


-
乔木作为森林生态系统的主体,其物种多样性是表征群落组成结构和稳定性的重要指标,越高的物种多样性越有利于形成稳定有序的耗散结构,在森林生态系统维持稳定发展过程中更具生态学意义[1-4]。研究尺度的改变影响着生境异质性,异质性随着研究尺度增加而增加,意味着不同的尺度存在更加多样的小生境,进而影响群落的物种多样性[5-6]。
森林结构是林分中各林木个体及其属性之间相互关系的体现[7],通过空间结构与非空间结构两大方面全面反映林分生长状态[8],进而阐述林分的发展潜力[9]。空间结构主要反映林木水平分布格局、物种混合度及林木大小差异,与林木的位置和个体属性密切相关[10]。基于参照树与最近4株相邻木构成空间结构单元[7]的林分空间结构参数主要为角尺度[11-14]、大小比数[15]、混交度[16-18]和密集度。研究尺度对于林分空间结构有较大影响,小尺度上林分空间结构变化较大,研究尺度上升的同时空间结构逐步趋稳[19]。
湖北省西南部山区(鄂西南)典型亚热带常绿落叶阔叶混交林中物种丰富,结构稳定,对维持该区域的生态系统稳定性以及实现森林可持续发展有重要意义[20]。然而该区作为喀斯特地貌典型地区,不同尺度上生境异质差异变化明显,与同纬度地区常态地貌的森林群落相比,鄂西南森林群落具有更高的丰富性和异质性[21]。目前在该区域的相关研究主要涉及物种组成及物种多样性[6, 22-24]、林分空间结构[25-26]、种群恢复及发展动态[27-29]等方面,而关于尺度变化下林分空间结构和多样性的变化规律及相互关系还尚未有相关报道。因此,本研究依托湖北恩施森林生态系统国家定位观测研究站,以湖北省鄂西南3个研究区(七姊妹山自然保护区、木林子自然保护区和金子山国有林场)为评价单元,通过分析不同研究尺度上的林分空间结构特征、物种多样性的变化,并在此基础上进一步探究林分空间结构特征与物种多样性的相互影响,以期为鄂西南常绿落叶阔叶混交林的结构研究和物种多样性的维持与提高提供一定科学依据。
-
研究区分别位于湖北省恩施州鹤峰县木林子国家级自然保护区、宣恩县七姊妹山国家级自然保护区和利川市团堡镇金子山国有林场。整个区域属典型亚热带季风性气候,夏季温湿多雨,冬季严寒,日照充足(表1)。研究区域内植物资源丰富,原生性强,区系组成复杂,具有保存较好的原生植被和受到人为干扰并处于不同恢复阶段的次生林[29]。其主要林分类型为亚热带常绿落叶阔叶混交林。
表 1 研究区概况
Table 1. Basic situation of study area
样地
Plot地理位置
Location海拔/m
Altitude土壤类型
Soil type年均气温/℃
Annual average
temperature年均降水量/mm
Annual
precipitation无霜期/d
Frost-free
period年日照时数/h
Annual
sunshine
hours树种数
Varieties of
trees七姊妹山 30°03'44″ N,109°72'08″ E 1 343~1 486 黄壤、黄棕壤、棕壤 13.7 1 667.5 249 1 212 257 木林子 30°06'21″ N,110°16'32″ E 1 588~1 780 黄棕壤、棕壤、黄壤 15.5 1 529.2 230 1 298 230 金子山 30°29'47″ N,109°06'67″ E 1 320~1 425 黄棕壤、棕壤 12.7 1 475.0 230 1 300 194 -
2014 年6 月至2016 年8 月,参照CTFS(The Center for Tropical Forest Sciences)样地建设技术规范,使用全站仪先后在木林子、七姊妹山和金子山建立15 、6 和6 hm2大样地,并记录坡度、坡向、坡位、土壤、样地中心点经纬度和海拔等生境因子。样地内所有DBH ≥ 1 cm 的木本植物挂牌并进行每木检尺,记录树种名、胸径、坐标、树高、冠幅、萌生等林木特征值。
依据3个大样地基础调查数据,每个大样地以东西方向为X轴,南北方向为Y轴,左下角为坐标原点,依次向右上角以20 m为间距划分10块不同尺度样地,最大尺度为200 m × 200 m。
-
重要值( IV )是确定群落中优势种反映其作用和地位的指标。重要值( IV ) = 相对密度( RA ) + 相对频度( RF ) + 相对显著度( RP )。
-
基于林分空间结构单元,以参照树及其最近4株相邻木构成的结构单元选取角尺度、大小比数、混交度分析林分空间结构特征。具体计算公式见表2。
表 2 空间结构参数公式
Table 2. Formulations of spatial structure parameters
参数 Parameter 公式 Formulation 备注 Notes 角尺度 Uniform angle index ${\mathit{W}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{z}}_{\mathit{i}\mathit{j}} $ 当第j个α角小于标准角α0时,Zij= 1;反之,Zij= 0 大小比数 Dominance ${\mathit{U}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{k}}_{\mathit{i}\mathit{j}} $ 当相邻木j比参照树i小时,Kij= 0;反之,Kij= 1 混交度 Mingling ${\mathit{M}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{v}}_{\mathit{i}\mathit{j}} $ 当相邻木j与参照树i非同种时,Vij= 1;反之,Vij= 0 -
对物种丰富度、多样性以及均匀度的测定选用Simpson指数、Shannon-Weiner指数和Pielou指数计算(表3)。
表 3 多样性指数
Table 3. Diversity index
指数 Index 公式 Formulation 备注 Notes Simpson指数 ${D}=1-\sum\limits_{{i}=1}^{{S}}{{P}}_{{i}}^{2} $ S为样地中某一层次物种的总数;Pi为物种 i 的个体数占总个体数的比例;
H为Shannon-Wiener指数Shannon-Weiner指数 ${H}=-\sum\limits_{ {i}=1}^{ {S} }{ {P} }_{ {i} }^{2}{ {{\rm{log}}} }_{2}{ {P} }_{ {i} }$ Pielou均匀度指数 ${J}={H}/{{\rm{ln}}}{S}$ -
筛选出DBH ≥ 5 cm的乔木,利用林木特征值计算乔木层的空间结构参数及多样性指数。采用林分空间结构分析软件Winkelmass[30-32]计算核心区林木空间结构参数W、U和M,R 4.1.1计算物种多样性指数和物种重要值;利用SPSS 24、Excel进行数据分析。不同尺度下林分空间结构与物种多样性的关系使用R 4.1.1代码进行Pearson显著相关性检验。使用R 4.1.1和Origin 2017作图。
-
重要值代表了各树种在林分空间中的地位和优势程度。3个研究区内物种丰富,其中重要值排位前4的乔木树种如表4所示。七姊妹山自然保护区以川陕鹅耳枥(Carpinus fargesiana H. Winkl.)、多脉青冈(Cyclobalanopsis multinervis W.C.Cheng & T.Hong)为主;木林子自然保护区以多脉青冈、川陕鹅耳枥占优势种地位,石灰花楸(Sorbus folgneri (Schneid.) Rehd.)和小叶青冈(Cyclobalanopsis myrsinifolia (Blume) Oersted)为亚优势种。该群落中优势种和亚优势种的相对密度、重要值较其余研究区差距都较小,表明群落组成中这些树种的个体数量都较多。金子山国有林场中锥栗(Castanea henryi (Skan) Rehd. et Wils.)的重要值(22.38%),明显高于其他乔木物种,其次为川陕鹅耳枥(8.24%)。金子山林分内有较多的锥栗大径级木,所以相对显著度高达44.88%。总的来看,3个研究区主要优势树种有一定差异,但是川陕鹅耳枥的重要值在3个研究区群落中都较高,这说明该种群在鄂西南常绿落叶阔叶混交林群落构建中起重要作用,群落中常见优势乔木树种为川陕鹅耳枥、多脉青冈和锥栗。
表 4 主要乔木树种重要值
Table 4. Importance value of main tree species
样地
Plot树种
Species相对密度RA/%
Relative abundance相对频度RF/%
Relative frequency相对显著度RP/%
Relative prominence重要值IV/%
Important value七姊妹山自然保护区
Qizimeishan National Nature Reserve川陕鹅耳枥
Carpinus fargesiana H. Winkl.16.33 4.77 16.70 12.60 多脉青冈
Cyclobalanopsis multinervis W.C.Cheng & T.Hong10.92 4.13 8.26 7.77 交让木
Daphniphyllum macropodum Miq.8.86 4.66 7.06 6.86 灯台树
Cornus controversa Hemsley4.03 3.98 9.27 5.76 木林子自然保护区
Mulinzi National Nature Reserve多脉青冈
Cyclobalanopsis multinervis W.C.Cheng & T.Hong13.71 6.49 14.66 11.62 川陕鹅耳枥
Carpinus fargesiana H. Winkl.11.46 6.43 13.72 10.54 石灰花楸
Sorbus folgneri (Schneid.) Rehd.10.14 5.27 8.87 8.09 小叶青冈
Cyclobalanopsis myrsinifolia (Blume) Oersted8.95 5.63 8.52 7.70 金子山国有林场
Jinzishan National Forest Farm锥栗
Castanea henryi (Skan) Rehd. et Wils.14.55 7.70 44.88 22.38 川陕鹅耳枥
Carpinus fargesiana H. Winkl.13.42 6.21 5.09 8.24 短柄枹栎
Quercus glandulifera Bl.9.23 7.30 4.35 6.96 大叶杨
Populus lasiocarpa Oliv.5.22 6.21 7.03 6.15 -
在不考虑小尺度样地(20 m × 20 m)出现极个别特殊数值的情况下,不同尺度上,3个研究区内林分多样性指数与尺度之间呈现出明显的非线性相关(图1),整体上Shannon指数和Simpson指数随着尺度增大呈现先快速增加后趋于缓慢增加或稳定的趋势,Pielou指数则随着尺度增加逐步下降然后趋于稳定,但在各个研究区呈现出一定差异。随着空间尺度的增加,七姊妹山多样性指数增幅逐渐趋缓;金子山波动变化较大,但整体趋势与七姊妹山接近;木林子多样性增幅逐渐趋于零,且在大尺度上(≥ 160 m × 160 m),多样性指数值显著低于其余2个研究区。金子山多样性指数在尺度40 m × 40 m、120 m × 120 m时骤降,这可能是由于金子山部分样地受到一定程度的人为干扰,林分内物种受到了损害,从而导致多样性指数的明显下降。鄂西南3个研究区的多样性指标在不同尺度上的变异系数整体呈现先减小后增大的趋势,其中Shannon-weiner指数变异系数在不同尺度上波动最剧烈,除在40 m × 40 m尺度时的Shannon-weiner指数,其余尺度不同指数都低于8%。Simpson指数和Pielou指数从尺度20 m × 20 m上升至140 m × 140 m,变异系数逐渐降低,随后达到160 m × 160 m以上,变异系数又出现上升趋势,且随着空间尺度增大,变异系数也随之增大。
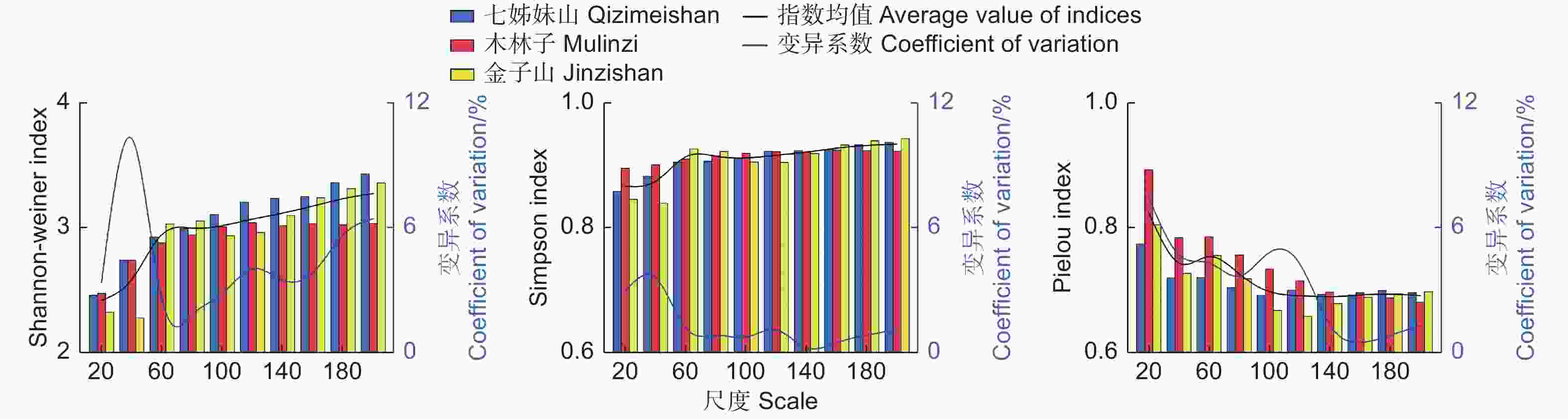
图 1 不同尺度多样性指数变化
Figure 1. Variations of diversity indices at different scales
-
七姊妹山自然保护区、木林子自然保护区和金子山国有林场在不同尺度上林分空间结构参数W、U、M变化如图2所示。从局部看,木林子角尺度曲线整体低于其他研究区,角尺度均值表现为七姊妹山(0.557)> 金子山(0.551)> 木林子(0.533),且各研究区角尺度均值达到0.536(> 0.517)以上,水平空间分布格局多属于轻微聚集分布,而木林子林木分布格局聚集程度最低。林分大小比数均值在0.500~0.510之间,随着尺度增加,各研究区差异变化不明显,均呈现中庸状态。说明各研究区林木大小分化处于中庸状态,空间结构单元中相邻木与参照树胸径大小差异化不明显,不同优势度树种数量相近,林分内树种生长竞争较为激烈。林分空间隔离程度在3个研究区有较明显差异,各研究区林分混交度均值达0.732以上,其中木林子整体处于极强度混交状态(平均混交度0.757),这表明研究区多数参照木周围相邻木不为同种,单种聚集发生概率低,林木整体混交良好。鄂西南3个研究区的林分空间结构参数在不同尺度上的变异系数变化,在20 m × 20 m至100 m × 100 m尺度范围内,结构参数指标变异系数波动较大(1.39%~10.82%),随着尺度上升至120 m × 120 m尺度,结构参数指标变异系数分别为:W(2.08%)、M(2.93%)和U(0.40%),逐渐趋于稳定。因此鄂西南常绿落叶阔叶混交林林分空间结构研究最小样地尺度范围为120 m × 120 m,该尺度能较全面的反映鄂西南林分空间结构变化规律。
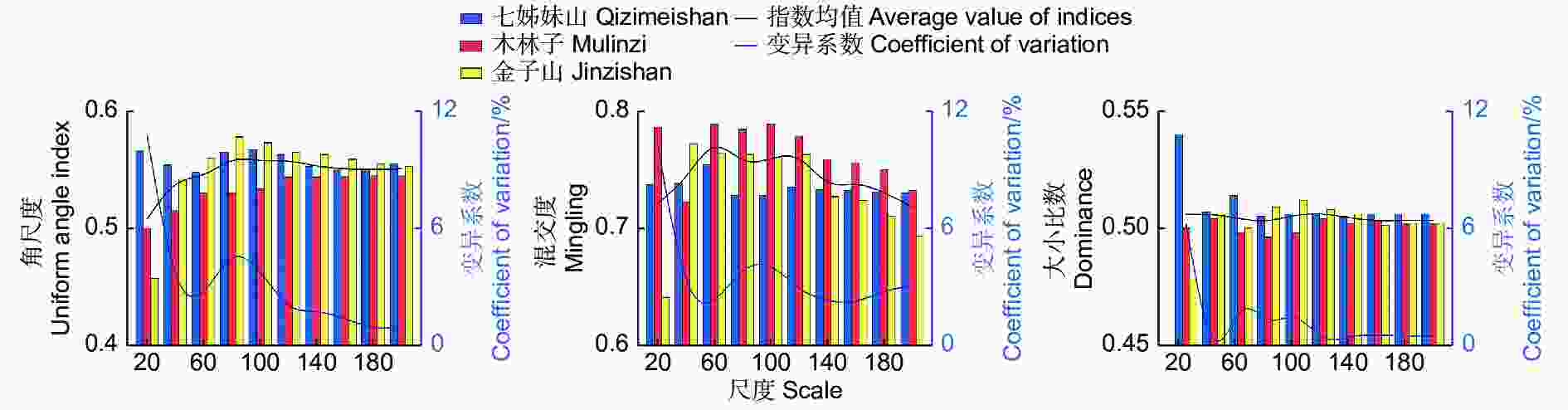
图 2 不同尺度空间结构参数变化
Figure 2. Variations of spatial structure parameters at different scales
-
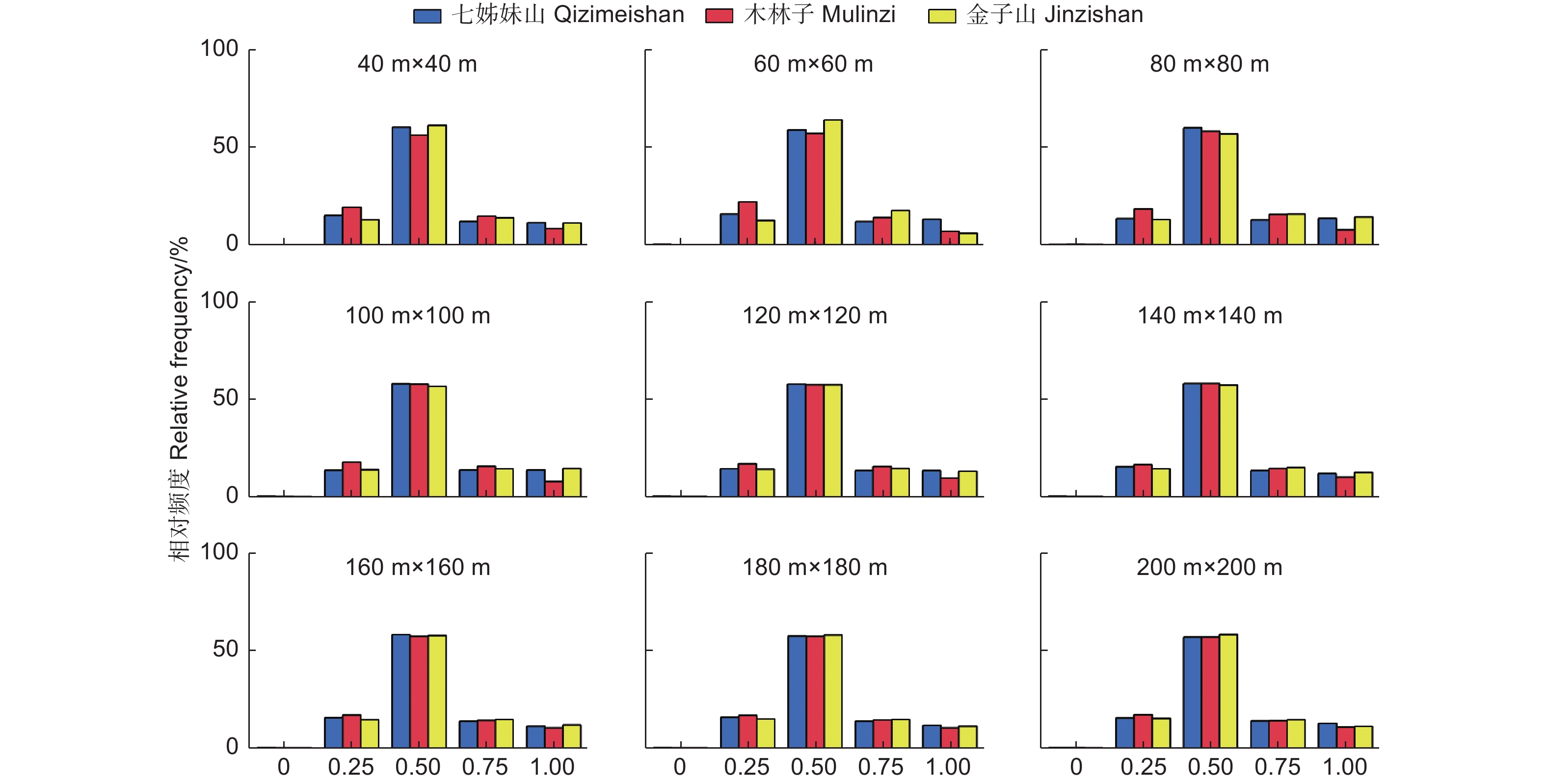
由图3可知,在鄂西南常绿落叶阔叶混交林中,不同尺度上角尺度频率分布取值集中在0.5的林木个体数量超过了55%,处于0.5右侧的取值频率高于0.5左侧的频率值,且W = 0(非常均匀)的频率值均低于1%,表明林分中处于随机分布状态的林木个体数量最多,总体聚集程度不高,因此鄂西南常绿落叶阔叶混交林林分的水平分布格局在各尺度上呈现出微聚集分布状态,且随着尺度增加,分布状态未呈现明显变化。结合林分不同树种的空间结构单元的平均角尺度分析发现,主要优势树种的平均角尺度与整体林分角尺度接近,这进一步说明优势种对于整个林分林木的分布中起着主导作用。
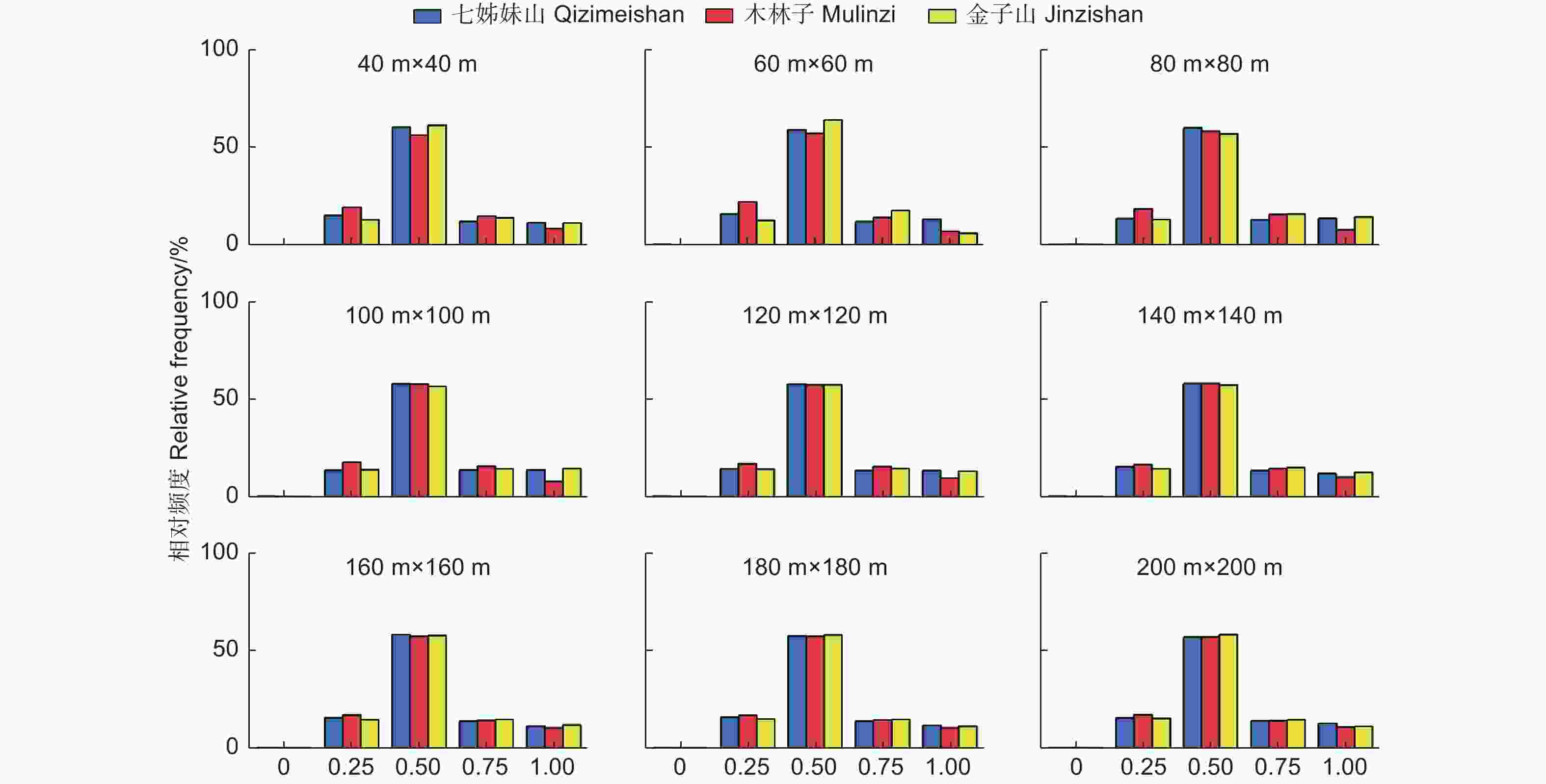
图 3 不同尺度林分角尺度(W)一元分布
Figure 3. The univariate distribution of stand uniform angle at different scales
3个研究区的结构参数取值频率分布表现出与尺度的极大相关性,即随着尺度的增加(40 m × 40 m → 200 m × 200 m),各研究区5个取值的频率值差异逐渐减小并趋于相等。例如:W = 0.25时,小尺度(40 m × 40 m)中,3个研究区频率值变异系数为28.4%,而在在大尺度(200 m × 200 m)中,3个研究区频率值变异系数为6.6%。这种现象同样适用于大小比数、混交度不同取值频率分布。可见,尺度观测增加到一定的范围时,鄂西南各研究区林分空间结构基本趋于一致。
-
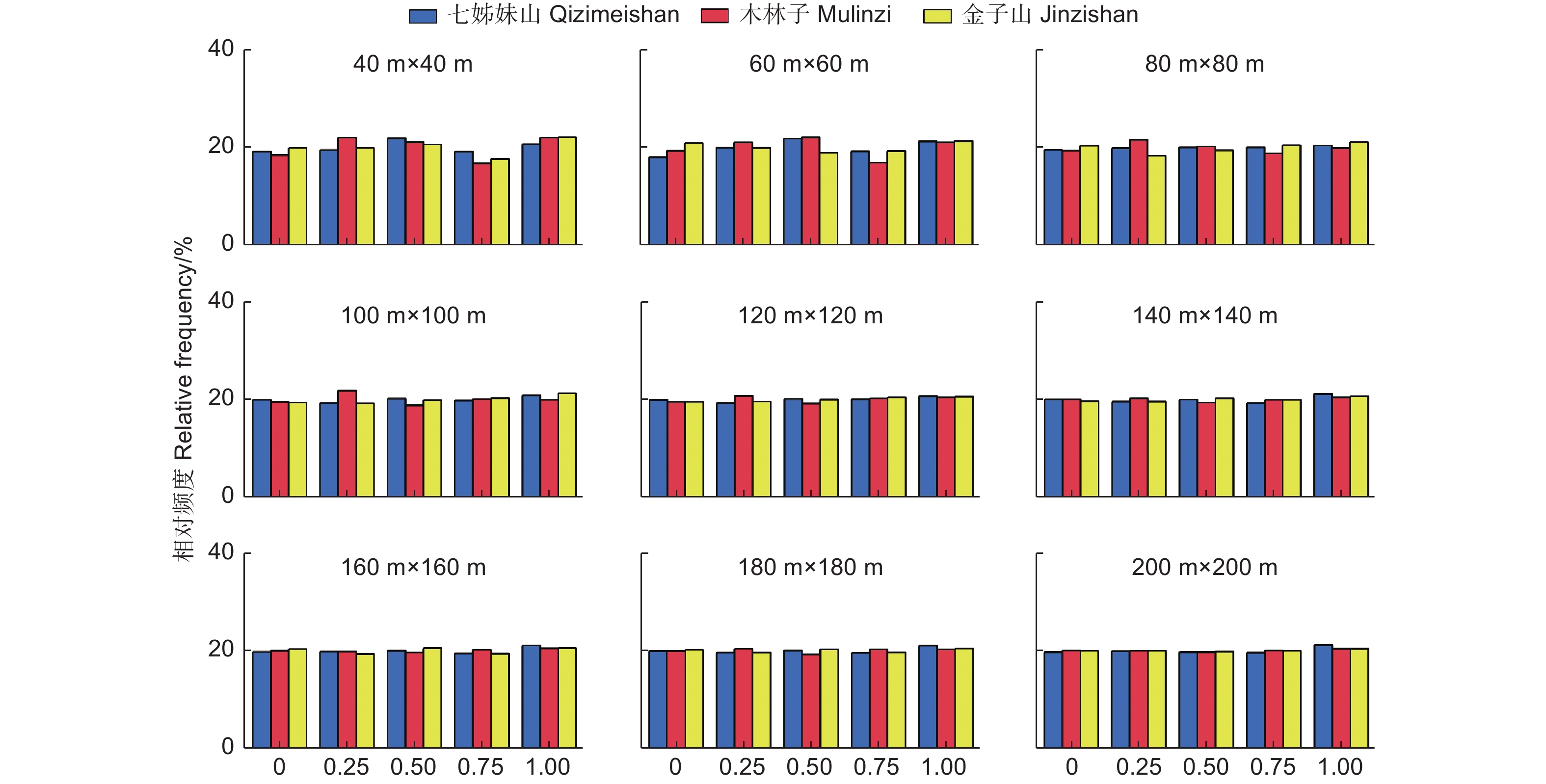
参照树与4株相邻木构成的最佳空间结构单元中,描述参照树与相邻木的相对优势程度,各优势程度林木占全部林木的比例,即大小比数的一元分布。图4可见,不同研究区不同尺度的大小比数频率值都在20%左右,且随着尺度增加到120 m × 120 m时,各大小比数频率趋于稳定,表明鄂西南常绿落叶阔叶混交林中不同优势度的树种数量大致相等,群落内竞争处于中庸状态,且随着尺度上升更加稳定。
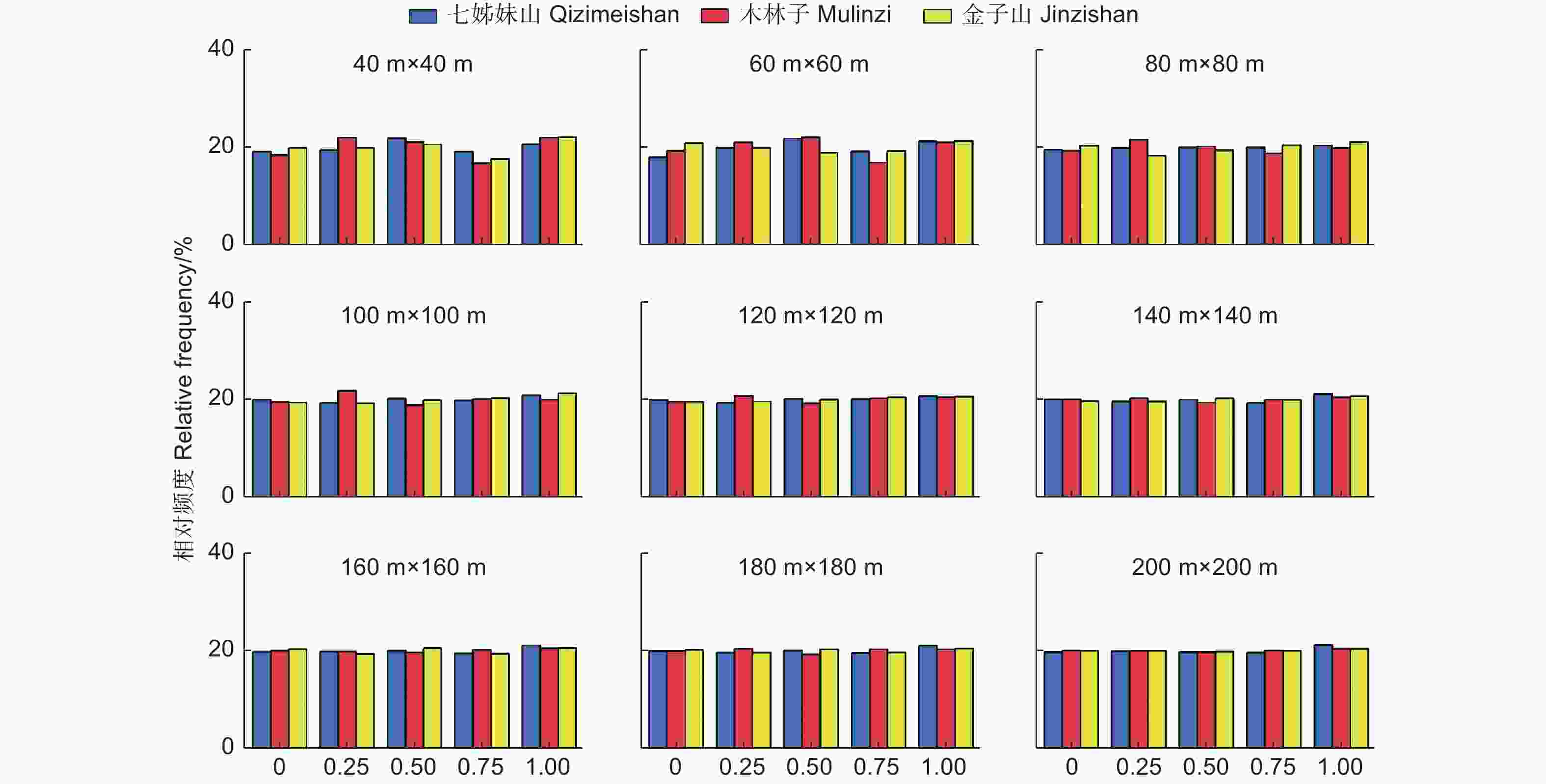
图 4 不同尺度林分大小比数(U)一元分布
Figure 4. The univariate distribution of stand dominance at different scales
-
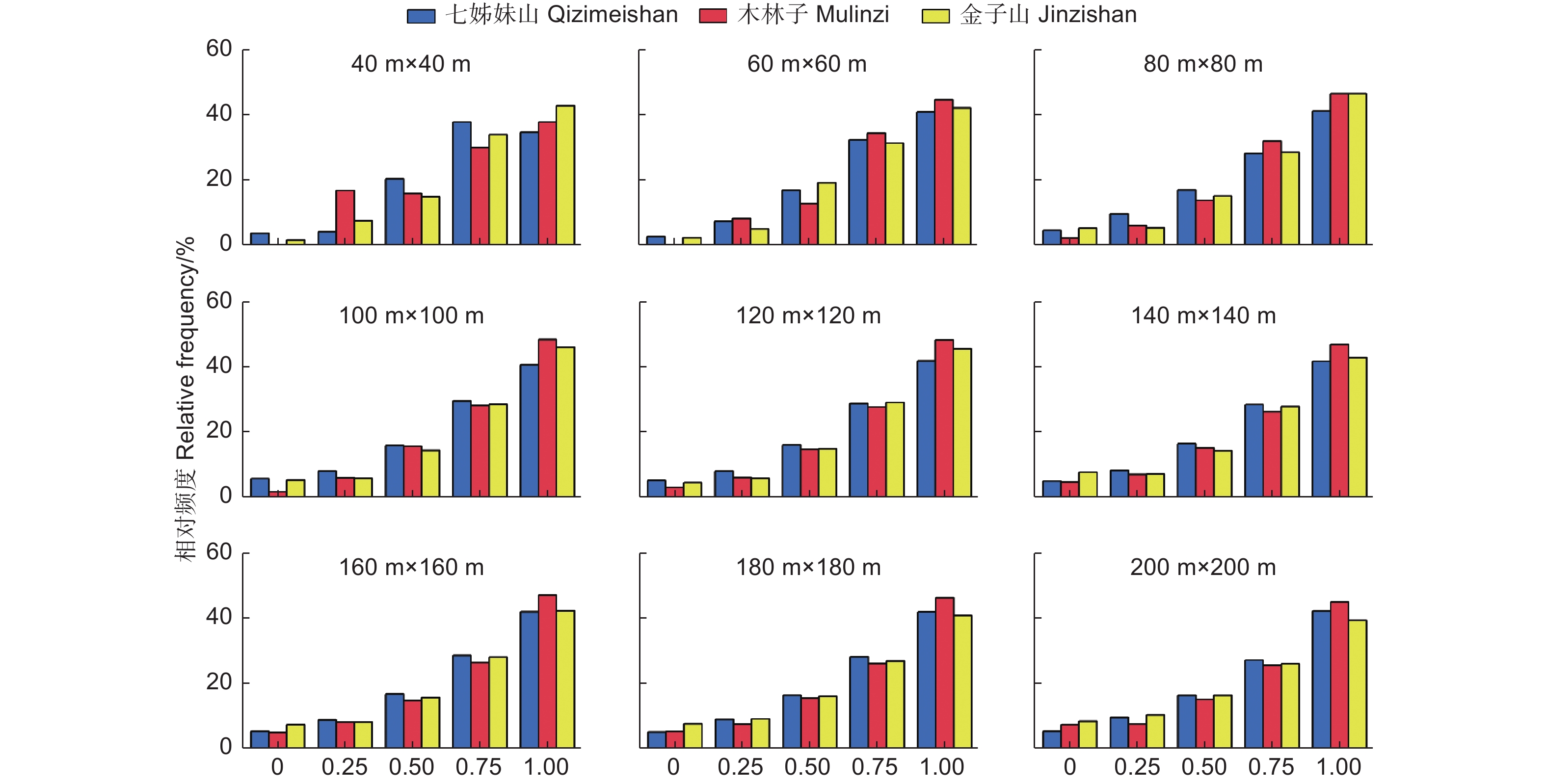
混交度说明了结构单元中的树种空间隔离程度,以是否为同种树种计算出5个取值等级,每个取值等级在林分中所占比例,即为混交度一元分布。如图5所示,鄂西南3个研究区不同尺度上林木混交度频率分布主要集中在0.75(强度混交)和1(极强度混交)状态,频率和均在64%以上,零度混交和弱度混交所占比例较少,表明3个研究区林分呈现较强的混交状态,树种组成丰富。
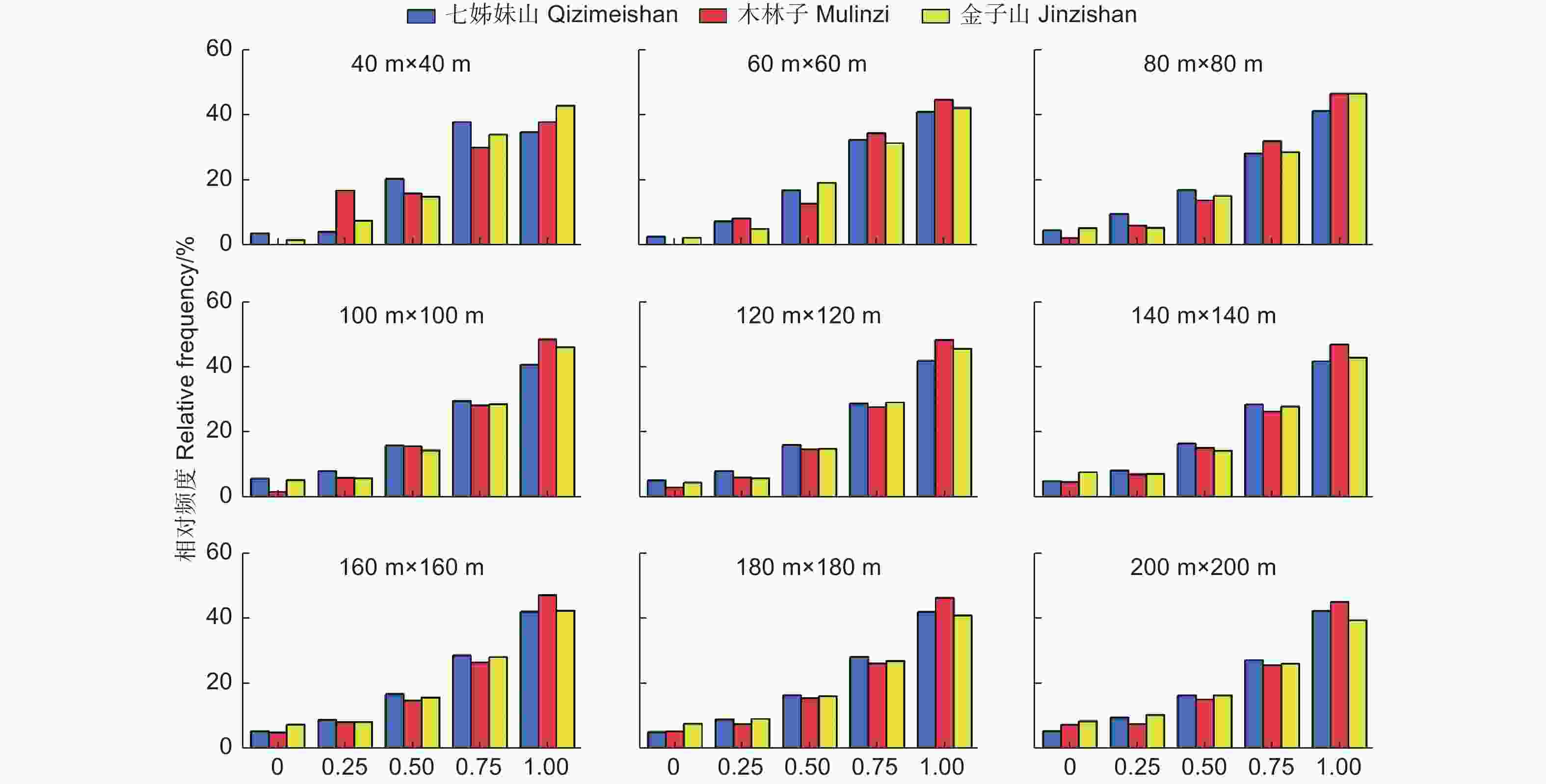
图 5 不同尺度林分混交度(M)一元分布
Figure 5. The univariate distribution of Mingling at different scales
-
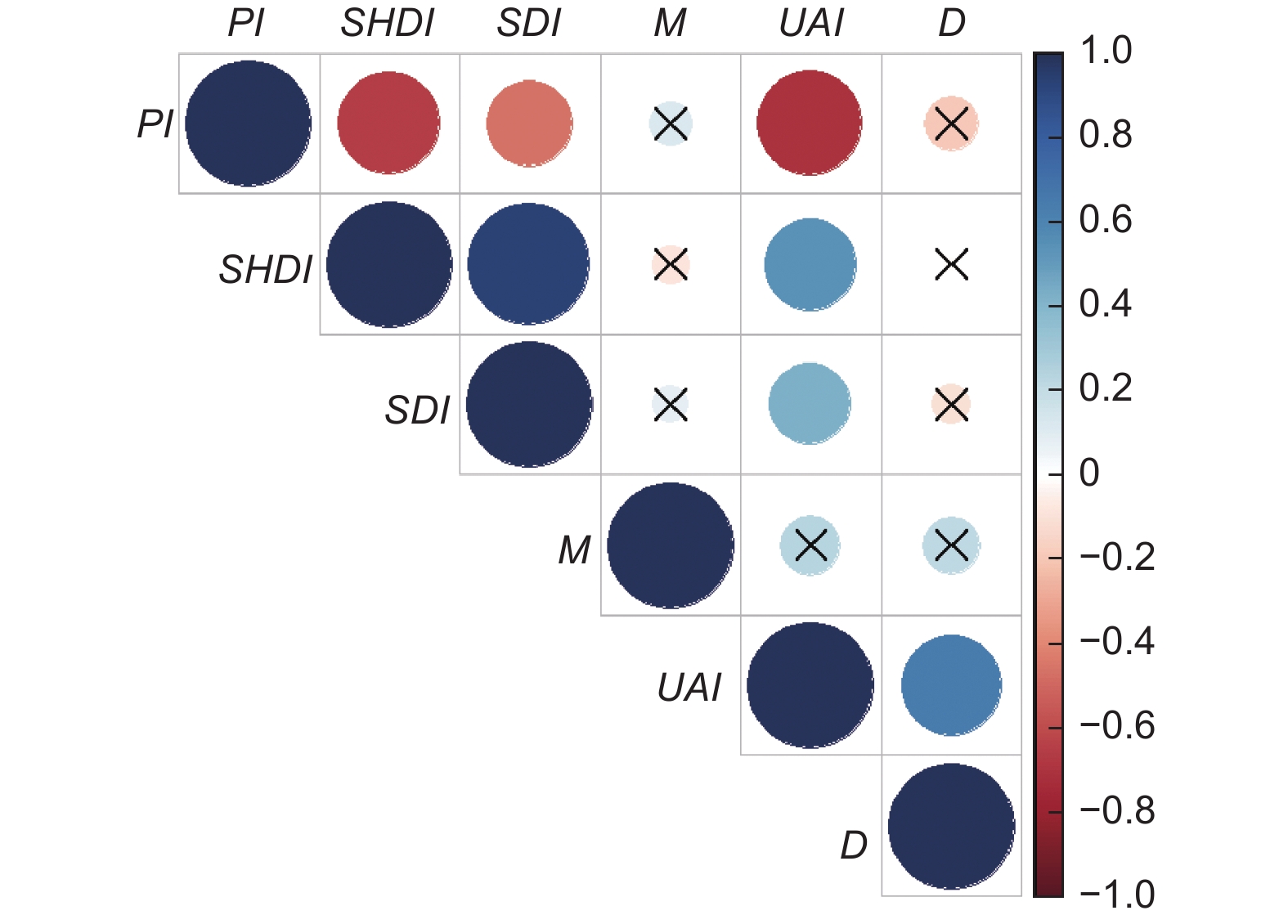
以七姊妹山、木林子、金子山3个大样地调查数据为基础,将不同尺度林分空间结构参数与乔木物种多样性指标进行相关性分析(图6)。亚热带常绿落叶阔叶混交林在不同尺度上,混交度和大小比数与物种多样性不存在显著相关性(P > 0.05),角尺度与乔木Pielou指数存在中度负相关性(P < 0.01),角尺度与Shannon-Weiner指数存在中度正相关性(P < 0.01),角尺度与Simpson指数存在低度相关性(P < 0.05)。综合来看,不同尺度变化中角尺度与乔木物种多样性存在显著相关性,即角尺度是影响鄂西南亚热带常绿落叶阔叶混交林林分乔木多样性的一个重要因子,随着林分角尺度的增大,林分内物种多样性程度越高,群落也越复杂,林木分布越趋向于不均匀。
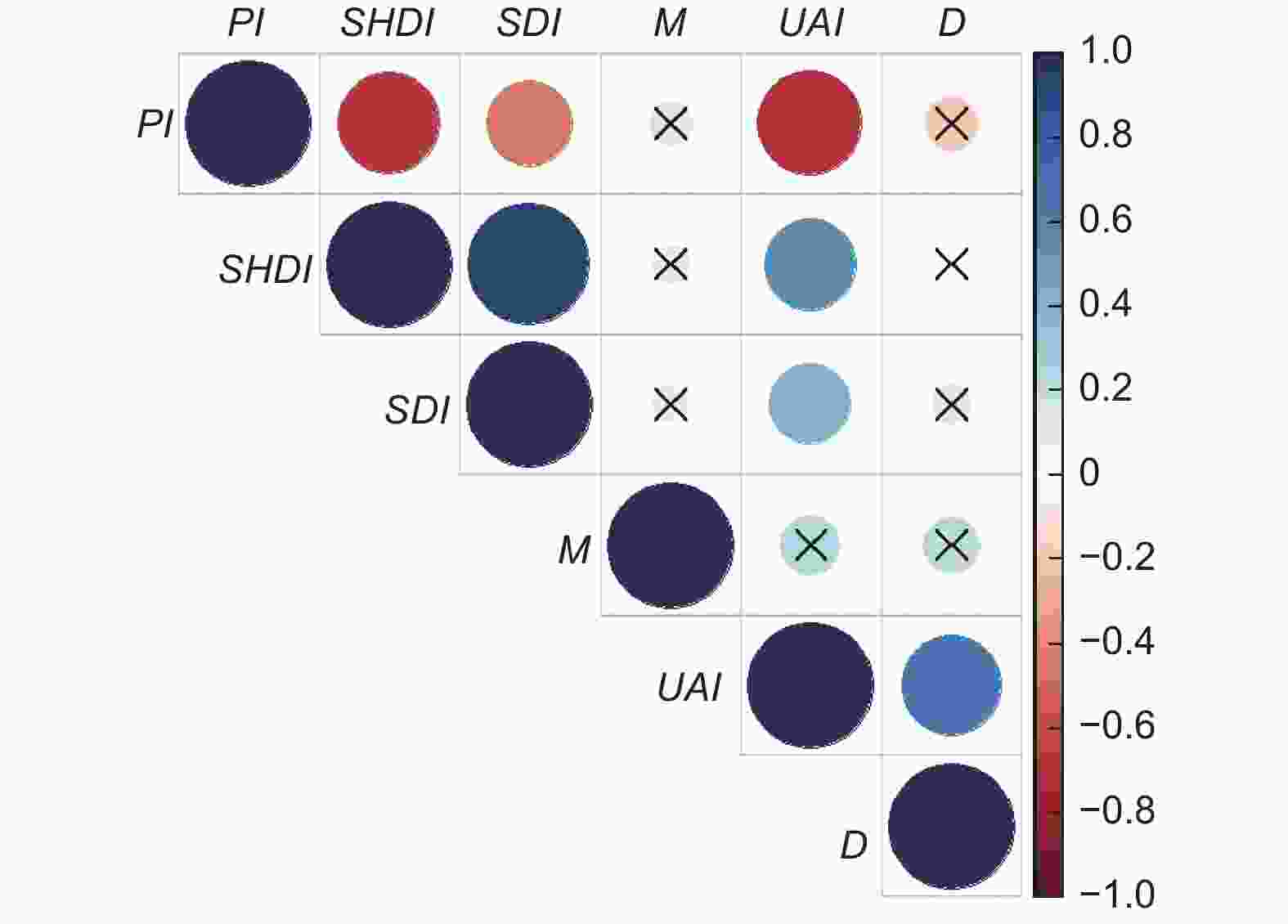
图 6 林分空间结构指标与乔木物种多样性指数之间的相关性
Figure 6. Correlation between stand spatial structure indices and tree species diversity index
-
本研究基于鄂西南3个研究区大样地清查数据中DBH ≥ 5 cm的乔木林木属性和位置信息,分析林分空间结构指标和乔木物种多样性指数随着尺度的变化趋势与相关性。研究结果一定程度上弥补了以往对大尺度林分空间结构研究及其与多样性相互关系的空缺[33-34],与近几年相关研究对比发现:
-
通过对鄂西南3个研究区常绿落叶阔叶混交林乔木层物种多样性指标进行分析,发现各研究区物种多样性较高,且不同区域优势树种的重要值存在一定的相似性,因此鄂西南常绿落叶阔叶混交林常见优势乔木树种为川陕鹅耳枥、多脉青冈和锥栗等,这与相关学者研究结果基本一致[28,35]。同时研究尺度的大小影响着物种空间分布格局,合适的样地尺度对于生物多样性和林分空间结构研究至关重要[36]。鄂西南3个研究区物种多样性随着研究尺度的上升,多样性与尺度之间呈现出明显的非线性相关。金子山Shannon指数和Simpson指数在40 m × 40 m、120 m × 120 m尺度上骤降,主要是由于部分样地受到了较强的人为干扰。随着尺度增加到120 m × 120 m以上时,3个研究区的多样性指数波动幅度也逐渐减小,这与He F等人[37]研究结果一致,表明小尺度范围内样地空间异质性表达较为明显,尺度增加带来更加多样的生境,但新生境出现速度也会减慢,异质性逐渐平均化,因此大尺度上很少出现剧烈波动。鄂西南作为喀斯特地貌发育最强烈的地区,创造了丰富多样的生境,使鄂西南地区森林生态系统维持着较高水平的物种多样性、异质性和复杂性[21],因此鄂西南七姊妹山、金子山、木林子有着鲜明的多样性差异,同时受限于目前的研究尺度,关于物种多样性最小面积还需进一步增加尺度以进行确定。
-
鄂西南地区不同尺度林分空间特征表现出微聚集分布、大小分化中庸且混交良好的状态,与薛卫星[38-39]等的研究结果相一致。林分中主要优势树种的结构参数均值与整体林分均值接近,表明主要优势树种的林木分布格局很大程度上决定了林分空间特征[40] 。同时3个空间结构参数存在区域性差异,木林子自然保护区处于微聚集、极强度混交状态,其余研究区林木聚集程度高于木林子,且呈强度混交状态。一般认为,天然林在演替初期会呈明显的聚集分布,这是由种源扩散和林中开放空间的形式决定的,随着演替的推进,林木个体对资源的需求导致了生存竞争,这种自然稀疏的过程促使林分朝着随机分布格局演替[41]。同时聚集分布格局相较于随机分布格局,总体混交度更低[18, 42]。因此木林子虽然聚集程度相对较低,但由于各树种种内排斥、同树种种间分布较均匀,因此林分整体混交度较高,演替阶段也高于七姊妹山和金子山。随着样地尺度的变化,其对应林木的空间分布格局也不同,这表明,取样面积应有一定的大小才能真实地反映林分特征[42]。研究显示能够稳定反映鄂西南亚热带常绿落叶阔叶混交林林分空间结构变化规律的最小样地尺度为120 m × 120 m(1.44 hm2),而胡中洋等[19]对新疆天山国家级自然保护区确立过80 m × 80 m(0.64 hm2)最小样地尺度,与本研究有一定差异。这可能是由于两个区域气候、地形、生境等有明显空间异质性,导致不同植被类型的样地尺度有所差别,并且鄂西南地区生物多样性非常丰富[19, 43],林分结构在尺度上表现出更复杂的特征,因此120 m × 120 m的尺度更适合鄂西南地区的空间结构样地调查。
-
林分空间结构与林分的物种多样性存在一定相关性[4]。同一林分类型不同演替阶段中,多种空间结构指标将综合影响物种多样性,其中混交度、角尺度为关键因子,但各个演替阶段的影响又各不相同。鄂西南3个区域不同尺度上林分空间结构参数与乔木物种多样性指标存在一定的相关性,且仅表现为角尺度与林分多样性存在中度相关性,其他结构参数与林分多样性则不存在相关性,而黎芳等[4]研究的飞播马尾松人工林,张亚昊等[44]研究的不同演替阶段马尾松林则表现出空间结构指标如角尺度、混交度[45]等对林分物种多样性有显著影响。导致研究结果差异首先是因为鄂西南常绿落叶阔叶混交林物种组成丰富,且处于基本无人为干扰状态,因此空间结构主要通过自然竞争、稀疏等形成,整体呈现结构稳定,多样性丰富维持的状态。此外,由于本研究的区域尺度较大,乔木物种分布格局影响因子如:地形、温度、光照、水分等发生异质性变化,物种多样性因此也呈现出相应变化,表现出一定的相关性。一般认为小尺度上,环境差异性较小,植物群落分布主要受种子扩散限制性影响[46];大尺度上植物群落分布更多是受到生境异质性的作用,尤其是喀斯特地区复杂的地形地貌,故在不同的尺度上反映出不同的生态学特征[47],这也成为影响物种多样性差异的关键因素之一[47]。结合本研究的结果,大尺度上的乔木物种多样性变化不仅受到生境异质性的作用,还受到空间结构参数角尺度的影响。
-
鄂西南地区亚热带常绿落叶阔叶混交林中林分空间结构特征表现出微聚集分布、大小分化中庸且混交良好的状态,该区进行林分空间结构调查的最适尺度为120 m × 120 m。不同尺度上林分空间结构参数角尺度与林分物种多样性存在中度相关性,其他结构参数与林分物种多样性不存在相关性,可见,在天然林中不同研究尺度下角尺度是林分空间结构影响物种多样性的一个关键因子。在今后鄂西南亚热带常绿落叶阔叶混交林可持续发展过程中,建议考虑适当调整林分空间结构,优化角尺度,利于该区乔木物种多样性的提高,以进一步促进该区森林生态系统的稳定发展。
林分空间结构与物种多样性随空间尺度变化的规律研究
Variations on Stand Spatial Structure and Species Diversity in Different Spatial Scales
-
摘要:
目的 探究林分空间结构、物种多样性随尺度变化的规律及林分空间结构与物种多样性的相互关系,为鄂西南常绿落叶阔叶混交林可持续经营提供科学依据。 方法 以鄂西南七姊妹山自然保护区、木林子自然保护区和金子山国有林场3个研究区的典型植被——亚热带常绿落叶阔叶混交林为研究对象,基于各研究区大样地调查数据,分析从20 m × 20 m到200 m × 200 m不同尺度下,林分空间结构指标和物种多样性指数及其随尺度变化的规律,结合Pearson相关系数法解析尺度变化影响下的空间结构指标与物种多样性的相关性。 结果 鄂西南地区常绿落叶阔叶混交林中物种丰富,常见优势乔木树种为川陕鹅耳枥、多脉青冈和锥栗。林分空间结构特征在尺度 ≥ 120 m × 120 m后变化趋于稳定,整体表现出微聚集分布、大小分化中庸且混交良好的状态。不同尺度上林分空间结构参数角尺度与林分物种多样性存在中度相关性(P < 0.01),其他结构参数与林分多样性不存在相关性。 结论 鄂西南亚热带常绿落叶阔叶混交天然林,林分空间结构调查的最适尺度为120 m × 120 m;不同研究尺度下,角尺度是林分空间结构影响物种多样性的一个关键因子。 Abstract:Objective To provide a scientific reference for the sustainable management of evergreen deciduous broad-leaved mixed forest in southwest Hubei. Method The stand spatial structure and species diversity as well as their correlations in the different spatial scales ranging from the sample size 20 m × 20 m to 200 m× 200 m in the subtropical evergreen deciduous broad-leaved mixed forests in Qizimeishan National Nature Reserve, Mulinzi National Nature Reseve and Jinzishan National forest farm in southwest Hubei province were explored. Result The tree species were abundant in this study, and Carpinus fargesiana, Cyclobalanopsis multinervis and Castanea henryi were dominant tree species. The spatial structure diversities tended to be stable when the sample size ≥ 120 m× 120 m, showing the state of micro-aggregation distribution, moderate differentiation and good mixing. The Uniform angle index was correlated with species diversity (p < 0.01) at different scales, while other structural parameters had no correlations with species diversity. Conclusion The optimum investigation scale for the stand spatial structure of subtropical evergreen deciduous broad-leaved mixed forest in southwest Hubei is 120 m × 120 m. In addition, the Uniform angle index was a key factor of stand spatial structure affecting species diversity in different spatial scales. -
Key words:
- stand spatial structure
- / species diversity
- / univariate distribution
- / scale effect
-
图 2 不同尺度空间结构参数变化
Figure 2. Variations of spatial structure parameters at different scales
图 3 不同尺度林分角尺度(W)一元分布
Figure 3. The univariate distribution of stand uniform angle at different scales
图 4 不同尺度林分大小比数(U)一元分布
Figure 4. The univariate distribution of stand dominance at different scales
图 5 不同尺度林分混交度(M)一元分布
Figure 5. The univariate distribution of Mingling at different scales
图 6 林分空间结构指标与乔木物种多样性指数之间的相关性
Figure 6. Correlation between stand spatial structure indices and tree species diversity index
表 1 研究区概况
Table 1. Basic situation of study area
样地
Plot地理位置
Location海拔/m
Altitude土壤类型
Soil type年均气温/℃
Annual average
temperature年均降水量/mm
Annual
precipitation无霜期/d
Frost-free
period年日照时数/h
Annual
sunshine
hours树种数
Varieties of
trees七姊妹山 30°03'44″ N,109°72'08″ E 1 343~1 486 黄壤、黄棕壤、棕壤 13.7 1 667.5 249 1 212 257 木林子 30°06'21″ N,110°16'32″ E 1 588~1 780 黄棕壤、棕壤、黄壤 15.5 1 529.2 230 1 298 230 金子山 30°29'47″ N,109°06'67″ E 1 320~1 425 黄棕壤、棕壤 12.7 1 475.0 230 1 300 194  下载: 导出CSV
下载: 导出CSV
表 2 空间结构参数公式
Table 2. Formulations of spatial structure parameters
参数 Parameter 公式 Formulation 备注 Notes 角尺度 Uniform angle index ${\mathit{W}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{z}}_{\mathit{i}\mathit{j}} $ 当第j个α角小于标准角α0时,Zij= 1;反之,Zij= 0 大小比数 Dominance ${\mathit{U}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{k}}_{\mathit{i}\mathit{j}} $ 当相邻木j比参照树i小时,Kij= 0;反之,Kij= 1 混交度 Mingling ${\mathit{M}}_{\mathit{i}}=\dfrac{1}{4}\sum\limits_{\mathit{j}=1}^{4}{\mathit{v}}_{\mathit{i}\mathit{j}} $ 当相邻木j与参照树i非同种时,Vij= 1;反之,Vij= 0
下载: 导出CSV
表 3 多样性指数
Table 3. Diversity index
指数 Index 公式 Formulation 备注 Notes Simpson指数 ${D}=1-\sum\limits_{{i}=1}^{{S}}{{P}}_{{i}}^{2} $ S为样地中某一层次物种的总数;Pi为物种 i 的个体数占总个体数的比例;
H为Shannon-Wiener指数Shannon-Weiner指数 ${H}=-\sum\limits_{ {i}=1}^{ {S} }{ {P} }_{ {i} }^{2}{ {{\rm{log}}} }_{2}{ {P} }_{ {i} }$ Pielou均匀度指数 ${J}={H}/{{\rm{ln}}}{S}$
下载: 导出CSV
表 4 主要乔木树种重要值
Table 4. Importance value of main tree species
样地
Plot树种
Species相对密度RA/%
Relative abundance相对频度RF/%
Relative frequency相对显著度RP/%
Relative prominence重要值IV/%
Important value七姊妹山自然保护区
Qizimeishan National Nature Reserve川陕鹅耳枥
Carpinus fargesiana H. Winkl.16.33 4.77 16.70 12.60 多脉青冈
Cyclobalanopsis multinervis W.C.Cheng & T.Hong10.92 4.13 8.26 7.77 交让木
Daphniphyllum macropodum Miq.8.86 4.66 7.06 6.86 灯台树
Cornus controversa Hemsley4.03 3.98 9.27 5.76 木林子自然保护区
Mulinzi National Nature Reserve多脉青冈
Cyclobalanopsis multinervis W.C.Cheng & T.Hong13.71 6.49 14.66 11.62 川陕鹅耳枥
Carpinus fargesiana H. Winkl.11.46 6.43 13.72 10.54 石灰花楸
Sorbus folgneri (Schneid.) Rehd.10.14 5.27 8.87 8.09 小叶青冈
Cyclobalanopsis myrsinifolia (Blume) Oersted8.95 5.63 8.52 7.70 金子山国有林场
Jinzishan National Forest Farm锥栗
Castanea henryi (Skan) Rehd. et Wils.14.55 7.70 44.88 22.38 川陕鹅耳枥
Carpinus fargesiana H. Winkl.13.42 6.21 5.09 8.24 短柄枹栎
Quercus glandulifera Bl.9.23 7.30 4.35 6.96 大叶杨
Populus lasiocarpa Oliv.5.22 6.21 7.03 6.15
下载: 导出CSV
-
[1] 陈 龙, 安明态, 任娇娇, 等. 贵州北盘江石漠化区植物群落物种多样性及主要乔木种间联结性[J]. 西南大学学报:自然科学版, 2020, 42(5):80-94. [2] 郭连金, 张文辉, 刘国彬. 黄土丘陵区沙棘人工林发育过程中物种多样性及种间关联变化[J]. 应用生态学报, 2007, 18(1):9-15. doi: 10.3321/j.issn:1001-9332.2007.01.002 [3] 茹文明, 张金屯, 张 峰, 等. 历山森林群落物种多样性与群落结构研究[J]. 应用生态学报, 2006, 17(4):4561-4566. [4] 黎 芳, 潘 萍, 宁金魁, 等. 飞播马尾松林林分空间结构对林下植被多样性的影响[J]. 东北林业大学学报, 2016, 44(11):31-35. doi: 10.3969/j.issn.1000-5382.2016.11.007 [5] Chase J M, McGill B J, McGlinn D J, et al. Embracing scale-dependence to achieve a deeper understanding of biodiversity and its change across communities[J]. Ecology Letters, 2018, 21(11): 1737-1751. doi: 10.1111/ele.13151 [6] 冯 广. 鄂西南天然林群落木本植被多样性研究[D]. 北京: 北京林业大学, 2020. [7] 惠刚盈. 基于相邻木关系的林分空间结构参数应用研究[J]. 北京林业大学学报, 2013, 35(4):1-9. [8] 胡艳波, 惠刚盈. 基于相邻木关系的林木密集程度表达方式研究[J]. 北京林业大学学报, 2015, 37(9):1-8. [9] Yuanfa L, Shaoming Y, Gangying H, et al. Spatial structure of timber harvested according to structure-based forest management[J]. Forest Ecology and Management, 2014, 322(6): 106-116. [10] Kint V, Lust N, Ferris R, et al. Quantification of forest stand structure applied to Scots pine (Pinus sylvestris L. ) forests[J]. Investigacion Agraria Sistemas Y Recursos Forestales, 2000, 10(1): 147-164. [11] 惠刚盈. 角尺度——一个描述林木个体分布格局的结构参数[J]. 林业科学, 1999, 35(1):39-44. [12] 惠刚盈, K. V. Gadow ,胡艳波. 林分空间结构参数角尺度的标准角选择[J]. 林业科学研究, 2004, 17(6):687-692. doi: 10.3321/j.issn:1001-1498.2004.06.001 [13] 惠刚盈, K. V. Gadow, 胡艳波. 林木分布格局类型的角尺度均值分析方法[J]. 生态学报, 2004, 24(6):1225-1229. doi: 10.3321/j.issn:1000-0933.2004.06.020 [14] 惠刚盈, 胡艳波. 角尺度在林分空间结构调整中的应用[J]. 林业资源管理, 2006, 28(2):31-35. doi: 10.3969/j.issn.1002-6622.2006.02.008 [15] 惠刚盈. 一个新的林分空间结构参数——大小比数[J]. 林业科学研究, 1999, 12(1):1-6. doi: 10.3321/j.issn:1001-1498.1999.01.001 [16] 胡艳波, 惠刚盈. 一种新的基于混交度的林木种群分布格局测度方法[J]. 北京林业大学学报, 2015, 37(1):9-14. [17] 惠刚盈, 胡艳波, 赵中华. 基于相邻木关系的树种分隔程度空间测度方法[J]. 北京林业大学学报, 2008, 30(4):131-134. doi: 10.3321/j.issn:1000-1522.2008.04.023 [18] 张 科, 冯思思, 黄 浪, 等. 树种混交度——一个基于树种谱系的林分混交度新概念[J]. 林业资源管理, 2021, 43(4):62-68. [19] 胡中洋, 刘锐之, 刘 萍. 不同尺度天山云杉活立木空间结构分析[J]. 西北林学院学报, 2020, 35(2):153-159. doi: 10.3969/j.issn.1001-7461.2020.02.23 [20] Huang J, Lu X, Huang J, et al. Conservation priority of endemic Chinese flora at family and genus levels[J]. Biodiversity& Conservation, 2016, 25(11): 23-35. [21] 安明态. 喀斯特森林土壤水分和养分格局及其植物物种多样性维持机制研究[D]. 贵阳: 贵州大学, 2019. [22] 陈思艺, 艾训儒, 姚 兰, 等. 鄂西南地区种子植物多样性与区系特征[J]. 西北植物学报, 2019, 39(2):330-342. [23] 朱 强. 鄂西南七姊妹山常绿落叶阔叶混交林物种多样性格局及维持机制[D]. 恩施: 湖北民族大学, 2020. [24] 朱 强, 艾训儒, 姚 兰, 等. 鄂西南亚热带山地常绿落叶阔叶混交林物种多度分布格局[J]. 西北植物学报, 2020, 40(6):1061-1069. [25] 朱 强, 艾训儒, 姚 兰, 等. 鄂西南川陕鹅耳枥种群结构与动态[J]. 中南林业科技大学学报, 2019, 39(8):93-100. [26] 王志鸣. 鄂西南亚热带常绿落叶阔叶混交林林分结构分析及优化经营研究[D]. 恩施: 湖北民族大学, 2019. [27] 汤景明. 鄂西南山地常绿落叶阔叶混交林恢复研究[D]. 北京: 北京林业大学, 2008. [28] 王 进, 姚 兰, 艾训儒, 等. 鄂西南不同区域亮叶桦种群结构与动态特征[J]. 应用生态学报, 2020, 31(2):357-365. [29] 冯 广, 艾训儒, 姚 兰, 等. 鄂西南亚热带常绿落叶阔叶混交林的自然恢复动态及其影响因素[J]. 林业科学, 2016, 52(8):1-9. [30] Gadow K V, Hui G Y, Albert M. Das Winkelma - Ein Strukturparameter zur Beschreibung der Individualverteilung in Waldbestnden[J]. Project: Forest Models,1998, 115(1): 1-10 [31] Hui G Y, Gadow K V. Das Winkelmass-Theoretische iiberlegungen zum optimalen Standardwinkel[J]. Project: Forest Models,2002, 173(1): 173-176. [32] 张岗岗, 刘瑞红, 惠刚盈, 等. 林分空间结构参数N元分布及其诠释——以小陇山锐齿栎天然混交林为例[J]. 北京林业大学学报, 2019, 41(4):21-31. [33] 赵中华, 惠刚盈, 胡艳波, 等. 树种多样性计算方法的比较[J]. 林业科学, 2012, 48(11):1-8. doi: 10.11707/j.1001-7488.20121101 [34] 吴 恒, 朱丽艳, 王海亮, 等. 乔木树种分布格局和林分空间结构分析[J]. 林业资源管理, 2020, 42(1):54-61. [35] 姚 兰. 湖北木林子保护区15 hm2大样地森林群落结构及多样性[D]. 北京: 北京林业大学, 2016. [36] Holt A R, Gaston K J, He F. Occupancy-abundance relationships and spatial distribution: A review[J]. Basic and Applied Ecology, 2002, 3(1): 1-13. doi: 10.1078/1439-1791-00083 [37] He F, Legendre P, Bellehumeur C, et al. Diversity pattern and spatial scale: a study of a tropical rain forest of Malaysia[J]. Environmental & Ecological Statistics, 1994, 1(4): 265-286. [38] 薛卫星, 郭秋菊, 艾训儒, 等. 鄂西南天然林主要乔木树种物种组成及林分空间结构动态变化研究[J]. 西北植物学报, 2021, 41(6):1051-1061. [39] 薛卫星, 郭秋菊, 艾训儒, 等. 鹅掌楸天然林物种组成与林分空间结构特征研究[J]. 林业科学研究, 2021, 34(2):166-173. [40] 薛卫星. 鄂西南野生鹅掌楸群落结构与人工促进鹅掌楸更新方法研究[D]. 恩施: 湖北民族大学, 2021. [41] 廖彩霞. 樟子松人工林空间结构的研究[D]. 哈尔滨: 东北林业大学, 2007. [42] 惠刚盈, Klaus von Gadow,等. 结构化森林经营原理[M]. 北京: 中国林业出版社, 2016. [43] 李玮宜, 艾训儒, 姚 兰, 等. 鄂西南3个国家级自然保护区生物多样性评价[J]. 湖北林业科技, 2021, 50(2):18-22. [44] 张亚昊, 佃袁勇, 黄光体, 等. 不同演替阶段马尾松林林分空间结构对物种多样性的影响[J]. 生态学杂志, 2021, 40(8):2357-2365. [45] 朱 军, 湛 斌, 陈 霞, 等. 紫金山针阔混交林空间结构特征与植物多样性分析[J]. 中南林业科技大学学报, 2018, 38(1):75-80. [46] 宋厚娟, 叶 吉, 蔺 菲, 等. 取样面积对森林木本植物空间分布格局分析的影响[J]. 科学通报, 2014, 59(24):2388-2395. [47] Levin S A. The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture[J]. Ecology, 1992, 73(6): 1943-1967. doi: 10.2307/1941447 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4340
- HTML全文浏览量: 2583
- PDF下载量: 88
- 被引次数: 0
