


-
杨树(Populus L.)作为一种快速生长的木本植物,是我国最主要的速生丰产林树种之一[1]。杨树具有速生、易繁殖、适应性强和生产力高等特性,在木材加工、碳汇造林、制浆造纸和生物燃料等方面发挥着重要作用[2-4]。目前,我国许多杨树人工林种植园生长在氮素贫瘠的地区[5],易导致杨树人工林生产力低下,原材料利用率不高,产品质量差等问题。因此,开展杨树根系响应氮素分子调控机制的研究,可以实现杨树人工林氮素高效利用的经济效益,意义重大。
氮素是植物生长发育所必需的大量矿质营养元素之一,是蛋白质、核酸、叶绿素等重要物质的组成成分[6]。植物根系是吸收硝态氮或铵态氮的主要器官,良好的根系形态对植物氮素高效吸收利用至关重要[7]。有研究表明,植物可通过对外界不同氮形态响应做出反应,从而改变根系形态并逐渐做出相应的适应[8-12]。Wei等研究表明,杨树根系形态对低硝态氮处理响应的初期表现是加快增加主根长度,随后加速侧根增殖,在低硝态氮处理2~4 d后,根系生物活性显著增加,其根长和侧根数显著高于对照[5]。类似的动态响应在玉米(Zea mays L.)中也有报道[13]。上述研究结果表明:植物根系有能够根据不同氮形态供应而改变其形态的能力[14]。
除植物根系形态发生变化外,分子水平调节在植物响应不同氮形态吸收同化过程中也发挥着重要作用,特别是来自具有调控功能的miRNAs的调节[15-16]。miRNAs是一类广泛存在于真核生物中,长约18~24个碱基(nt)的高度保守小分子RNAs[17]。利用小RNA高通量测序技术并结合生物信息学分析,前人研究中已鉴定出大量参与氮素吸收同化响应相关的miRNAs[16, 18-19],其中,部分miRNAs也参与了植物根生长和发育过程[16, 20-21]。如在拟南芥( Arabidopsis thaliana (L.) Heynh.)根中柱鞘细胞中,5 mmol·L−1硝态氮可以抑制miR167的表达,从而提高靶基因auxin response factor 8 (ARF8)的表达水平,促进拟南芥 侧根起始和随后的萌发[20,22]。在水稻(Oryza sativa L.)中,miR396的靶基因growth-regulating factor 4 (GRF4) 能够促使水稻根系吸收铵态氮从而改变水稻根系结构[23];而miRNA调控机制在不同氮形态处理杨树根尖上存在怎样的差异表达模式尚不清楚,值得深入研究。
综上所述,本研究以灰杨(Populus × canescens)根尖为试材,利用小RNAs高通量测序技术,分析根尖差异表达的miRNAs,并结合降解组和转录组测序结果,分析miRNAs靶基因的差异表达模式,阐明杨树根尖对硝态氮或铵态氮处理条件下miRNAs及其靶基因的表达调控机制。本研究成果可为培育氮素吸收利用效率高的林木奠定理论基础,具有理论与实践意义。
-
将灰杨组培苗在组培室 (白天/黑夜温度25℃/18℃,相对湿度50%~60%,光照长度14 h,光照强度150 µmol·m−2·s−1) 中培养4周。随后,将生根的灰杨植株种植在10 L的花盆里进行砂培。每株植株隔天浇50 mL Long Ashton (LA)营养液(0.5 mmol·L−1 NH4NO3、0.5 mmol·L−1 KCl、0.9 mmol·L−1 CaCl2、0.3 mmol·L−1 MgSO4、0.6 mmol·L−1 KH2PO4、42 µmol·L−1 K2HPO4、10 µmol·L−1 Fe-EDTA、2 µmol·L−1 MnSO4、10 µmol·L−1 H3BO3、7 µmol·L−1 Na2MoO4、0.05 µmol·L−1 CoSO4、0.2 µmol·L−1 ZnSO4和0.2 µmol·L−1 CuSO4, pH 5.5)。砂培植株在温室(与组培室气候条件相同)中培养14 d后,具有相似高度(株高约30 cm)的植株被选择并分成2组(每组24株植物)。水培1~2周后,将这2组植株移植到分别添加等量氮供应的0.5 mmol·L−1 NaNO3 (硝态氮)和0.5 mmol·L−1 NH4Cl (铵态氮) 代替0.5 mmol·L−1 NH4NO3 的改良LA营养液中进行水培,培养时间为10 d。在处理10 d后对其进行根系形态特征分析,然后选取根尖40 mm进行收获。所有收获样品迅速置于液氮中,放于−80℃冰箱备用。为了获得足够的材料进行进一步分析,将8株植物的样本均匀混合作为一个重复,每种处理水平3个生物学重复。
-
试验苗进行硝态氮或铵态氮处理后,对灰杨根系形态构型进行监测,包括测量根系长度,二级可见侧根发生位置等指标。
-
不同处理水平根尖小RNAs测序按照Illumina公司提供的标准步骤执行。简单来说,将2种不同氮形态处理下的样品,每个处理3个重复分别利用RNA提取试剂盒提取总RNAs (TRK1001, 联川生物技术公司, 杭州, 中国),将提取的总RNAs质量检测后,利用miRNA特有的5′端磷酸基团和3′端羟基基团属性,将一个腺苷化单链DNA 3′接头和5′接头相继连接到小RNAs上,通过与3′端互补的RT引物进行反转录反应,对反转录产生的cDNA序列进行PCR扩增。随后通过6% polyacrylamideTris-borate-EDTA切胶回收的方式对140~160 bp长度范围的PCR产物进行胶回收,最后利用杭州联川生物技术公司Illumina HiSeqTM 2000高通量测序技术进行小RNA测序分析。
-
将测得的RNAs序列使用联川生物技术公司开发的软件进行分析。首先,去除低质量的序列,保留干净的唯一的序列进行分析,将剩余序列比对RFam数据库和重复序列数据库用以去除rRNA、tRNA、snRNA和snoRNA等,过滤后的数据使用已有的灰杨基因组数据库(
http://aspendb.uga.edu/index.php/databases/spta-717-genome )、miRNAs数据库(http://www.mirbase.org/ )进行比对,筛选出已知miRNAs。针对新的miRNAs序列,利用miRNA前体的标志性发夹结构,通过Mfold软件(http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi )分析RNA二级结构,鉴定新的miRNAs。 -
把等量的每个处理水平的3个重复样品合并,共2个处理水平的合并样品利用联川生物技术公司Illumina HiSeq2500技术平台进行降解组测序,采用CleaveLand软件(3.0.1版本)预测miRNAs的靶基因。为了进一步挖掘靶基因的表达水平,6个cDNA文库被构建,利用转录组数据Hiseq测序平台分析相关miRNAs靶基因的表达模式,同时结合生物信息学Kyoto Encyclopedia of Genes and Genomes (KEGG)软件和MapMan软件,分析差异表达miRNAs靶基因的生物学功能。
-
根据灰杨数据库,利用Primer Premier 3.0软件设计引物并由上海生工生物工程有限公司合成相关miRNAs和靶基因的引物。5.8s rRNA和Actin 分别作为miRNAs和靶基因的内参基因。序列见表1。
表 1 RT-qPCR引物序列
Table 1. Primers used for RT-qPCR
miRNAs/靶基因ID
miRNAs/targets ID引物序列5′-3′
Primer sequence 5′-3′PC-5p-35885_222 TCTACTGAAAGTGAAAGCTAT ptc-MIR166e-p5 GGATTGTCGTCTGGTTCGATG ptc-MIR166m-p5_1ss9TC GAATGTCGCCTGACTCGAGAC ath-miR166e-5p_2ss4AC16AT GGACTGTTGTCTGGCTCGAGG mtr-miR166e-5p_2ss4AC10GT GGACTGTTGTCTGGCTCGAGG ptc-MIR1450-p3_2ss19AG21TC CGGTCAGGTTACACAAGCGAC ptc-miR6478_R+2_2ss5CT21GA CCGATCTTAGCTCAGTTGGTAGA ptc-MIR2111a-p3 GTCCTTGGGTTGCAGATTACC ptc-MIR2111b-p3 GTCCTTGGGTTGCAGATTACC vvi-MIR399d-p3_1ss13GA TCTGCCAAAGGAAATTTGCTC ptc-miR396g-5p TTCCACGGCTTTCTTGAACTT ath-miR858a_L-2R+1 TCGTTGTCTGTTCGACCTTG ptc-miR476b_2ss8TC17CT TAGTAATCCTTCTTTGTAAAA ptc-miR395b CTGAAGTGTTTGGGGGAACTC PC-3p-42422_177 TTTATGCGTTTTTGCCCCTCG ptc-miR399b TGCCAAAGGAGATTTGCCCGG URT primer 由Mir-X miRNA First-Strand Synthesis和SYBR RT-qPCR试剂盒提供 U6 F′/ R′ 均由Mir-X miRNA First-Strand Synthesis和SYBR RT-qPCR试剂盒提供 protein binding F′ TGAGATCAGCGAGGGACAAA protein binding R′ AAGGTTTGAGTGAGCAAGGC GRF9 F′ TCCCTCCCTCTCTCTGTCAA GRF9 R′ CATGATCACCAGCACCAATC GRF1 F′ TGCTGTTTCTTGGCCTGAAC GRF1 R′ CAATACCACCACCTCCACCT APS2 F′ CCAGAACCTGTCCCAGCTTA APS2 R′ GCTCCCAACACACATGCTAG phosphate 2 F′ AGGAAGAGCTGAGGGAGAGA phosphate 2 R′ CCTGTCGAACCACCCTTTTG MYB4 F′ CGCGTGTGGTGATTGTAGAG MYB4 R′ GTTGCCTCTCTTGATGTCCG ATP binding F′ GCGGGTGAATGAGGCTTTAG ATP binding R′ GCCTCTGAAACCTTGCCTTC 5.8s rRNA F′ CTCGGCTCTCGCATCGATGA 5.8s rRNA R′ AGACGTGCCCTCGACCAAGA Actin F′ CCCATTGAGCACGGTATTGT Actin R′ TACGACCACTGGCATACAGG -
使用Statgraphics软件(STN, St Louis, MO, USA)进行数据统计分析。在进行统计分析之前,先对数据进行正态性检验。所有数据均采用单因素方差分析(ANOVA),以P < 0.05作为统计意义上的显著水平。
利用transcripts per million(TPM)对miRNAs表达水平进行定量。使用miRNA在铵态氮处理下的TPM除以硝态氮处理下的TPM来计算miRNA的差异倍数(FC)。差异表达miRNAs筛选阈值为P < 0.05。
采用Fragments Per Kilobase of exon per Million mapped (FPKM)对靶基因mRNAs表达水平进行定量。基于FPKM值,使用Ballgown package计算mRNAs的差异表达水平。使用mRNAs在铵态氮处理下的FPKM除以硝态氮处理下FPKM来计算这个基因的差异倍数(FC)。差异表达mRNAs筛选阈值为log2(FC) ≥ 1或 ≤ −1,P < 0.05。
对于RT-qPCR的测定,将qPCR得到的Ct值进行归一化,计算miRNAs及其靶基因的相对表达量。
-
由图1可知:在不同氮形态处理条件下灰杨根的形态特征具有显著差别。测量表明,硝态氮处理下的根长为27.5 cm,比铵态氮处理的14.3 cm几乎长1倍(p < 0.001),但2种处理水平下其二级侧根发生位置比较一致。这说明不同氮形态处理对根系生长发育的影响不同。选取二级侧根形成前的根尖部分(40 mm)进行后续分子实验。
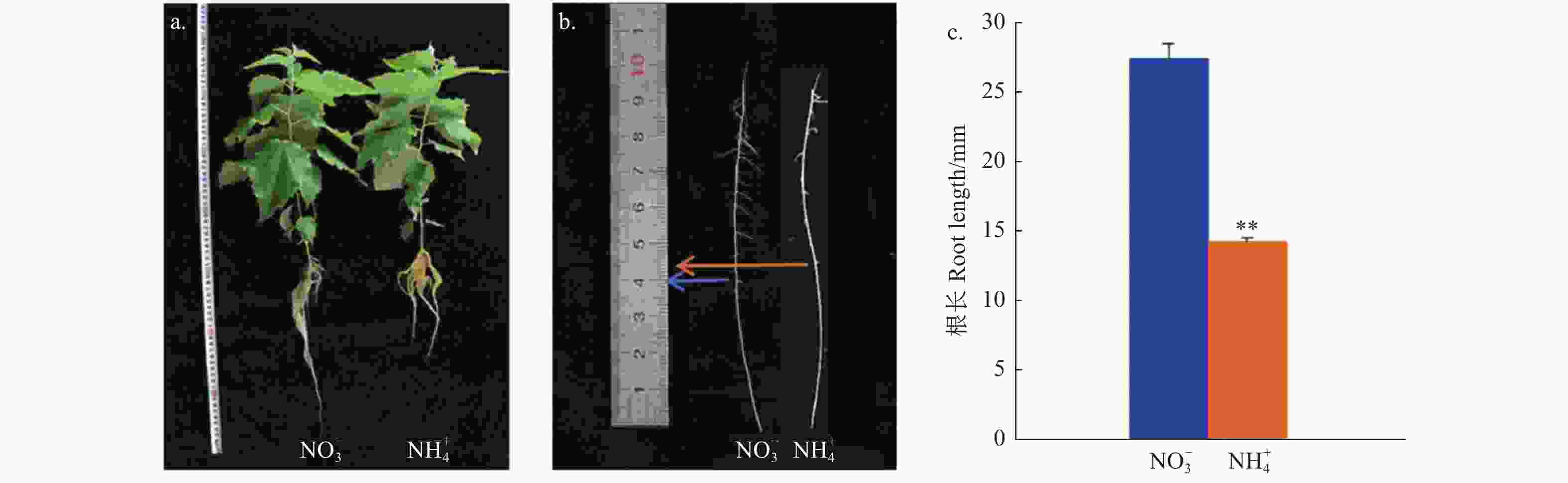
图 1 不同氮形态处理灰杨根系的表型
Figure 1. Morphological parameters of P. × canescens roots with different nitrogen forms
-
基于上述根尖形态特征分析的结果,进行高通量小RNAs测序分析。测序结果显示:2个处理水平的测序分别得到的原始序列读数为5 752 458和7 800 546,进行数据处理和杂质的过滤,去掉低质量序列和3′、5′缺失序列等,分别获得1 996 697和2 608 788条干净序列(表2)。长度在18~25 nt的序列被保留下来做进一步研究。
表 2 小RNA文库测序数据统计
Table 2. Distribution of small RNAs in different categories
类别
category硝态氮
NO3−铵态氮
NH4+Total Unique Total Unique raw reads 11 320 294 2 909 511 12 316 643 3 519 988 valid reads 5 752 458 1 996 697 7 800 546 2 608 788 Rfam 668 833 14 694 817 912 16 666 mRNA 169 342 2 779 343 819 5 169 other Rfam RNA 28 770 1 020 40 909 1 157 注:Rfam代表除miRNAs外许多常见的非编码RNA家族的集合。
Note: Rfam: collection of many common non-coding RNA families except miRNAs. -
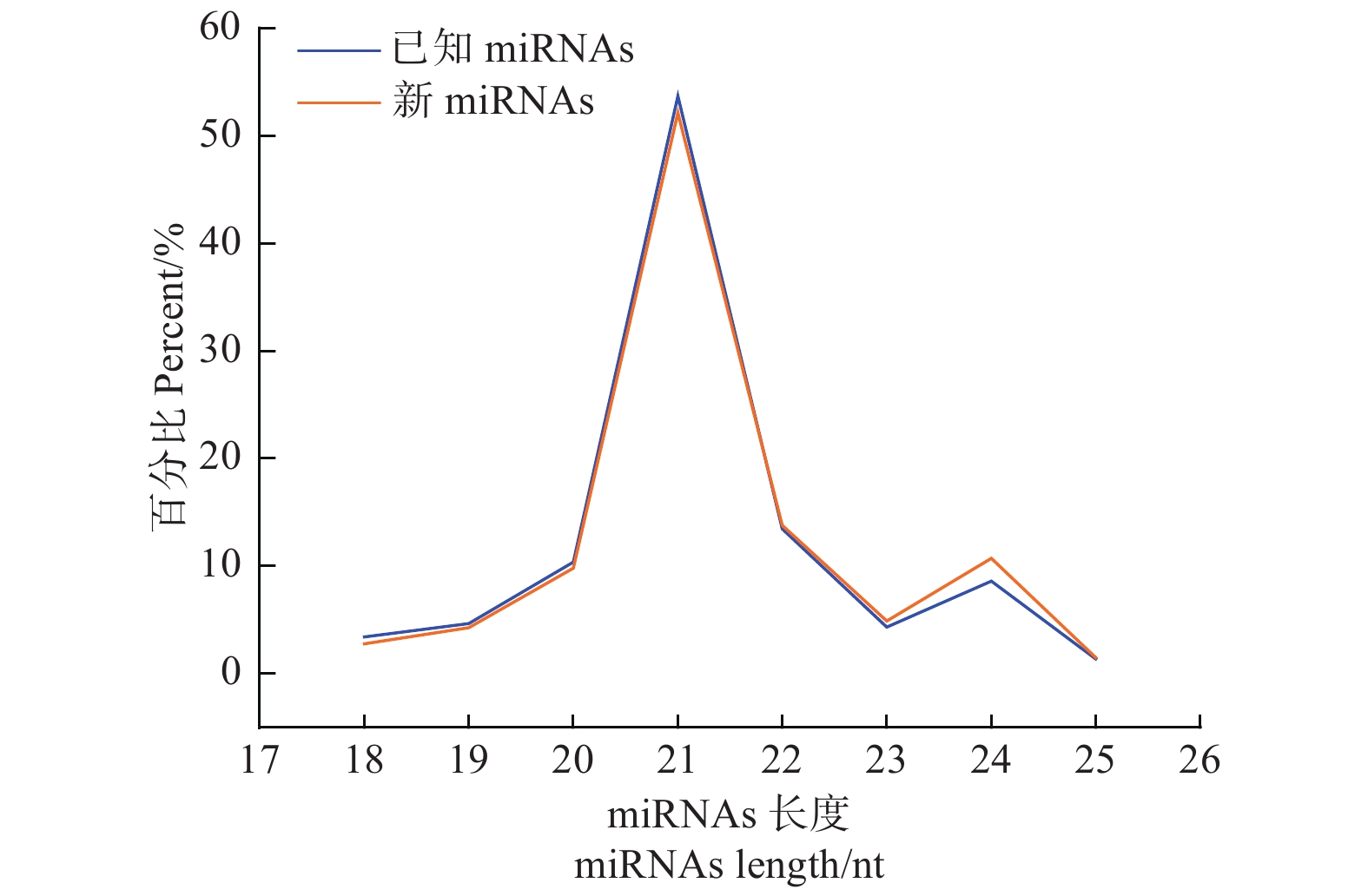
在不同氮形态处理下,共鉴定灰杨523个已知的miRNAs和42个新的miRNAs (表3)。研究表明,植物miRNAs长度大多在21 nt或22 nt,统计本实验2个处理水平miRNAs文库中所有18~25 nt的miRNAs长度,21-nt miRNA出现频率最高 (图2),这说明该测序数据可靠。
表 3 基于高通量测序技术鉴定灰杨根尖已知和新的miRNAs数量
Table 3. The number of identified known and novel miRNAs in root tips of P. × canescens based on high-throughput sequencing
样本
Samples已知/新
miRNAs known/
novelmiRNAsmiRNA前体
miRNA PremiRNA成熟体
miRNA mature硝态氮 NO3− 已知 Known 460 469 新的 Novel 37 39 铵态氮 NH4+ 已知 Known 502 514 新的 Novel 68 69 合计 Total 已知 Known 523 592 新的 Novel 42 42 
图 2 灰杨根尖已知和新miRNAs长度分布
Figure 2. Lengths of known and novel miRNAs in root tips of P. × canescens.
-
为了解不同氮形态处理下,灰杨根尖对miRNAs的响应,对筛选出来的灰杨中已知miRNAs和新miRNAs的表达量进行计算。分析表明:灰杨根尖共有96个miRNAs (p < 0.05)在不同氮形态处理下展示出不同的表达模式,其中,包括33个已知miRNAs家族和9个新的miRNAs。相比较于硝态氮处理,铵态氮处理有44个上调表达的miRNAs和52个下调表达的miRNAs (图3a),其中29个显著差异表达的miRNAs (p < 0.001)见表4。
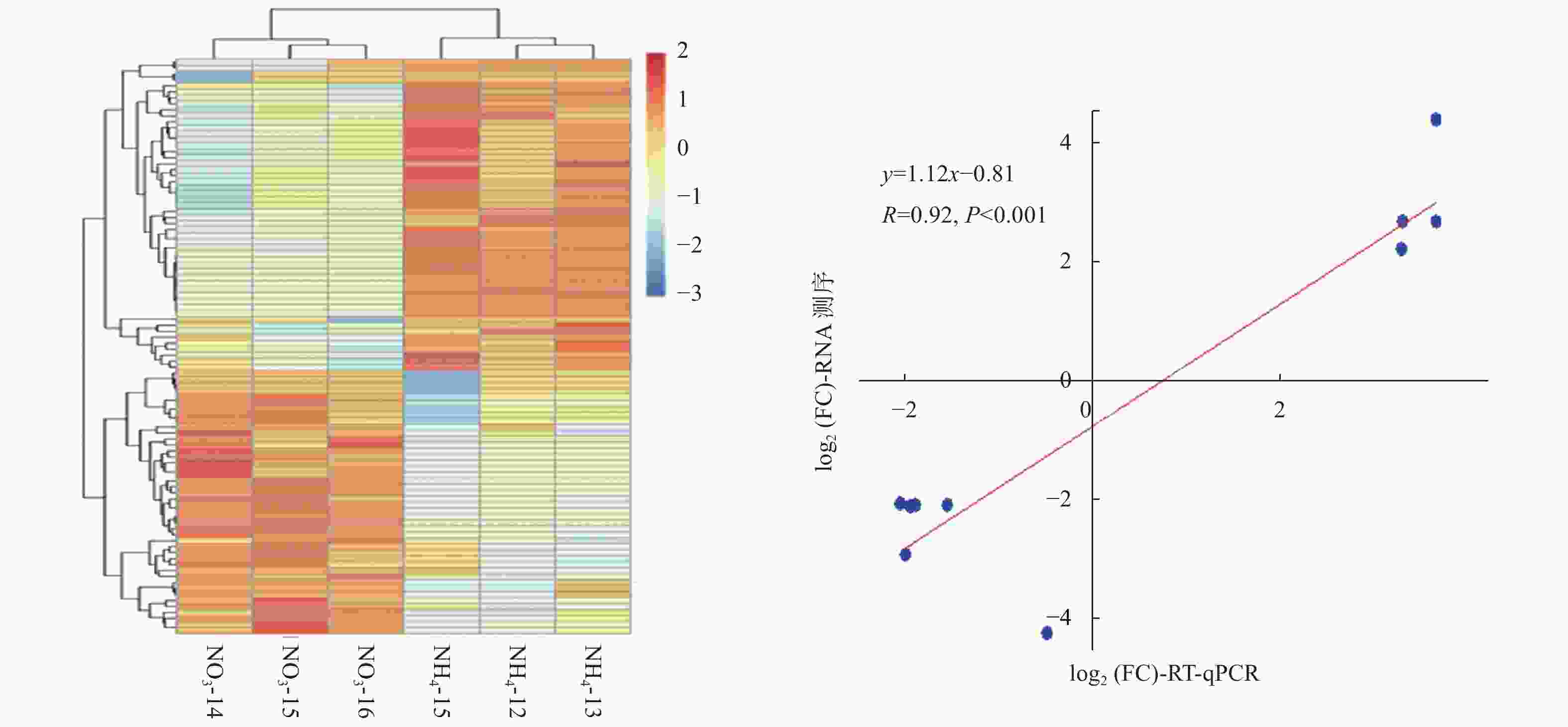
图 3 不同氮形态处理条件下灰杨根尖差异表达的miRNAs
Figure 3. Significantly differentially expressed miRNAs in root tips of P. × canescens under different nitrogen forms treatments
表 4 29个显著差异表达miRNAs
Table 4. twenty-nine significantly differentially expressed miRNAs
miRNAs ID
miRNAs IDmiRNAs成熟体序列
sequence of miRNAs上调/下调
Up/downlog2(FC) P-value 是否RT-qPCR验证
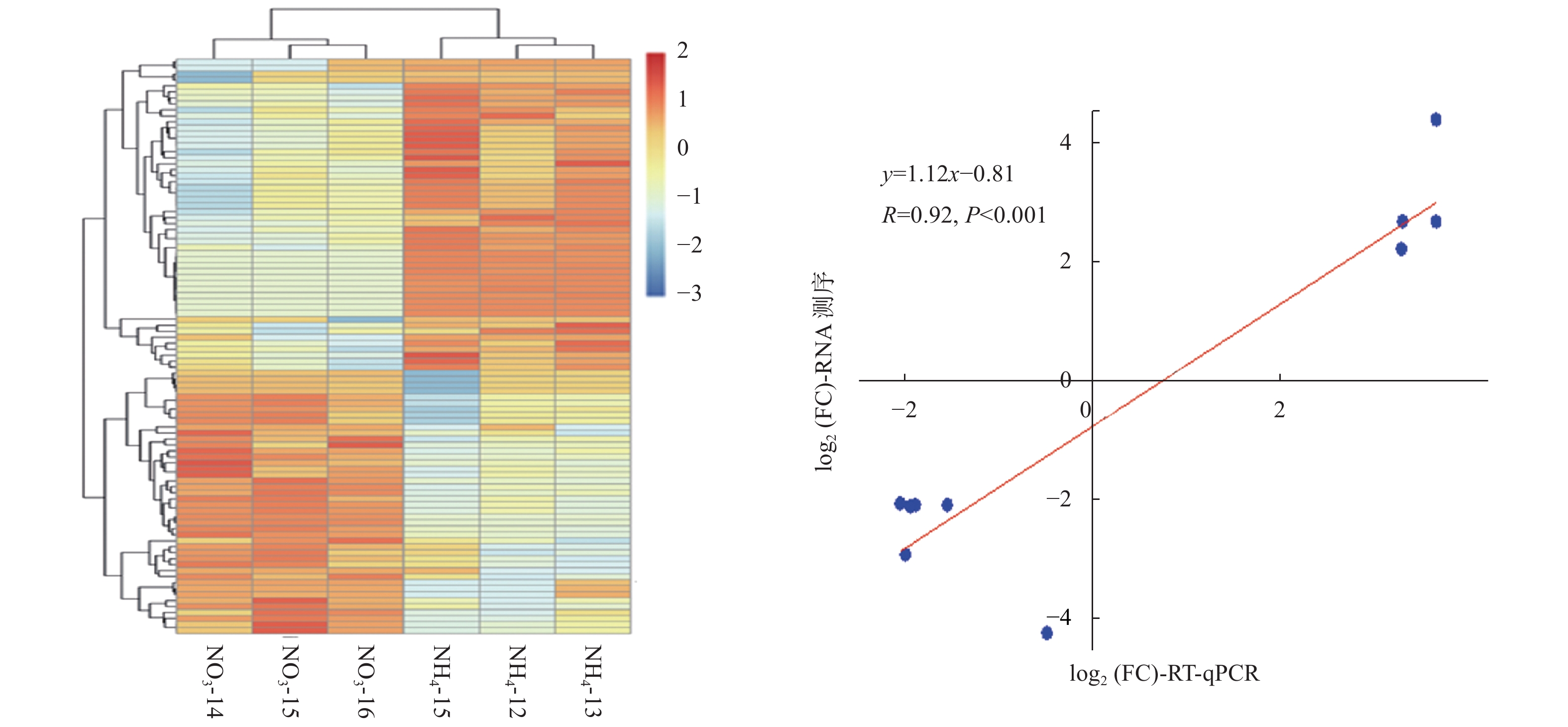
RT-qPCR verifiedPC-5p-35885_222 TCTACTGAAAGTGAAAGCTAT down −4.50 0.003 21 是 ptc-MIR166e-p5 GGATTGTCGTCTGGTTCGATG down −3.15 0.007 14 是 ptc-MIR166m-p5_1ss9TC GAATGTCGCCTGACTCGAGAC down −2.32 0.008 37 是 ath-miR166e-5p_2ss4AC16AT GGACTGTTGTCTGGCTCGAGG down −2.30 0.003 18 是 mtr-miR166e-5p_2ss4AC10GT GGACTGTTGTCTGGCTCGAGG down −2.30 0.003 18 是 ptc-MIR1450-p3_2ss19AG21TC CGGTCAGGTTACACAAGCGAC down −2.27 0.006 95 是 mtr-miR171g_R+1 CGAGCCGAATCAATATCACTCC down −2.25 0.008 97 否 ptc-MIR169ae-p3_1ss20GA TGGGCAAGCACCTTGGCTAAC down −1.55 0.009 87 否 mtr-miR166c_2ss1TG20TC GCGGACCAGGCTTCATTCCCC down −1.30 0.003 29 否 ptc-MIR6438b-p5_1ss11AG TCGACTGAAAGTGAAAGCTAT down −0.96 0.003 68 否 PC-5p-166_26156 TAGCCAAGAGCGACTTGCCCA down −0.76 0.007 70 否 ptc-MIR396d-p3 CTCAAGAAAGCCGTGGGAGA down −0.54 0.002 63 否 ptc-miR166n_L+2R-2 TCTCGGACCAGGCTTCATTCC down −0.53 0.006 92 否 ptc-miR1447_1ss21TC CAGAATTGCAGTGCCTTGATC up 0.43 0.004 66 否 ptc-miR476b_2ss8TC17CT TAGTAATCCTTCTTTGTAAAA up 0.60 0.002 85 否 ptc-MIR1447-p5_1ss6GT AATCATGGCACTGCAATTCTA up 0.68 0.005 60 否 ptc-miR319e_R+1 TTGGACTGAAGGGAGCTCCTT up 1.00 0.001 08 否 vvi-miR172d_L-2R+1 AGAATCTTGATGATGCTGCATT up 1.05 0.000 50 否 ptc-miR530a_R+1 TGCATTTGCACCTGCACCTTA up 1.65 0.005 57 否 ptc-miR2111a TAATCTGCATCCTGAGGTTTG up 1.67 0.006 39 否 PC-3p-42422_177 TTTATGCGTTTTTGCCCCTCG up 1.95 0.003 47 否 ptc-miR6478_R+2_2ss5CT21GA CCGATCTTAGCTCAGTTGGTAGA up 2.11 0.008 94 是 ptc-MIR2111a-p3 GTCCTTGGGTTGCAGATTACC up 2.58 0.001 48 是 ptc-MIR2111b-p3 GTCCTTGGGTTGCAGATTACC up 2.58 0.001 48 是 vvi-MIR399d-p3_1ss13GA TCTGCCAAAGGAAATTTGCTC up 4.33 0.000 15 是 gma-miR166m_L+1_1ss5AC GCGGCCCAGGCTTCATTCCCC down −inf 0.003 18 否 PC-3p-82747_59 CCGATCATTATTGGTGACCTT down −inf 0.004 57 否 ptc-miR171k_1ss12CT GGATTGAGCCGTGCCAATATC down −inf 0.001 50 否 ptc-MIR319b-p5_1ss4TC TAGCTACCGACTCATTCATCCA down −inf 0.004 30 否 同时,随机挑选了10个显著差异表达miRNAs,利用RT-qPCR进一步证实了测序结果的可靠性 (图3b)。虽然miRNAs表达水平的差异与测序得到的差异并不一致,但上调或下调表达的趋势是相似的。2种技术在比率方面的差异是由2种技术的算法和灵敏度的本质不同造成的。总体而言,本研究测序结果的可靠性较高。
-
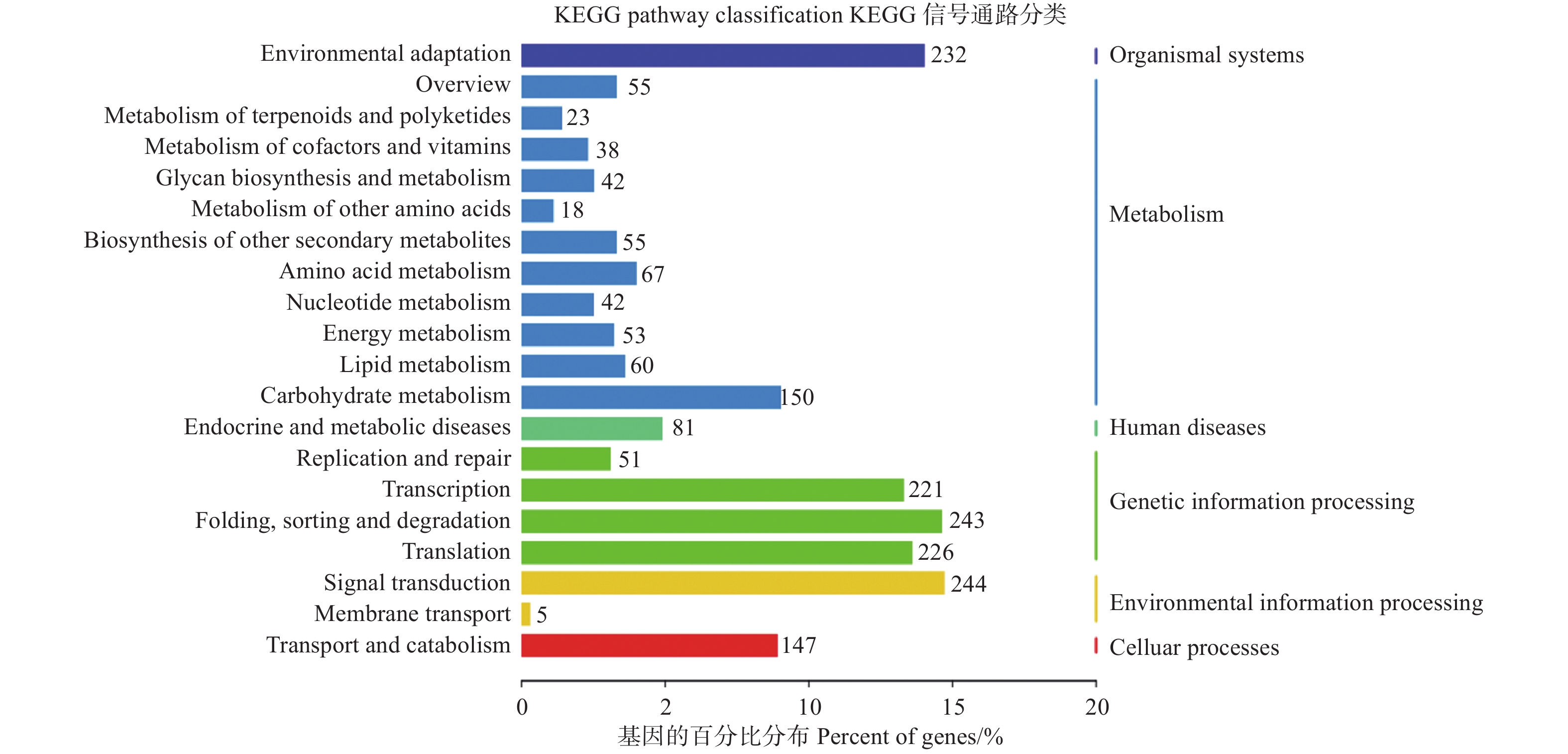
为了进一步了解miRNAs的作用,分别使用等量的总RNA样本对硝态氮和铵态氮2个混合降解组池进行测序,以识别miRNAs靶基因。结果表明:分别有67.13%和65.37%去除冗余的reads可以比对到灰杨数据库中。从降解组中共鉴定出2729个靶基因位点,其中,2104个靶基因显著差异表达。为了进一步了解上述靶基因的功能,对识别出的2104个显著差异表达靶基因进行KEGG通路分析 (图4);同时MapMan分析进一步揭示了这些差异靶基因在氮代谢过程中以及植物生长发育过程中参与的各种生物学过程,如ptc-miR166i-p5有4个靶基因potri.015g017500.1/2/3/4均属于NADH依赖的谷氨酸合成酶(NADH-dependent glutamate synthase family) 蛋白参与氮代谢过程。gra-MIR8723b-p3_2ss6TC21TC有3个靶基因Potri.003G111500.1/2/3均作为硝态氮转运体基因(nitrate transmembrane transporter1.1/NRT1.1) 在硝态氮运输中起到关键作用。ptc-MIR6462a-p5_1ss14TC的靶基因Potri.005G079200.1、gma-miR6300_1ss5TG和gma-miR6300_R+1_1ss5TG的靶基因Potri.001G300900.1/2均编码氨基酸运输或代谢相关蛋白,也对氮代谢有所响应 (表5);同时,与植物生长发育相关的一些miRNA的靶基因也相继被发现。如miR164家族的多个靶基因均属于NAC基因家族成员,其在植物生长发育过程中发挥重要功能 (表5)。
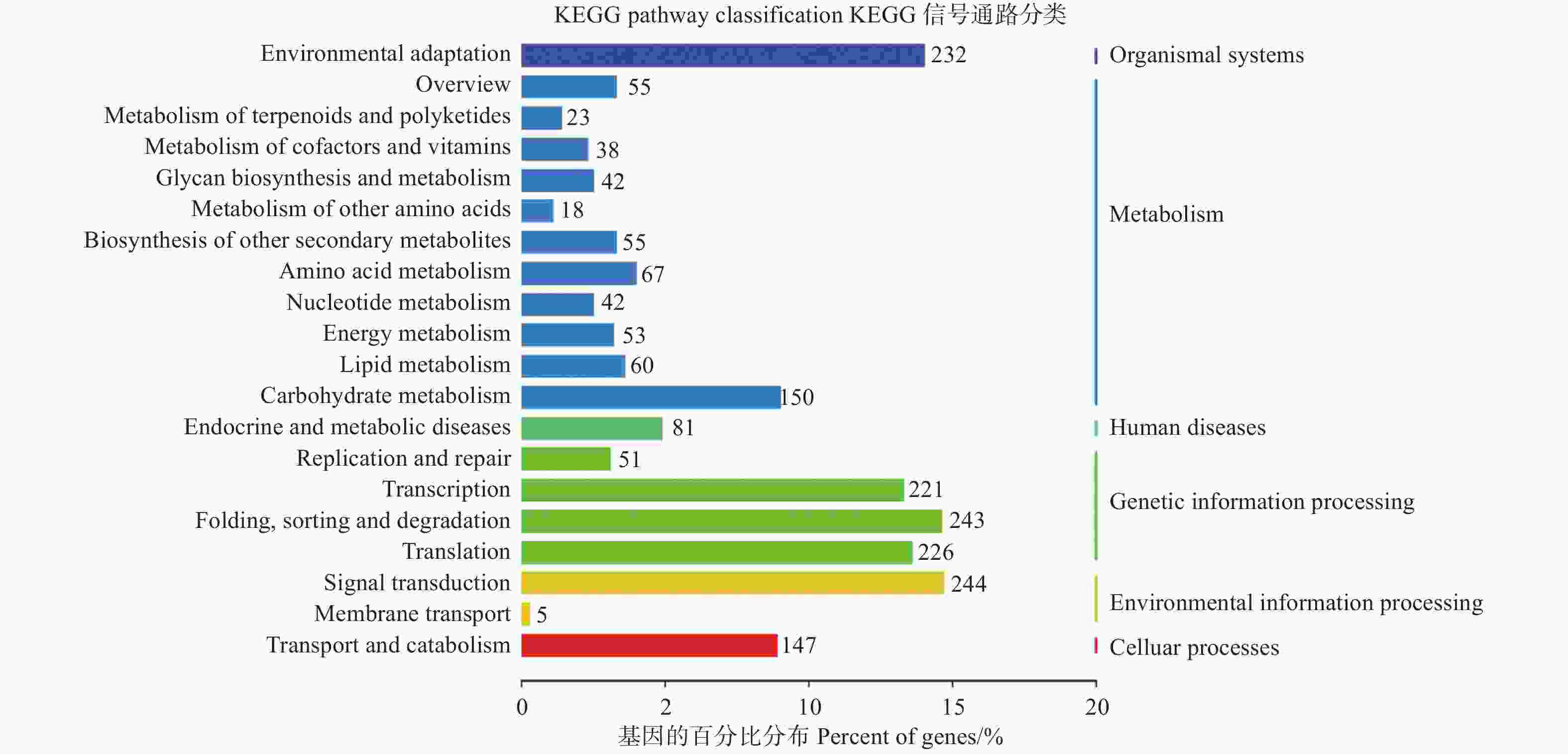
图 4 KEGG通路分析鉴定差异表达靶基因
Figure 4. KEGG pathway analysis of significantly differentially expressed target genes
表 5 灰杨根尖氮代谢以及生长发育相关差异表达靶基因
Table 5. Significantly differentially expressed target genes related to nitrogen metabolism and growth and development of P. × canescens root tips
miRNAs ID
miRNAs ID靶基因ID
Targets ID功能注释
Functional annotationptc-MIR166i-p5 Potri.015G017500.4 glutamate synthase (NADH) ptc-MIR6462a-p5_1ss14TC Potri.005G079200.1 aminotransferase class Ⅰ and Ⅱ family protein gma-miR6300_1ss5TG Potri.001G300900.1 embryo sac development arrest 9 gma-miR6300_R+1_1ss5TG Potri.001G300900.1 embryo sac development arrest 9 gma-miR6300_1ss5TG Potri.001G300900.2 embryo sac development arrest 9 gma-miR6300_R+1_1ss5TG Potri.001G300900.2 embryo sac development arrest 9 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.1 nitrate transmembrane transporter,NRT1.1 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.2 nitrate transmembrane transporter,NRT1.1 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.3 nitrate transmembrane transporter,NRT1.1 mtr-miR164a_1ss21AG Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) ptc-miR164a Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) vvi-miR164a_R+1 Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) ptc-MIR7813-p5_2ss13AG20CT Potri.017G016700.1 NAC domain-containing protein 73 (NAC073) cca-miR164 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) -
为了研究不同氮形态处理下,杨树根尖差异表达miRNAs与靶基因调控网络,利用上述结果进行联合分析。表6表明:有23个差异表达靶基因 (log2(FC) ≥ 1或 ≤ −1,P < 0.05) 隶属于5个miRNA家族和一个新的miRNA (P < 0.05)被鉴定,且miRNA-靶基因呈现负相关调控关系,表明这些靶基因可能是通过miRNAs转录抑制作用而被降解。
表 6 不同氮形态处理条件下灰杨根尖差异表达miRNA-靶基因
Table 6. Significantly differentially expressed miRNAs-target pairs in root tips of P. × canescens under different nitrogen forms treatments
miRNAs ID
miRNAs ID上调/下调
Up/down靶基因ID
Targets IDlog2(FC)
log2(FC)靶基因注释信息
Target Functional annotationPC-3p-42422_177 up Potri.003G152300.2 −3.177 77 NA PC-3p-42422_177 up Potri.003G152000.1 −3.177 77 NA PC-3p-42422_177 up Potri.003G152300.1 −3.177 77 NA ptc-miR396g-5p down Potri.014G012800.1 2.346 49 growth-regulating factor 1 (GRF1) ptc-miR396g-5p down Potri.013G077500.1 inf growth-regulating factor 5 (GRF5) ptc-miR396g-5p down Potri.001G132600.1 1.613 36 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.014G071800.1 2.436 61 growth-regulating factor 9 (GRF9) ptc-miR396g-5p down Potri.003G100800.4 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.3 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.1 1.00154 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.2 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.007G007100.1 1.463 15 growth-regulating factor 1 (GRF1) ptc-miR395b up Potri.008G159000.3 −1.159 23 ATP sulfurylase 1 ptc-miR395b up Potri.010G081200.2 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.010G081200.1 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.010G081200.3 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.008G159000.2 −1.159 23 ATP sulfurylase 1 ptc-miR395b up Potri.008G159000.1 −1.159 23 ATP sulfurylase 1 ptc-miR399d up Potri.011G052600.1 −1.157 53 phosphate 2 ptc-miR399b up Potri.011G052600.1 −1.157 53 phosphate 2 ath-miR858a_L-2R+1 down Potri.006G221800.1 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ath-miR858a_L-2R+1 down Potri.006G221800.2 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ath-miR858a_L-2R+1 down Potri.006G221800.3 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ptc-miR476b_2ss8TC17CT up Potri.006G242500.1 −1.285 67 ATP binding; nucleic acid binding; helicases 通过RT-qPCR实验验证了其中7个miRNA-靶基因的表达模式,结果显示:差异表达miRNAs及其靶基因RT-qPCR分析与高通量测序产生的表达模式相似,且表达模式呈负相关关系 (图5)。
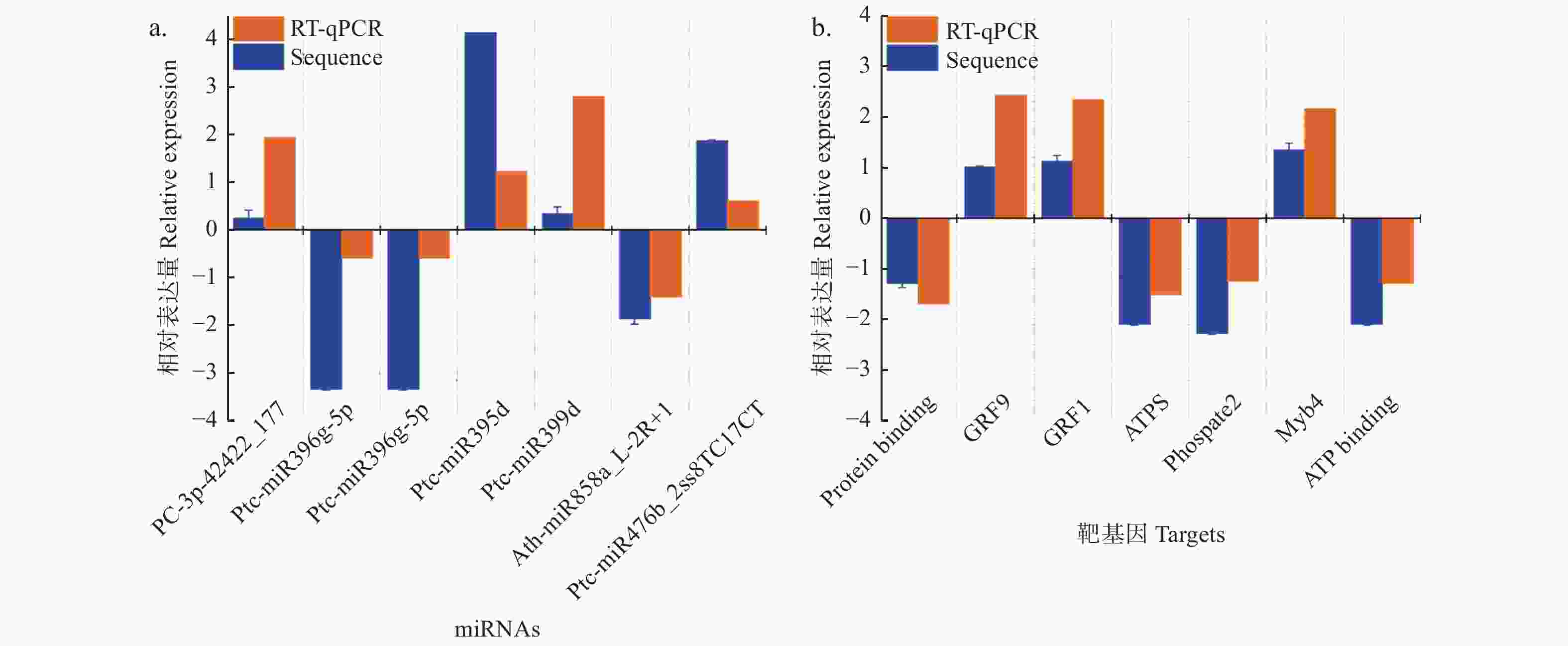
图 5 不同氮形态处理下miRNAs-靶基因对差异表达分析
Figure 5. Validation of significantly differentially expressed miRNAs and their targets under different nitrogen forms treatments by sRNA-seq and RT-qPCR
-
研究表明,基因的表达受miRNAs调节,不同氮形态可能导致miRNAs及其调控的靶基因差异表达,从而影响植物根尖生长和发育[24]。本研究利用高通量测序平台研究不同氮形态处理下,杨树根尖差异miRNAs和靶基因的表达模式。
在不同氮形态处理10 d后,收获杨树根尖40 mm区段材料,经过小RNA高通量测序分析,获得96个显著差异表达miRNAs,其中,vvi-MIR399d-p3_1ss13GA上调表达倍数最高。前人研究表明,miR399参与调控水稻多种营养饥饿反应[25]。在本研究中,miR399靶基因为泛素化连接酶,且与miR399呈负相关关系。有研究证明,泛素分子主要通过泛素活化酶、泛素结合酶和泛素化连接酶将靶蛋白泛素化,泛素化的蛋白最后被26S蛋白酶体识别和降解[26],该过程对植物营养缺乏等非生物胁迫发挥着重要作用[27]。因此,可以推测miR399及其靶基因泛素化连接酶在不同氮形态胁迫响应中发挥着重要的调控作用。在本试验条件下,部分差异表达miRNAs与杨树根尖生长发育相关,这与在水稻中的研究结果相似[15]。如miR171家族成员,其靶基因之一是DELLA protein RHT-1。DELLA蛋白家族是GA信号途径中的负调控因子,可以抑制GA途径的基因表达从而抑制植物生长[28]。另有研究表明,DELLA-GRF4介导植物生长与氮代谢的协同调控机制,DELLA的积累不仅导致水稻生长矮化,而且降低了氮素利用效率[23]。在本研究中,相比较于硝态氮处理,铵态氮处理诱导miR171靶基因DELLA表达,从而导致杨树根尖发育受到抑制,根长变短。该研究说明杨树根尖部分差异表达miRNAs及其靶基因有通过对不同氮形态处理的响应而改变其根尖形态结构的能力。
对差异靶基因KEGG通路分析表明,一些靶基因富集参与氮代谢途径,如NADH-dependent glutamate synthase family protein和nitrate transmembrane transporter (NRT1.1)。NADH-dependent glutamate synthase family protein酶作为铵态氮吸收同化过程中的关键酶对植物幼苗期根系的初级铵同化很重要[29]。另有研究表明,在水稻NADH-gogat1突变体中,相比较于1 mmol·L−1硝态氮处理,1 mmol·L−1铵态氮处理抑制其根系的生长[29],该研究结果与本研究结果相一致。相比较于硝态氮处理,铵态氮处理抑制靶基因NADH-dependent glutamate synthase family protein表达,从而导致杨树根尖发育受到抑制,这表明NADH-dependent glutamate synthase family protein可能在植物根尖响应不同氮形态时发挥重要的作用。NRT1.1作为硝态氮转运体对硝态氮的反应范围广泛,从初始硝酸盐反应到长期发育变化,且对侧根发育的影响发挥着重大作用,即NRT1.1在低浓度硝态氮条件下抑制侧根生长,而在高浓度硝态氮条件下促进植物侧根生长[30],该结论与本文研究结果一致。在本研究中,相比较于硝态氮处理,铵态氮处理下靶基因NRT1.1上调表达,抑制了杨树侧根的伸长生长。另外,对灰杨根尖氮代谢以及生长发育相关差异表达靶基因进行MapMan分析表明,NAC基因家族出现频率最多。在拟南芥中,NAC4基因作为protein auxin signaling F-BOX 3 (AFB3)的下游基因,参与硝态氮响应,从而影响根系形态结构[31]。在小麦中,TaNAC2-5A可直接结合硝酸盐转运体和谷氨酰胺合成酶基因的启动子区域,过表达TaNAC2-5A可促进小麦根系生长和硝酸盐流入[32]。在本研究中,相比较于铵态氮处理,硝态氮处理中NAC1基因高表达,从而促进根生长,这一结论与前人的研究结果相一致。
在联合分析中,共发现24对miRNA-靶基因呈负相关关系,其中,miR396-GRF是联合分析中出现最多的一组关系对。GRF4作为miR396的靶基因能够驱动水稻根对铵态氮的吸收[23],同时GRF4能够驱动硝态氮转运体的转录水平,例如NRT1.1B和NRT2.3a、GRF4还能够驱动硝态氮同化酶基因nitrate reductase 1 (NIA1)、NIA3和nitrite reductase 1 (NiR1)的合成从而去调节氮代谢,且在水稻中,GRF4突变抑制植物生长[23]。在本研究中,相比较于硝态氮处理,在铵态氮处理中miR396下调表达,其靶基因GRF1/2/5/9均上调表达,从而促进杨树根生长,这一结论与前人的研究结果相一致。该结果说明miR396-GRF在不同氮形态胁迫响应中发挥着重要的调控作用。另外,表达差异最显著的靶基因为新miRNAPC-3p-42422_177的3个靶基因,但其功能未知,有待进一步研究证实。
-
本研究从2种不同氮形态处理杨树根尖miRNAs文库中鉴定出523个已知miRNAs和42个新miRNAs,其中,有96个miRNAs显著差异表达。与硝态氮处理相比,铵态氮处理下有44个miRNAs上调表达,52个下调表达,其中,miR396-GRF模块引起笔者的关注。研究表明,miR396-GRF模块可能通过响应不同氮形态调控杨树根系形态构型。该研究不仅可以为我国转基因杨树研究储备基因资源信息,还可以加快氮吸收能力强的杨树优质速生良种的培育。
基于硝态氮或铵态氮条件下杨树根尖miRNAs特征分析
miRNAs Analysis of Poplar Root Tips Treated with Nitrate- or Ammonium-Nitrogen
-
摘要:
目的 通过小RNAs高通量测序技术,在硝态氮或铵态氮处理下,对杨树根尖差异表达miRNAs及其靶基因调控机制进行了研究,分析和描述了杨树根尖生长和形态特征的规律,为进一步筛选和鉴定硝态氮或铵态氮吸收利用效率高的林木新品种提供依据。 方法 以灰杨水培苗根尖为研究对象,应用高通量测序技术进行小RNA文库构建,获得miRNAs表达谱,鉴定差异表达miRNAs,同时利用降解组测序技术鉴定miRNAs靶基因,并对差异表达靶基因进行功能注释以及代谢通路富集等分析。最后通过联合分析,阐明不同氮形态处理下,灰杨根尖miRNA-靶基因调控网络。 结果 从2种不同氮形态处理杨树根尖miRNAs文库中鉴定出523个已知miRNAs和42个新miRNAs,其中,有96个miRNAs显著差异表达。与硝态氮处理相比,铵态氮处理下有44个miRNAs上调表达,52个下调表达,其中,vvi-MIR399d-p3_1ss13GA上调倍数最大,新的miRNA PC-5p-35885_222下调最显著。对miRNAs靶基因进行KEGG通路富集分析表明,部分显著差异表达靶基因参与杨树根尖氮响应,从而影响杨树根尖生长发育和形态特征。利用RT-qPCR对7组随机挑选的miRNA-靶基因进行验证,结果与测序数据一致,说明测序结果可靠。 结论 miRNAs及其靶基因在不同氮形态处理后对杨树根尖氮响应起着重要的调控作用,从而使硝态氮处理下的灰杨根长比铵态氮处理的几乎长1倍。 Abstract:Objective To screen the differentially expressed miRNAs from root tips of poplar through small RNAs high-throughput sequencing technology, to study the regulation mechanism of miRNAs and their target genes related to nitrate or ammonium responses and try to identify and explain the morphological characteristics of poplar root tips growth and development. Method Using root tips of poplar as the research object, the high-throughput sequencing technology for small RNA library construction was applied to reveal the genetic differences of nitrogen forms in miRNA level, and to identify the differentially expressed miRNAs. Meanwhile, degradation sequencing technology was used to identify the miRNA target genes, and the functional annotation, clustering and metabolic pathway enrichment of the differentially expressed target genes were analyzed. Through conjoint analysis, the regulation network of miRNA-target pairs in the root tips of poplar was clarified under different nitrogen forms treatments. Result 523 known and 42 novel miRNAs were identified from miRNA libraries. A total of 96 miRNAs with significantly different expression were identified. Under ammonium treatment, 44 miRNAs were upregulated and 52 downregulated. Among them, the upregulation of vvi-MIR399d-p3_1ss13GA and the downregulation of novel miRNA PC-5p-35885_222 were the most significant. The KEGG pathway analysis of differentially expressed target genes of miRNAs in the libraries showed that some significantly different expression target genes were involved in nitrogen response pathway. Real time fluorescence quantitative PCR confirmed 7 randomly selected miRNA-target pairs, and the results were consistent with the sequencing data. Conclusion miRNAs and their target genes may play important roles in nitrogen response of poplar root tips after treatments with different forms of nitrogen, thus the root length under nitrate-nitrogen treatment is almost twice that under ammonium-nitrogen treatment. -
Key words:
- nitrate-nitrogen
- / ammonium-nitrogen
- / miRNA
- / mRNA
- / poplar
- / root
-
图 1 不同氮形态处理灰杨根系的表型
Figure 1. Morphological parameters of P. × canescens roots with different nitrogen forms
图 2 灰杨根尖已知和新miRNAs长度分布
Figure 2. Lengths of known and novel miRNAs in root tips of P. × canescens.
图 3 不同氮形态处理条件下灰杨根尖差异表达的miRNAs
Figure 3. Significantly differentially expressed miRNAs in root tips of P. × canescens under different nitrogen forms treatments
图 4 KEGG通路分析鉴定差异表达靶基因
Figure 4. KEGG pathway analysis of significantly differentially expressed target genes
图 5 不同氮形态处理下miRNAs-靶基因对差异表达分析
Figure 5. Validation of significantly differentially expressed miRNAs and their targets under different nitrogen forms treatments by sRNA-seq and RT-qPCR
表 1 RT-qPCR引物序列
Table 1. Primers used for RT-qPCR
miRNAs/靶基因ID
miRNAs/targets ID引物序列5′-3′
Primer sequence 5′-3′PC-5p-35885_222 TCTACTGAAAGTGAAAGCTAT ptc-MIR166e-p5 GGATTGTCGTCTGGTTCGATG ptc-MIR166m-p5_1ss9TC GAATGTCGCCTGACTCGAGAC ath-miR166e-5p_2ss4AC16AT GGACTGTTGTCTGGCTCGAGG mtr-miR166e-5p_2ss4AC10GT GGACTGTTGTCTGGCTCGAGG ptc-MIR1450-p3_2ss19AG21TC CGGTCAGGTTACACAAGCGAC ptc-miR6478_R+2_2ss5CT21GA CCGATCTTAGCTCAGTTGGTAGA ptc-MIR2111a-p3 GTCCTTGGGTTGCAGATTACC ptc-MIR2111b-p3 GTCCTTGGGTTGCAGATTACC vvi-MIR399d-p3_1ss13GA TCTGCCAAAGGAAATTTGCTC ptc-miR396g-5p TTCCACGGCTTTCTTGAACTT ath-miR858a_L-2R+1 TCGTTGTCTGTTCGACCTTG ptc-miR476b_2ss8TC17CT TAGTAATCCTTCTTTGTAAAA ptc-miR395b CTGAAGTGTTTGGGGGAACTC PC-3p-42422_177 TTTATGCGTTTTTGCCCCTCG ptc-miR399b TGCCAAAGGAGATTTGCCCGG URT primer 由Mir-X miRNA First-Strand Synthesis和SYBR RT-qPCR试剂盒提供 U6 F′/ R′ 均由Mir-X miRNA First-Strand Synthesis和SYBR RT-qPCR试剂盒提供 protein binding F′ TGAGATCAGCGAGGGACAAA protein binding R′ AAGGTTTGAGTGAGCAAGGC GRF9 F′ TCCCTCCCTCTCTCTGTCAA GRF9 R′ CATGATCACCAGCACCAATC GRF1 F′ TGCTGTTTCTTGGCCTGAAC GRF1 R′ CAATACCACCACCTCCACCT APS2 F′ CCAGAACCTGTCCCAGCTTA APS2 R′ GCTCCCAACACACATGCTAG phosphate 2 F′ AGGAAGAGCTGAGGGAGAGA phosphate 2 R′ CCTGTCGAACCACCCTTTTG MYB4 F′ CGCGTGTGGTGATTGTAGAG MYB4 R′ GTTGCCTCTCTTGATGTCCG ATP binding F′ GCGGGTGAATGAGGCTTTAG ATP binding R′ GCCTCTGAAACCTTGCCTTC 5.8s rRNA F′ CTCGGCTCTCGCATCGATGA 5.8s rRNA R′ AGACGTGCCCTCGACCAAGA Actin F′ CCCATTGAGCACGGTATTGT Actin R′ TACGACCACTGGCATACAGG  下载: 导出CSV
下载: 导出CSV
表 2 小RNA文库测序数据统计
Table 2. Distribution of small RNAs in different categories
类别
category硝态氮
NO3−铵态氮
NH4+Total Unique Total Unique raw reads 11 320 294 2 909 511 12 316 643 3 519 988 valid reads 5 752 458 1 996 697 7 800 546 2 608 788 Rfam 668 833 14 694 817 912 16 666 mRNA 169 342 2 779 343 819 5 169 other Rfam RNA 28 770 1 020 40 909 1 157 注:Rfam代表除miRNAs外许多常见的非编码RNA家族的集合。
Note: Rfam: collection of many common non-coding RNA families except miRNAs.
下载: 导出CSV
表 3 基于高通量测序技术鉴定灰杨根尖已知和新的miRNAs数量
Table 3. The number of identified known and novel miRNAs in root tips of P. × canescens based on high-throughput sequencing
样本
Samples已知/新
miRNAs known/
novelmiRNAsmiRNA前体
miRNA PremiRNA成熟体
miRNA mature硝态氮 NO3− 已知 Known 460 469 新的 Novel 37 39 铵态氮 NH4+ 已知 Known 502 514 新的 Novel 68 69 合计 Total 已知 Known 523 592 新的 Novel 42 42
下载: 导出CSV
表 4 29个显著差异表达miRNAs
Table 4. twenty-nine significantly differentially expressed miRNAs
miRNAs ID
miRNAs IDmiRNAs成熟体序列
sequence of miRNAs上调/下调
Up/downlog2(FC) P-value 是否RT-qPCR验证
RT-qPCR verifiedPC-5p-35885_222 TCTACTGAAAGTGAAAGCTAT down −4.50 0.003 21 是 ptc-MIR166e-p5 GGATTGTCGTCTGGTTCGATG down −3.15 0.007 14 是 ptc-MIR166m-p5_1ss9TC GAATGTCGCCTGACTCGAGAC down −2.32 0.008 37 是 ath-miR166e-5p_2ss4AC16AT GGACTGTTGTCTGGCTCGAGG down −2.30 0.003 18 是 mtr-miR166e-5p_2ss4AC10GT GGACTGTTGTCTGGCTCGAGG down −2.30 0.003 18 是 ptc-MIR1450-p3_2ss19AG21TC CGGTCAGGTTACACAAGCGAC down −2.27 0.006 95 是 mtr-miR171g_R+1 CGAGCCGAATCAATATCACTCC down −2.25 0.008 97 否 ptc-MIR169ae-p3_1ss20GA TGGGCAAGCACCTTGGCTAAC down −1.55 0.009 87 否 mtr-miR166c_2ss1TG20TC GCGGACCAGGCTTCATTCCCC down −1.30 0.003 29 否 ptc-MIR6438b-p5_1ss11AG TCGACTGAAAGTGAAAGCTAT down −0.96 0.003 68 否 PC-5p-166_26156 TAGCCAAGAGCGACTTGCCCA down −0.76 0.007 70 否 ptc-MIR396d-p3 CTCAAGAAAGCCGTGGGAGA down −0.54 0.002 63 否 ptc-miR166n_L+2R-2 TCTCGGACCAGGCTTCATTCC down −0.53 0.006 92 否 ptc-miR1447_1ss21TC CAGAATTGCAGTGCCTTGATC up 0.43 0.004 66 否 ptc-miR476b_2ss8TC17CT TAGTAATCCTTCTTTGTAAAA up 0.60 0.002 85 否 ptc-MIR1447-p5_1ss6GT AATCATGGCACTGCAATTCTA up 0.68 0.005 60 否 ptc-miR319e_R+1 TTGGACTGAAGGGAGCTCCTT up 1.00 0.001 08 否 vvi-miR172d_L-2R+1 AGAATCTTGATGATGCTGCATT up 1.05 0.000 50 否 ptc-miR530a_R+1 TGCATTTGCACCTGCACCTTA up 1.65 0.005 57 否 ptc-miR2111a TAATCTGCATCCTGAGGTTTG up 1.67 0.006 39 否 PC-3p-42422_177 TTTATGCGTTTTTGCCCCTCG up 1.95 0.003 47 否 ptc-miR6478_R+2_2ss5CT21GA CCGATCTTAGCTCAGTTGGTAGA up 2.11 0.008 94 是 ptc-MIR2111a-p3 GTCCTTGGGTTGCAGATTACC up 2.58 0.001 48 是 ptc-MIR2111b-p3 GTCCTTGGGTTGCAGATTACC up 2.58 0.001 48 是 vvi-MIR399d-p3_1ss13GA TCTGCCAAAGGAAATTTGCTC up 4.33 0.000 15 是 gma-miR166m_L+1_1ss5AC GCGGCCCAGGCTTCATTCCCC down −inf 0.003 18 否 PC-3p-82747_59 CCGATCATTATTGGTGACCTT down −inf 0.004 57 否 ptc-miR171k_1ss12CT GGATTGAGCCGTGCCAATATC down −inf 0.001 50 否 ptc-MIR319b-p5_1ss4TC TAGCTACCGACTCATTCATCCA down −inf 0.004 30 否
下载: 导出CSV
表 5 灰杨根尖氮代谢以及生长发育相关差异表达靶基因
Table 5. Significantly differentially expressed target genes related to nitrogen metabolism and growth and development of P. × canescens root tips
miRNAs ID
miRNAs ID靶基因ID
Targets ID功能注释
Functional annotationptc-MIR166i-p5 Potri.015G017500.4 glutamate synthase (NADH) ptc-MIR6462a-p5_1ss14TC Potri.005G079200.1 aminotransferase class Ⅰ and Ⅱ family protein gma-miR6300_1ss5TG Potri.001G300900.1 embryo sac development arrest 9 gma-miR6300_R+1_1ss5TG Potri.001G300900.1 embryo sac development arrest 9 gma-miR6300_1ss5TG Potri.001G300900.2 embryo sac development arrest 9 gma-miR6300_R+1_1ss5TG Potri.001G300900.2 embryo sac development arrest 9 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.1 nitrate transmembrane transporter,NRT1.1 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.2 nitrate transmembrane transporter,NRT1.1 gra-MIR8723b-p3_2ss6TC21TC Potri.003G111500.3 nitrate transmembrane transporter,NRT1.1 mtr-miR164a_1ss21AG Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) ptc-miR164a Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) vvi-miR164a_R+1 Potri.017G086200.1 NAC domain-containing protein 77 (NAC077) ptc-MIR7813-p5_2ss13AG20CT Potri.017G016700.1 NAC domain-containing protein 73 (NAC073) cca-miR164 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.007G065400.2 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.007G065400.1 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.3 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.2 NAC domain-containing protein 1 (NAC1) cca-miR164 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mdm-miR164a_1ss17AT Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mdm-miR164b_R+3 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) mtr-miR164a_1ss21AG Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) ptc-miR164a Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) ptc-miR164f Potri.005G098200.1 NAC domain-containing protein 1 (NAC1) vvi-miR164a_R+1 Potri.005G098200.1 NAC domain-containing protein 1 (NAC1)
下载: 导出CSV
表 6 不同氮形态处理条件下灰杨根尖差异表达miRNA-靶基因
Table 6. Significantly differentially expressed miRNAs-target pairs in root tips of P. × canescens under different nitrogen forms treatments
miRNAs ID
miRNAs ID上调/下调
Up/down靶基因ID
Targets IDlog2(FC)
log2(FC)靶基因注释信息
Target Functional annotationPC-3p-42422_177 up Potri.003G152300.2 −3.177 77 NA PC-3p-42422_177 up Potri.003G152000.1 −3.177 77 NA PC-3p-42422_177 up Potri.003G152300.1 −3.177 77 NA ptc-miR396g-5p down Potri.014G012800.1 2.346 49 growth-regulating factor 1 (GRF1) ptc-miR396g-5p down Potri.013G077500.1 inf growth-regulating factor 5 (GRF5) ptc-miR396g-5p down Potri.001G132600.1 1.613 36 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.014G071800.1 2.436 61 growth-regulating factor 9 (GRF9) ptc-miR396g-5p down Potri.003G100800.4 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.3 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.1 1.00154 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.003G100800.2 1.001 54 growth-regulating factor 2 (GRF2) ptc-miR396g-5p down Potri.007G007100.1 1.463 15 growth-regulating factor 1 (GRF1) ptc-miR395b up Potri.008G159000.3 −1.159 23 ATP sulfurylase 1 ptc-miR395b up Potri.010G081200.2 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.010G081200.1 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.010G081200.3 −1.727 46 Pseudouridine synthase (ATPS) ptc-miR395b up Potri.008G159000.2 −1.159 23 ATP sulfurylase 1 ptc-miR395b up Potri.008G159000.1 −1.159 23 ATP sulfurylase 1 ptc-miR399d up Potri.011G052600.1 −1.157 53 phosphate 2 ptc-miR399b up Potri.011G052600.1 −1.157 53 phosphate 2 ath-miR858a_L-2R+1 down Potri.006G221800.1 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ath-miR858a_L-2R+1 down Potri.006G221800.2 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ath-miR858a_L-2R+1 down Potri.006G221800.3 2.165 71 Duplicated homeodomain-like superfamily protein (myb4) ptc-miR476b_2ss8TC17CT up Potri.006G242500.1 −1.285 67 ATP binding; nucleic acid binding; helicases
下载: 导出CSV
-
[1] 国家林业局森林资源管理司, 全国森林资源统计-第七次全国森林资源清查[R]. 北京: 国家林业局, 2010, 37(2): 1-7. [2] Castro-Rodríguez V, García-Gutiérrez A, Canales J, et al. Poplar trees for phytoremediation of high levels of nitrate and applications in bioenergy[J]. Plant Biotechnology, 2016, 14(1): 299-312. doi: 10.1111/pbi.12384 [3] Rennenberg H, Wildhagen H, Ehlting B. Nitrogen nutrition of poplar trees[J]. Plant Biol (Stuttg), 2010, 12(2): 275-291. doi: 10.1111/j.1438-8677.2009.00309.x [4] Lu Y, Deng S, Li Z, et al. Competing endogenous RNA networks underlying anatomical and physiological characteristics of poplar wood in acclimation to low nitrogen availability[J]. Plant Cell Physiology, 2019, 60(11): 2478-2495. doi: 10.1093/pcp/pcz146 [5] Wei H, Yordanov Y S, Georgieva T, et al. Nitrogen deprivation promotes Populus root growth through global transcriptome reprogramming and activation of hierarchical genetic networks[J]. New Phytology, 2013, 200(2): 483-497. doi: 10.1111/nph.12375 [6] O'Brien J A, Vega A, Bouguyon E, et al. Nitrate transport, sensing, and responses in plants[J]. Molecular Plant, 2016, 9(6): 837-856. doi: 10.1016/j.molp.2016.05.004 [7] Zhou J, Fan Y, Chen H. Analyses of long non-coding RNA and mRNA profiles in the spinal cord of rats using RNA sequencing during the progression of neuropathic pain in an SNI model[J]. RNA Biology, 2017, 14(12): 1810-1826. doi: 10.1080/15476286.2017.1371400 [8] Liu C, Li C, Deng Z, et al. Long non-coding RNA BC168687 is involved in TRPV1-mediated diabetic neuropathic pain in rats[J]. Neuroscience, 2018, 374: 214-222. doi: 10.1016/j.neuroscience.2018.01.049 [9] Peng H, Zou L, Xie J, et al. lncRNA NONRATT021972 siRNA decreases diabetic neuropathic pain mediated by the P2X3 receptor in dorsal root ganglia[J]. Molecular Neurobiol, 2017, 54(1): 511-523. doi: 10.1007/s12035-015-9632-1 [10] Campa C, Diouf D, Ndoye I, et al. Differences in nitrogen metabolism of Faidherbia albida and other N-2-fixing tropical woody acacias reflect habitat water availability[J]. New Phytologist, 2000, 147: 571-578. doi: 10.1046/j.1469-8137.2000.00714.x [11] Barzegari M, Sepaskhah A R, Ahmadi S H. Irrigation and nitrogen managements affect nitrogen leaching and root yield of sugar beet[J]. Nutrient Cycling in Agroecosystems, 2017, 108(2): 211-230. doi: 10.1007/s10705-017-9853-y [12] Gunn K M, Baule W J, Frankenberger J R, et al. Modeled climate change impacts on subirrigated maize relative yield in northwest Ohio[J]. Agricultural Water Management, 2018, 206: 56-66. doi: 10.1016/j.agwat.2018.04.034 [13] Gaudin A C, McClymont S A, Holmes B M, et al. Novel temporal, fine-scale and growth variation phenotypes in roots of adult-stage maize (Zea mays L.) in response to low nitrogen stress[J]. Plant Cell Environment, 2011, 34(12): 2122-2137. doi: 10.1111/j.1365-3040.2011.02409.x [14] Zhang H, Rong H, Pilbeam D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana[J]. J Exp Bot, 2007, 58(9): 2329-2338. doi: 10.1093/jxb/erm114 [15] Li H, Hu B, Wang W, et al. Identification of microRNAs in rice root in response to nitrate and ammonium[J]. J Genet Genomics, 2016, 43(11): 651-661. doi: 10.1016/j.jgg.2015.12.002 [16] Zhou J, Lu Y, Shi W, et al. Physiological characteristics and RNA sequencing in two root zones with contrasting nitrate assimilation of Populus × canescens[J]. Tree Physiology, 2020, 40(10): 1392-1404. doi: 10.1093/treephys/tpaa071 [17] Tang G, Reinhart B J, Bartel D P, et al. A biochemical framework for RNA silencing in plants[J]. Genes & Development, 2003, 17(1): 49-63. [18] Ren Y, Sun F, Hou J, et al. Differential profiling analysis of miRNAs reveals a regulatory role in low N stress response of Populus[J]. Funct Integr Genomics, 2015, 15(1): 93-105. doi: 10.1007/s10142-014-0408-x [19] Lu S, Sun Y, Shi R, et al. Novel and mechanical stress–responsive microRNAs in Populus trichocarpa that are absent from Arabidopsis[J]. The Plant Cell Online, 2005, 17(8): 2186-2203. doi: 10.1105/tpc.105.033456 [20] Gifford M L, Dean A, Gutierrez R A, et al. Cell-specific nitrogen responses mediate developmental plasticity[J]. Proceedings of the National Academy of Sciences, 2008, 105(2): 803-808. doi: 10.1073/pnas.0709559105 [21] Vidal E A, Araus V, Lu C, et al. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences, 2010, 107(9): 4477-4482. doi: 10.1073/pnas.0909571107 [22] Gutierrez R A. Systems biology for enhanced plant nitrogen nutrition[J]. Science, 2012, 336(6089): 1673-1675. doi: 10.1126/science.1217620 [23] Li S, Tian Y, Wu K, et al. Modulating plant growth-metabolism coordination for sustainable agriculture[J]. Nature, 2018, 560(7720): 595-600. doi: 10.1038/s41586-018-0415-5 [24] Guo Q, Love J, Roche J, et al. A RootNav analysis of morphological changes in Brassica napus L. roots in response to different nitrogen forms[J]. Plant Growth Regulation, 2017, 83(1): 83-92. doi: 10.1007/s10725-017-0285-0 [25] Hu B, Wang W, Deng K, et al. MicroRNA399 is involved in multiple nutrient starvation responses in rice[J]. Front Plant Sci, 2015, 6: 188. [26] 许传俊, 李 玲. 泛素_26S蛋白酶体途径与植物的生长发育[J]. 西北植物学报, 2007, 27(3):635-643. [27] 朱美娇, 张海玲, 徐香玲, 等. 泛素_26S蛋白酶体途径调节非生物胁迫的研究进展[J]. 北方园艺, 2015(3):188-192. [28] Harberd N P, Belfield E, Yasumura Y. The angiosperm gibberellin-GID1-DELLA growth regulatory mechanism: how an "inhibitor of an inhibitor" enables flexible response to fluctuating environments[J]. Plant Cell, 2009, 21(5): 1328-1339. doi: 10.1105/tpc.109.066969 [29] Tamura W, Hidaka Y, Tabuchi M, et al. Reverse genetics approach to characterize a function of NADH-glutamate synthase1 in rice plants[J]. Amino Acids, 2010, 39(4): 1003-1012. doi: 10.1007/s00726-010-0531-5 [30] Maghiaoui A, Gojon A, Bach L. NRT1.1-centered nitrate signaling in plants[J]. J Exp Bot, 2020, 71(20): 6226-6237. doi: 10.1093/jxb/eraa361 [31] Zhao M, Ding H, Zhu J, et al. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis[J]. New Phytol, 2011, 190(4): 906-915. doi: 10.1111/j.1469-8137.2011.03647.x [32] He X, Qu B, Li W, et al. The nitrate-Inducible NAC transcription factor TaNAC2-5A controls nitrate response and increases wheat yield[J]. Plant Physiology, 2015, 169(3): 1991-2005. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5861
- HTML全文浏览量: 3013
- PDF下载量: 83
- 被引次数: 0
