




-
国槐作为我国的特色树种,树冠硕大,姿态优美,具有极高的园林观赏价值,在城市绿化中发挥着重要作用。国槐带化病是近年国槐苗木繁育及园林景观营造中危害比较严重的一种侵染性病害,其发病后导致国槐生长减缓,枝条扁茎、顶端小叶丛生,并最终发生枯死,不仅给园林生产带来较大的经济损失,而且近年来该病发生日趋严重,有些苗圃中国槐受害率已达30%,严重威胁园林绿化生态效益与观赏价值[1-2]。自上世纪90年代首次证实国槐带化病为植原体侵染造成以来,国内外对其关注较少且研究并不深入。直到近年随着分子检测技术的快速发展,杜银银[3]通过采用PCR快速扩增及生物信息学技术首次明确了国槐带化病植原体的分类地位,并对国槐带化病植原体2个株系(16Sr I-D与16Sr I-B)之间的遗传相关性和亲缘关系进行了鉴定分析。但由于植原体观察及体外培养难度较大[4],因而很难对植原体病害的致病机理及病菌-寄主间互作机制进行深入探究。转录组学研究可以比较精准的反映寄主受到侵染后不同功能基因的响应机制,现已逐渐成为互作解析和差异基因分析的主要技术手段[5]。Fan和Mardi等人通过转录组分析发现,泡桐和墨西哥柠檬在被植原体侵染后,植原体-病菌互作基因表达上调,并且功能注释结果表明植株的相关代谢活动和信号通路主要与植物激素有关[6-7];万倩芸[8-9]等人在对茅苍术扁茎机制进行转录组分析时也发现扁茎症状可能与多种激素代谢通路协同作用相关,并且可能主要与病菌寄生后诱发植物代谢紊乱。而张舒仪等[10]通过对枣疯病植株进行转录组检测及qPCR后期验证后,认为植原体在侵染寄主后可能会迅速激起激素合成及相关保护酶合成基因的表达上升,进而导致激素含量和酶活性代谢紊乱,最终引发丛生、小叶等症状。前人结果表明转录组学与生理功能验证相结合可以较为准确、高效地揭示出植原体病害的致病机理,但目前关于国槐带化病内源激素与转录组学研究尚无报道。
本研究通过运用高效液相色谱对病健组织中的生长素、赤霉素、脱落酸、玉米素含量进行比较分析,并对感病和健康国槐枝条进行转录组测序和对比,结合GO和KEGG功能注释,筛选出可能导致国槐激素代谢紊乱的关键差异基因,旨在通过转录组学与生理水平相结合,揭示出国槐感病枝条扁茎机理。为国槐带化病的防治和抗病品种选育提供理论依据。
-
试验样品在2019年5、8、9、11月中每个月份的20日于保定地区东林水村国槐苗圃园选择6年生国槐进行取样,随机采取6组样品(3株健康树、3株感病树及3株感病树的无症状枝条)用消毒高枝剪剪下树冠顶端2年生的健康与带病枝,并将枝条用无菌剪刀剪成1 cm小段脱去木质部立即放入2 mL离心管,并用标签标记后迅速置于液氮冷冻,分成两组,一组带回实验室于零下20℃~80℃超低温冰箱保存,用于测定激素含量。另一组送转录组高通量测序。
-
参考杜绍华等[11]的HPLC法进行提取、纯化、分离略有改动。称取样品1 g用液氮研磨,加入8 mL 80%的预冷甲醇,4℃避光浸提12 h;避光超声1 h,4℃ 12000 rpm下离心15 min,将上清液转移至10 mL离心管;加入0.2 g PVPP去除酚杂质,超声0.5 h,4℃ 12000 rpm离心5 min,转移上清液到新的离心管中;用冰乙酸调节pH值为2.9,用C18纯化移入10 mL试管,加入1 mL pH=8的甲醇冲洗即进样瓶。试验中以生长素(IAA)、赤霉素(GA)、玉米素(ZT)、脱落酸(ABA)为标样,并建立标样的标准曲线。本试验采用高压液相色谱仪为日本产HI-TACHI L-7420型,流动相为100%甲醇:0.7%乙酸体积比为55∶40∶5,紫外检测仪检测波长为254 nm,流速0.7 mL·min−1,柱温30℃,进样量为10 uL。
-
将国槐韧皮部用液氮进行研磨,用天根生化科技有限公司试剂盒进行总RNA的提取。用琼脂糖凝胶电泳的方法检测总RNA是否存在污染、降解。用Nanodrop检测总RNA的纯度(OD260/280比值)、浓度核酸吸收峰是否正常。用Agilent 2100精确检测RNA的完整性,检测指标包括:RIN值、28S/18S、图谱基线有无上抬、5S峰。样品总RNA检测合格后寄到北京组学生物科技有限公司,进行cDNA文库的构建,文库构建完成之后使用Agilent 2100对文库的insert size进行检测,insert size符合预期后用qPCR方法对文库的有效浓度进行准确定量(文库有效浓度> 2 nM),完成库检。库检合格后,用Illumina测序平台进行测序。
-
对Raw Data进行数据过滤,去除其中的接头序列及低质量Reads获得高质量的CleanData。然后用Quality(Score或Q-score)进行碱基识别,使用Phred的公式(Q-score=−10 log10p)来获得Phred碱基质量值,准确计算出碱基质量值与碱基识别出错的概率的对应关系。碱基质量值越高表明碱基识别越可靠,准确度越高。经过质量控制之后得到的高质量的Clean Data,用一款专门为高通量转录组测序设计的组装软件Trinity[12]对其进行序列组装拼接。以Trinity拼接得到的转录本作为参考序列(Ref),将每个样品的Clean Reads对Ref 做比对分析。并根据基因在不同样品或不同样品组中的表达量进行差异表达分析、差异表达基因功能注释和功能富集等分析。
-
通过对转录组测序所得内源激素相关数据进行筛选差异基因,采用SYBR法对筛选的差异基因进行实时荧光定量PCR验证。并以Actin 作为内参基因。Actin-F(5′- GAGGGCCCAGGTTCTGATTT-3′)和 Actin-R(5′-GCACCCACCTGCATTTTACC-3′)。通过NCBI设计引物并进行荧光定量分析。实时荧光定量PCR按照以下步骤进行扩增:95℃预变性3 min, 95℃变性5 s, 72℃退火30 s,40个循环。按照2−ΔΔCT[13] 计算基因相对表达量。用 SPSS20.0 对数据进行单因素方差分析(ANOVA),采用LSD 和 Duncan’s 方法进行两两比较、差异显著性分析(P < 0.05)。
-
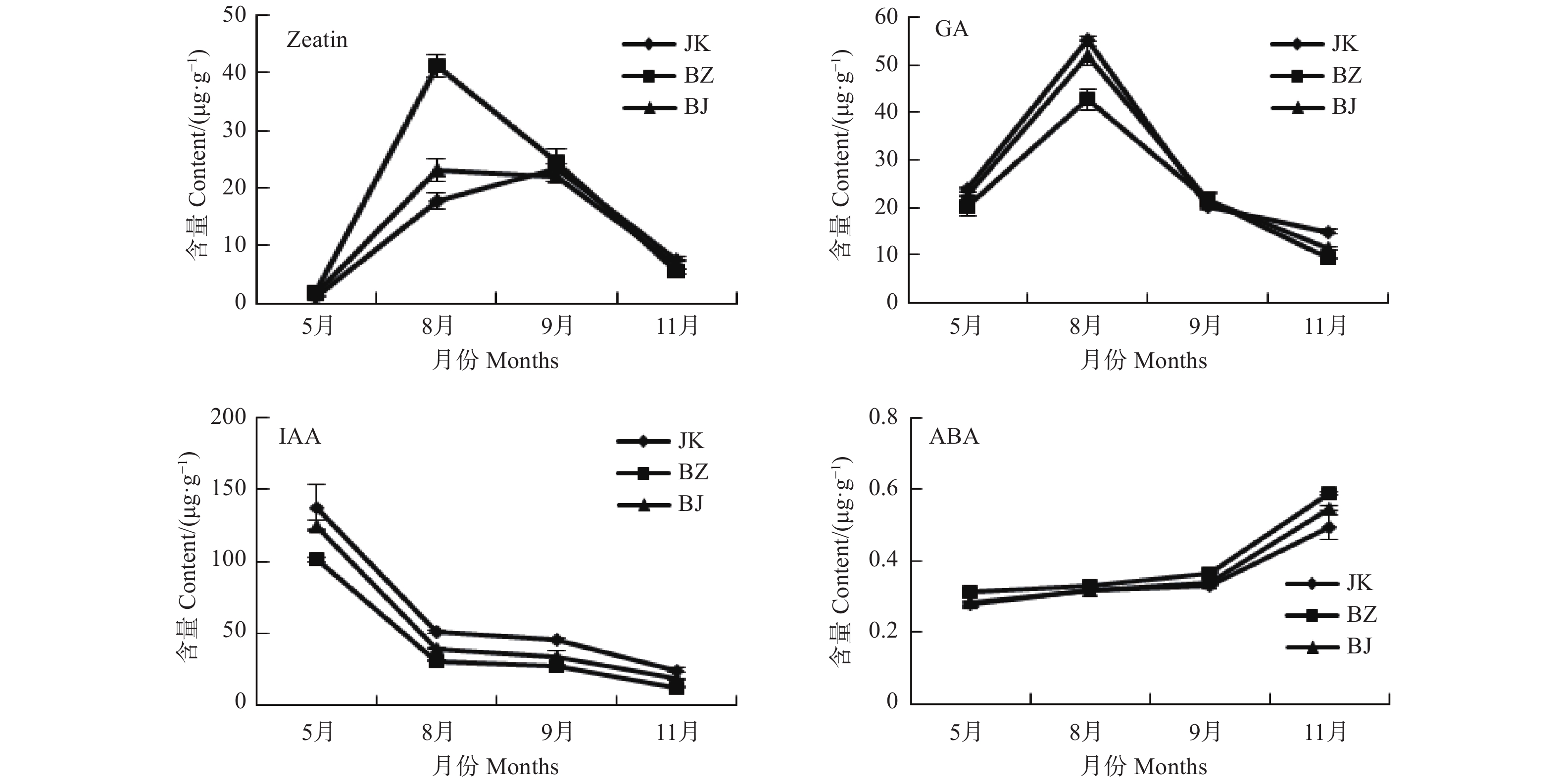
通过对处于生长期的感病国槐(BZ)、健康国槐(JK)和感病国槐树上的表象健康枝(BJ)树皮的内源激素含量的测定,研究该病害发生期对国槐内源激素的影响。由图1可以看出,BZ、BJ和JK间的Zeatin、GA、IAA及ABA含量变化趋势基本一致。Zeatin含量在8、9月份达到高峰,之后呈下降趋势,其中8月份BZ中的Zeatin含量显著高于BJ和JK,BJ、JK无差异。其它月份中三者间无显著差异;GA含量与Zeatin含量年变化较为相似,在8月份达到最高,之后处于下降趋势。但其中8月份BZ的GA含量显著低于JK和BJ,BJ其次,BZ中的GA含量最低,其它月份三者间并无差异;而IAA和ABA的含量变化呈相反趋势,IAA除5月份最高外,之后一直呈下降趋势,并在11月份达到最低,其中JK含量最高,BJ其次,BZ中的IAA含量最低。ABA则是从5月份开始一直呈上升趋势,在11月份达到最高,BZ中的含量显著高于JK和BJ,BJ与JK的ABA含量在5、8、9月份中均无差异。
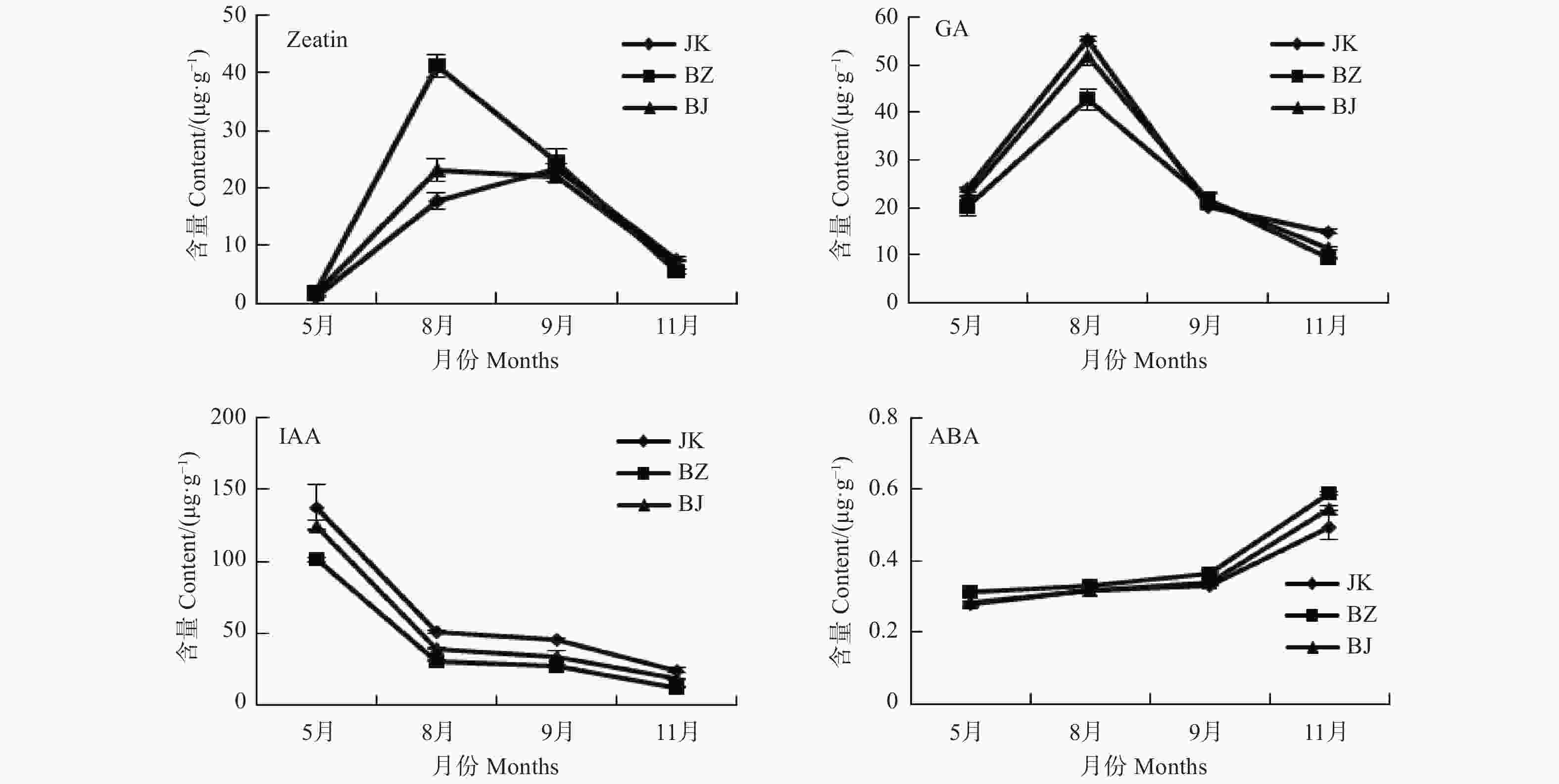
图 1 病健国槐内源激素含量
Figure 1. Content of endogenous hormones in the diseased and heathy
-
通过对国槐感病样本(BZ1、BZ2、BZ3)和健康样本(JK1、JK2、JK3)进行高通量转录组测序,分别获得26499450、22444628、33026072、37221613、28464069和27856864条Raw Reads,在去除含有接头的Reads以及低质量的Reads(包括去除N的比例大于10%的Reads)后,最终得到20427416、17402101、25556038、28443464、21878012和21192357条Clean reads。其中Q20碱基百分比分别是98.20%、98.16%、98.05%、97.95%、97.93%和97.99%,GC碱基含量则均约达到44%(表1)。经过过滤后,最终得到可靠的转录本数据,并进行下一步生物学分析。
表 1 每个样品有效测序数据统计
Table 1. Statistics of valid sequencing data of each sample
SampleID Raw Reads Clean reads Q20/% GC/% BZ1 26499450 20427416 98.20 44.22 BZ2 22444628 17402101 98.16 44.47 BZ3 33026072 25556038 98.05 44.69 JK1 37221613 28443464 97.95 44.26 JK2 28464069 21878012 97.93 44.13 JK3 27856864 21192357 97.99 44.26 -
采用BLAST[14]软件对Unigene序列与NR、Swiss-Prot[15]、GO[16]、COG/KOG[17]、KEGG[18]数据库进行比对,共获得Unigne注释功能信息有72637条,占全部Unigene的60%,其中NR数据库中注释的信息最多,而COG数据库中被注释信息最少。通过使用KOBAS2.0[15]获得Unigene在KEGG中的KEGG Orthology结果,在预测Unigene的氨基酸序列之后使用HMMER[17]软件与Pfam[19]数据库比对,获得Unigene的注释信息(表2)。
表 2 Unigene功能注释统计
Table 2. Unigene function annotation statistics
Anno_Database Annotated_Number 300<=length<1000 length>=1000 COG 19618 5730 5909 GO 38153 17363 12172 KEGG 42076 19494 12612 KOG 35012 15753 10062 Pfam 32551 10775 13098 Swissprot 47071 19687 13780 nr 67685 31769 17211 All 72637 32877 17313 -
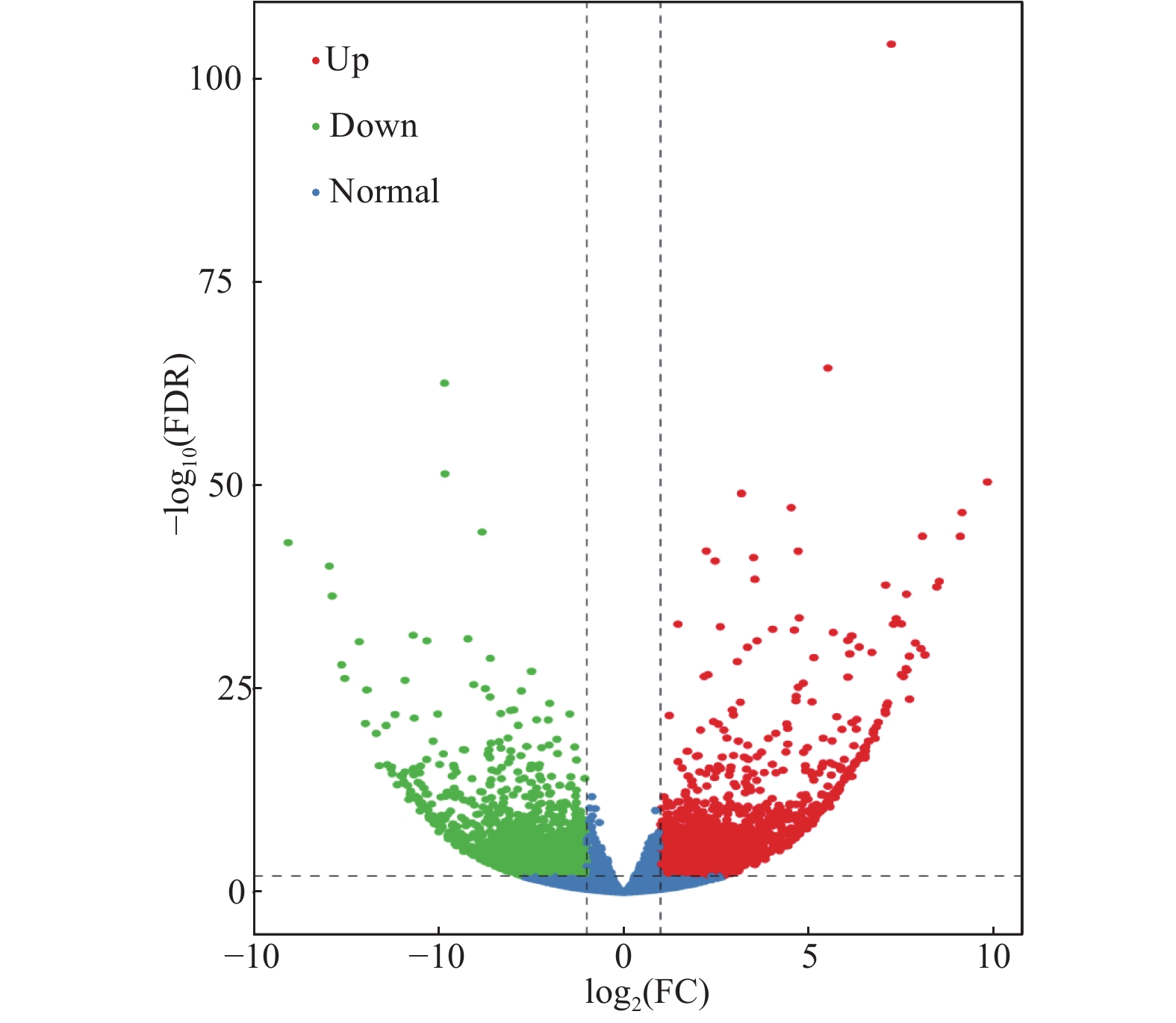
通过采用EBSeq[20]对所得数据进行差异分析,并采取Fold Change≥2且FDR<0.01作为筛选标准。如图2所示,绿点代表下调差异表达基因,红点代表上调差异表达基因,蓝色的点代表非差异表达基因。其中共得到差异表达基因数目4714条,国槐带化病样品相对于健康国槐样品上调基因有2283条,下调基因有2431条。这些结果表明,国槐在受到植原体侵染后其植物体内多数的基因表达受到抑制。
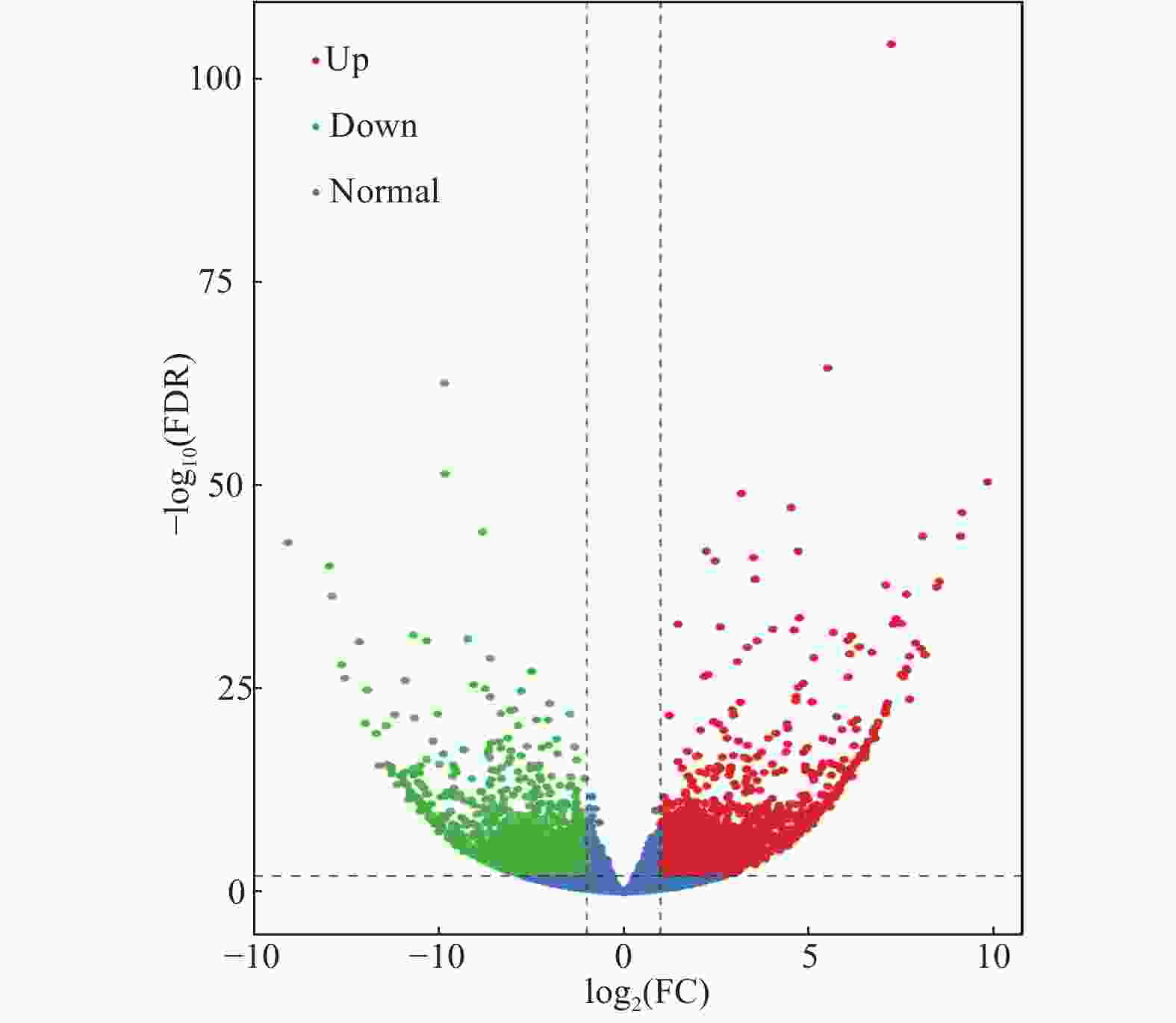
图 2 国槐带化病株和健康株之间差异表达基因的火山图
Figure 2. Volcanic diagram of differentally expressed genes between locust striated and healthy plants
-
通过对感病与健康国槐转录组差异表达基因进行GO功能注释富集分析,结果可分为3大类:细胞组成(cellular component)、分子功能(molecular function)、生物学过程(biological process)。其中注释到细胞组成有7个,分子功能10个,生物学过程3个(图3)。细胞组成中以膜的组成成分(783个)和质膜(141)中差异表达基因最多,另外膜的组成成分、运动蛋白复合物等离子组分以及细胞壁等差异表达基因富集表达均达到显著水平;分子功能中以蛋白丝氨酸苏氨酸激酶(136个)和DNA结合转录(84个)差异表达基因最多,而在木糖葡聚糖、木糖葡聚糖转化和DNA结合转录等富集基因中达到显著水平;生物学过程中蛋白磷酸化中的差异表达基因居多并达到显著水平。结果表明细胞组成和分子功能的基因富集更为显著,这说明在国槐受到植原体侵染以后,细胞和分子功能方面均发生明显的差异变化,并可能受到上述富集差异基因的调控。

图 3 差异表达基因GO富集分析
Figure 3. GO enrichment analysis of differentially expressed genes
-
在生物体内,不同的基因产物相互协调来行使生物学功能,对差异表达基因的通路(Pathway)注释分析有助于进一步解读基因的功能。通过对差异基因(2539个)进行KEGG功能注释分析,发现差异基因被富集到48条代谢通路,本研究挑选了富集最显著的20条pathway条目在图中进行展示(图4)。其中代谢通路从低到高依次为:戊糖和葡萄糖醛酸互相转化、卟啉和叶绿素代谢、AMPK信号通路、癌变中的微型核糖核酸、多巴胺能受体、半光氨酸和蛋氨酸代谢、泛素介导的蛋白水解、MAPK信号通路、溶酶体、卵母细胞减数分裂、RNA降解、抗原加工和传递、苯丙氨酸合成、Ras信号通路、内噬作用、植物激素信号转导、植物-病原互作、淀粉和蔗糖代谢、Toll受体信号转导途径和NF-KB信号传导途径。其中,汇集差异基因最多的Toll受体信号转导途径和NF-KB信号传导途径均表现为显著下调,另外,控制细胞内噬作用、减数分裂以及抗原加工与传递同样表现为显著下调。
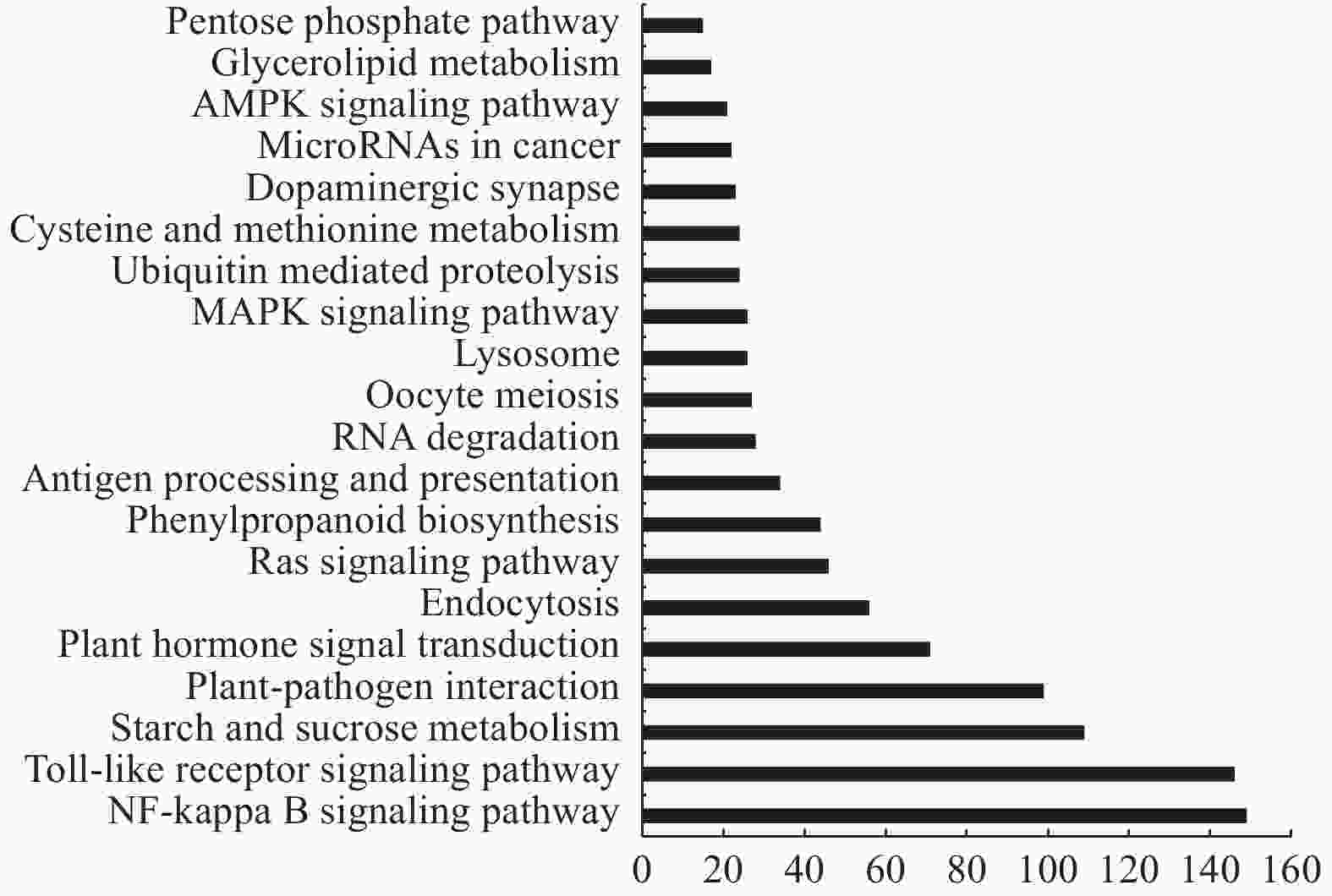
图 4 差异表达基因KEGG富集
Figure 4. Distribution of differentially expressed gene KEGG
-
感病国槐和正常国槐激素代谢通路的差异基因被注释到25个DEGs(图5)。由图可知,其中有7个下调基因(绿色标注),8个上调基因(红色标注)。其中,在色氨酸代谢通路上(生长素),AUX1节点上有3个差异表达基因下调,而在GH3节点有3个差异表达基因上调。而在玉米素信号传导代谢通路上(细胞分裂素),CRE1基因节点上涉及到7个差异基因下调;AHP节点上有3个差异基因下调;ARR-A节点处有3个差异基因下调。二萜生物合成(赤霉素)代谢通路上关键调控基因TF涉及13个差异基因上调。生长素、玉米素合成关键基因的下调以及赤霉素合成基因的上调,可能是导致国槐枝条顶端生长停滞、发生横向卷曲生长的主要原因。类胡萝卜素生物合成(脱落酸)代谢途径中,ABF调控关键基因涉及10个差异基因上调,产生这种原因可能与植原体的致病因子的基因干扰或植物体受到胁迫产生的自激反应有关。

图 5 植物激素传导(Ko04075)路径
Figure 5. Pathway map of phytohormone transmission (ko04075)
-
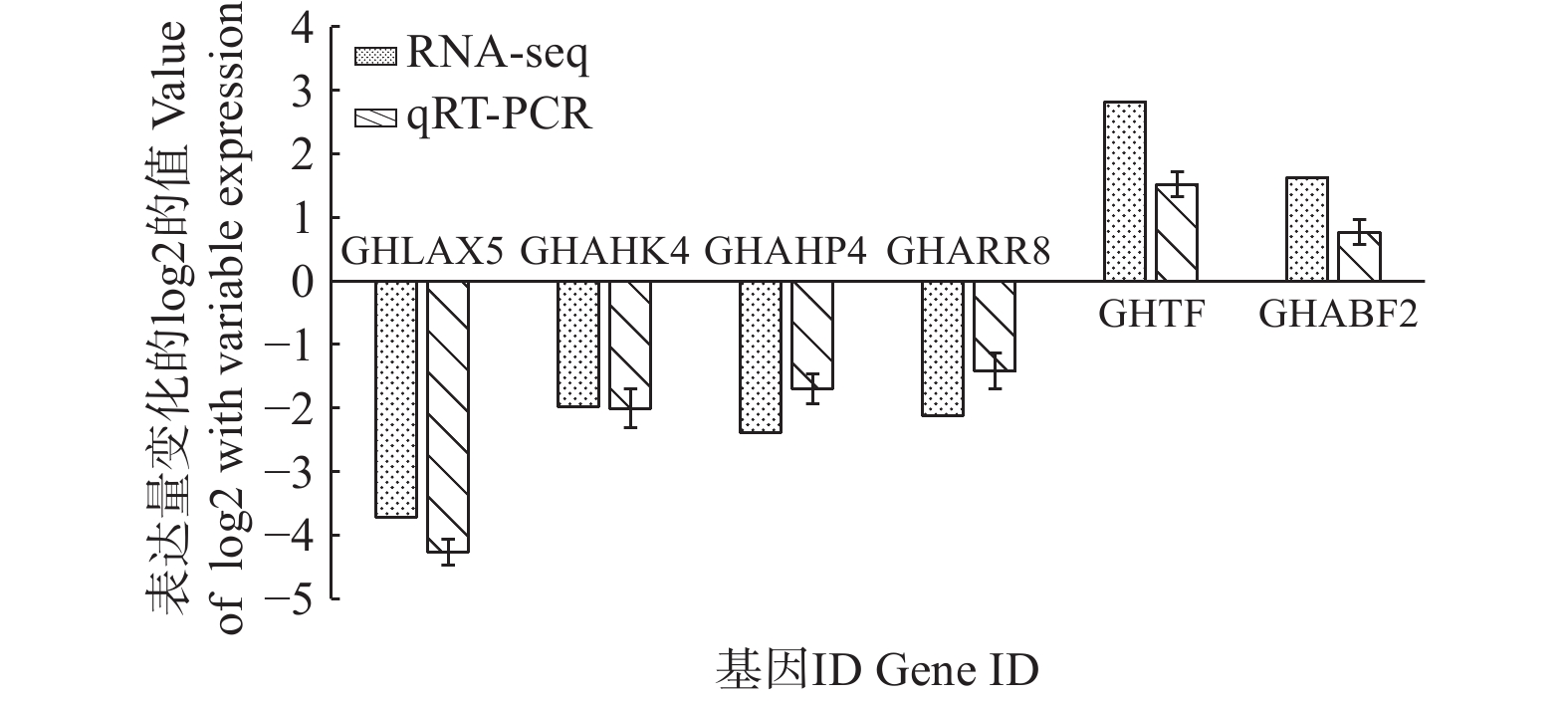
qRT-PCR验证本研究通过对转录组差异基因数据进行筛选获得与生长素、细胞分裂素、赤霉素和脱落酸代谢相关基因共6个(表3),其中,与生长素代谢相关基因AUX 1个(GHLAX5),与细胞分裂素信号传导受体CRE1相关基因3个(GHAHK4、GHAHP4和GHARR8),调控赤霉素信号传导的相关基因TF 1个(GHTF),脱落酸信号转导调控因子相关基因ABF 1个(GHABF2)。通过对上述6个激素代谢关键基因进行荧光定量表达验证,如图6所示,虽然定量表达与转录组结果具有一定差异,但表达趋势较为一致,生长素、细胞分裂素代谢关键基因表现为显著下调,赤霉素和脱落酸代谢关键基因表现为显著上调。
表 3 差异表达基因qRT-PCR 引物
Table 3. Primers of differentially expressed genes QRT-PCR
基因名称
Gene name序列 ID
Sequence ID上游引物
Forward sequence(5′→ 3′)下游引物
Reverse sequence(3′→ 5′)GHLAX5 DN122098_c0_g1 TCTCCCAACCTTCGAGAACC AATGCTAGGTGGGTGTGGAG GHAHK4 DN95401_c0_g1 CGCGGATGAAGAAAAGTGGTC CCAATTCCAGTGTCCTCCACA GHAHP4 DN115292_c1_g2 TGTTGGTCCGTTTGGGGATA AAGTGGAAGAGCAAGATTCCTACA GHARR8 DN107272_c1_g2 TGTTGGTCCGTTTGGGGATA AAGTGGAAGAGCAAGATTCCTACA GHTF DN111522_c0_g1 TCCCTTCCCATTTTTCCAGCA TTTTGTCTTGCTTCCACGCTC GHABF2 DN119624_c4_g2 GAGTTCTGAGGCCGTCCATT GAGCATACGCAATCTGCACT 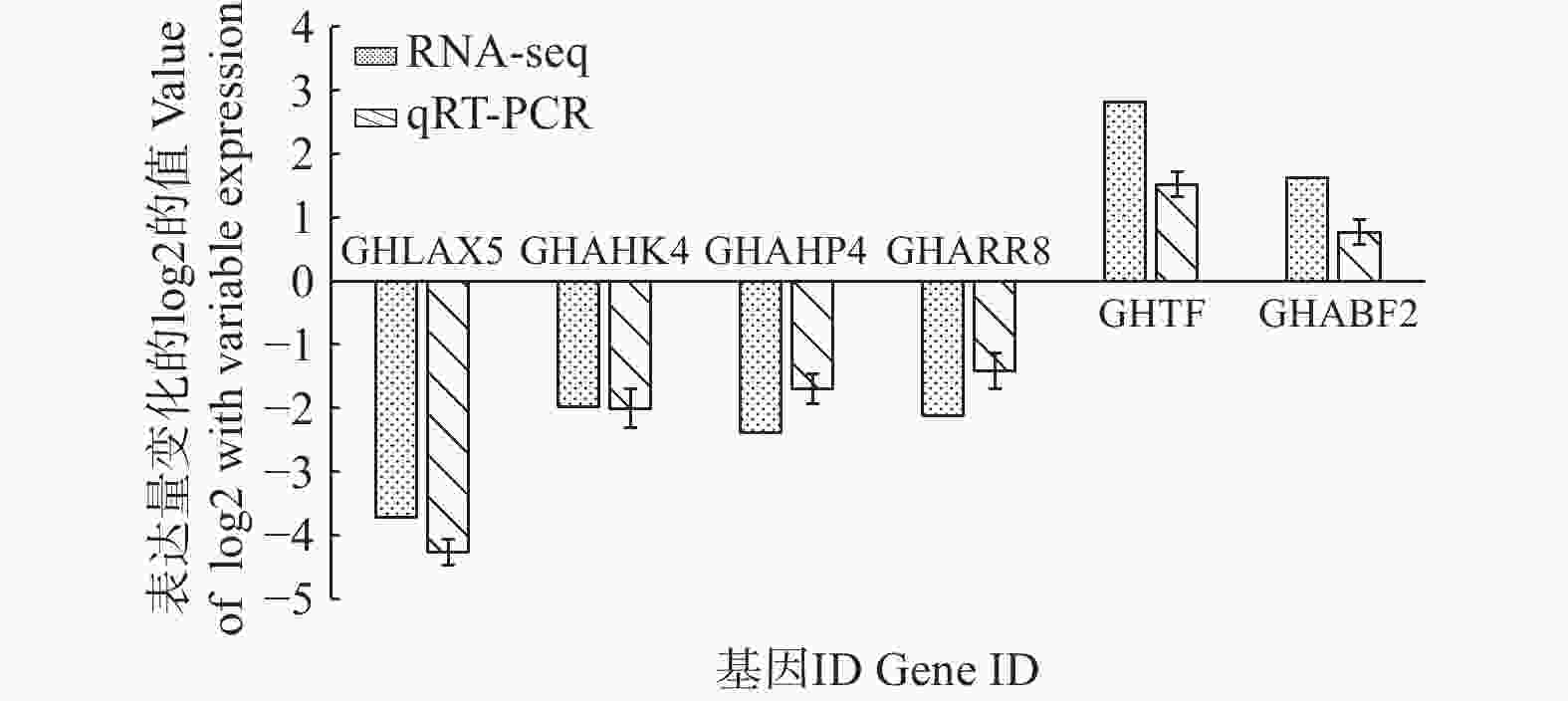
图 6 差异表达基因的qRT-PCR 验证
Figure 6. Validation of differentially expressed genes using qRT-PCR
-
多数研究认为,植原体在侵入植物体后,产生的某种产物会导致植物体内激素失衡,进而导致植物体表型生长发生改变,出现小叶、丛生等症状[21-22]。因而通过研究病株机体内源激素含量进而解释植原体病害的致病机理逐渐成为植原体病害研究的主要手段。赵锦等[23]研究发现枣疯病株在1年中的发育旺盛期内玉米素含量表现为异常显著升高,而且在生长后期内玉米素含量也显著高于健株。Kesumawati[24]的研究结果也表明在八仙花受到植原体感染的组织中含有较高含量的玉米素。另外,有研究认为玉米素和生长素的比例变化失调可能是导致植物体发生小叶、丛枝等病状的主要因素[23,25-26]。本研究发现国槐在受到植原体侵染以后,其生长旺盛的8月份期间,病枝内玉米素显著高于健树和病树上的无症状枝,而在其他月份并未产生较大差异;但在旺盛生长期内病枝中的赤霉素、生长素含量则均显著低于健树和病树上的无症状枝,在旺盛生长期内的玉米素含量与生长素比例的严重失衡可能是导致国槐感病枝条发生小叶和丛枝的主要原因,与前人得出的玉米素含量的显著增加是导致器官分化方向改变,进而引起丛枝的结果较为一致[23]。而且生长素、赤霉素的显著降低也可能是导致国槐病枝向内卷曲和停止伸长生长的主要原因[27]。
植原体的功能基因组较小,在侵染寄主后,其基因组片段会整合到植物的细胞基因组中,研究发现植原体基因组缺乏脂肪酸和氨基酸合成等基本的生化途径[23],因而可能主要通过基因表达调控帮助其完成寄生,并会对寄主植物产生影响。有研究发现,植原体的简单基因组中存在编码致病效应因子蛋白的基因,部分致病效应因子通过与标靶蛋白相结合干扰寄主植物的正常代谢以达到干扰寄主植物防御功能的目的[4],翠菊黄化植原体丛枝株系所产生的致病效应蛋白SAP11AYWB能够通过靶标作用寄主的TCP转录因子,进而影响寄主植物体内的抗体活性甚至使得该类转录因子失活,导致丛枝症状的产生[28]。本研究通过转录组测序发现国槐带化病枝条内的多数基因表达受到抑制,细胞组分与分子功能富集差异基因最多,并且差异基因多数注释到植物的免疫应答机制、植物-病原互作和激素信号转导等方面,其中Toll受体信号转导途径和NF-KB信号传导途径汇集差异基因最多,且均表现为显著下调,有研究指出Toll受体在动物体内主要通过识别来自细菌和病毒的侵入信号及细胞损伤状况,进而对免疫应答起到关键调控作用[29],而NF-KB信号通路作为Toll受体信号传导激活的下游通路,常与Toll受体在免疫机制中形成联控作用[30],这说明国槐枝条在受到植原体侵染以后,机体调控免疫应激的能力显著降低,细胞衰亡程度加快。另外,转录组结果表明在激素代谢通路中调控生长素代谢的响应因子AUX1的表达显著下调,这与前人得到的在表达植原体致病蛋白“天狗蛋白”的植物中生长素响应因子显著下调,且生长素含量显著降低的结果较为一致[31]。同样,Jennifer研究表明拟南芥组氨酸激酶(CRE)作为一种玉米素受体,可能在玉米素功能信号传导中发挥着关键作用[32],而Riefler等人研究结果也指出植物体内的玉米素含量可能受到CRE受体的反馈抑制,在CRE受体突变的植株中积累了更高含量的细胞分裂素[33],本研究结果表明在玉米素代谢通路中,CRE1受体基因表达显著下调,另外负调控细胞分裂发育的A型ARRs基因[32]表达也显著下调,这与前人报道称泡桐丛枝病枝发病后,病枝内玉米素代谢通路基因表达显著上调[34]的研究结果相反,但这些结果说明国槐带化病枝内积累的较高含量细胞分裂素以及细胞增生进而发生横向生长可能与CRE1和A型ARRs等基因的显著下调表达有关,具体原因则需要对这两个基因进行进一步功能验证。另外本研究通过对转录组筛选得到的生长素、玉米素、赤霉素和脱落酸代谢通路中的关键基因进行荧光定量表达验证,结果表明表达趋势与转录组数据较为一致,这说明本研究所进行的转录组验证准确性较高。综上,可能正是由于国槐病株内源激素代谢关键基因表达显著上调或下调,进而导致生长素、玉米素、赤霉素和脱落酸的含量积累发生变化,这与前人在对筛选得到的关键基因进行定量表达验证后[10,34]所得到的结果一致。
-
本研究通过探究国槐带化病株内激素含量与基因表达之间的变化联系,认为国槐植株在受到病原侵染以后,其病枝内玉米素、生长素和赤霉素的代谢关键调控基因表达显著上调或下调,进而导致病枝内的玉米素和生长素比例失衡,以及赤霉素显著降低,这可能是导致国槐病枝表现丛生、小叶及枝条带化生长的主要原因。本工作为进一步深入研究国槐带化病及其防治工作提供了理论基础。
感染国槐带化病植株内源激素及转录组分析
Analysis of Endogenous Hormones and Transcriptome in Sophora japonica Infected with Fasciation Disease
-
摘要:
目的 为探究国槐带化病的致病机理,进而为国槐带化病的防治研究提供理论依据。 方法 以国槐健康与感病枝条作为研究材料,对其生长期间的5、8、9及11月份中枝条生长素、玉米素、赤霉素和脱落酸含量进行连续测定,通过选取8月份国槐的病、健组织进行转录组测序筛选差异表达基因,并进行qPCR验证分析。 结果 在5、8、9、11月中,健康、感病枝条及感病树上的健康枝条中的生长素、玉米素、赤霉素和脱落酸含量变化趋势基本一致,但在8月份国槐旺盛生长期间,感病枝条内的玉米素含量显著高于健康对照;生长素和赤霉素含量显著低于健康对照。而在11月份植株的生长后期,仅感病植株脱落酸含量显著高于健康对照。转录组测序结果表明差异基因下调表达明显高于上调,并且差异基因GO功能注释分析显示,差异基因主要汇集在细胞组分、分子功能和生物学过程3个方面,其中以细胞组分和分子功能差异基因富集更为显著;而KEGG代谢通路分析表明,感病植株的差异基因主要汇集在调控植株免疫应答的信号传导、植物-病原互作和植物激素信号传导等方面,并且其中调控植株免疫应答的信号传导富集差异基因最多且表现为显著下调;内源激素代谢通路注释结果表明,生长素、玉米素受体基因AUX1和CRE表达显著下调,而调控赤霉素、脱落酸代谢的基因TF和ABF表达显著上调;通过对转录组数据进行筛选获得调节生长素、玉米素、赤霉素和脱落酸代谢通路代谢及信号转导的相关基因(GHLAX5、GHAHK4、GHAHP4、GHARR8、GHTF和GHABF2)进行qPCR验证,结果虽与转录组数据存在一定差异,但表达趋势基本一致,这些结果表明,上述基因的差异表达可能对于调控国槐内源激素含量变化具有重要作用。 结论 认为玉米素含量的显著上升和生长素、赤霉素含量的显著降低,阻碍植株的免疫机制应答并调控这3种激素代谢的关键基因差异表达,可能是导致国槐带化病枝条发病且横向生长形成扁茎的主要原因。 Abstract:Objective To investigate the pathogenesis of banding disease of Sophora japonica, so as to provide data for the prevention and control of the disease. Method In this study, the contents of auxin, zeatin, gibberellin and abscisic acid in the healthy and diseased branches of Sophora japonica were determined continuously in May, August, September and November. The differentially expressed genes were screened by transcriptome sequencing and the qPCR analysis was performed. Result In May, August, September and November, the contents of auxin, zeatin, gibberellin and abscisic acid in healthy tree, infected and symptomless shoots of the diseased trees were basically the same, but during the vigorous growth period of S. japonica in August, the content of zeatin in the infected branches was significantly higher than that in the healthy control; the contents of auxin and gibberellin were significantly lower than that of the healthy control. However, in the late growth stage of the plants in November, only the abscisic acid content of the diseased plants was significantly higher than that of the healthy control. Transcriptome sequencing results showed that the down-regulation of differential genes was significantly higher than up-regulation, and GO functional annotation analysis of differential genes showed that the differential genes were mainly concentrated in cell component, molecular function and biological process, especially in cell component and molecular function, while the KEGG metabolic pathway analysis showed that the differentially expressed genes in susceptible plants were mainly concentrated in signal transduction, plant pathogen interaction and plant hormone signal transduction. The results of endogenous hormone metabolic pathway annotation showed that AUX1 and CRE were down regulated, while the TF and ABF were up-regulated. The genes (GHLAX5, GHAHK4, GHAHP4,GHARR8, GHTF and GHABF2) regulating auxin, zeatin, gibberellin and abscisic acid metabolism and signal transduction were obtained by screening the transcriptome data for qPCR verification. Although there were some differences between the results and the transcriptome data, the expression trend was basically the same. These results suggested that the differential expression of these genes may play an important role in regulating the endogenous hormone content of S. japonica. Conclusion The significant increase of zeatin content and the significant decrease of auxin and gibberellin contents may hinder the immune response of plants and the differential expression of key genes regulating the metabolism of these three hormones, which may be the main reason for the occurrence and lateral growth of the diseased branches of S. japonica. -
Key words:
- Sophora japonica
- / fasciation disease
- / pathogenesis
- / endogenous hormones
- / transcriptome
-
图 2 国槐带化病株和健康株之间差异表达基因的火山图
Figure 2. Volcanic diagram of differentally expressed genes between locust striated and healthy plants
图 6 差异表达基因的qRT-PCR 验证
Figure 6. Validation of differentially expressed genes using qRT-PCR
表 1 每个样品有效测序数据统计
Table 1. Statistics of valid sequencing data of each sample
SampleID Raw Reads Clean reads Q20/% GC/% BZ1 26499450 20427416 98.20 44.22 BZ2 22444628 17402101 98.16 44.47 BZ3 33026072 25556038 98.05 44.69 JK1 37221613 28443464 97.95 44.26 JK2 28464069 21878012 97.93 44.13 JK3 27856864 21192357 97.99 44.26  下载: 导出CSV
下载: 导出CSV
表 2 Unigene功能注释统计
Table 2. Unigene function annotation statistics
Anno_Database Annotated_Number 300<=length<1000 length>=1000 COG 19618 5730 5909 GO 38153 17363 12172 KEGG 42076 19494 12612 KOG 35012 15753 10062 Pfam 32551 10775 13098 Swissprot 47071 19687 13780 nr 67685 31769 17211 All 72637 32877 17313
下载: 导出CSV
表 3 差异表达基因qRT-PCR 引物
Table 3. Primers of differentially expressed genes QRT-PCR
基因名称
Gene name序列 ID
Sequence ID上游引物
Forward sequence(5′→ 3′)下游引物
Reverse sequence(3′→ 5′)GHLAX5 DN122098_c0_g1 TCTCCCAACCTTCGAGAACC AATGCTAGGTGGGTGTGGAG GHAHK4 DN95401_c0_g1 CGCGGATGAAGAAAAGTGGTC CCAATTCCAGTGTCCTCCACA GHAHP4 DN115292_c1_g2 TGTTGGTCCGTTTGGGGATA AAGTGGAAGAGCAAGATTCCTACA GHARR8 DN107272_c1_g2 TGTTGGTCCGTTTGGGGATA AAGTGGAAGAGCAAGATTCCTACA GHTF DN111522_c0_g1 TCCCTTCCCATTTTTCCAGCA TTTTGTCTTGCTTCCACGCTC GHABF2 DN119624_c4_g2 GAGTTCTGAGGCCGTCCATT GAGCATACGCAATCTGCACT
下载: 导出CSV
-
[1] Duduk B, Tian J, Contaldo N, et al. Occurrence of phytoplasmas related to stolbur and to ‘Candidatus Phytoplasmajaponicum’ in woody host plants in China[J]. J Phytopathol, 2010, 158: 100-104. doi: 10.1111/j.1439-0434.2009.01586.x [2] Yu Z C, Cao Y, Zhang Q, et al. ‘Candidatus Phytoplasmaziziphi’ associated with Sophora japonica withes’broom disease in China[J]. J Gen Plant Pathol, 2012, 78: 298-300. doi: 10.1007/s10327-012-0385-7 [3] 杜银银. 国槐和紫薇带化病植原体的分子鉴定[D]. 杨凌: 西北农林科技大学, 2014. [4] 李继东, 陈 鹏, 倪 静, 等. 植原体致病分子机理研究进展[J]. 园艺学报, 2019, 46(9):1691-1700. [5] Wang Z, Gerstein M, Snyder M. RNA-Seq: a revolutionary tool for transcriptomics[J]. Nature Reviews. Genetics, 2009, 10(1): 57. doi: 10.1038/nrg2484 [6] Fan G. G, Cao X. B, Zhao Z. L, et al. Transcriptome analysis of the genes related to the morphological changes of Paulownia tomentosa plantlets infected with phytoplasma[J]. Acta Physiologiae Plantarum, 2015, 37(10): 1-12. [7] Mardi M, Farsad L. K, Gharechahi J, et al. Indepthtranscriptome sequencing of mexican lime trees infected with candidatus Phytoplasma aurantifolia[J]. PloS One, 10(7):e, 0130: 425. [8] 万倩芸, 陈丽娜, 邓 娟, 等. 植物扁茎形成机制研究进展[J]. 分子植物育种, 2017, 15(9):3717-3722. [9] 万倩芸. 基于转录组学的茅苍术扁茎变异机制研究[D]. 武汉: 湖北中医药大学, 2019. [10] 张舒怡, 张 钟, 张春梅, 等. 基于转录组水平的枣疯病发病机理研究[J]. 园艺学报, 2017, 44(7):1287-1298. [11] 杜绍华, 卜志国, 刘 洋. 植原体浸染对枣树内源激素含量的影响[J]. 北方园艺, 2013(13):12-15. [12] Grabherr M G, Haas B J, Yassour M, et al. Trinity: reconstructing a full-length transcriptome without a genome from RNA-Seqdata[J]. Nature Biotechnology, 2011, 29(7): 644-652. doi: 10.1038/nbt.1883 [13] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔ CT method[J]. Methods, 2001, 25(4): 402-408. doi: 10.1006/meth.2001.1262 [14] Altschul S F, Madden T L, Zhang J, et al. Gapped BLAST and PSI BLAST: A New Generation of Protein Database Search Programs[J]. Nucleic Acids Research, 1997, 25(17): 3389-3402. doi: 10.1093/nar/25.17.3389 [15] Apweiler R, Bairoch A, Wu C H, et al. UniProt: the universal protein knowledgebase[J]. Nucleic Acids Research, 2004, 32: 115-119. doi: 10.1093/nar/gkh151 [16] Xie C, Mao X, Huang J, et al. KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases[J]. Nucleic Acids Research, 2011, 39(WebServerissue): 316-322. [17] Finn R D, Coggill P, Eberhardt R Y, et al. Nucleic Acids Research[J]. Database Issue, 2016, 44: 279-285. [18] Tatusov R L, Galperin M Y, Natale D A. The COG database: a tool for genome scale analysis of protein functions and evolution[J]. Nucleic Acids Research., 2000, 28(1): 33-36. doi: 10.1093/nar/28.1.33 [19] Kanehisa M, Goto S, Kawashima S, Okuno Y, et al. The KEGG resource for deciphering the genome[J]. Nucleic Acids Research, 2004, 32: 277-280. doi: 10.1093/nar/gkh063 [20] Leng N, Dawson J A, Thomson J A, et al. EBSeq: an empirical Bayes hierarchical model for inference in RNA-seqexperiments[J]. Bioinformatics., 2013, 29(8): 1035-1043. doi: 10.1093/bioinformatics/btt087 [21] Bertamini M, Grando M S, Muthuchelian K, et al. Effect of phytoplasmal infection on photosystem II efficiency and thylakoid membrane protein changes in field grown apple (Maluspumila) leaves[J]. Physiological & Molecular Plant Pathology, 2002, 61(6): 349-356. [22] MirnaĆ P, Hrvoje L, Martina Š M. Effect of indole-3-butyric acid on phytoplasmas in infected Catharanthusroseus shoots grown in vitro[J]. FEMS Microbiology Letters, 2007, 268: 171-177. doi: 10.1111/j.1574-6968.2006.00577.x [23] 赵 锦, 刘孟军, 代 丽, 等. 枣疯病病树中内源激素的变化研究[J]. 中国农业科学, 2006, 39(11):2255-2255. doi: 10.3321/j.issn:0578-1752.2006.11.014 [24] Kesumawati E, Kimata T, Uemachi T, et al. Correlation of phytoplasma concentration in Hydrangea macrophylla with green-flowering stability[J]. Entia Horticulturae, 2006, 108(1): 74-78. [25] 何放亭, 戴 群. C/A值与甘薯丛枝病症状发生的关系[J]. 植物病理学报, 1997, 27(1):43-46. doi: 10.3321/j.issn:0412-0914.1997.01.012 [26] 田国忠, 黄钦才, 袁巧平, 等. 感染MLO泡桐组培苗代谢变化与致病机理的关系[J]. 中国科学(B辑化学生命科学地学), 1994(5):484-490. [27] 李合生. 现代植物生理学: Modern Plant Physiology[M]. 北京: 高等教育出版社, 2012. [28] Sugio A, Maclean A M, Kingdom H N, et al. Diverse Targets of Phytoplasma Effectors: From Plant Development to Defense Against Insects[J]. Annual Review of Phytopathology, 2011, 49(1): 175. doi: 10.1146/annurev-phyto-072910-095323 [29] 鲍璐璐, 崔立红. TLR4/MyD88/NF-κB信号通路的研究进展[J]. 胃肠病学和肝病学杂志, 2019, 28(5):568-572. doi: 10.3969/j.issn.1006-5709.2019.05.019 [30] 何金鑫, 黄丽华. LPS-TLR4/NF-kB信号通路与抗炎免疫研究进展[J]. 中外健康文摘, 2013(37). [31] Minato N, Himeno M, Hoshi A, et al. The phytoplasmal virulence factor TENGU causes plant sterility by downregulating of the jasmonic acid and auxinpathways[J]. Scientific Reports, 2014, 4: 7399. [32] To J P C, Kieber J J. Cytokinin signaling: two-components and more[J]. Trends Plant, 2008, 13(2): 85-92. doi: 10.1016/j.tplants.2007.11.005 [33] Michael R, Ondrej N, Miroslav S, et al. Arabidopsis Cytokinin Receptor Mutants Reveal Functions in Shoot Growth, Leaf Senescence, Seed Size, Germination, Root Development, and Cytokinin Metabolism[J]. The Plant Cell, 2006, 18(1): 40-54. [34] 韩良泽, 陈明利, 陈永华, 等. 泡桐根系应答铅锌矿胁迫的转录组分析[J]. 中南林业科技大学学报, 2020, 40(8):128-138. -

 点击查看大图
点击查看大图
图(6) / 表(3)
计量
- 文章访问数: 3441
- HTML全文浏览量: 1607
- PDF下载量: 55
- 被引次数: 0
