



-
榛(Hazelnut)隶属于桦木科(Betulaceae)榛属(Corylus Linn.),是世界上重要的木本油料树种,榛果亦被誉为“坚果之王”,中国先民自古以来就采集野生榛子食用[1]。世界上的榛属植物约有20种,我国是榛属植物多样性和分布中心,原产8个种和2个变种。近年来,国内关于榛属的研究主要涉及到榛种质资源研究、育种、育苗与栽培、形态发育、生理和分子生物学研究及榛仁营养、综合利用与榛产业发展现状等[2-5]。目前,在榛子育种方面已经培育出众多优良的平欧杂种榛品种(系),填补了榛子生产无栽培品种(系)的空白,但因大多数品种(系)均为灌木,基部萌蘖多,在实际生产中除萌要耗费大量人力物力,不利于机械化采摘,因此,培育乔木状单干树形品种是当前榛子产业发展的研究方向之一。
华榛(Corylus chinensis Franch.)作为我国特有的珍稀濒危树种,具有极高的经济、营养与生态价值:首先,其树体高大,树干通直,无根蘖发生,是培育无根蘖砧木及乔木状单干树形品种的优良亲本材料[6-7];其次,华榛木材纹理致密、结构细腻,质地坚韧,可制作建筑及家具;再者,华榛种子及果仁中含有多种营养物质,微量元素含量也比较丰富,果实中富含淀粉和油脂,含油率达50%[8],味美可口,营养价值丰富[9-10]。华榛喜温凉、湿润的气候环境,其自然种群间断分布于我国亚热带地区的中高山地带(海拔800~3 500 m),分布区北起秦岭淮河一线,南抵横断山区,内部山脉河流交错,环境异质化程度高。近年来,人类活动不断影响自然环境,引起环境质量的变化,严重影响物种的生存和繁衍[11-12]。曹策等[13]指出,全球的气候条件在18世纪工业革命以来发生了巨大变化,总体气候的变化趋势是全球变暖,并且预测气候的变化速率呈持续增大趋势。气候变化通过影响物种的生理生态特性从而影响物种的地理分布格局,同时也影响着物种现有栖息地的适宜度[14-16]。华榛受气候变化与人为干扰的影响,其生境被破坏、资源锐减,分布区内不仅大树罕见,残存幼株也较稀少,而且果实易被鸟兽啃食,自然更新困难,有被其它阔叶树种替代而陷入濒危绝灭的趋势。目前,华榛已被列为国家三级保护树种,同时也被世界自然保护联盟(IUCN)列入濒危物种红色名录。鉴于此,展开华榛与气候环境变化关系的探究具有必要性和紧迫性。
随着生态环境的变化,人们的生物多样性保护意识不断加强。在对生物多样性保护的研究中,生态位模型被广泛使用,不仅被用来研究入侵害虫对植物物种的潜在威胁,同时也用于预测物种分布对气候变化的响应机制[17]。生态位模型通过收集物种有限的分布点即可预测物种潜在的适宜生境,这对于仅有少量分布记录的珍稀濒危树种具有重要的实际意义[18-19]。基于MaxEnt软件的最大熵模型是目前应用最广泛的物种分布模型之一[20],该模型准确度较高,得以广泛使用[21],也被称为是预测物种地理分布最可靠的三种技术之一[22]。多数研究认为,最大熵模型与GIS相结合是目前效果最好且稳定性较强的一种预测方法[23-26]。王运生等[27]对比其他几种模型发现,用来判定模型准确性的ROC曲线下面积值中,MaxEnt模型值最大,由此看来,MaxEnt模型的准确度更高、模拟效果更好。在对黄顶菊适生区分布进行预测时,应用不同的生态位模型进行预测,对比发现,MaxEnt模型的模拟精度最好[28]。孙文涛等[29]也认为,在物种分布数据相对较少情况下,应用MaxEnt模型进行模拟的预测结果优于其他模型的预测效果。
目前,我国关于华榛的研究大多集中于人工造林、生理生化及形态分类等方面,而对华榛适生区的潜在地理分布以及华榛如何应对气候变化尚无系统报道。因此,基于华榛的现代分布数据模拟其在第四纪以来气候变化下的潜在适生区分布及地理分布变迁,可为华榛自然资源的保护提供理论依据。本研究结合华榛野外调查与文献记录,利用ArcGis空间分析技术与MaxEnt最大熵模型,选取影响华榛分布的相关环境因子建模,模拟华榛在不同时期的地理分布,对中国特有珍稀濒危树种华榛的生境进行评价,以期解决如下问题:(1)影响华榛分布的关键气候因子是什么,其地理分布和生物气候因子之间有什么关系?(2)华榛在末次间冰期、末次盛冰期、全新世中期、现代及未来2种不同情景下2050s和2070s的潜在适生区分布?(3)华榛在当前全球气候变暖背景下的生存前景如何?
-
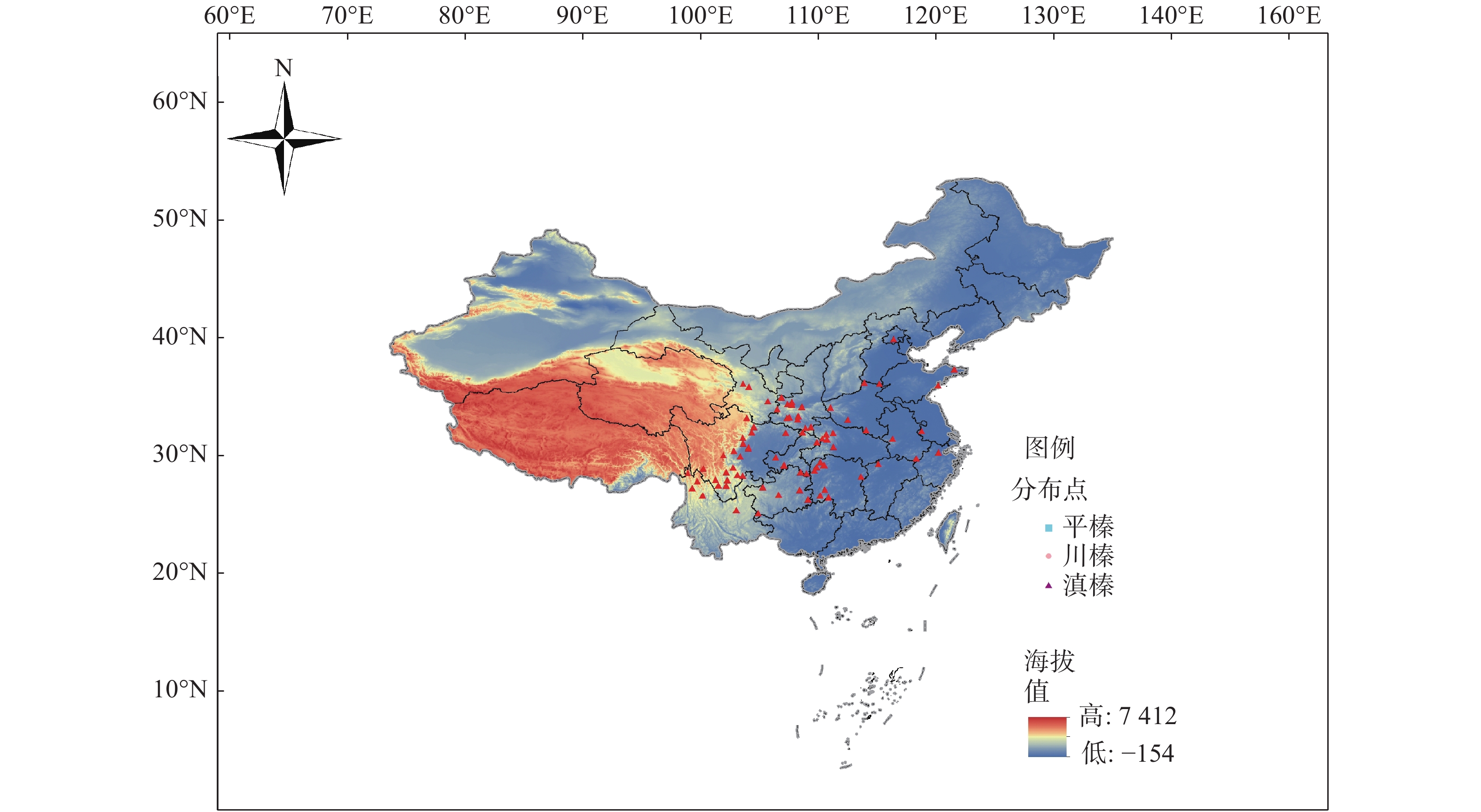
华榛的分布点(图1)数据主要来源于国家标本平台(
http://www.nsii.org.cn )、全球生物多样性信息网(http://www.gbif.org ),去除鉴定错误和一些地名无法核实的记录,并利用本课题组采集的华榛叶片的地理信息进行补充,共获得鉴定较为明确的华榛分布记录数据83个用于后续研究,对筛选得到的标本信息进行经纬度确认,输入Excel表格,保存成csv格式。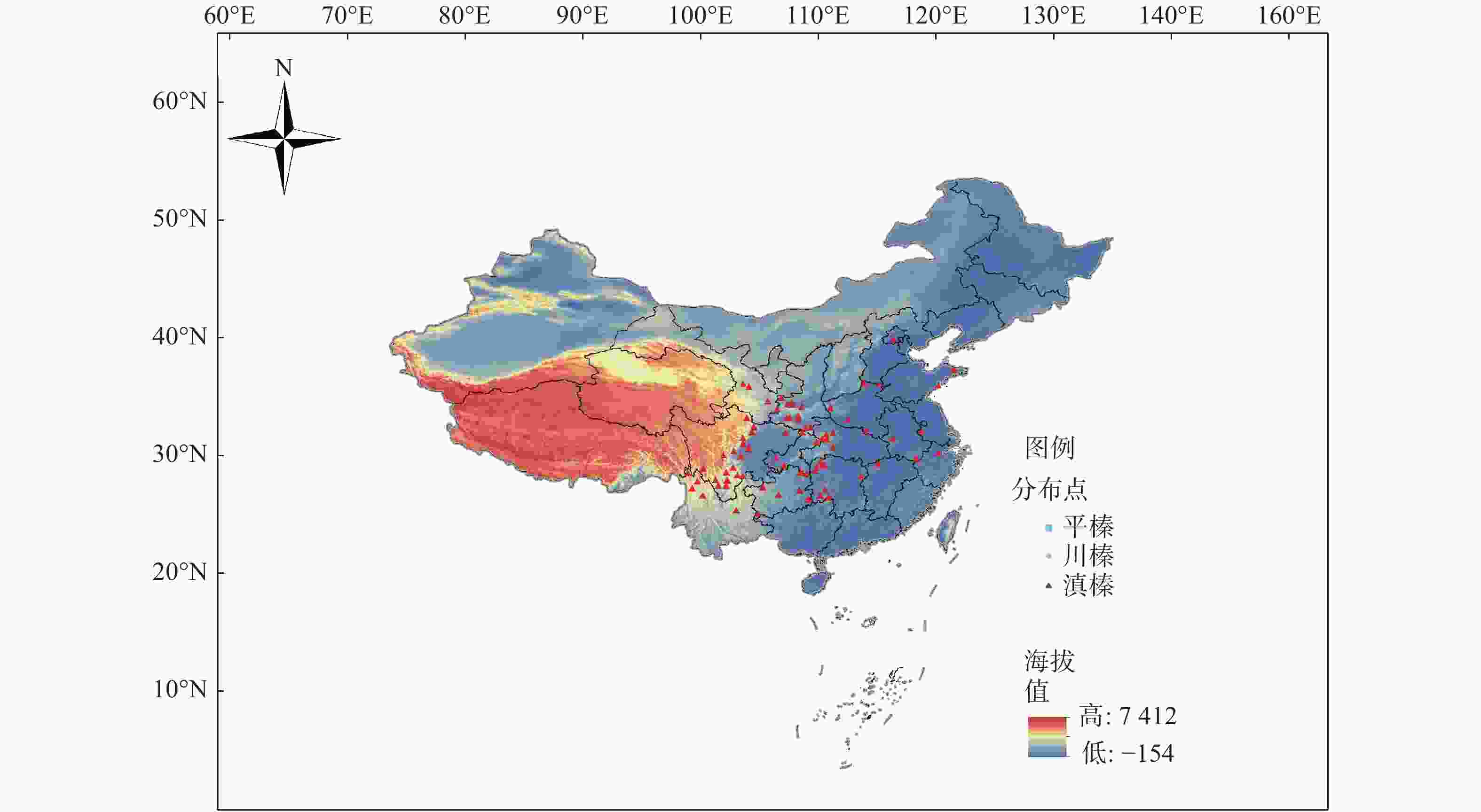
图 1 华榛地理分布点
Figure 1. Distribution of Corylus chinensis Franch.
-
本研究使用30个生态因子,包括世界气候数据库(
http: //world-clim.org/ )提供的不同时期末次间冰期(Last inter-glacial, LIG, 140-120 Ka BP)、末次盛冰期(Last Glacial Maximum, LGM, 22 Ka BP)、全新世中期(Mid Holocene, MH, 6 Ka BP)、现代(1970-2000年)、未来2050s(2041-2060年)和2070s(2061-2080年),2种温室气体排放情景RCP4.5和RCP8.5)的19个气候变量因子和海拔高度、全球UV-B辐射数据库中的6个UV-B变量以及世界土壤数据库中4个土壤因子(养分有效性、养分保持能力、生根条件和根系的氧气有效性)。由于极端气候变化可能对地理分布格局有较大的影响,所以,本研究选取2050s和2070s一种气候模式(CCSM4)下2种温室气体排放情景RCP4.5和RCP8.5的数据。气候数据采用30″空间分辨率,标本数据设置为在每个30″ × 30″网格中只取唯一分布点,上述数据坐标系均为WGS84。采用ENVI和Arcmap10.2对紫外线及土壤数据进行格式转换,得到与气候数据和海拔数据相同的数据格式,使用ArcGIS10.2中的掩膜裁剪工具裁剪并提取这30个环境变量在中国范围内的气候数据,将裁剪后的气候数据转为ASCII格式用于MaxEnt模型[30]。在MaxEnt模型中根据刀切法测试30个环境变量的重要性,然后利用ArcGIS软件中的Spatial Ananlyst工具,提取分布样点的环境变量数值。当模拟一个物种的潜在分布时,环境变量之间的多重共线性可能会妨碍物种与环境关系的分析[31],所以,通过SPSS软件对环境变量进行Pearson相关系数分析,首先剔除贡献率和重要性较低的环境变量,优先选择相关系数小于0.8的环境因子,在相关性系数大于0.8的环境因子中选择贡献率较高的因子构建模型,在经过一系列操作后,最终筛选出18个环境因子构建模型,具体选取的生物变量详见表1。
表 1 用于模型建立的环境变量
Table 1. environment variables for modeling
变量代码
Variable code环境变量
Environment variablesbio 2 每月平均值(最高温度-最低温度)
Mean Diurnal Range (Mean of monthly (max temp - min temp))bio 4 温度季节性(标准偏差 × 100)
Temperature Seasonality (standard deviation × 100)bio 5 最暖月的最高温度
Max Temperature of Warmest Monthbio 6 最冷月份的最低温度
Min Temperature of Coldest Monthbio 7 温度年度范围(bio5-bio6)
Temperature Annual Rangebio 9 最干燥季度的平均温度
Mean Temperature of Driest Quarterbio 11 最冷季的平均温度
Mean Temperature of Coldest Quarterbio 12 年降水量
Annual Precipitationbio 14 最干旱月份的降水
Precipitation of Driest Monthbio 17 最干燥区的降水
Precipitation of Driest Quarterbio 19 最冷季的降水
Precipitation of Coldest QuarterUV-B3 最高月份的平均UV-B
Mean UV-B of Highest MonthUV-B4 最低月份的平均UV-B
Mean UV-B of Lowest MonthAlt 海拔 Elevation 土壤因子SQ1 养分有效性 Nutrient availability 土壤因子SQ2 养分保持能力 Nutrient retention capacity 土壤因子SQ3 生根条件 Rooting conditions 土壤因子SQ4 根系的氧气有效性 Oxygen availability to roots -
将筛选经裁剪后的6个不同时期的环境因子图层、华榛分布点数据(.csv格式)加载到MaxEnt软件中,设置参数进行建模,设分布数据的25%被随机抽取作为测试集,其余75%作为训练集,最大迭代次数是10000,输出文件类型选为.asc,其他参数不作改变,为MaxEnt软件默认值,本次模拟运行10次,选取10次模拟的平均预测结果作为对华榛潜在分布区预测的最终结果。应用ArcGIS10.2软件,对MaxEnt软件运行结果图进行处理,利用平均间距法对华榛的适生区等级进行重分类,将其分为非适生区(适生值为0%~20%)、低度适生区(适生值为20%~40%)、一般适生区(适生值为40%~60%)、中度适生区(适生值为60%~80%)及高度适生区(适生值为80%~100%)五个等级,并利用ArcGIS10.2中SDM toolbox工具中“Centroid Changes (Lines)”工具计算不同时期预测分布的几何中心位移情况,检测华榛分布区的总体变迁趋势,同时得到几何中心变化的矢量重叠密度。
MaxEnt模型预测的准确性程度使用AUC值(受试者工作特征曲线下面积)来评估[32],受试者工作特征曲线下面积的取值一般为0~1,其值越大,表明选取的环境变量与预测的物种地理分布模型间的相关性越大,值越接近1则模型的预测结果越准确。AUC值为0~0.5,模型预测视为失败;AUC值为0.6~0.7,模型预测结果视为较差;AUC值为0.7~0.8,模型预测结果视为一般;AUC值为0.8~0.9,模型预测结果视为良好;AUC值大于0.9时,模型预测结果视为优秀。
在使用MaxEnt模型对物种分布进行预测时经常用到刀切法,刀切法结果有助于分析选取的各环境因子对模型预测结果的影响程度,进而确定影响物种分布的主要环境因子,应用刀切法能够反映各变量对模型预测华榛适生区的影响程度,判断不同的环境因子与所模拟物种适生区分布之间的关系[33]。本次运用MaxEnt3.4.1软件自带的刀切法工具,利用环境变量贡献率与置换重要值,检验不同环境变量限制华榛地理分布的重要性大小,并利用响应曲线评价华榛的适宜气候条件[34]。
-
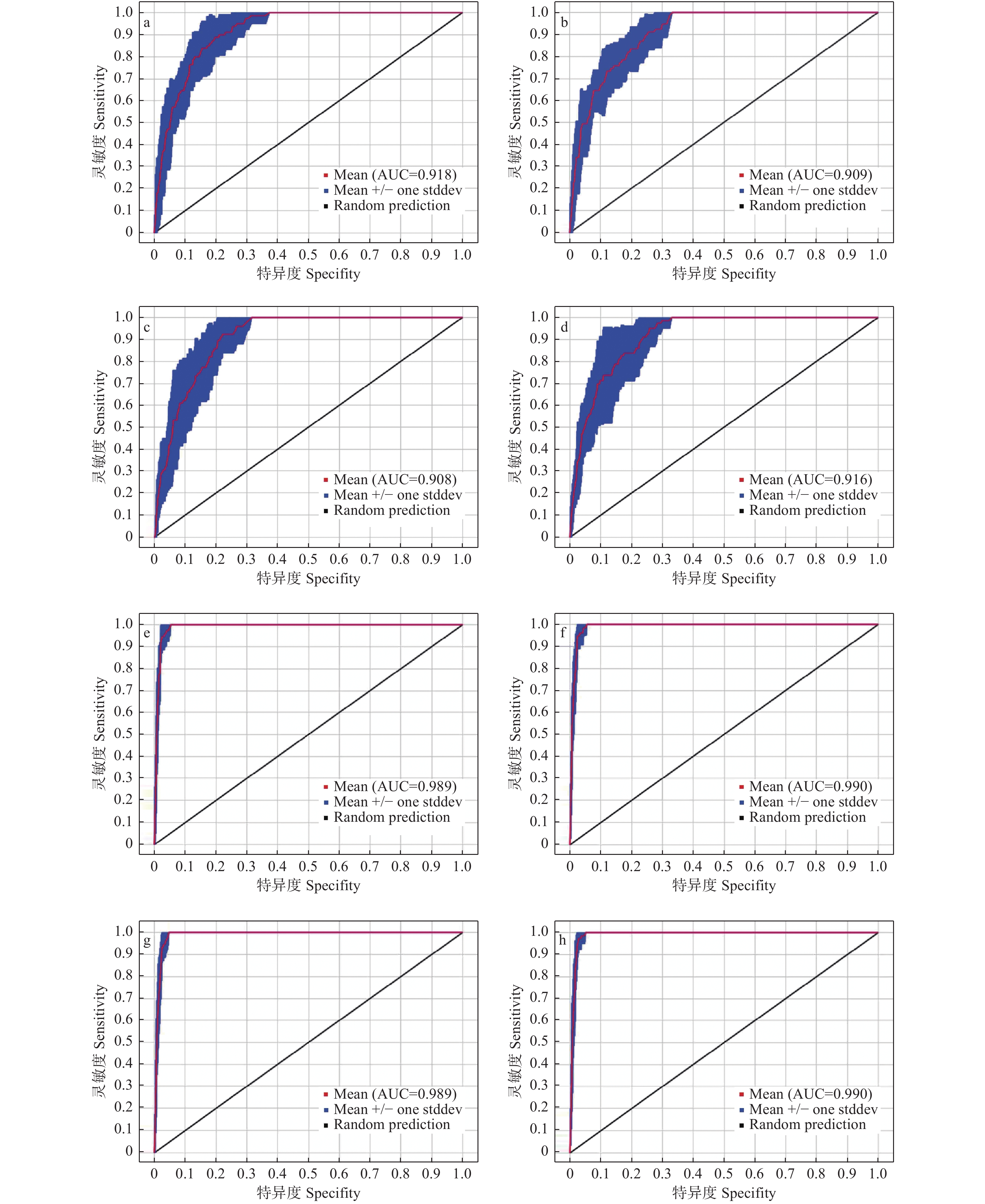
图2表明:6个不同时期的模拟结果显示模型具有较好的预测能力,对华榛现代地理分布预测10次重复的平均AUC值为0.915,标准差为0.032;末次间冰期预测10次重复的平均AUC值为0.916,标准差为0.036;末次盛冰期预测10次重复的平均AUC值为0.908,标准差为0.028;全新世中期预测10次重复的平均AUC值为0.914,标准差为0.025;RCP4.5情境下未来(2050s和2070s)预测的10次重复的平均AUC值分别为0.991、0.990,标准差均为0.004;RCP8.5情境下未来(2050s和2070s)预测的10次重复的平均AUC值分别为0.990、0.989,标准差分别为0.004、0.002,不同时期10次重复的平均AUC值均明显大于随机分布模型的AUC值(0.5),表明此次运用MaxEnt模型对华榛不同时期适生区分布的预测结果较为优秀。
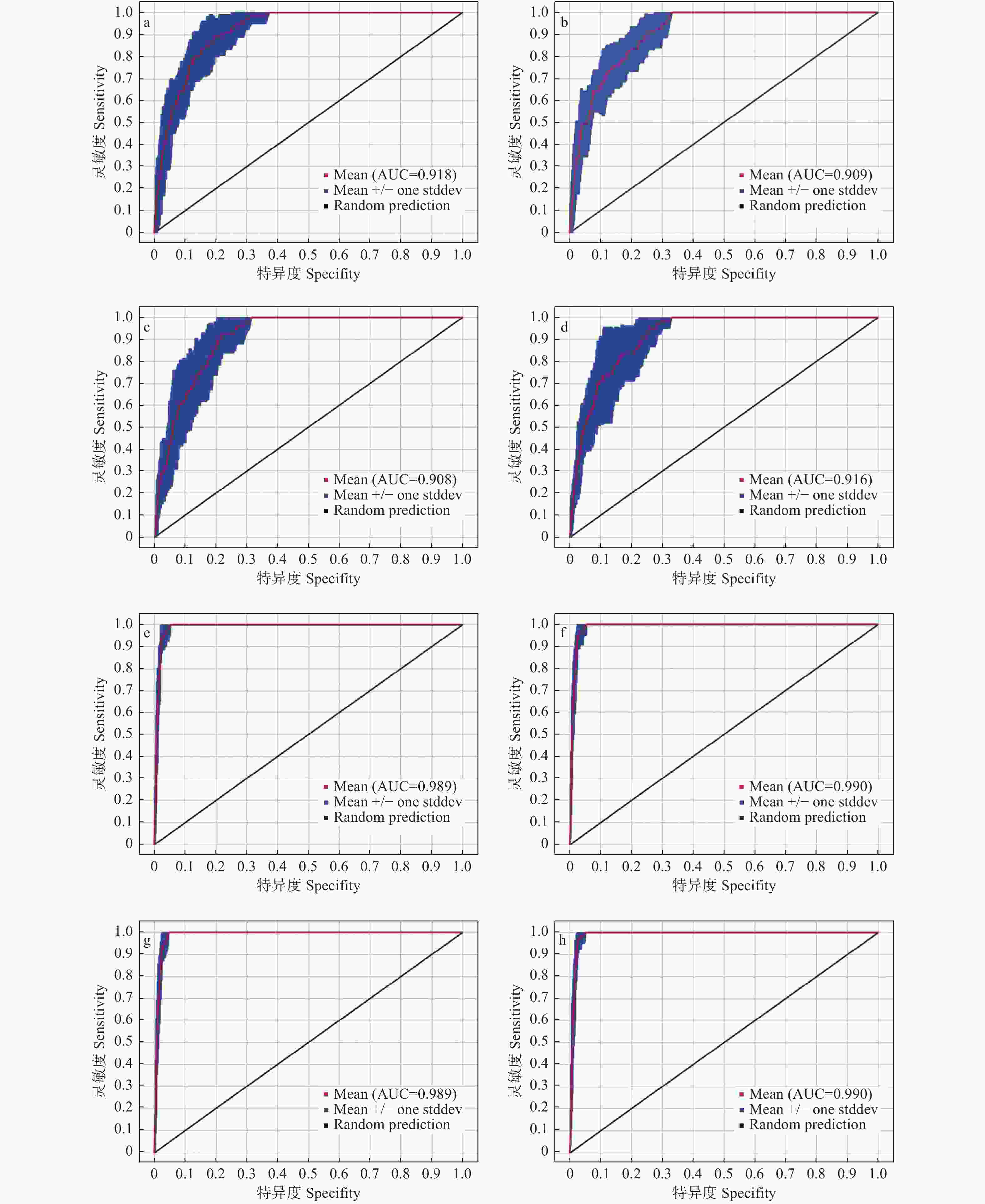
图 2 接受者操作特征曲线检验模型预测的精准度
Figure 2. Receiver operator characteristic curve tests the accuracy of Maxent model
-
本次预测结果的环境因子训练增益结果显示:影响华榛在6个不同时期地理分布的气候因子主要为最冷月份的最低温度(bio6)、最冷季的平均温度(bio11)、最干燥季度的平均温度(bio9),均属于温度因子;其中,使用单独变量进行模型预测时,最冷月份的最低温度的正规化训练增益值、测试增益值和AUC值最高,因此,最冷月份的最低温度被认为是影响华榛分布的主要气候限制条件。
影响华榛分布的主要环境因子的响应曲线(图3)表明:随着环境条件的改变,华榛适生区分布概率会发生一定的变化。以现代为例,其中最冷月份的最低温度(bio6)对预测华榛的潜在分布的贡献较大,其次为最冷季的平均温度(bio11)、最干燥季度的平均温度(bio9)、年降水量(bio12)。根据华榛主要气候因子的响应曲线,划分存在概率大于0.5的气候因子适宜值范围可知:对于华榛的分布及生长,最冷月份最低温度的最适范围是-8~4℃,最冷季平均温度的最适范围是-2~8℃,最干燥季度平均温度的最适范围是-1~9℃,年降水量的最适范围是700~1 500 mm。主要气候因子中,气温因子方面,最冷月份的最低温度在-20~-3℃时,华榛的适生概率随温度的升高而增大,最冷月份最低温度大于−3℃后适生概率随温度的升高而下降;最干燥季度的平均温度在-10~4℃时,华榛的适生概率随温度的升高而增大,在最干燥季度的平均温度大于4℃后适生概率随温度的升高而下降;最冷季的平均温度在-10~3℃时,华榛的适生概率随温度的升高而增大,在最冷季的平均温度大于3℃后适生概率随温度的升高而下降。降水因子方面,年降水量在800 mm以下,华榛的适生概率随降水量的增多而上升,在800 mm以上,华榛的适生概率随降水量的增多逐渐下降。
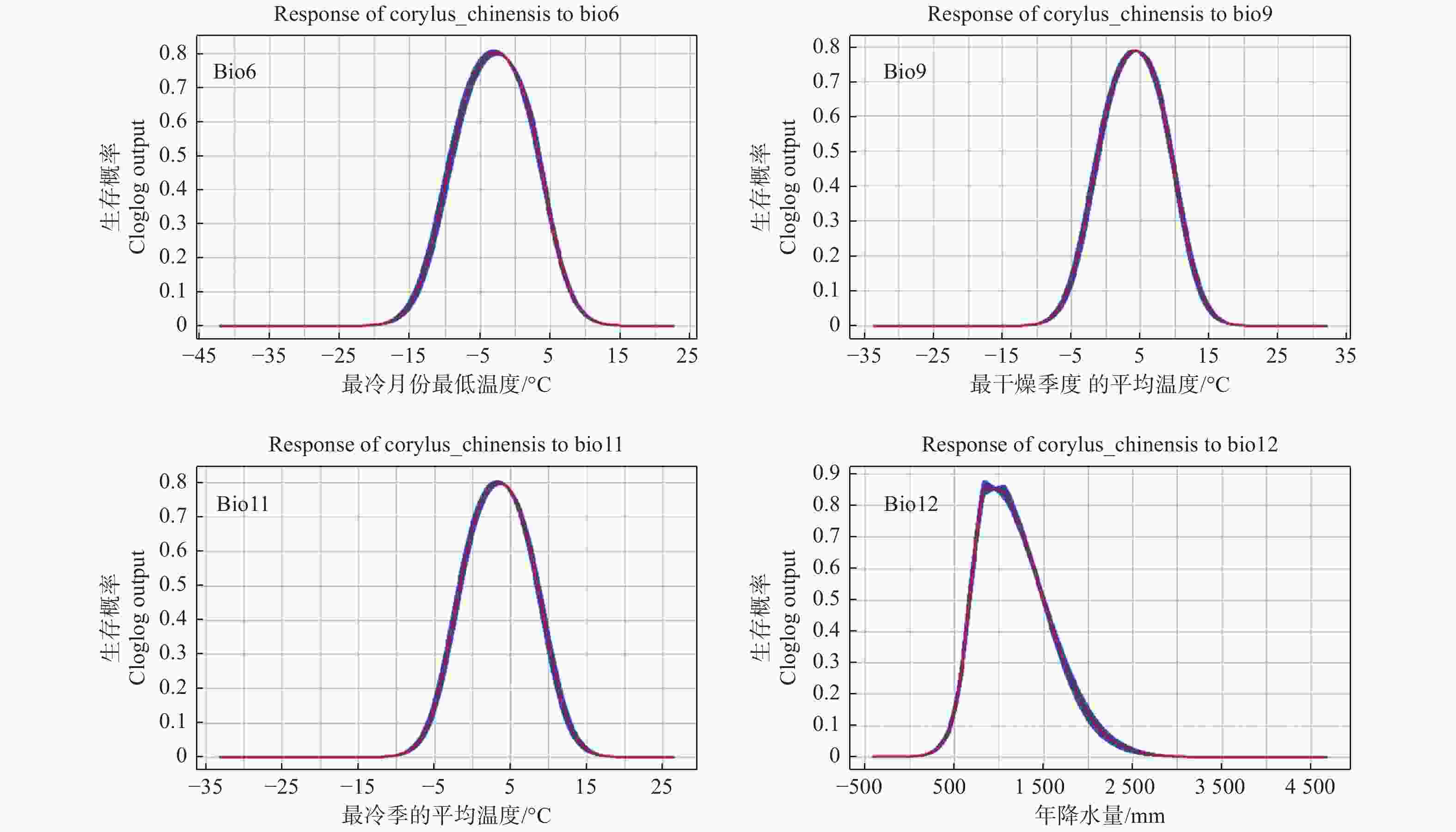
图 3 预测分布概率与主要气候因子的响应曲线
Figure 3. Response curves between prediction value and bioclimatic variables
-
运用ArcGIS10.2软件的重分类工具对华榛不同时期不同气候条件下的分布区域进行适生区等级划分,得到华榛在6个时期不同气候条件下的适生等级分布图及华榛的分布中心转移图(图4、5)。
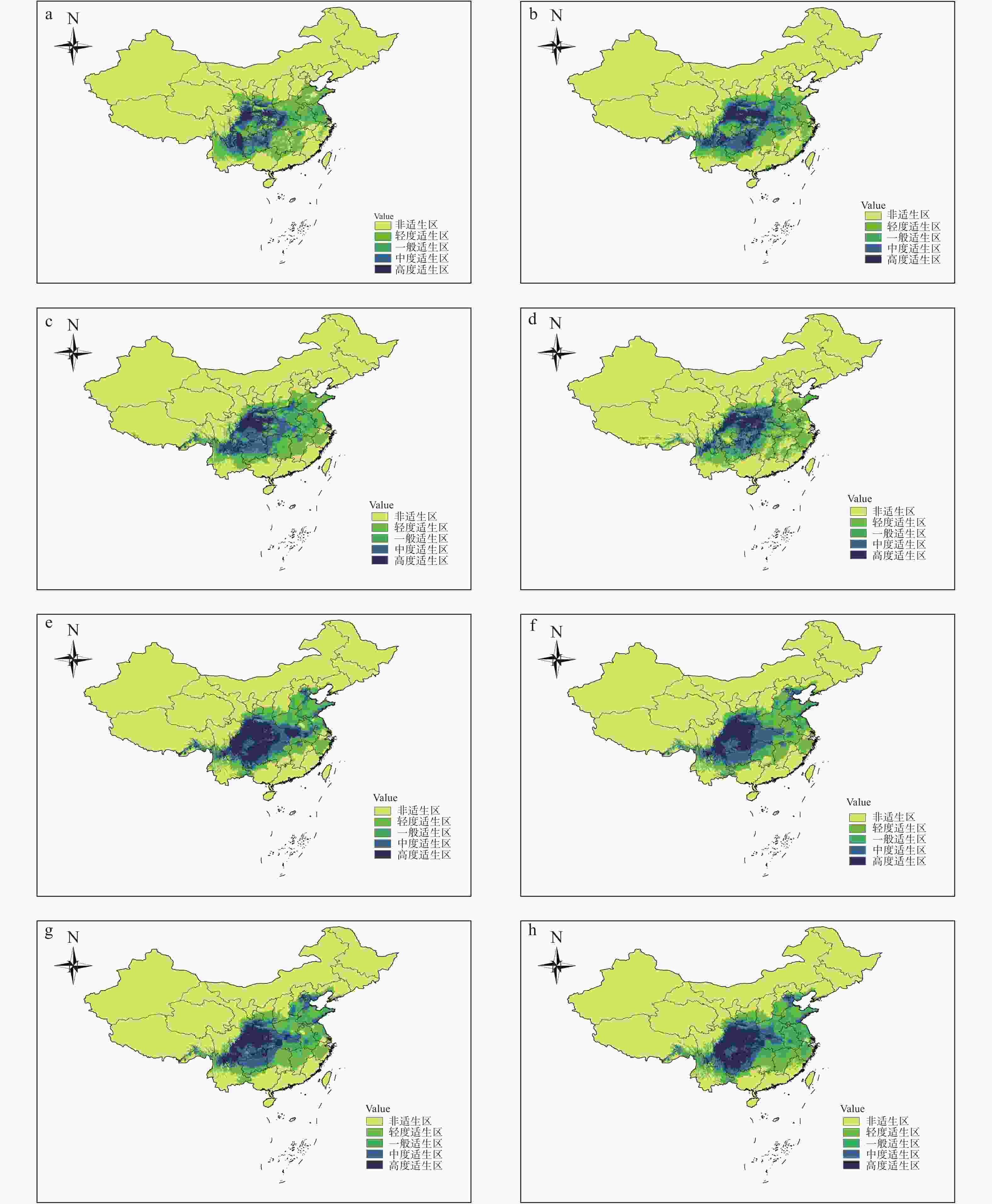
图 4 华榛6个时期适生区分布变迁
Figure 4. Historical changes of potential distribution areas of Corylus chinensis Franch. in six periods
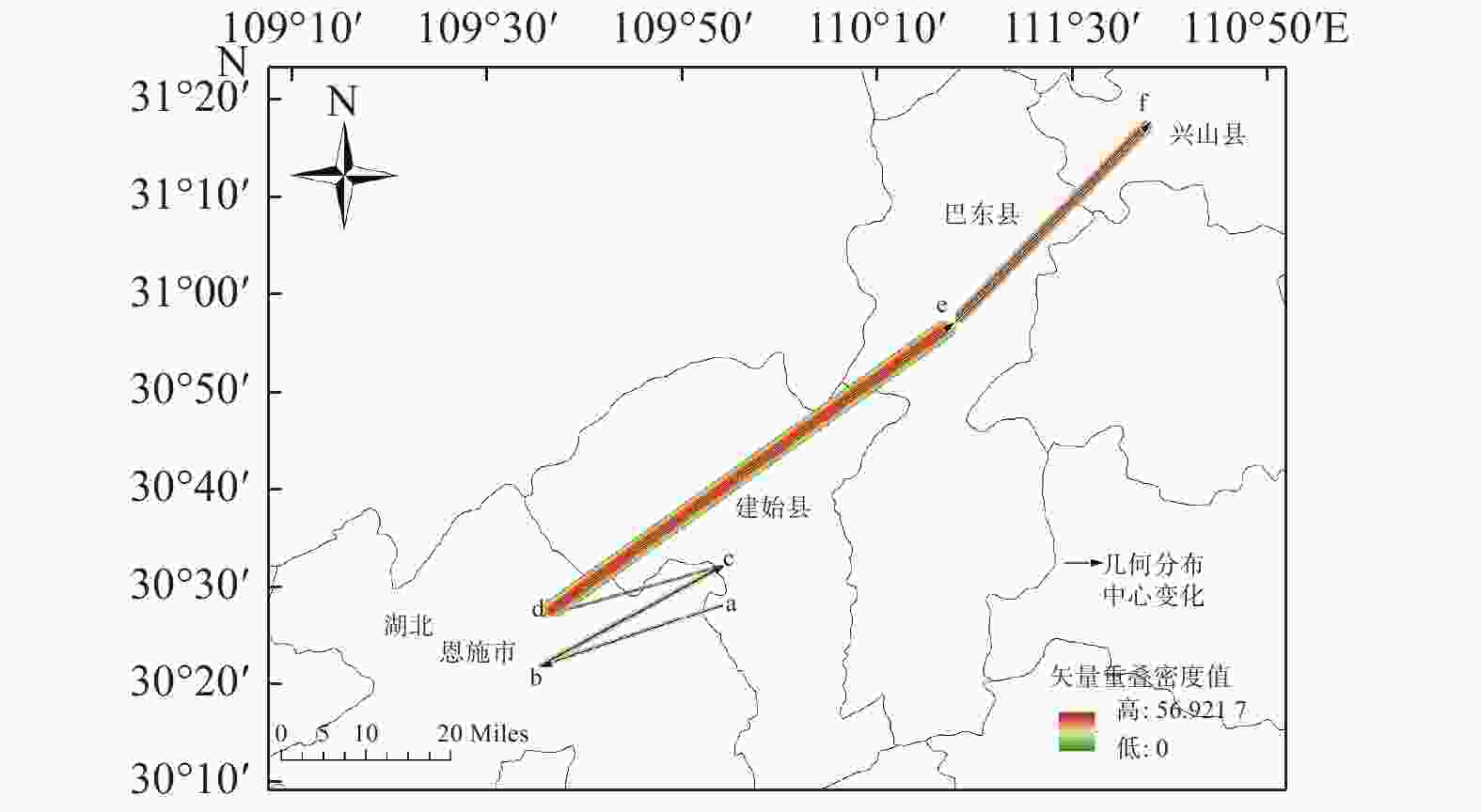
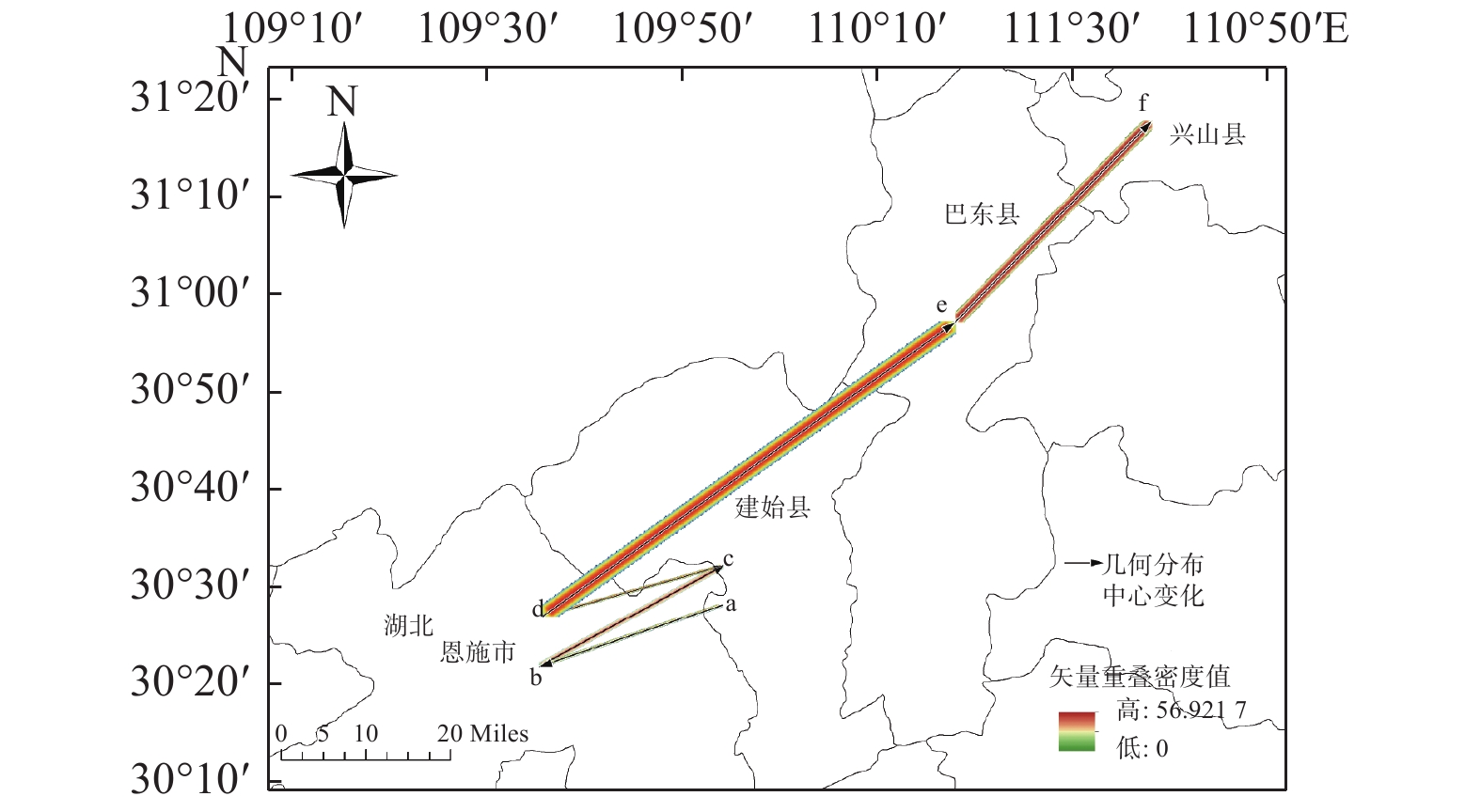
图 5 华榛的分布中心变化
Figure 5. Changes in the distribution center of Corylus chinensis Franch.
-
模型模拟结果显示(图4-d),生态位模型模拟的华榛现代分布结果同华榛实际分布点区域基本相符,模拟的华榛现代适生区主要分布于云南、四川、陕西、河南、湖北、贵州六省,其中,高度适生区位于四川东北部、甘肃东南部、陕西西南部、重庆东北部、湖北西北部,主分布在大巴山、米仓山附近及汉中盆地一带,在四川主要分布于四川盆地北部,湖北神农架林区一带也存在华榛的高度适生区。
-
末次间冰期(图4-a)华榛的高度适生区主要分布在云南、四川东北部,甘肃东南部、陕西西南部、湖北中部,主分布在鄂西山地东侧、云南五连峰、乌蒙山附近,甘肃临江、陕西安康汉中一带以及贵州佛顶山地区。表2表明:华榛在末次间冰期的适生区总面积比现代的减少,适生区总面积相比现代减少9%左右。分布区内东部与西部面积收缩,高度适生区位置相对现代南移。
表 2 华榛不同时期在中国适生区预测
Table 2. Prediction of habitat of Corylus chinensis Franch. in China at different periods
万km2 时间段
Periods总适生区
Total suitable region低度适生区
Lowly suitable region一般适生区
General suitable region中度适生区
Moderately suitable region高度适生区
Highly suitable region末次间冰期 LIG 203.167 8 87.782 5 54.881 4 38.131 5 22.372 4 末次盛冰期 LGM 224.189 2 80.691 0 65.107 6 49.633 7 28.756 9 全新世中期 MH 232.977 4 81.121 5 68.816 0 61.010 4 22.029 5 现代 Current 223.244 8 94.470 5 64.887 1 38.243 1 25.644 1 2050s,RCP4.5 231.046 9 61.677 1 64.529 5 55.783 0 49.057 3 2050s,RCP8.5 237.201 4 83.809 0 59.081 6 42.668 4 51.642 4 2070s,RCP4.5 253.359 4 74.031 3 79.732 6 59.671 9 39.923 6 2070s,RCP8.5 252.427 1 73.055 6 79.722 2 55.404 5 44.244 8 末次盛冰期(图4-b),华榛的高度适生区主要分布在四川东北部、贵州东部部分区域、湖北西北部,陕西、甘肃南部及湖南少许区域,主分布于秦岭及米仓山一带。与末次间冰期对比,华榛的总适生区面积呈扩大趋势,一般适生区分布向北移动。华榛适生区总面积相对现代变化不大,其中,高度适生区面积比现代多12.1%。末次间冰期到末次盛冰期,华榛总适生区面积增加10.3%。
全新世中期(图4-c),华榛的适生区在四川、重庆、陕西、贵州、湖南张家界、湖北襄阳武汉地带、河南信阳、山东济宁、安徽、江苏盐城、湖南郴州、江西、浙江均有分布,高度适生区位于四川东北部、四川南部及陕西西南部,少许位于喜马拉雅山脉附近及横断山脉东南部及大巴山和秦岭一带。在全新世中期,华榛适生区总面积相对现代多4.4%;从末次盛冰期到全新世中期,总适生区面积增加3.9%。
-
在未来气候条件下,高度适生区主要分布在四川东北部、东南部,重庆大部分区域,贵州北部,湖北恩施附近,在大巴山、巫山及四川盆地附近。2种不同情景RCP4.5和RCP8.5下,华榛适生区分布总体差距不大,比现代适生区面积均有增加。RCP4.5情景下,相对于现代分布,2050s(图4-e)华榛高度适生区向西南方向移动,中度适生区及高度适生区面积有所扩大,低度适生区面积减少,预测潜在适宜分布区面积增大,与现代相比总适生区面积增加了3.5%,高度适生区面积增加了91.3%;2070s(图4-g)华榛的高度适生区主要分布在横断山脉、大巴山、巫山、四川盆地及芙蓉江、大娄山、清江地区。华榛总适生区面积比现代增加13.5%,高度适生区面积增加了55.7%。华榛在贵州地区部分高度适生区及湖北地区部分高度适生区转为中度适生区,通过和海拔图叠加发现,在未来气候变暖的情况下,该物种往高海拔地区收缩。
图5预测结果显示:华榛不同时期的分布中心均位于湖北省,分布在恩施、建始、巴东、兴山4县,在未来气候变暖情况下分布中心呈向高纬度地区迁移的趋势。总体来说,由末次间冰期到末次盛冰期,华榛的高度适生区向喜马拉雅山脉一带延伸,高度适生区面积增多。从末次盛冰期到全新世中期,贵州、湖南地区高度适生区消失。从全新世中期到现代,华榛在云南与四川交界处高度适生区面积大幅度缩小,大巴山脉附近湖北地区高度适生区面积增多。2050s华榛高度适生区分布相对现代向西南方向移动,2070s高度适生区分布面积较2050s缩小。
-
因自然及人为原因共同作用所导致的全球气候变化是目前我们所面临的较严重的环境问题,全球气候变暖会导致北极气温的改变、冰覆盖量的减少、永久冻土的融化及冰原面积的缩小等各种问题。近百年来,气温升高速度明显增加并且有越来越快的趋势,气候的变化也影响着二氧化碳的排放及降水量的变化,导致极端天气增多,也因此对生态系统及生物多样性产生了显著的影响,进而可能引起物种分布的变化,如物种随着气候条件的改变其适生区会发生一定的迁移。气候的变化对不同物种的影响作用可能不同,李昂[35]对樟子松的研究发现,水分逐渐成为我国东北地区樟子松林分布的主要限制因素。张文秀等[36]对濒危植物白豆杉进行模拟预测时发现,降水因子是影响白豆杉分布的主要因子,这与白豆杉更适合潮湿多雨的气候特性有关。张爱平等[37]对3种云杉属植物的研究发现,温度是影响3种云杉属植物的主要环境因子,与前人对绝大多数云杉属植物的主要环境影响因子的研究相符。对于被子植物来说,同样潜在适生区位于亚热带的伯乐树的分布受到最暖季度降雨量的影响最大,降水丰富且潮湿的环境更适合其生存[38]。李璇等[39]对白栎潜在分布区的预测发现,水热条件共同制约着白栎的分布格局,白栎喜温暖环境且抗干旱能力较好,温度和水分在不同的维度上影响着白栎的分布。毛榛(Corylus mandshurica)与华榛同属榛属植物,王琦等[40]对毛榛的潜在适生区预测发现,温度、海拔、降水量都影响着毛榛的分布,是影响其分布的重要环境因子,影响其分布的各因子的最适范围与毛榛耐寒、耐荫的生物学习性相符。
本次对华榛潜在分布区模拟结果显示:受试者工作特征曲线下面积均大于0.9,表明对华榛的潜在分布区进行的模拟预测是有效预测。MaxEnt模型相对其他预测软件更适合模拟分布数据有限的物种,由分析结果可知,影响末次间冰期华榛分布的气候因子中最冷月份的最低温度居首位,末次盛冰期和全新世中期影响华榛分布的主要气候因子均为最冷月份的最低温度和最冷季的平均温度,当前气候分析结果表明,影响华榛现代时期分布最主要的气候因子是最冷月份的最低温度。未来气候条件下,最干燥季度的平均温度及最冷季的平均温度是影响华榛分布的主要气候因子,总体来说,温度是影响华榛适宜区分布的主要环境因子。对于华榛的分布及生长,最冷月份最低温度的最适范围是-8~4℃,最冷季平均温度的最适范围是-2~8℃,最干燥季度平均温度的最适范围是-1~9℃,年降水量的最适范围是700~1 500 mm。在所选取的环境因子中,影响华榛的首要环境因子虽然是温度,但降水量也同样影响着华榛的潜在适生区分布,这与华榛主要分布区地处中亚热带至北亚热带,多生于中山地带,喜温凉、湿润的气候环境的实际生长特性相符。总体来说,气候影响着物种的分布,无论是裸子植物还是被子植物,它们的潜在适生区分布都受到不同环境因子的影响,温度与水分的影响比重较大,影响物种的具体主导环境因子与物种本身的生物学习性息息相关,在对这些物种进行开发利用及保护的过程中应该考虑其地理分布与环境之间的关系,为其开发利用提供更好的科学依据。虽然本文所选取的18个环境因子涉及到气候、光照、海拔、土壤几个因素,但未考虑人为因素的影响,之后的研究中应综合考虑多种因素的影响,这样有助于对华榛的潜在分布区进行更精准的模拟预测。
-
第四纪以来,物种的分布格局受到剧烈变化的地球气候的不断影响[41-42]。末次盛冰期全球大降温背景下,不同地区森林面积在不同程度上呈减少和破碎化趋势,中国东部暖温带常绿阔叶林和混交林向南推移;在全新世中期(暖期),中国东部暖温带常绿阔叶林和混交林向北推移。就现代气候情况而言,大多物种面临着全球气候变暖的威胁[43-44]。本次对华榛不同时期潜在分布区的预测结果具有较高的可信度,由此可以推测第四纪气候变化对华榛地理分布的影响,结果显示,华榛在末次间冰期的适生区面积少于现代华榛的适生区面积,在末次盛冰期时期的适生区面积与现代适生区总面积相差无几,全新世中期华榛的适生区面积高于现代华榛的适生区面积。研究发现,在末次盛冰期时期,冰川未直接作用于中国亚热带,中国亚热带地区虽温度有所下降,但受到地形因素的影响,亚热带山区的湿度变化范围不大,中国南方亚热带是亚热带与热带的交界地区,降水丰富,形成的水热条件适合森林植被的生长[45],华榛主要生活在亚热带地区,这可能是华榛在末次盛冰期适生区面积没有大幅度变化的原因。
由末次盛冰期至全新世中期,华榛的总适生区面积增加。全新世中期气候温暖湿润,降水强度增大,在此种气候条件下喜此种气候的常绿阔叶林呈分布区小幅度向北推进的趋势。郑益群等[46]认为,未来由二氧化碳浓度增加而引起的全球增温现象与全新世中期的增温效应相似。将模型模拟的华榛过去3个时期的适生区与收集到的孢粉数据做对照发现,预测到的华榛分布区部分省份有孢粉数据相对应,如在末次间冰期模拟的华榛分布地与收集到的陕西、云南孢粉数据对应,末次盛冰期模拟的华榛分布地与收集到的陕西、云南、湖北、甘肃、四川孢粉数据相对应,全新世中期模拟的华榛分布地与收集到的四川、甘肃、湖北、湖南、贵州孢粉数据相对应。由于收集到的孢粉数据有限且不区分具体种,因此,孢粉信息只具有部分参考性。
在2050s和2070s,相对现代,华榛的适生区面积都呈增加趋势,和全新世中期适生区面积变化趋势相似。2个增温的环境下,华榛的分布区均表现为增加,其中,全新世中期的增幅略大,说明气候变暖对华榛的生长有一定的正面影响。研究发现,在现今全球气候不断变暖的情况下,许多动植物的迁移趋势呈现向高纬度和高海拔地区的移动,在对华榛未来潜在分布区预测中发现,对比于现代,在2050s及2070s华榛的高度适生区都有向高纬度迁移的趋势,其不同时期的分布中心在未来气候变暖情况下,分布中心呈现向高纬度地区迁移的趋势,这与之前的研究结果相符。对于同属榛属的毛榛来说,未来气候变暖对毛榛的地理分布也产生了一定的影响,其高度适生区也有向高纬度及高海拔地区扩增的趋势[40],与本文华榛在未来气候变暖情境下的地理变迁趋势相同。
-
本研究利用生态位模型中的MaxEnt模型并结合ArcGIS软件模拟了华榛自末次间冰期以来6个不同时期的适生区分布。根据华榛地理分布和环境因子之间的联系,发现最冷月份的最低温度(bio6)、最冷季的平均温度(bio11)、最干燥季度的平均温度(bio9)3个与温度相关的气候因子为决定华榛6个不同时期地理分布的主导气候因子,温度条件对华榛地理分布的影响巨大,气候因子中华榛的最适生存范围也印证了华榛喜温凉、湿润环境的生物特性。对华榛适生区分布模拟显示,对比现代分布,华榛在末次间冰期适生区面积减少,末次盛冰期适生区面积变化不大,全新世中期及未来华榛的适生区面积高于现代华榛的适生区面积。在当前全球气候变暖背景下,华榛的适生区分布呈增加趋势,同时高度适生区有向高纬度迁移的趋势。
气候变化下濒危树种华榛的潜在适生区预测
Prediction of Potential Distribution of Endangered Species Corylus chinensis Franch. in Climate Change Context
-
摘要:
目的 探究华榛(Corylus chinensis Franch.)地理分布格局随气候的变化趋势,划分不同等级的华榛适生区,为华榛的保护利用提供科学依据。 方法 本研究基于华榛83条现代分布记录信息和筛选后的18个环境变量数据,采用物种分布模型中的最大熵模型(MaxEnt)模拟华榛末次间冰期以来6个时期的潜在分布区。 结果 受试者工作特征曲线下方面积(AUC)结果显示:模拟结果较为准确,温度是影响华榛在6个不同时期地理分布的重要因素。末次间冰期华榛的适生区总面积相比现代少9%左右,末次盛冰期变化不大,全新世中期适生区总面积相对现代多4.4%,预测未来2050s和2070s适生区面积均增加。华榛不同时期的分布中心均位于湖北省,在未来气候变暖情况下分布中心呈向高纬度地区迁移的趋势。从适生区面积来看,华榛在冰期时适生区面积呈收缩趋势,在全新世中期及未来全球气候变暖情境下适生区面积呈扩张趋势。 结论 华榛在气候变化的情境下适生区分布区域不同,温度是影响其分布的主要因子,未来气候变暖条件下分布中心有向高纬度方向迁移的趋势。 Abstract:Objective To explore the change trend of the geographical distribution pattern of Corylus chinensis with climate, divide the suitable areas and provide references for the protection and utilization of C. chinensis. Method Based on 83 modern distribution records of C. chinensis and the screened data of 18 environmental variables, the maximum entropy model (MaxEnt) in the species distribution model was used to simulate the potential distribution areas of C. chinensis in six periods since the last interglacial period. Result The results of the area under receiver operating curve (AUC) show that the simulation results are more accurate, and temperature is an important factor affecting the geographical distribution of C. chinensis in six different periods. During the last interglacial period, the total suitable area of C. chinensis decreased by about 9% compared with modern times, and there was little change during the last glacial maximum period. The total suitable area increased by 4.4% compared with modern times in the middle Holocene. It is predicted that the suitable area will increase in 2050s and 2070s. The distribution centers of C. chinensis in different periods are all located in Hubei Province. In the future, the distribution center will show a tendency to migrate to high latitudes under the condition of future climate warming. From the perspective of the suitable area, C. chinensis shows a shrinking trend in the glacial period, and shows an expansion trend in the mid-Holocene and future global warming scenarios. Conclusion The distribution area of C. chinensis suitable for climate change is different. Temperature is the main factor affecting its distribution. Under the condition of climate warming in the future, the distribution center will migrate to high latitude. -
图 2 接受者操作特征曲线检验模型预测的精准度
Figure 2. Receiver operator characteristic curve tests the accuracy of Maxent model
图 3 预测分布概率与主要气候因子的响应曲线
Figure 3. Response curves between prediction value and bioclimatic variables
图 4 华榛6个时期适生区分布变迁
Figure 4. Historical changes of potential distribution areas of Corylus chinensis Franch. in six periods
图 5 华榛的分布中心变化
Figure 5. Changes in the distribution center of Corylus chinensis Franch.
表 1 用于模型建立的环境变量
Table 1. environment variables for modeling
变量代码
Variable code环境变量
Environment variablesbio 2 每月平均值(最高温度-最低温度)
Mean Diurnal Range (Mean of monthly (max temp - min temp))bio 4 温度季节性(标准偏差 × 100)
Temperature Seasonality (standard deviation × 100)bio 5 最暖月的最高温度
Max Temperature of Warmest Monthbio 6 最冷月份的最低温度
Min Temperature of Coldest Monthbio 7 温度年度范围(bio5-bio6)
Temperature Annual Rangebio 9 最干燥季度的平均温度
Mean Temperature of Driest Quarterbio 11 最冷季的平均温度
Mean Temperature of Coldest Quarterbio 12 年降水量
Annual Precipitationbio 14 最干旱月份的降水
Precipitation of Driest Monthbio 17 最干燥区的降水
Precipitation of Driest Quarterbio 19 最冷季的降水
Precipitation of Coldest QuarterUV-B3 最高月份的平均UV-B
Mean UV-B of Highest MonthUV-B4 最低月份的平均UV-B
Mean UV-B of Lowest MonthAlt 海拔 Elevation 土壤因子SQ1 养分有效性 Nutrient availability 土壤因子SQ2 养分保持能力 Nutrient retention capacity 土壤因子SQ3 生根条件 Rooting conditions 土壤因子SQ4 根系的氧气有效性 Oxygen availability to roots  下载: 导出CSV
下载: 导出CSV
表 2 华榛不同时期在中国适生区预测
Table 2. Prediction of habitat of Corylus chinensis Franch. in China at different periods
万km2 时间段
Periods总适生区
Total suitable region低度适生区
Lowly suitable region一般适生区
General suitable region中度适生区
Moderately suitable region高度适生区
Highly suitable region末次间冰期 LIG 203.167 8 87.782 5 54.881 4 38.131 5 22.372 4 末次盛冰期 LGM 224.189 2 80.691 0 65.107 6 49.633 7 28.756 9 全新世中期 MH 232.977 4 81.121 5 68.816 0 61.010 4 22.029 5 现代 Current 223.244 8 94.470 5 64.887 1 38.243 1 25.644 1 2050s,RCP4.5 231.046 9 61.677 1 64.529 5 55.783 0 49.057 3 2050s,RCP8.5 237.201 4 83.809 0 59.081 6 42.668 4 51.642 4 2070s,RCP4.5 253.359 4 74.031 3 79.732 6 59.671 9 39.923 6 2070s,RCP8.5 252.427 1 73.055 6 79.722 2 55.404 5 44.244 8
下载: 导出CSV
-
[1] 王贵禧. 中国榛属植物资源培育与利用研究(Ⅰ)——榛种质资源研究[J]. 林业科学研究, 2018, 31(1):105-112. [2] 王贵禧. 中国榛属植物资源培育与利用研究(Ⅲ)——育种、育苗与栽培[J]. 林业科学研究, 2018, 31(1):122-129. [3] 王贵禧. 中国榛属植物资源培育与利用研究(Ⅱ)——形态发育、生理和分子生物学研究[J]. 林业科学研究, 2018, 31(1):113-121. [4] 王贵禧. 中国榛属植物资源培育与利用研究(Ⅳ)——榛仁营养、综合利用与榛产业发展现状[J]. 林业科学研究, 2018, 31(1):130-136. [5] 赵天田, 梁丽松, 马庆华, 等. 平榛ChWRKY28基因克隆及表达模式分析[J]. 林业科学研究, 2016(2):250-255. doi: 10.3969/j.issn.1001-1498.2016.02.016 [6] 蒲发光, 黄武刚, 肖正东, 等. 安徽植物地理分布新纪录树种——华榛[J]. 安徽林业科技, 2013, 29(2):51-52. doi: 10.3969/j.issn.2095-0152.2013.02.014 [7] 李翩翩, 张国禹, 黄桂云, 等. 华榛播种育苗技术[J]. 中国园艺文摘, 2018, 34(6):187-188. doi: 10.3969/j.issn.1672-0873.2018.06.068 [8] 姜在民, 蔡 靖, 崔宏安. 华榛, 羽叶丁香种子形态构造特点的研究[J]. 陕西林业科技, 1999(3):14-16. doi: 10.3969/j.issn.1001-2117.1999.03.004 [9] 杨亚莉, 蒋丽娟, 向祖恒, 等. 龙山县八面山华榛人工林造林成效分析[J]. 湖南林业科技, 2014, 41(3):46-50. doi: 10.3969/j.issn.1003-5710.2014.03.011 [10] 刘祝祥, 陈功锡, 欧阳姝敏, 等. 华榛种仁油提取及GC-MS分析[J]. 中国油脂, 2011, 36(009):14-17. [11] 王玉君, 李玉杰, 张晋东. 气候变化对大熊猫影响的研究进展[J]. 野生动物学报, 2018, 39(3):709-715. doi: 10.3969/j.issn.1000-0127.2018.03.046 [12] 汪青春, 刘彩红, 李万志. 全球气候变化中的可可西里[J]. 世界遗产, 2017(4):60-62. doi: 10.3969/j.issn.1812-0946.2017.04.015 [13] 曹 策, 蒋 昕. 植物分布对气候变化的响应[J]. 绿色科技, 2017(16):111-113. [14] Zeng J R, Li C M, Liu J Z, et al. Ecological assessment of current and future Pogostemon cablin Benth. potential planting regions in China based on MaxEnt and ArcGIS models[J]. Journal of Applied Research on Medicinal and Aromatic Plants, 2021, 24: 100308. doi: 10.1016/j.jarmap.2021.100308 [15] Villén‐Peréz S, Heikkinen J, Salemaa M, et al. Global warming will affect the maximum potential abundance of boreal plant species[J]. Ecography, 2020, 43(6): 801-811. doi: 10.1111/ecog.04720 [16] Lippmann R, Babben S, Menger A, et al. Development of Wild and Cultivated Plants under Global Warming Conditions[J]. Current Biology, 2019, 29(24): R1326-R1338. doi: 10.1016/j.cub.2019.10.016 [17] Xu D P, Zhuo Z H, Wang R L, et al. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling[J]. Global Ecology and Conservation, 2019, 19: e00691. doi: 10.1016/j.gecco.2019.e00691 [18] 朱耿平, 刘国卿, 卜文俊, 等. 生态位模型的基本原理及其在生物多样性保护中的应用[J]. 生物多样性, 2013, 21(1):90-98. [19] Sharma S, Arunachalam K, Bhavsar D, et al. Modeling habitat suitability of Perilla frutescens with MaxEnt in Uttarakhand—A conservation approach(Article)[J]. Journal of Applied Research on Medicinal and Aromatic Plants, 2018, 10: 99-105. doi: 10.1016/j.jarmap.2018.02.003 [20] Tang J H, Li J H, Lu H, et al. Potential Distribution of an Invasive Pest, Euplatypus parallelus, in China as predicted by Maxent[J]. Pest management science, 2019, 75(6): 1630-1637. doi: 10.1002/ps.5280 [21] Xu N, Meng F Y, Zhou G F, et al. Assessing the suitable cultivation areas for Scutellaria baicalensis in China using the Maxent model and multiple linear regression[J]. Biochemical Systematics & Ecology, 2020, 90: 104052. [22] 贺水莲, 孟 静, 洪明伟, 等. 基于MaxEnt软件的忍冬与灰毡毛忍冬的适生分布区预测[J]. 云南农业大学学报:自然科学, 2015, 30(5):777-783. [23] Yan H Y, Feng L, Zhao Y F, et al. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt[J]. Global Ecology and Conservation, 2020, 21: e00856. doi: 10.1016/j.gecco.2019.e00856 [24] Namgung H, Kim M J, Baek S, et al. Predicting potential current distribution of Lycorma delicatula (Hemiptera: Fulgoridae) using MaxEnt model in South Korea[J]. Journal of Asia-Pacific Entomology, 2020, 23(2): 291-297. doi: 10.1016/j.aspen.2020.01.009 [25] Qin A L, Jin K, Batsaikhan M E, et al. Predicting the current and future suitable habitats of the main dietary plants of the Gobi Bear using MaxEnt modeling[J]. Global Ecology and Conservation, 2020, 22: e01032. doi: 10.1016/j.gecco.2020.e01032 [26] 赵 健, 李志鹏, 张华纬, 等. 基于MaxEnt模型和GIS技术的烟粉虱适生区预测[J]. 植物保护学报, 2019, 46(6):1292-1300. [27] 王运生, 谢丙炎, 万方浩, 等. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4):365-372. doi: 10.3321/j.issn:1005-0094.2007.04.005 [28] 曹向锋, 钱国良, 胡白石, 等. 采用生态位模型预测黄顶菊在中国的潜在适生区[J]. 应用生态学报, 2010, 21(12):3063-3069. [29] 孙文涛, 刘雅婷. 生物入侵风险分析的研究进展[J]. 中国农学通报, 2010, 26(7):233-236. [30] 杨冬臣, 王佳颖, 李 静, 等. 基于Maxent生态位模型的外来入侵植物刺果瓜在我国的适生区预测[J]. 河北农业大学学报, 2019, 42(3):45-50. [31] Ji W, Han K, Lu Y Y, et al. Predicting the potential distribution of the vine mealybug, Planococcus ficus under climate change by MaxEnt[J]. Crop Protection, 2020, 137: 105268. doi: 10.1016/j.cropro.2020.105268 [32] 弓晓敏, 王建军, 白建伟. 基于Maxent和GIS的历山国家级自然保护区原麝栖息地预测[J]. 山西林业科技, 2015, 44(3):7-9,23. doi: 10.3969/j.issn.1007-726X.2015.03.003 [33] Peterson A T, Cohoon K P. Sensitivity of distributional prediction algorithms to geographic data completeness[J]. Ecological Modelling, 1999, 117(1): 159-164. doi: 10.1016/S0304-3800(99)00023-X [34] 张 梅, 禄彩丽, 魏喜喜, 等. 基于MaxEnt模型新疆枣潜在适生区预测[J]. 经济林研究, 2020, 38(1):157-166. [35] 李 昂. 应用ArcGIS软件和最大熵模型分析樟子松潜在分布及其气候适宜性[D]. 沈阳: 沈阳农业大学, 2016. [36] 张文秀, 寇一翾, 张 丽, 等. 采用生态位模拟预测濒危植物白豆杉五个时期的适宜分布区[J]. 生态学杂志, 2020, 39(2):600-613. [37] 张爱平, 王 毅, 熊勤犁, 等. 末次间冰期以来3种云杉属植物的历史分布变迁及避难所[J]. 应用生态学报, 2018, 29(7):2411-2421. [38] 龚 维, 夏 青, 陈红锋, 等. 珍稀濒危植物伯乐树的潜在适生区预测[J]. 华南农业大学学报, 2015, 36(4):98-104. doi: 10.7671/j.issn.1001-411X.2015.04.018 [39] 李 璇, 李 垚, 方炎明. 基于优化的Maxent模型预测白栎在中国的潜在分布区[J]. 林业科学, 2018, 54(8):153-164. doi: 10.11707/j.1001-7488.20180817 [40] 王 绮, 樊保国, 赵光华. 气候变化下毛榛在中国的潜在适生区预测[J]. 生态学杂志, 2020, 39(11):3774-3784. [41] Hewitt G M. Genetic consequences of climatic oscillations in the Quaternary[J]. Philosophical Transactions of the Royal Society B:Biological Sciences, 2004, 359(1442): 183-195. doi: 10.1098/rstb.2003.1388 [42] 胡 菀, 张志勇, 陈陆丹, 等. 末次盛冰期以来观光木的潜在地理分布变迁[J]. 植物生态学报, 2020, 44(1):48-59. [43] 陈 瑜, 倪 健. 利用孢粉记录定量重建大尺度古植被格局[J]. 植物生态学报, 2008, 32(5):1201-1212. doi: 10.3773/j.issn.1005-264x.2008.05.025 [44] N J, Y G, Harrison S P, et al. Palaeovegetation in China during the late Quaternary: Biome reconstructions based on a global scheme of plant functional types[J]. Palaeogeography, 2010, 289(1-4): 44-61. doi: 10.1016/j.palaeo.2010.02.008 [45] 吕 燕. 中国南方亚热带山地10-40 ka BP古植被与古气候特征探讨[D]. 南京: 南京师范大学, 2014. [46] 郑益群, 于 革, 薛 滨, 等. 6kaB. P. 东亚区域气候模拟及其变化机制探讨[J]. 第四纪研究, 2004, 24(1):28-38. doi: 10.3321/j.issn:1001-7410.2004.01.004 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4492
- HTML全文浏览量: 2905
- PDF下载量: 84
- 被引次数: 0
