧光强度(Fi),最后达到最大荧光强度P相(Fm)。通过JIP-test分析并计算得出叶片的PSII最大光化学效率(Fv/Fm)、光合性能指数(PIABS)、单位反应中心活性(ABS/RC)等参数[33]。
-
所有数据用RStudio软件(Version 1.3.1093,
http://www.rstudio.com )进行三因素(种源、光周期和光质)方差分析、多重比较,以及显著性检验(α = 0.05),用RStudio及OriginPro 2019b制图。 -
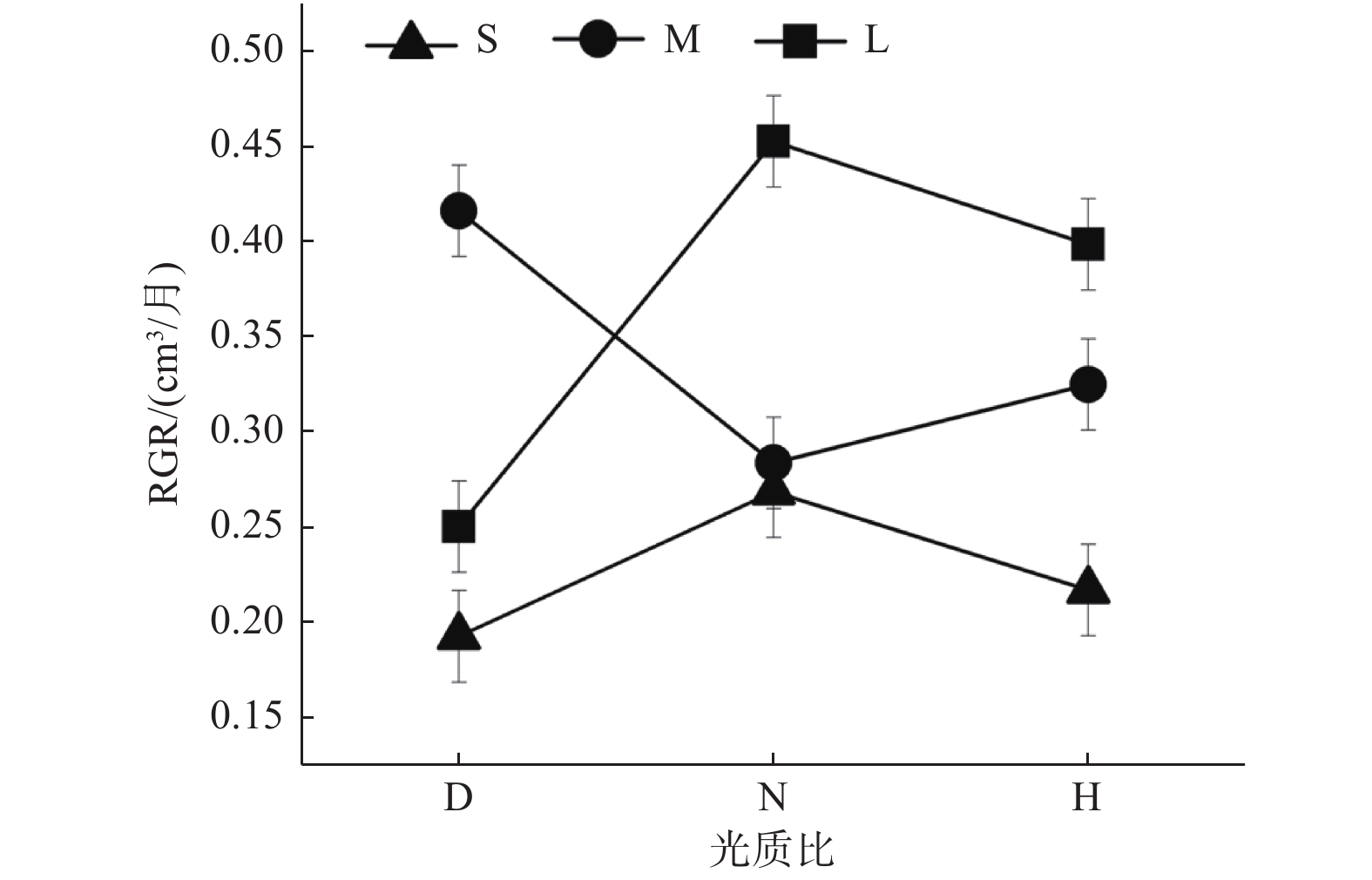
栓皮栎的不同种源间(指栓皮栎种源辽宁大连LN、陕西太白SX、云南安宁YN之间,以下简称为“种源间”)的RGR有显著差异(p < 0.05)(表3)。光周期对RGR有显著影响,短光周期S下的RGR分别比中光周期M、长光周期L低8.11%、37.84%(p < 0.05)(图1)。3个种源的RGR均随着光周期延长而呈现增加的趋势,种源LN的RGR则在中光周期M下最高。光质比处理对种源间RGR无显著影响(p > 0.05)。
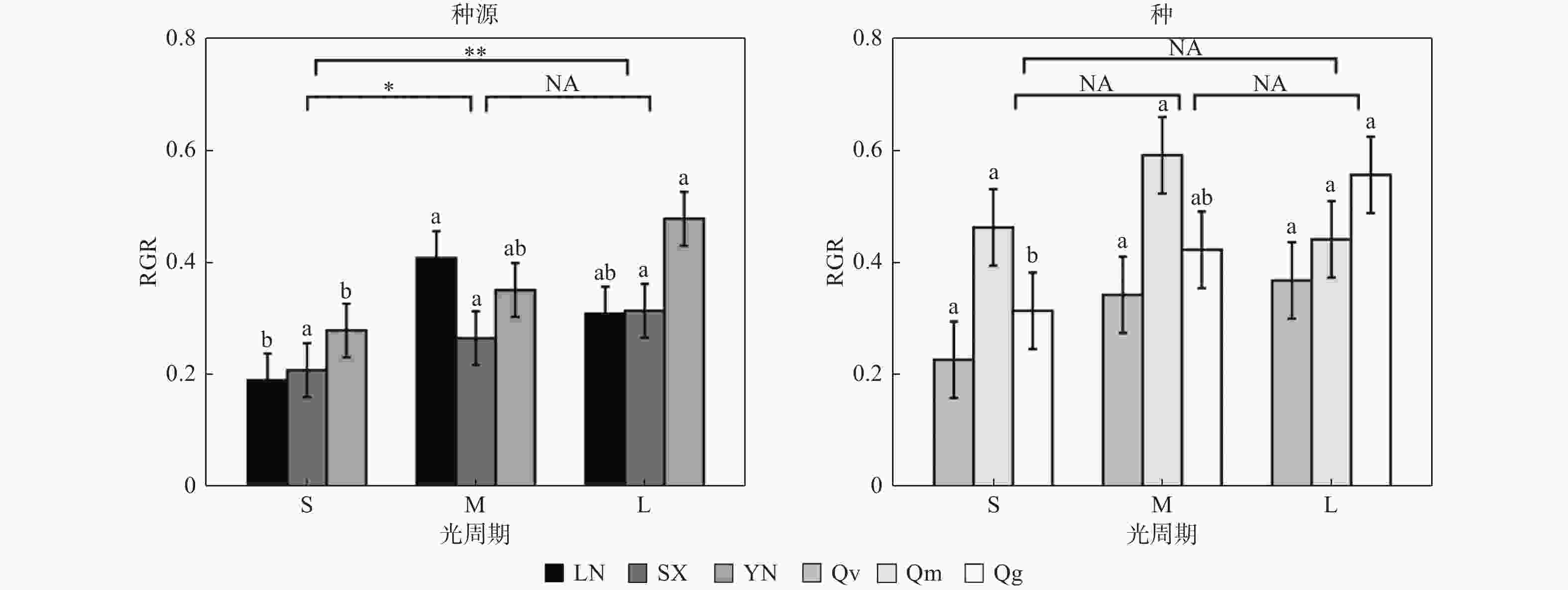
图 1 相对生长速率(RGR)对不同处理的响应
Figure 1. Response of relative growth rate (RGR) to different treatments
表 2 各栎属种的种源地地理位置与气候概况
Table 2. Geographic location and climate characteristics of different provenances
种源
Provenance种
Species纬度N
Latitude经度E
Longitude海拔
Altitude/
m年均温
Annual
mean
temperature/
℃最热月最高温
Maximum temperature
of the warmest
month/
℃最冷月最低温
Minimum temperature
of the coldest
month/
℃年降水量
Annual
precipitation/
mm年均日照时数
Annual
meansunshine
hours/
h辽宁大连 LN 栓皮栎 39°07′ 121°47′ 77.2 10.7 27.2 −8.2 619.4 2095.6 陕西太白 SX 栓皮栎 34°06′ 107°41′ 1774.3 8.2 24.0 −7.5 815.9 1379.4 云南安宁 YN 栓皮栎 24°33′ 102°17′ 1766.9 15.6 24.7 2.5 1011.0 1694.0 黑龙江加格达奇 HLJ 蒙古栎 50°14′ 124°04′ 390.3 −1.8 25.2 −33.6 492.2 2289.1 河南内乡 HN 短柄枹栎 33°02′ 111.31′′ 861.0 14.8 31.7 −3.8 813.7 1468.6 表 3 光周期、光质比及不同种源/种对RGR与基础荧光参数的三因素方差分析
Table 3. Two-way ANOVA on RGR and basic fluorescence parameters by photoperiod,light quality ratio and different provenances/species(F value)
类别
Type处理
Treatmentdf RGR 基础荧光参数
Basic fluorescence parametersFo Fj Fi Fm Vj Vi Fv/Fo Mo IP PP 2 7.36** 5.5** 8.98*** 7.45** 5.5** 4.08* 5.36** 1.59 4.93* LQ 2 0.78 4.86* 7.52** 13.54*** 14.57*** 0.58 0.06 3.44* 0.01 IP 2 3.82* 24.8*** 16.37*** 10.09*** 10.8*** 4.37* 0.51 13.82*** 12.38*** PP : LQ 4 3.32* 0.65 0.67 1.67 2.08 0.76 1.57 3.87** 0.47 PP : IP 4 1.72 1.16 0.37 0.5 0.38 0.27 2.32 1.45 1.41 LQ : IP 4 1.28 0.84 0.65 0.34 0.39 0.73 0.02 0.77 0.94 PP : LQ : IP 8 1.02 0.89 0.68 1.07 1.09 0.42 0.41 0.68 0.46 IS PP 2 3.1 7.16** 7.47** 6.29** 4.87* 4.18* 6.87** 10.31*** 4.3* LQ 2 0.95 1.62 4.07* 8.02*** 9.25*** 1.29 0.22 0.43 1.52 IS 2 5.8** 16.13*** 17.47*** 18.75*** 17.23*** 6.06** 12.02*** 11.28*** 11.67*** PP : LQ 4 1.26 0.35 1.02 2.13 2.31 0.53 3.05* 2.24 0.58 PP : IS 4 1.39 1.24 0.91 0.51 0.45 0.84 1.29 1.48 0.62 LQ : IS 4 2.26 0.82 0.59 0.72 0.84 0.23 0.54 1 0.88 PP : LQ : IS 8 1.02 0.4 0.31 0.29 0.37 0.31 0.4 0.62 0.56 注 *P < 0.05;**P < 0.01;***P > 0.001;NA:P>0.05. 下同
Notes: *P < 0.05; **P < 0.01; ***P > 0.001; NA: P > 0.05. The same below.3个栎属植物种间(指栓皮栎Qv、蒙古栎Qm、短柄枹栎Qg之间,以下简称为“种间”;栓皮栎Qv数据为栓皮栎3个种源LN、SX、YN数据取均值)的RGR有显著差异(p < 0.05)(表3)。光周期对种间RGR无显著影响,光周期S下的RGR虽然分别比光周期M、L低26.67 %、26.67 %但差异不显著(图1)。光周期处理对栓皮栎Qv、蒙古栎Qm的RGR无显著影响,但短柄枹栎的RGR随光周期延长而升高。光质比处理对种间RGR无显著影响(p > 0.05)。
光周期与光质比的交互效应仅在栓皮栎种源间显著(表3)。在中光周期M下,低光质比D的RGR最高,且显著高于正常光质比;但短光周期S与长光周期L下,均是正常光质比N下RGR最高(图2)。
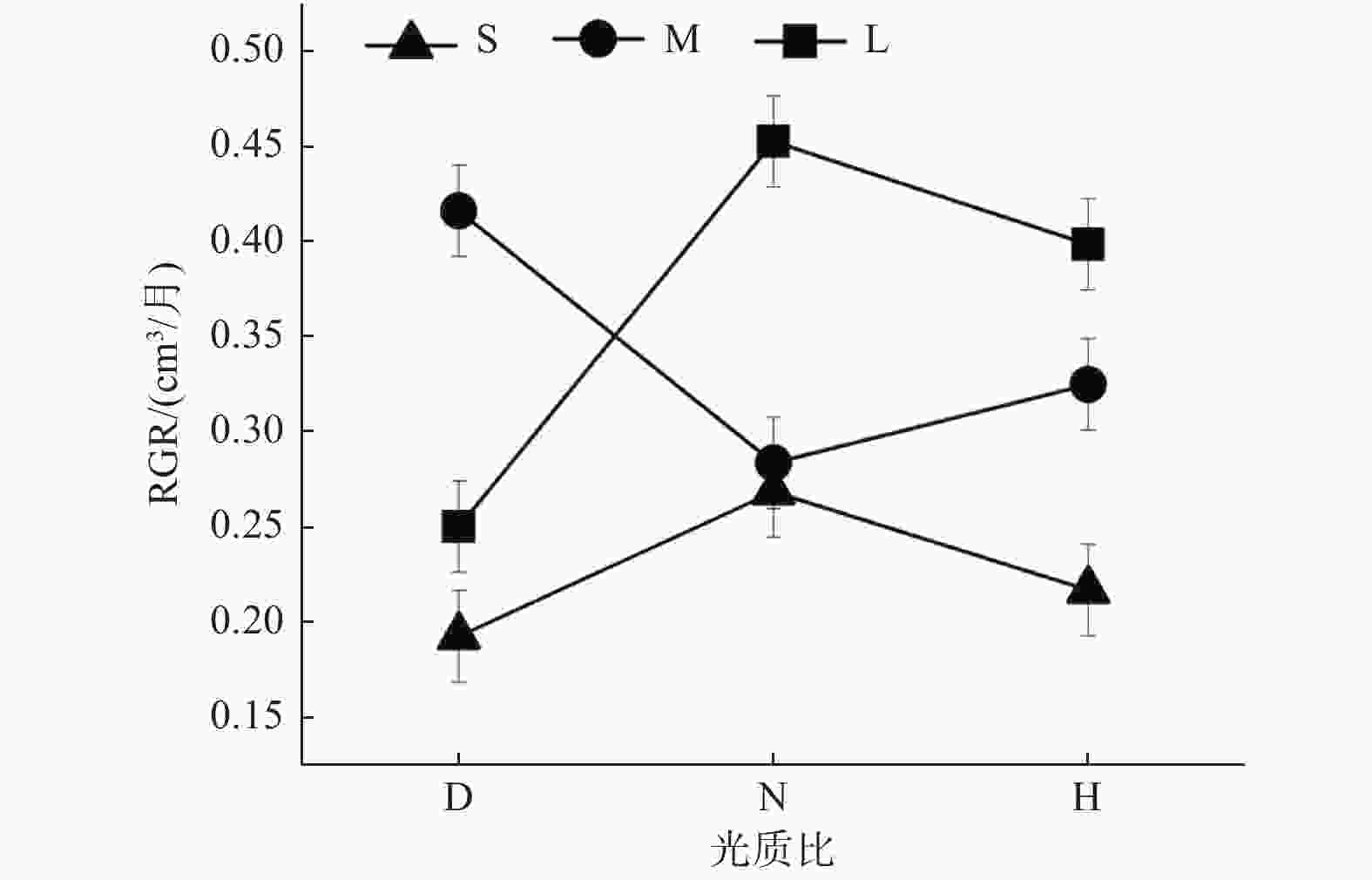
图 2 栓皮栎种源间不同光周期幼苗的RGR对光质比处理的响应
Figure 2. Response of RGR of Qv seedlings under different photoperiods to light quality ratios
-
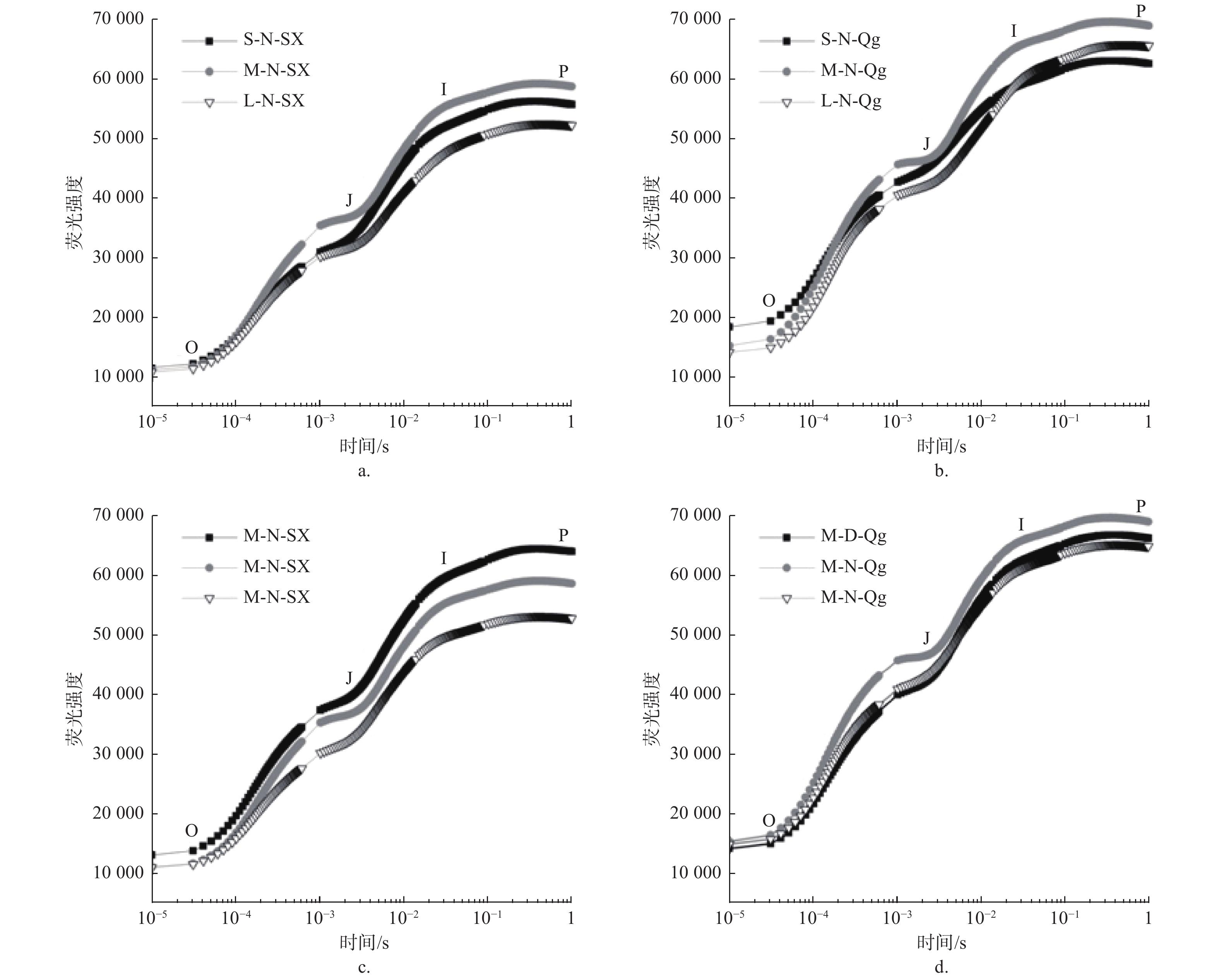
光周期、光质比以及种源对O、J、I、P4个拐点的荧光强度(Fo、Fj、Fi、Fm)均有显著或极显著影响,但3个因素的交互作用均不显著(表3)。光周期处理中,中光周期M的Fo、Fj、Fi、Fm值显著高于长光周期L (p < 0.05);光质比处理中,低光质比D下Fj、Fi、Fm均显著高于正常光质比N和高光质比H(p < 0.05)。在种间,除光质比处理对初始荧光(Fo)无显著影响外,光周期、光质比以及种对O、J、I、P4个拐点的荧光强度(Fo、Fj、Fi、Fm)均有显著或极显著影响,但3个因素的交互作用均差异不显著(表3)。由于篇幅限制,仅展示部分处理组合下(固定两个因素,对第三个因素进行比较)的OJIP曲线图(见图3)。
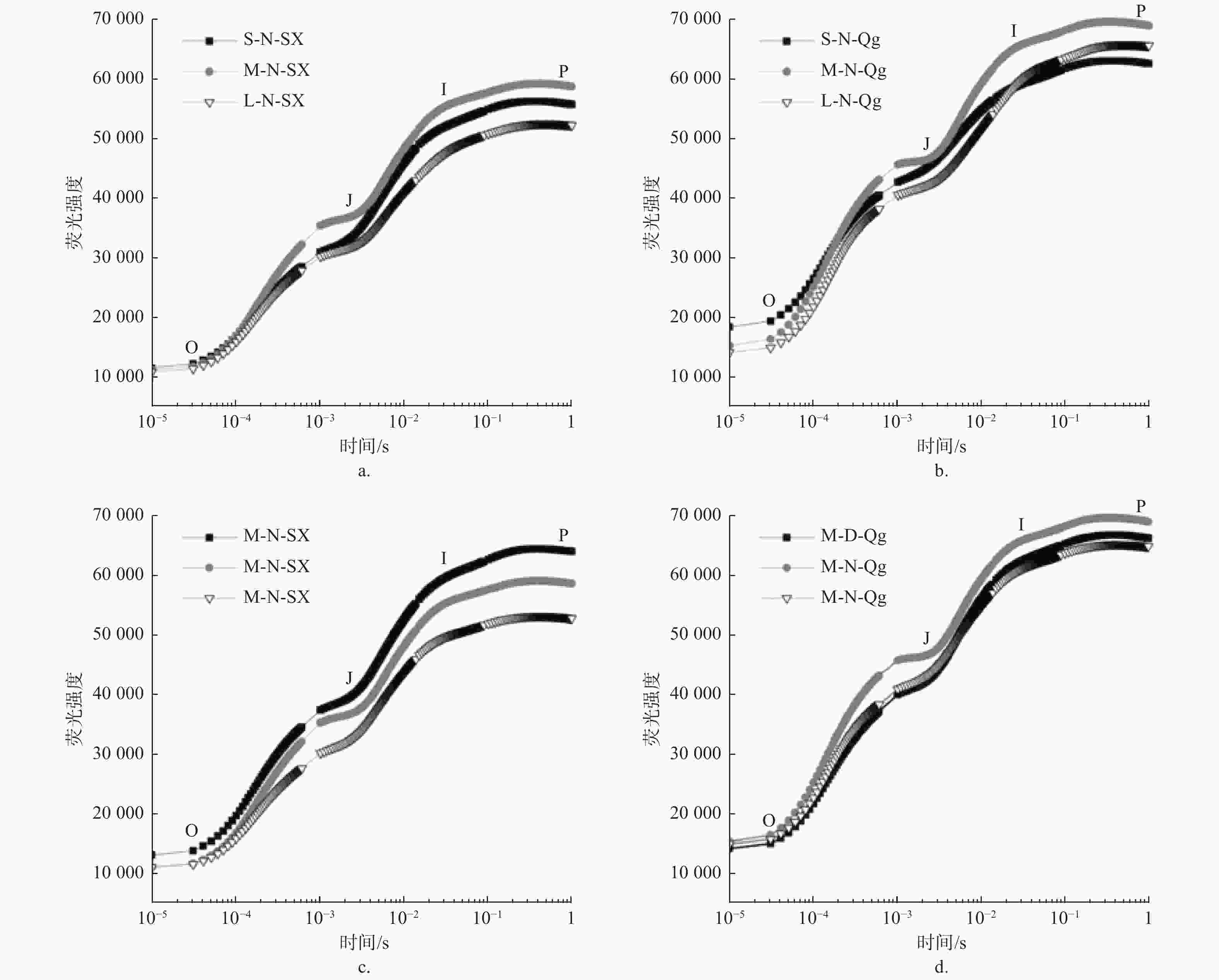
图 3 不同处理下的OJIP曲线
Figure 3. OJIP curve under different treatments
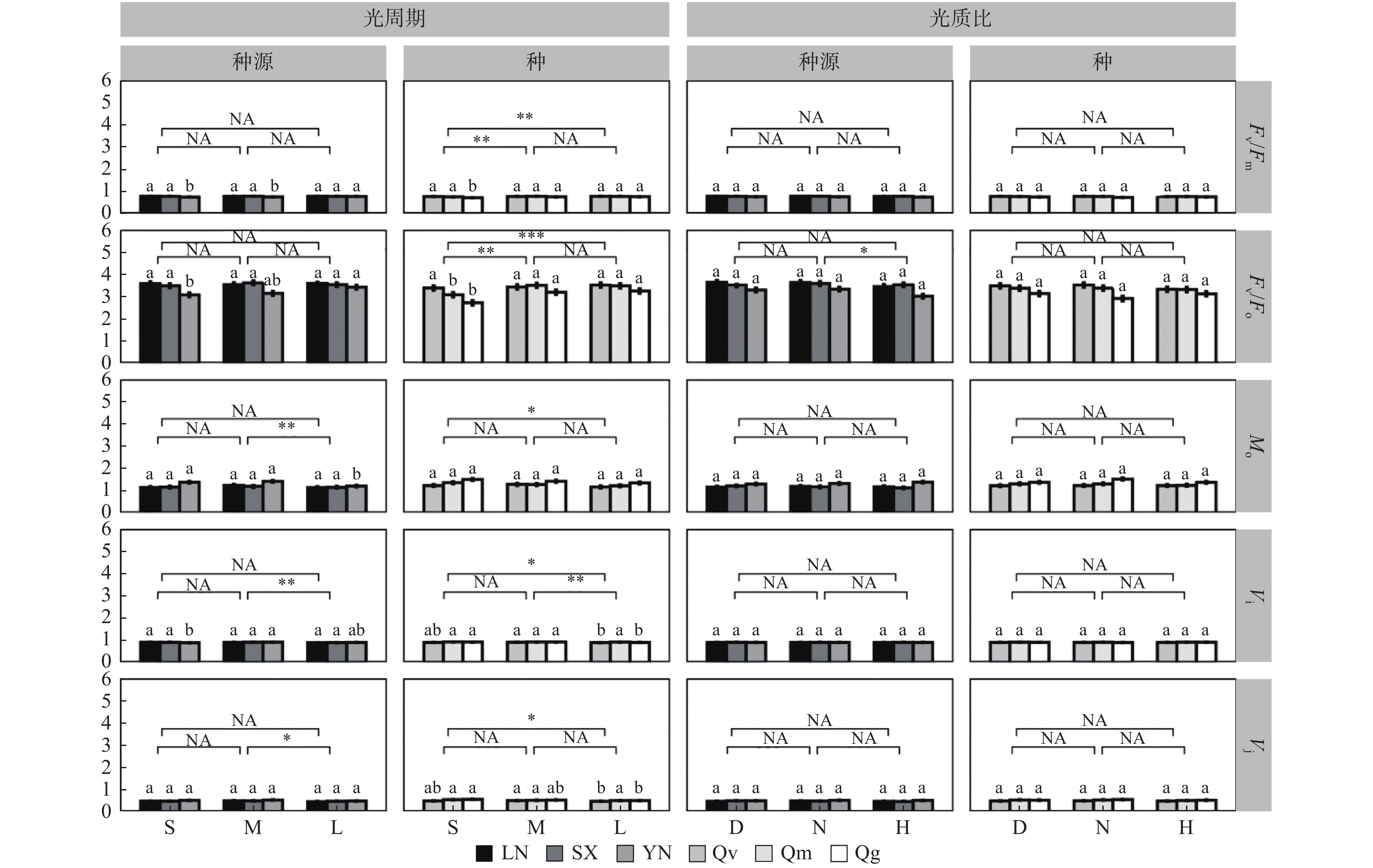
在种源水平,光周期处理主要影响Vj、Vi和Mo,如中光周期M处理下的Mo、Vi、Vj分别比长光周期L高10.34 %、2.25 %、6.25 %(p < 0.05)(图4)。光质比处理主要影响Fv/Fo值,正常光质比N处理下的Fv/Fo比高光质比H高5.41 %(N:3.51;H:3.48)。
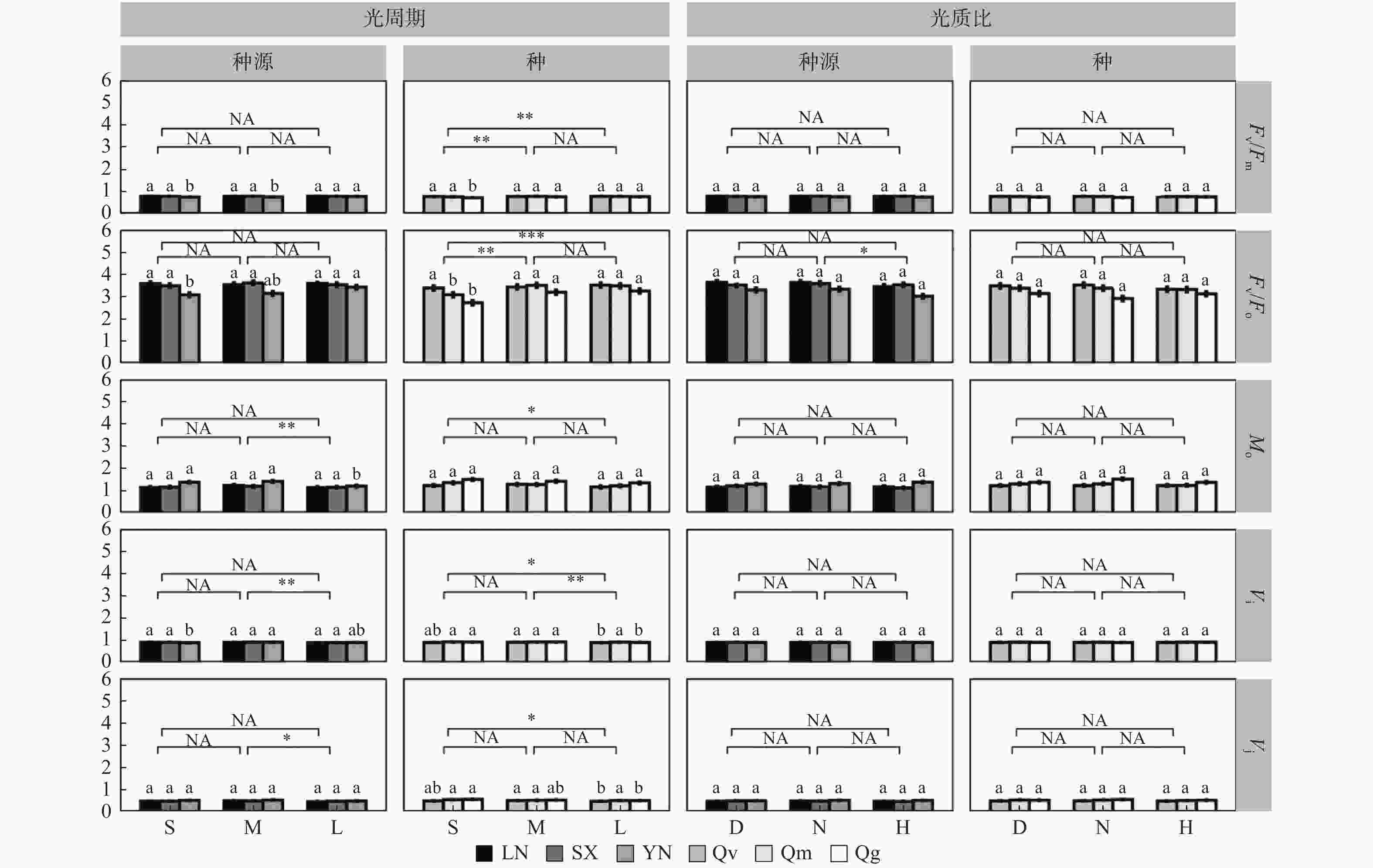
图 4 部分基础荧光参数对不同处理的响应
Figure 4. Response of some basic fluorescence parameters to different treatments
光周期处理在种间对基础荧光参数均有显著影响,中光周期M、长光周期L处理下的Fv/Fo显著高于短光周期S(S:3.06;M:3.37;L:3.41),短光周期S下Mo(S:1.36;L:1.24)、Vi(S:0.91;L:0.9)、Vj(S:0.53;L:0.5)均显著高于长光周期L(p < 0.05),中光周期M下的Vi也显著高于长光周期L(M:0.91;L:0.9)(图4)。光质比对Vj、Vi、Fv/Fo以及Mo作用均不显著。
在种源间(图4),南方种源YN在长光周期L下的的Fv/Fo、Fv/Fm显著高于短光周期S(p < 0.05),Mo显著低于短光周期S(p < 0.05),中光周期M下Vi最高。但种源LN、SX在不同光周期下Fv/Fo、Fv/Fm、Mo、Vi、Vj均差异不显著(p > 0.05)。各种源幼苗的Fv/Fo、Fv/Fm、Mo、Vi、Vj对光质比处理的响应均不显著。
在种间(图4),短柄枹栎的Fv/Fo、Fv/Fm在短光周期S下均显著低于光周期M、L,Vj、Vi显著高于光周期L;但栓皮栎的Fv/Fo、Fv/Fm、Mo、Vj对光周期处理响应均不显著,Vi在中光周期M下最高。蒙古栎的Fv/Fm、Mo、Vi、Vj对光周期处理响应均不显著,Fv/Fo在短光周期S下显著低于光周期M、L。3个栎属种的Fv/Fo、Fv/Fm、Mo、Vi、Vj对光质比处理的响应均不显著。
-
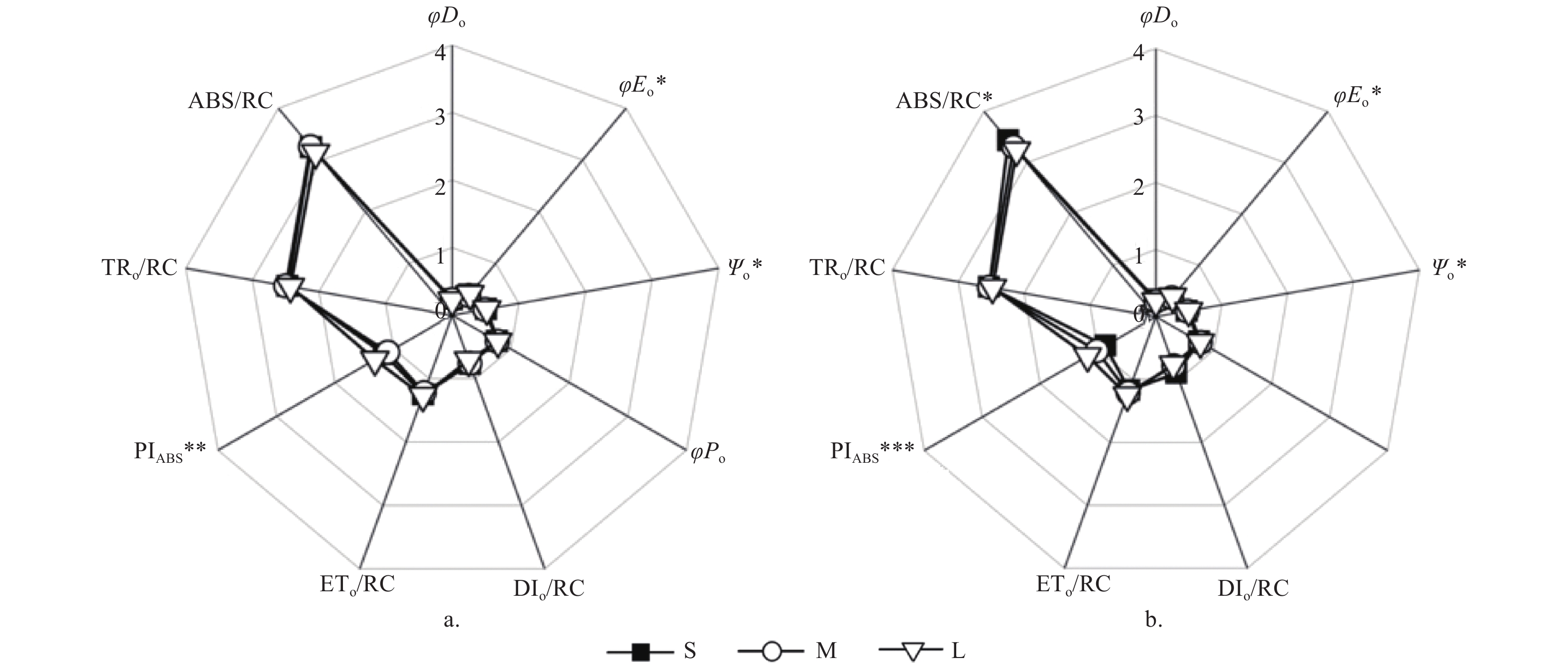
光周期处理对种源间各能量分配参数无显著影响,但对Ψo、φEo有显著影响(p < 0.05)。长光周期L的Ψo(S:0.51;M:0.49;L:0.52)、φEo(S:0.39;M:0.38;L:0.41)显著高于中光周期M(p < 0.05)(图5)。光质比处理对种源间各能量分配参数及量子产额参数均无显著影响。
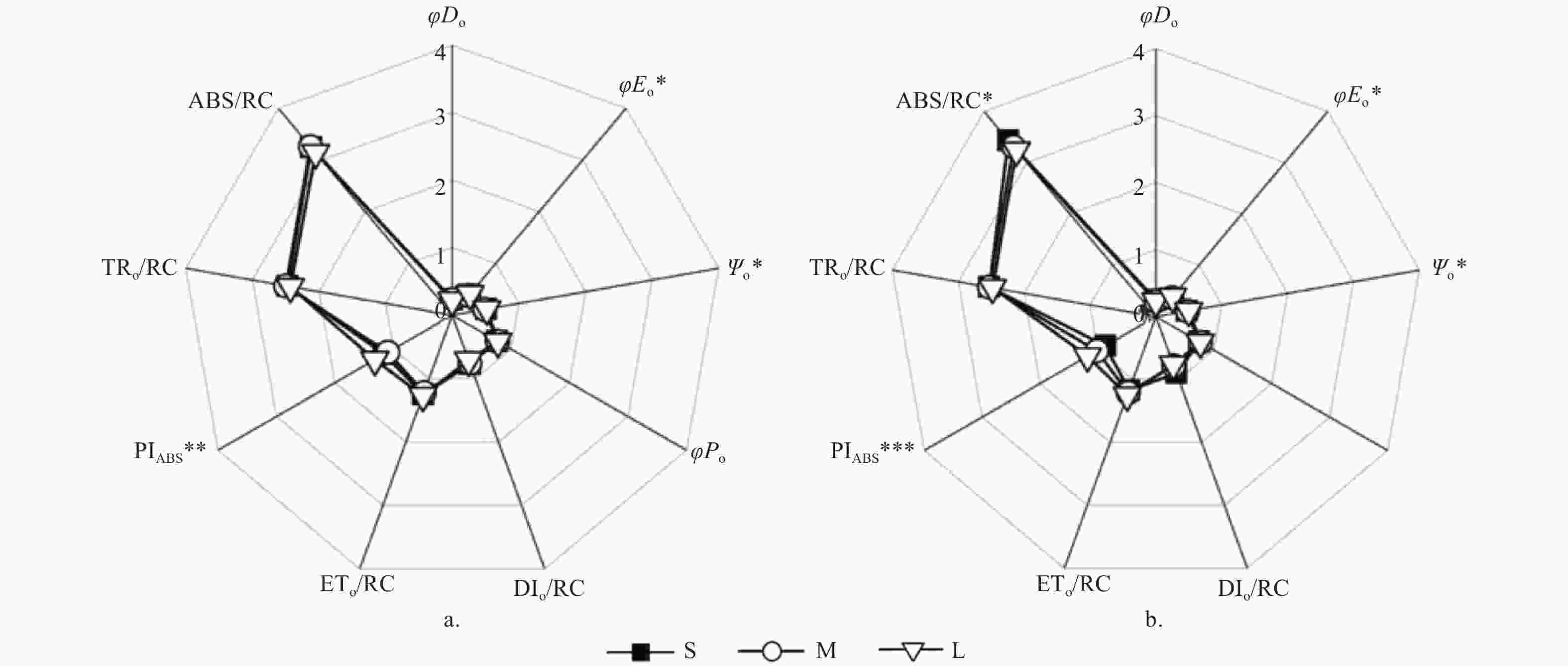
图 5 不同光周期处理下PSII反应中心活性和电子传递的变化
Figure 5. Changes of PSII reaction center activity and electron transfer under different photoperiod
除ETo/RC以及TRo/RC外,光周期对种间各能量分配参数及量子产额参数均有显著影响(p < 0.05)。短光周期S下ABS/RC(S:3.43;M:3.31;L:3.24)、DIo/RC(S:0.9;M:0.78;L:0.76)显著高于长光周期L(p < 0.05),虽也高于中光周期M但差异不显著。长光周期L的Ψo(S:0.47;M:0.48;L:0.5)、φEo(S:0.35;M:0.37;L:0.39)显著高于短光周期S(p < 0.05)。中光周期M、长光周期L的φDo显著低于短光周期S(S:0.25;M:0.23;L:0.23)(p < 0.05)。此外,长光周期L的PIAbs显著高于短光周期S和中光周期M(S:0.87;M:1.02;L:1.18)。光质比处理对种间的各能量分配参数及量子产额参数均无显著影响。
-
光周期作为环境信号因子激活植物体内的不同信号途径的发生,同时通过影响叶片光合作用制造同化物的时长来影响同化物合成和分配[34-35]。本研究结果表明,延长光周期可提高栎属植物的相对生长速率RGR,这与前人关于光周期[36-37]的研究结果一致。如朱开元等[37]的研究也表明,延长光周期显著增加了罗汉松(Podocarpus macrophylus(Thunb.)D.Don)和鸡爪槭(Acer palmatumThunb.)的株高,表明光合作用制造的同化物主要用在了植物茎的高生长上;缩短光周期会导致干物质积累不足,导致幼苗高生长缓慢。光质比处理对栎属植物幼苗RGR均无显著影响。但在中光周期下,低光质比的幼苗RGR最高,且显著高于正常光质比,表明避阴反应(即低光质比会促进茎伸长的现象)并非在任何环境下均有效,在本实验中仅在中光周期下表现出了明显的避阴反应,缩短、延长光周期均会使这一效应不明显,进一步说明光周期对避阴反应存在介导效应。这可能是由于短光周期下干物质积累不足,无法供应茎伸长所需养分,而长光周期下茎生长速度均已达到较高水平,无需光质比信号的诱导就有充足的养分供应高生长。郑芬等[4]通过监测天然林林下光照及幼树株高等指标,也发现R/FR变化对林下幼树的高径比无显著影响,认为复杂的天然林生境下植株高生长会更多地受到其他因素(如凋落物和种子特性等)的影响[38]。因此,虽然许多研究[1,39-42]表明植物存在避阴反应,但不同植物对其敏感度不同,同种植物在不同环境条件下(如光周期、凋落物等)也会对避阴反应有不同的响应。
荧光的快速上升反映了光合电子传递链中几种不同氧化还原反应的动力学所影响的
${\rm{Q}}_{\rm{A}}^ - $ 的浓度。因此,OJIP瞬态可用于表征PSII光化学的量子产率和电子传递活性[43]。本研究中,延长光周期会导致栎属植物幼苗的Fo、Fj、Fi、Fm值的降低,表明延长光周期会影响光合机构的功能,比如增加反应中心复合体的数量、增加QA到QB的电子传递速率等。φPo(Fv/Fm)和PIABS是研究植物光合生理状态的重要指标,Fv/Fm反映了植物叶片PSII原初光能转化效率[43],PIABS可从光能吸收、捕获和电子传递3方面综合反映光系统II的活性[44]。本研究中,除缩短光周期会显著降低栎属植物种水平的Fv/Fm外,其他光周期处理下Fv/Fm均变化不明显,表明缩短光周期会阻碍叶片转化能力。延长光周期会提高PIABS,表明延长光周期会增强PSII反应中心活性。Mo、Vj、ψo、φEo等参数可以反映出叶片PSII供体侧变化[45]。例如,Vj可以反映QA到QB的积累量[46-48]。本研究发现,延长光周期处理会使得Vj和Mo明显降低,说明QA被还原的速率下降,${\rm{Q}}_{\rm{A}}^ - $ 积累量减少甚至无积累。延长光周期会促进PSIIQA向QB的电子传递更通畅。缩短光周期导致φDo增加,说明缩短光周期会增加热耗散比例,降低PSII电子传递速率。与此同时,延长光周期造成ψo、φEo增加,这进一步促进了PSII中的电子传递。在种水平上的ABS/RC、DIo/RC随着光周期的延长逐渐降低,但光周期处理对TRo/RC、ETo/RC均无显著影响,表明随着光周期的延长,单位反应中心吸收的光能逐渐降低,但光合系统会通过降低热耗散的方式,优先保障传递到QA的能量以及QA传递到QB的能量,来维持光合作用的能力。李冬梅等[49]和刘杰等[50]的研究也表明,缩短光周期会降低φPo、ψo、PIABS,这与本研究发现的变化规律一致。综上,叶片的光合过程对光周期较为敏感,随着光周期延长,QA被还原的速率下降,${\rm{Q}}_{\rm{A}}^ -$ 积累量减少甚至无积累,PSIIQA向QB的电子传递更通畅,增加PSII电子传递速率。虽然单位反应中心吸收的光能逐渐降低,但光合系统通过降低热耗散,优先保障传递到QA的能量以及QA传递到QB的能量,来维持光合作用的能力。有研究认为在植物对干旱等外界环境变化不敏感时,Fo和Fm可能发生相同趋势的变化,进而维持Fv/Fm值不变[51],这与本研究的结果一致。降低光质R/FR比值会导致不同种源栓皮栎幼苗的Fo、Fj、Fi、Fm值的升高,Fv/Fm则无显著差异。因此可以认为在种源水平上叶片对光质比变化不敏感。光质比处理对栎属植物幼苗的能量分配参数及量子产额参数均无显著影响,表明光质比处理并未显著影响光合作用的结构。总体上,本研究中南部分布的种(短柄枹栎和栓皮栎)或种源(如栓皮栎南方种源YN)相比北方分布的种(如蒙古栎)或种源对光周期处理更敏感,延长光周期会显著提高南方种或种源的Fv/Fo、Fv/Fm,增加PSII光能转换效率和潜在活性,促进光系统线性电子传递,进而提高光合同化效率。这与本课题组前期的研究结果相吻合[52],即当栓皮栎南方种源(适应于当地短日照环境)移植到北方环境(长日照)后,其羧化效率、光化学猝灭系数和相对生长速率均显著高于南方环境。
-
本研究通过对栎属植物幼苗生长及荧光参数对光周期和光质比的响应分析,发现其对光周期和光质比存在种内和种间差别。栓皮栎南方种源对光周期变化响应较之中部与北部种源更加敏感,延长光周期会显著增加南方种源的Fv/Fo、Fv/Fm及RGR。在种间水平上,短柄枹栎和栓皮栎较之蒙古栎对光周期的响应更强烈,缩短光周期会显著降低短柄枹栎的Fv/Fm及RGR,延长光周期显著提高了栓皮栎和短柄枹栎的RGR,但蒙古栎的Fv/Fm及RGR对光周期的响应均不显著。R/FR比值降低会导致栎属植物幼苗的Fo、Fj、Fi、Fm值同步升高,但Fv/Fm不变,对各能量分配参数、量子产额参数及RGR也无显著影响。栎属植物幼苗总体对光质比处理不敏感。此外,光质比与光周期对RGR存在显著交互效应。在中光周期下,幼苗表现出明显的避阴反应,即低光质比下RGR最高,且显著高于正常光质比和高光质比。
光周期与光质对栎属幼苗生长及叶绿素荧光的影响
Effects of Photoperiod and Light Quality on Seedling Growth and Chlorophyll Fluorescence Kinetics of Quercus L.
-
摘要:
目的 通过探讨光周期与光质对栎属(Quercus L.)植物幼苗生长及对幼苗叶片光化学反应的影响机制,为栎属植物育苗壮苗及人工辅助更新提供科学参考。 方法 以栎属植物幼苗(3栓皮栎种源 + 1蒙古栎 + 1短柄枹栎)为研究对象,通过调控人工LED光源的方式,研究3种光周期(日夜时长分别为10 h/14 h,14 h/10 h,18 h/6 h)、3种红光/远红光比(R : FR分别为0.83 ± 0.04、1.37 ± 0.04、4.69 ± 0.04)对栎属植物幼苗的相对生长速率及叶绿素荧光参数的影响。 结果 (1)随着光周期延长,总体上栎属植物幼苗叶片的Fv/Fo、Fv/Fm、Ψo、φEo、PIAbs增加,Mo、Vi、Vj、ABS/RC、DIo/RC、φDo降低,其中栓皮栎幼苗在中光周期下Mo、Vi、Vj最高,Ψo、φEo最低;在种间水平上,短柄枹栎和栓皮栎较之蒙古栎对光周期的响应更强烈,缩短光周期会显著降低短柄枹栎的Fv/Fm及RGR,延长光周期显著提高了栓皮栎和短柄枹栎的RGR,但蒙古栎的Fv/Fm及RGR对光周期的响应均不显著。在种源水平,延长光周期显著提高了栓皮栎南方种源(YN)的Fv/Fo和Fv/Fm,且栓皮栎3个种源的RGR均随着光周期延长呈增加趋势。(2)栎属植物幼苗总体对光质比处理不敏感。光质比对3个栎属种间和栓皮栎不同种源间的RGR以及能量分配参数、量子产额参数影响不显著。(3)光质比与光周期对RGR存在显著交互效应。在中光周期下,幼苗表现出明显的避阴反应,即低光质比下RGR最高,且显著高于正常光质比和高光质比。 结论 幼苗对光周期和光质比响应存在种内和种间差别:栓皮栎南方种源对光周期变化响应更加敏感,短柄枹栎和栓皮栎较之蒙古栎对光周期的响应更强烈;栎属植物幼苗总体对光质比处理不敏感,幼苗只在特定光周期下有避阴反应。 Abstract:Objective To study the growth and photochemical mechanism of leaves in Quercus L. seedlings responding to photoperiod and light quality in order to provide reference for seedling cultivation and artificial assisted regeneration of Quercus L. Method The growth parameter and chlorophyll fluorescence parameters of Quercus spp. seedlings (3 Q. variabilis provenances + 1 Q. mongolica + 1 Q. glandulifera) under three photoperiods (the lengths of light and dark were 10 h/14 h, 14 h/10 h, 18 h/6 h, respectively) and three red - far red light ratios (R : FR = 0.83 ± 0.04, 1.37 ± 0.04, 4.69 ± 0.04, respectively) were investigated. Result (1) At the species' level, the Fv/Fo, Fv/Fm, Ψo, φEo and PIAbs increased, while Mo, Vi, Vj, ABS/RC, DIo/RC and φDo decreased with the prolonging of photoperiod. Specially, Mo, Vi and Vj of Q. variabilis seedlings were the highest, Ψo and φEo were the lowest under medium photoperiod. Q. glandulifera and Q. variabilis showed a stronger response to photoperiod than Q. mongolica. The shortening photoperiod would significantly reduce the Fv/Fm and relative growth rate (RGR) of Q. glandulifera, and the prolonging photoperiod would significantly improve the RGR of Q. glandulifera and Q. variabilis, but the Fv/Fm and RGR of Q. mongolica showed no significant response to photoperiod changes. At the intra-species level, long photoperiod significantly increased the Fv/Fo and Fv/Fm of the Yunnan provenance and the RGR of all the three provenances of Q. variabilis increased with photoperiod. (2) Light quality ratio had no significant effects on RGR, energy allocation parameters and quantum yield parameters among three Quercus species and different provenances of Q. variabilis. (3) The significantly interactive effects between light quality ratio and photoperiod on RGR were observed. Under medium photoperiod, the seedlings showed an obvious shade-avoidance response, namely, the RGR of seedlings with low light quality ratio was the highest, which was significantly higher than that in the normal and high light quality ratio. Conclusion The results show that there are intraspecific and interspecific differences in response to photoperiod and light quality ratio. The south provenance of Q. variabilis are more sensitive to photoperiod. Q. glandulifera and Q. variabilis have a stronger response to photoperiod than Q. mongolica. Light quality ratio generally has no significant effects among Quercus L. seedlings. The seedlings show obvious shade-avoidance responses only under specific photoperiod. -
Key words:
- photoperiod
- / light quality ratio
- / Quercus L.
- / chlorophyll fluorescence
- / photochemical efficiency
-
图 1 相对生长速率(RGR)对不同处理的响应
Figure 1. Response of relative growth rate (RGR) to different treatments
图 2 栓皮栎种源间不同光周期幼苗的RGR对光质比处理的响应
Figure 2. Response of RGR of Qv seedlings under different photoperiods to light quality ratios
图 4 部分基础荧光参数对不同处理的响应
Figure 4. Response of some basic fluorescence parameters to different treatments
图 5 不同光周期处理下PSII反应中心活性和电子传递的变化
Figure 5. Changes of PSII reaction center activity and electron transfer under different photoperiod
表 1 主要缩略词中英对照
Table 1. List of main abbreviations
缩略词
Abbreviations英文全称/含义
English name/meaning中文全称/含义
Chinese name/meaningPAR Photosynthetically active radiation 光合有效辐射 R:FR Red / far red light ratios 红光/远红光的光质比 SAS Shade avoidance syndrome 避阴反应 PP Photoperiod 光周期 LQ Light quality ratio 光质比 IS Interspecies 种间 IP Inter provenances 种源间 Qv Q. Variabilis 栓皮栎 Qm Q.Mongolica 蒙古栎 Qg Q. Glandulifera 短柄枹栎 LN Liaoning 辽宁大连,北方栓皮栎种源 SX Shaanxi 陕西太白,中部栓皮栎种源 YN Yunnan 云南安宁,南方栓皮栎种源 S Short photoperiod 短光周期,日夜时长分别为10 h/14 h M Medium photoperiod 中光周期,日夜时长分别为14 h/10 h L Long photoperiod 长光周期,日夜时长分别为8 h/6 h D Low light quality ratio 低光质比,R/FR为0.83 ± 0.04 N Normal light quality ratio 正常光质比(即日光下光质比),R/FR为1.37 ± 0.04 H High light quality ratio 高光质比,R/FR为4.69 ± 0.04 RGR Relative growth rate 相对生长速率 Fo、Fj、Fi、Fm / O、J、I、P四个拐点的荧光强度 Vj Relative variable fluorescence at the J-step J 相的相对可变荧光 Vi Relative variable fluorescence at 30 ms I 相的相对可变荧光 Fv/Fo Conformation term for primary photochemistry PS II潜在活性 Mo Approximated initial slope (in m·s−1) of the fluorescence
transient normalized on the maximal variable fluorescence FvOJIP 荧光诱导曲线的初始斜率 Ψo Openness of active reaction center 有活性反应中心的开放程度。 φPo Maximum photochemical efficiency 最大光化学效率 φEo Quantum yield 电子传递链的量子产额 φDo Maximum quantum yield for non-photochemical quenching 非光化学淬灭最大量子产额 ABS/RC Light energy absorbed by unit reaction center; 单位反应中心吸收的光能 TRo/RC Light energy captured by unit reaction center; 单位反应中心捕获的光能 ETo/RC Unit reaction center for electron transfer energy 单位反应中心用于电子传递的能量 DIo/RC The energy dissipated per unit reaction center 单位反应中心耗散掉的能量 PIAbs Photosynthetic performance index 以吸收光能为基础的光合性能指数  下载: 导出CSV
下载: 导出CSV
表 2 各栎属种的种源地地理位置与气候概况
Table 2. Geographic location and climate characteristics of different provenances
种源
Provenance种
Species纬度N
Latitude经度E
Longitude海拔
Altitude/
m年均温
Annual
mean
temperature/
℃最热月最高温
Maximum temperature
of the warmest
month/
℃最冷月最低温
Minimum temperature
of the coldest
month/
℃年降水量
Annual
precipitation/
mm年均日照时数
Annual
meansunshine
hours/
h辽宁大连 LN 栓皮栎 39°07′ 121°47′ 77.2 10.7 27.2 −8.2 619.4 2095.6 陕西太白 SX 栓皮栎 34°06′ 107°41′ 1774.3 8.2 24.0 −7.5 815.9 1379.4 云南安宁 YN 栓皮栎 24°33′ 102°17′ 1766.9 15.6 24.7 2.5 1011.0 1694.0 黑龙江加格达奇 HLJ 蒙古栎 50°14′ 124°04′ 390.3 −1.8 25.2 −33.6 492.2 2289.1 河南内乡 HN 短柄枹栎 33°02′ 111.31′′ 861.0 14.8 31.7 −3.8 813.7 1468.6
下载: 导出CSV
表 3 光周期、光质比及不同种源/种对RGR与基础荧光参数的三因素方差分析
Table 3. Two-way ANOVA on RGR and basic fluorescence parameters by photoperiod,light quality ratio and different provenances/species(F value)
类别
Type处理
Treatmentdf RGR 基础荧光参数
Basic fluorescence parametersFo Fj Fi Fm Vj Vi Fv/Fo Mo IP PP 2 7.36** 5.5** 8.98*** 7.45** 5.5** 4.08* 5.36** 1.59 4.93* LQ 2 0.78 4.86* 7.52** 13.54*** 14.57*** 0.58 0.06 3.44* 0.01 IP 2 3.82* 24.8*** 16.37*** 10.09*** 10.8*** 4.37* 0.51 13.82*** 12.38*** PP : LQ 4 3.32* 0.65 0.67 1.67 2.08 0.76 1.57 3.87** 0.47 PP : IP 4 1.72 1.16 0.37 0.5 0.38 0.27 2.32 1.45 1.41 LQ : IP 4 1.28 0.84 0.65 0.34 0.39 0.73 0.02 0.77 0.94 PP : LQ : IP 8 1.02 0.89 0.68 1.07 1.09 0.42 0.41 0.68 0.46 IS PP 2 3.1 7.16** 7.47** 6.29** 4.87* 4.18* 6.87** 10.31*** 4.3* LQ 2 0.95 1.62 4.07* 8.02*** 9.25*** 1.29 0.22 0.43 1.52 IS 2 5.8** 16.13*** 17.47*** 18.75*** 17.23*** 6.06** 12.02*** 11.28*** 11.67*** PP : LQ 4 1.26 0.35 1.02 2.13 2.31 0.53 3.05* 2.24 0.58 PP : IS 4 1.39 1.24 0.91 0.51 0.45 0.84 1.29 1.48 0.62 LQ : IS 4 2.26 0.82 0.59 0.72 0.84 0.23 0.54 1 0.88 PP : LQ : IS 8 1.02 0.4 0.31 0.29 0.37 0.31 0.4 0.62 0.56 注 *P < 0.05;**P < 0.01;***P > 0.001;NA:P>0.05. 下同
Notes: *P < 0.05; **P < 0.01; ***P > 0.001; NA: P > 0.05. The same below.
下载: 导出CSV
-
[1] Kurepin L V, Emery R J, Pharis R P, et al. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: putative roles for plant hormones in leaf and internode growth[J]. Journal of Experimental Botany, 2007, 58(8): 2145-57. doi: 10.1093/jxb/erm068 [2] Montgomery R A, Chazdon R L. Forest structure, canopy architecture, and light transmittance in tropical wet forests[J]. Global Ecology & Biogeography, 2001, 82(10): 2707-2718. [3] Mejia-Dominguez N R, Meave J A, Diaz-Avalos C. Spatial structure of the abiotic environment and its association with sapling community structure and dynamics in a cloud forest[J]. International Journal of Biometeorology, 2012, 56(2): 305-18. doi: 10.1007/s00484-011-0434-5 [4] 郑 芬, 李兆佳, 邱治军, 等. 广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响[J]. 生态学报, 2020, 40(13):4516-4527. [5] Terfa M T, Solhaug K A, Gislerod H R, et al. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa x hybrida but does not affect time to flower opening[J]. Physiologia Plantarum, 2013, 148(1): 146-59. doi: 10.1111/j.1399-3054.2012.01698.x [6] Sevillano I, Short I, Grant J, et al. Effects of light availability on morphology, growth and biomass allocation of Fagus sylvatica and Quercus robur seedlings[J]. Forest Ecology and Management, 2016, 374: 11-19. doi: 10.1016/j.foreco.2016.04.048 [7] 贺顺钦, 王发其. 辽东栎苗木早期生长与光的关系[J]. 林业科学研究, 2001, 14(6):697-700. doi: 10.3321/j.issn:1001-1498.2001.06.018 [8] Wei H, Hauer R J, Chen G, et al. Growth, nutrient assimilation, and carbohydrate metabolism in Korean pine (Pinus koraiensis) seedlings in response to light spectra[J]. Forests, 2019, 11(1). [9] Tylewicz S, Petterle A, Marttila S. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication[J]. Science, 2018, 360(6385): 212-215. doi: 10.1126/science.aan8576 [10] Navidad H, Fløistad I S, Olsen J E, et al. Subalpine fir (Abies laciocarpa) and Norway spruce (Picea abies) seedlings show different growth responses to blue light[J]. Agronomy, 2020, 10(5): 712. doi: 10.3390/agronomy10050712 [11] Bauerle W L, Oren R, Way D A, et al. Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(22): 8612-8617. doi: 10.1073/pnas.1119131109 [12] Kurepin L V, Shah S, Reid D M. Light quality regulation of endogenous levels of auxin, abscisic acid and ethylene production in petioles and leaves of wild type and ACC deaminase transgenic Brassica napus seedlings[J]. Plant Growth Regulation, 2007, 52(1): 53-60. doi: 10.1007/s10725-007-9176-0 [13] Li W, Liu S-W, Ma J-J, et al. Gibberellin signaling is required for far-red light-induced shoot elongation in Pinus tabuliformis seedlings[J]. Plant Physiology, 2020, 182(1): 658-668. doi: 10.1104/pp.19.00954 [14] Kurepin L V, Walton L J, Hayward A, et al. Interactions between plant hormones and light quality signaling in regulating the shoot growth of Arabidopsis thaliana seedlings[J]. Botany, 2012, 90(3): 237-246. doi: 10.1139/b11-108 [15] Tao Y, Ferrer J L, Ljung K, et al. Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants[J]. Cell, 2008, 133(1): 164-76. doi: 10.1016/j.cell.2008.01.049 [16] 王庆燕. 不同群体结构下玉米避阴反应的生理生化机制及其调控研究[D]. 北京: 中国农业大学, 2015. [17] Zheng L, Van Labeke M C. Long-Term effects of red- and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants[J]. Front Plant Sci, 2017, 8: 917. doi: 10.3389/fpls.2017.00917 [18] Centofante A R. Light quality on the morphoanatomy and physiology of Campomanesia pubescens (DC. ) O. Berg. seedlings[J]. Scientia Horticulturae, 2020: 259. [19] Ma X F, Hall D, Onge K R, et al. Genetic differentiation, clinal variation and phenotypic associations with growth cessation across the Populus tremula photoperiodic pathway[J]. Genetics, 2010, 186(3): 1033-44. doi: 10.1534/genetics.110.120873 [20] Franklin K A. Light and temperature signal crosstalk in plant development[J]. Curr Opin Plant Biol, 2009, 12(1): 63-68. doi: 10.1016/j.pbi.2008.09.007 [21] Chiang C, Hoch D B G. Reaching natural growth light quality effects on plant performance in indoor growth facilities[J]. Plants (Basel, Switzerland), 2020, 9(10): 1312. [22] 国家林业和草原局. 中国森林资源报告(2014-2018)[M]. 北京: 中国林业出版社, 2019. [23] 厉月桥, 李迎超, 吴志庄. 中国北方栎属植物资源调查与区划[J]. 林业资源管理, 2013(4):88-93. doi: 10.3969/j.issn.1002-6622.2013.04.017 [24] 郑金萍, 杨学东, 郭忠玲, 等. 蒙古栎林天然更新状况及影响因素研究[J]. 北华大学学报: 自然科学版, 2015, 16(5):652-658. [25] Lupke B V. Silvicultural methods of oak regeneration with special respect to shade tolerant mixed species[J]. Forest Ecology and Management, 1998, 106: 19-26. doi: 10.1016/S0378-1127(97)00235-1 [26] 姜玲玲, 许中旗. 东北东部山地蒙古栎林的年龄结构及天然更新研究[J]. 林业与生态科学, 2020, 35(4):365-376. [27] 王 巍, 李庆康, 马克平. 东灵山地区辽东栎幼苗的建立和空间分布[J]. 植物生态学报, 2000, 24(5):595-600. doi: 10.3321/j.issn:1005-264X.2000.05.014 [28] 于世川, 张建国, 叶权平, 等. 抚育间伐对黄龙山辽东栎林分质量的影响[J]. 西北林学院学报, 2018, 33(3):52-60. doi: 10.3969/j.issn.1001-7461.2018.03.09 [29] 李迎超, 厉月桥, 王利兵, 等. 木本淀粉能源植物栓皮栎与麻栎的资源调查以及分布规律[J]. 林业资源管理, 2013(2):94-101. doi: 10.3969/j.issn.1002-6622.2013.02.018 [30] Ballare, C L, Scopel, A L, Sanchez, R A. Far-red radiation reflected from adjacent leaves- an early signal of competition in plant canopies.[J]. Science, 1990, 247(4940): 329-332. doi: 10.1126/science.247.4940.329 [31] Smith H. Phytochromes and light signal perception by plants—an emerging synthesis[J]. Nature, 2000, 407(6804): 585-591. doi: 10.1038/35036500 [32] Kohyama T, Hotta M. Significance of allometry in tropical saplings.[J]. Functional Ecology, 1990, 4(4): 515-521. doi: 10.2307/2389319 [33] J. Strasser R, Alakasrivastava, Govindjee. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria[J]. Photochemistry and Photobiology, 1995, 61(1): 32-42. doi: 10.1111/j.1751-1097.1995.tb09240.x [34] Dong W, Zhang Y, Zhang Y, et al. Short-day Photoperiod effects on plant growth, flower bud differentiation, and yield formation in Adzuki Bean (Vigna angularis)[J]. International Journal of Agriculture and Biology, 2016, 18(2): 337-345. doi: 10.17957/IJAB/15.0091 [35] Hendriks J H, Kolbe A, Gibon Y, et al. ADP-glucose pyrophosphorylase is activated by posttranslational redox-modification in response to light and to sugars in leaves of Arabidopsis and other plant species[J]. Plant Physiology, 2003, 133(2): 838-49. doi: 10.1104/pp.103.024513 [36] Wei H, Ren J, Zhou J. Effect of exponential fertilization on growth and nutritional status in Buddhist pine [Podocarpus macrophyllus(Thunb. ) D. Don] seedlings cultured in natural and prolonged photoperiods[J]. Soil Science and Plant Nutrition, 2013, 59(6): 933-941. doi: 10.1080/00380768.2013.864957 [37] 朱开元, 刘慧春, 周江华, 等. 延长光周期对罗汉松和鸡爪槭苗期生长及养分吸收利用的影响[J]. 浙江大学学报: 农业与生命科学版, 2016, 42(2):190-198. [38] 郑维娜, 王孝安, 郭 华, 等. 微生境对辽东栎幼苗生长的影响[J]. 干旱区研究, 2013, 30(6):1049-1055. [39] Sessa G, Carabelli M, Possenti M, et al. Multiple pathways in the control of the shade avoidance response[J]. Plants (Basel), 2018, 7(4): 102. [40] Ammer C. Growth and biomass partitioning of Fagus sylvatica L. and Quercus robur L. seedlings in response to shading and small changes in the R/FR-ratio of radiation[J]. Annals of Forest Science, 2003, 60(2): 163-171. doi: 10.1051/forest:2003009 [41] Casal J J. Photoreceptor signaling networks in plant responses to shade[J]. Annual Review of Plant Biology, 2013, 64: 403-27. doi: 10.1146/annurev-arplant-050312-120221 [42] Hussain S, Iqbal N, Rahman T, et al. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes[J]. Environmental and Experimental Botany, 2019, 162: 374-382. doi: 10.1016/j.envexpbot.2019.03.011 [43] Stirbet A, Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: basics and applications of the OJIP fluorescence transient[J]. Journal of Photochemistry and Photobiology B-Biology, 2011, 104(1-2): 236-57. doi: 10.1016/j.jphotobiol.2010.12.010 [44] 张璐颖, 文 笑, 林勇明, 等. 盐胁迫对台湾桤木幼苗光合作用和荧光特性的影响[J]. 福建林学院学报, 2013, 33(3):193-199. doi: 10.3969/j.issn.1001-389X.2013.03.001 [45] 李鹏民. 快速叶绿素荧光诱导动力学在植物逆境生理研究中的应用[D]. 泰安: 山东农业大学, 2007. [46] 李英浩, 刘景辉, 赵宝平, 等. 干旱胁迫对燕麦生长及叶片光系统Ⅱ活性的影响[J]. 西北植物学报, 2020, 40(10):1706-1713. [47] Zhang H, Zhong H, Wang J, et al. Adaptive changes in chlorophyll content and photosynthetic features to low light in Physocarpus amurensis Maxim and Physocarpus opulifolius "Diabolo"[J]. Peer J, 2016, 4: e2125. doi: 10.7717/peerj.2125 [48] Haldimann P, Strasser R J. Effects of anaerobiosis as probed by the polyphasic chlorophyll a fluorescence rise kinetic in pea (Pisum sativum L. )[J]. Photosynthesis Research, 1999, 62(1): 67-83. doi: 10.1023/A:1006321126009 [49] 李冬梅, 谭秋平, 高东升, 等. 光周期对休眠诱导期桃树光合及PSII光系统性能的影响 J]. 应用生态学报, 2014, 25(7):1933-1939. [50] 刘 杰, 胡笑涛, 王文娥, 等. 光强和光周期对水培生菜光合及叶绿素荧光特性的影响[J]. 西南农业学报, 2019, 032(8):1784-1790. [51] 滕志远, 张会慧, 代 欣, 等. 干旱对桑树叶片光系统II活性的影响[J]. 浙江农业学报, 2016, 28(1):1-8. doi: 10.3969/j.issn.1004-1524.2016.01.01 [52] 王小菲, 高文强, 刘建锋, 等. 不同生境对栓皮栎幼苗光合生理特性的影响[J]. 生态学报, 2016, 36(24):8062-8070. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3513
- HTML全文浏览量: 1572
- PDF下载量: 66
- 被引次数: 0




