


-
氮素是农林植物生长发育必不可少的大量元素之一,也是农林植物根系从土壤中吸收最多的矿质元素,对其生长发育具有重要意义[1-2]。但在土壤中无机氮含量往往较低[3],从而导致农林植物生产力低下,限制了其经济效益的发挥。大量含硝态氮或铵态氮无机氮肥的施用,成为维持农林土壤肥力的主要手段之一。但盲目、过量施肥往往造成土壤养分失衡、氮素流失等诸多环境问题[4]。因此,深入挖掘农林植物响应氮素的关键基因,解析其分子调控机制,提高农林植物对氮素吸收同化能力,既可以实现农林生态系统氮素高效利用的经济效益,也能实现减少土壤氮肥施加,保护生态环境的环保效益。
杨树(Populus L.)是我国主要的速生丰产用材林树种之一[5 ]。由于杨树生长迅速、对养分消耗大,对氮素有着很强的需求[3]。已有研究表明,不同氮形态处理,能够导致小黑杨(P. simonii × P. nigra)根系形态发生改变,从而影响其对氮素的吸收同化能力[6]。作者前期研究表明,硝态氮处理,能够改变灰杨(Populus × canescens)根尖形态,并影响根尖不同区段对硝态氮的吸收速率[7]。上述研究均表明,杨树根尖能够通过响应不同氮形态,改变其自身形态特征,从而影响根尖对不同氮形态的吸收同化能力[6-8]。
近年来,随着高通量测序技术的快速发展,转录组测序成为研究基因差异表达的重要手段。利用高通量测序技术对杨树根尖响应氮素的研究也取得了显著成果,但是研究大多集中在氮素亏缺方面[9-10]。例如,通过转录组重测序以及生物信息学分析表明,低氮处理导致灰杨根系中特异表达的 PtaNAC1(NAC-domain protein)上调表达,且增加了杨树根系生物量,并显著改变了PtaNAC1下游基因的表达,从而影响杨树根系形态结构[9]。Dash等的研究也表明,低氮处理条件下,在灰杨根系中,PtaHWS(Hawaiian Skirt),PtaNAC1和PtaRAP2.11(subfamilies of ERF/AP2 TF family)显著差异表达,并影响杨树根系形态建成[10]。上述研究均通过转录组学和生物信息学方法,对杨树根系响应低氮胁迫的过程进行了研究。而不同氮形态处理条件下,杨树根尖存在怎样的差异表达模式尚不清楚,值得深入研究。
本研究以灰杨根尖为试材,利用Illumina测序平台,筛选不同氮形态处理条件下,杨树根尖差异表达基因,并结合生物信息学分析,构建杨树根尖对不同氮形态响应过程的互作调控网络,挖掘关键基因。最终,阐明杨树根尖响应硝态氮或铵态氮,从而影响根尖生长发育过程的分子调控机制。研究成果为后续开发高氮素吸收利用效率的杨树新种质提供了科学依据。
-
将生根的灰杨组培苗(4 周)转移至水培,隔天换水浇灌改良的LA(Long Ashton)营养液(0.5 mmol·L−1NH4NO3、0.5 mmol·L−1KCl、0.9 mmol·L−1 CaCl2、0.3 mmol·L−1 MgSO4、0.6 mmol·L−1 KH2PO4、42 µmol·L−1 K2HPO4、10 µmol·L−1 Fe-EDTA、2 µmol·L−1 MnSO4、10 µmol·L−1 H3BO3、7 µmol·L−1 Na2MoO4、0.05 µmol·L−1 CoSO4、0.2 µmol·L−1 ZnSO4和 0.2 µmol·L−1 CuSO4, pH 5.5)。2周后,选取具有相似高度的48株植株,分成2组。分别添加0.5 mmol·L−1硝态氮(NO3−)和0.5 mmol·L−1铵态氮(NH4 + )代替LA营养液中的NH4NO3,进行水培,培养时间为10 d[7]。对根系形态特征进行分析。同时,收获根尖0~4 cm样品,迅速置于液氮中,放于−80℃冰箱备用。将8株植物的样本等量混合作为1个重复,每种处理水平3个生物学重复。
-
试验苗进行不同氮形态处理10 d后,对杨树根系构型进行观察测量,包括测量主根长度,以及测量可见侧根距根尖的距离。每个处理水平测量8株植株,且每种处理水平3个生物学重复。进一步,对数据统计进行分析,利用Statgraphics软件,先对数据进行正态性检验,采用单因素方差分析(ANOVA),以P < 0.05作为统计意义上的显著水平。
-
mRNAs 测序文库构建和测序按照Illumina 公司提供的标准步骤执行。将上述收获的2种不同氮形态处理条件下的根尖(0~4 cm)样品,每个处理3个重复,分别利用RNA提取试剂盒提取总RNAs (TRK1001, 联川生物技术公司,杭州,中国)。将提取的总RNAs经过DNase I消化后,富集含有polyA尾的mRNA,反转录成cDNA。合成第二链cDNA后,用QIA quick PCR extraction 试剂盒进行纯化。经过回收纯化、PCR富集后,获得cDNA文库。将构建好的文库,利用杭州联川生物技术公司IlluminaHiSeqTM4000高通量测序技术,进行转录组测序分析。
-
将测得的RNAs序列,使用联川生物技术公司开发的软件进行分析。首先,将得到的原始序列进行去杂质处理,包括接头序列的读取片段、质量值Q ≤ 5的低质量序列读取片段和未知碱基数比例>10%的序列读取片段。经过筛选得到的序列读取片段使用SOAPaligner/soap2将测序结果比对到灰杨基因组数据库(
http://aspendb.uga.edu/index.php/databases/spta-717-genome )[11]。采用FPKM (Fragments Per Kilobase of exon per Million mapped)对基因的表达水平进行定量[12]。基于FPKM值,使用Ballgown package计算mRNAs的差异表达水平。利用mRNAs在硝态氮处理条件下的FPKM除以铵态氮处理条件下FPKM来计算基因的差异倍数(Fold change,FC)。差异表达mRNAs筛选阈值为log2(FC)≥1或 ≤ −1,且P < 0.05。 -
以上述提取的不同氮形态处理杨树根尖样品为模板,使用Takara公司PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒反转录合成cDNA第一链,作为荧光定量PCR反应的模板。从转录组数据中选取10个差异表达的基因,根据灰杨数据库,利用Primer Premier 3.0软件设计引物,并由上海生工生物工程有限公司合成相关引物。Actin为RT-qPCR的内参基因。序列见表1。使用SYBR Premix Ex TaqTM荧光定量试剂盒进行荧光定量检测。每个样品进行3次重复。将qPCR得到的Ct值进行归一化,计算差异表达基因的相对表达量[7]。
表 1 RT-qPCR引物序列
Table 1. Primers used for RT-qPCR
基因 ID gene ID 正向引物 Forward primer 反向引物 Reverse primer Potri.007G102400 ATTCCTGACATGGGCACTGA AGAAGTGATCCAGTCCCTGC Potri.001G010700 TACCATGGGAAAGCTGCTGA AGGCCTTGGAGCAGATCTTT Potri.009G045100 ACAATACCTGCCTCTCCACC GTGGTGCCGGCTTTTCTAAA Potri.003G152300 ATATCCACGAACCGACCTCC TATCGTTTTGACGCTCCTGC Potri.004G116500 CCCGTTATTGCAGCCTCATC AATGAATCCCAGAGCCAGCT Potri.001G391900 GCCTGGCAGTTATACAAGGC AAGTGCTCCTCTCCGAAACA Potri.006G123200 TGGTGACCAAGGCCATATGT AGAATCGTGTGAACCCGGAT Potri.019G131500 AATGGAGAATGTGGGGAGGG TTCCCCTAGCAGTTCCAAGG Potri.005G232600 GTCAATCTACGTGCCGGAAC CCATTGCTGCCATGTCAAGT Potri.007G045300 GGAGTGGACAAGTAGCCCTT TATCACCAGCAGAGCAACCA Potri.005G172400 CTGCTGACAACTGGATCGAA TTTTAACCAGACCGCAAACC Potri.002G113600 GGATGAGTTTCATCGGGCTA CCCGCTTGTAGCAGGAGTAG Potri.008G009500 AGGTTGTAGGAAGGGCTGGT GCCAACGTCTTCCCAACTAA Potri.014G022800 AATTTGGGAGACACCTGTGC CAGCCATTCTGCAGTTCGTA Potri.005G213100 TCCTCTCTGGAGTGCCAAGT ATATCCCTTGCCTCACATGC Potri.001G327000 CGACCAAGATGGGAAGAAAA TTGCAGTGGAGGTCTGTGAG Actin CCCATTGAGCACGGTATTGT TACGACCACTGGCATACAGG -
通过Blast2GO软件,对所有差异基因进行GO功能注释[13]。随后进行GO显著性富集分析,以上述得到目的GO条目(term)为单位,通过P值Bonferroni校正(P < 0.05)来定义差异表达基因中显著富集的GO term。同时,结合生物信息学KEGG软件,分析差异表达基因的生物学功能[14]。同样以corrected P < 0.05为标准,筛选在差异表达基因中显著富集的代谢途径。利用MapMan软件、Popgenie数据库(
https://popgenie.org/exnet )以及Cytoscape程序,构建杨树根尖响应不同氮形态相关差异表达基因的互作调控网络[15]。 -
在不同氮形态处理条件下,灰杨根系形态特征具有显著差别(图1)。硝态氮处理下的主根长度几乎是铵态氮处理条件下的一倍,这说明不同氮形态处理影响杨树根系生长发育过程。同时,在两种处理水平下,能够发挥吸收营养和水分作用的可见侧根发生的位置相对一致。因此选取侧根形成前的0~4 cm根尖部分进行后续研究。

图 1 不同氮形态处理后,灰杨根系表型
Figure 1. Morphological parameters of P. × canescens roots with different nitrogen forms
-
基于上述形态特征分析的结果,以不同氮形态处理杨树根尖为材料,进行高通量转录组测序分析。测序结果显示,两个处理水平的测序分别获得57 001 409和57 517 359条有效序列(表2),占各自文库的比例均超过了99.20%(表2)。两个处理水平的测序数据,能够比对到灰杨数据库中的碱基数比例分别为81.59%和82.60%,说明测序所得序列与灰杨数据库比对性较好,可以用于后续分析。
表 2 转录组文库测序数据统计
Table 2. Distribution of RNA-seq in different categories
样品
Sample原始数据
Raw data有效数据
Valid data有效比例
Valid ratio(reads) /%比对碱基数
Mapped readsQ20% Q30% GC含量
GC content/%NO3− 57407539 57001409 99.29 46521558(81.59%) 99.51 95.89 45 NH4+ 57945941 57517359 99.26 47517203(82.60%) 99.44 95.95 45 -
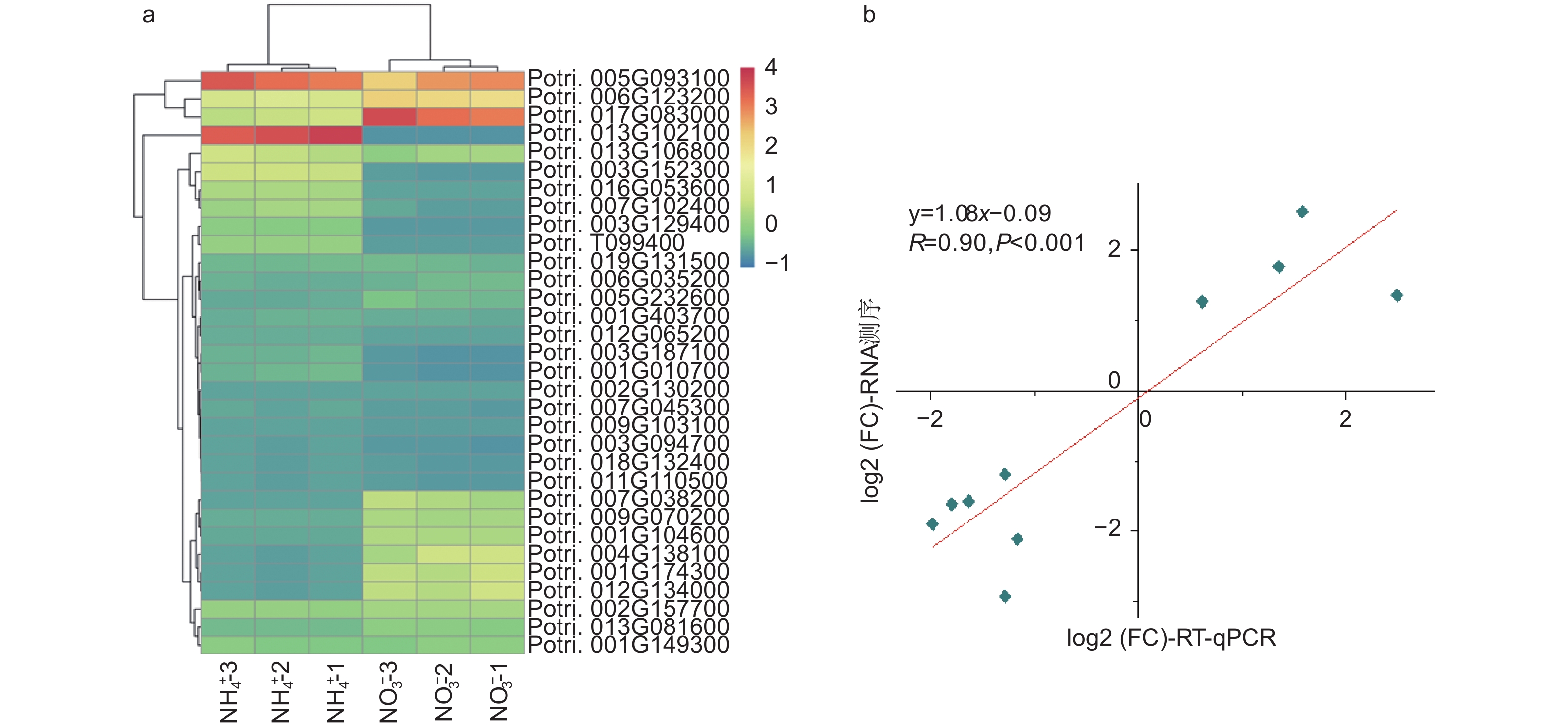
在不同氮形态处理条件下,以铵态氮处理水平为对照,在硝态氮处理条件下,共筛选出2207个差异表达基因(p < 0.05)。相比较于铵态氮处理,硝态氮处理有1414个基因上调表达,793个基因下调表达。有32个基因显著差异表达(p < 0.001),见图2a。其中,Potri.013G102100基因差异变化最显著,log2(FC)为−6.56,该基因注释为II型过氧化物酶(peroxiredoxin type 2);Potri.004G138100基因上调倍数最显著,log2(FC)为6.18,然而该基因尚未被注释,功能未知。同时,筛选出71个与植物生长发育相关的差异表达基因。其中,转录因子作为一大类调控基因,在杨树根尖通过响应不同氮形态而参与其生长发育过程中发挥着重要作用。例如,NFYA(nuclear transcription factor Y subunit alpha)、GRF(growth-regulating factor)和ARF(auxin response factor)类转录因子,它们均能够通过调控硝态氮转运体NRT(nitrate transporters)基因的表达响应不同氮形态,从而调节植物根系生长发育过程[16-17]。进一步,挑选了10个显著差异表达基因,利用RT-qPCR进一步证实了测序结果的可靠性(图2b)。
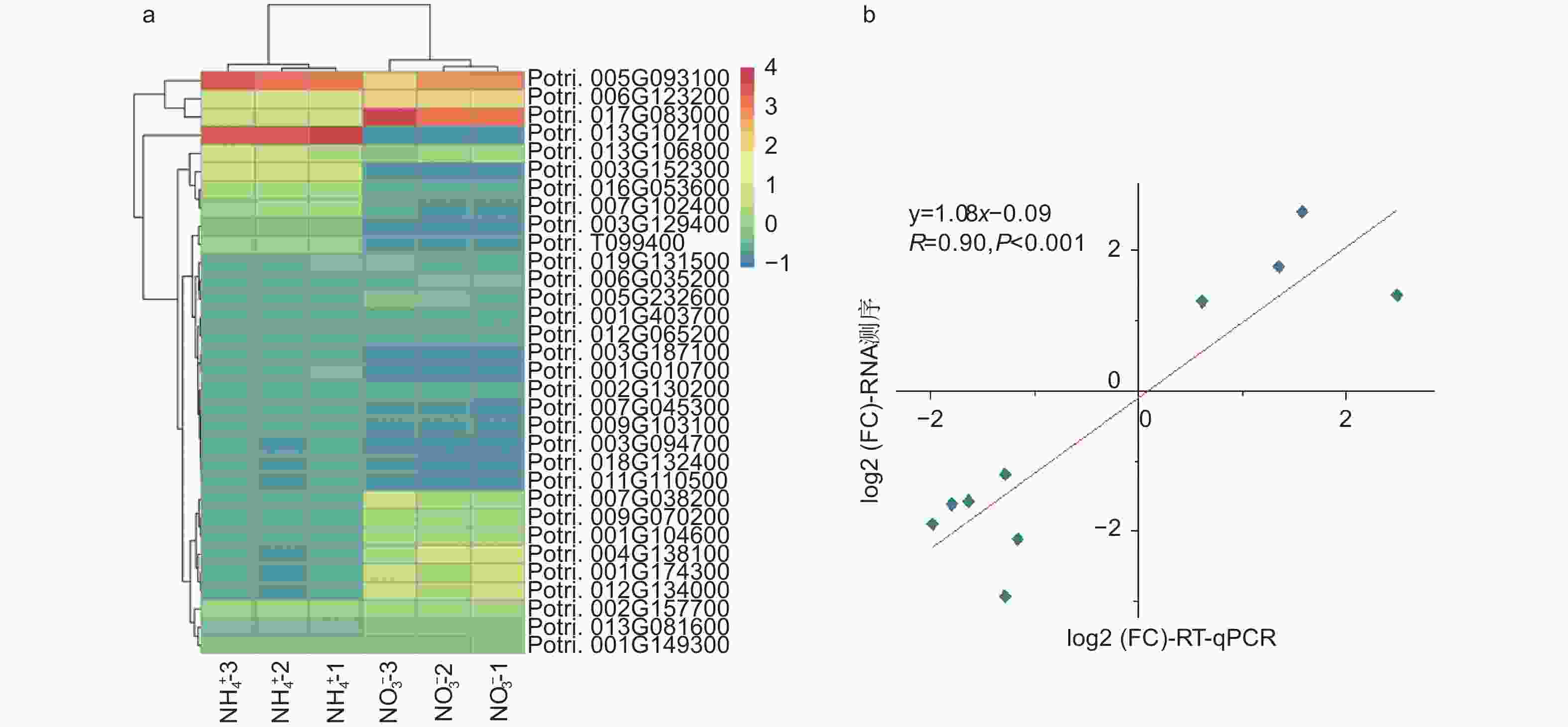
图 2 不同氮形态处理条件下,杨树根尖显著差异表达的基因
Figure 2. Significantly differentially expressed mRNAs in root tips of P. × canescens under different nitrogen form treatments
-
为进一步了解差异表达基因的功能,将差异表达基因映射GO数据库,根据序列同源性,将不同氮形态处理条件下,2207个差异表达基因分为3大类总共50个功能组,包括生物学过程(biological process,BP)、细胞组分(cellular component,CC)和分子功能(molecular function,MF)(图3)。结果显示,生物学过程主要集中在转录调控、刺激反应和代谢过程。在细胞组分类别中,差异基因被注释到最多的亚类依次为细胞核、质膜和细胞质部分。在分子功能中,注释为ATP结合和蛋白结合的差异基因最多,其次为转录因子活性和DNA结合。以上注释较多的过程所涉及到的差异基因,可能参与杨树根尖对不同氮形态的响应,从而影响根尖生长发育过程。进一步挖掘这些基因的功能,研究其响应不同氮形态的作用机制,将有助于为培育高氮素吸收利用效率的杨树新种质奠定基础。

图 3 不同氮形态处理条件下,灰杨根尖差异表达基因的GO功能聚类注释
Figure 3. Gene Ontology classification annotation of differentially expressed genes in root tips of P. × canescens under different nitrogen form treatments
-
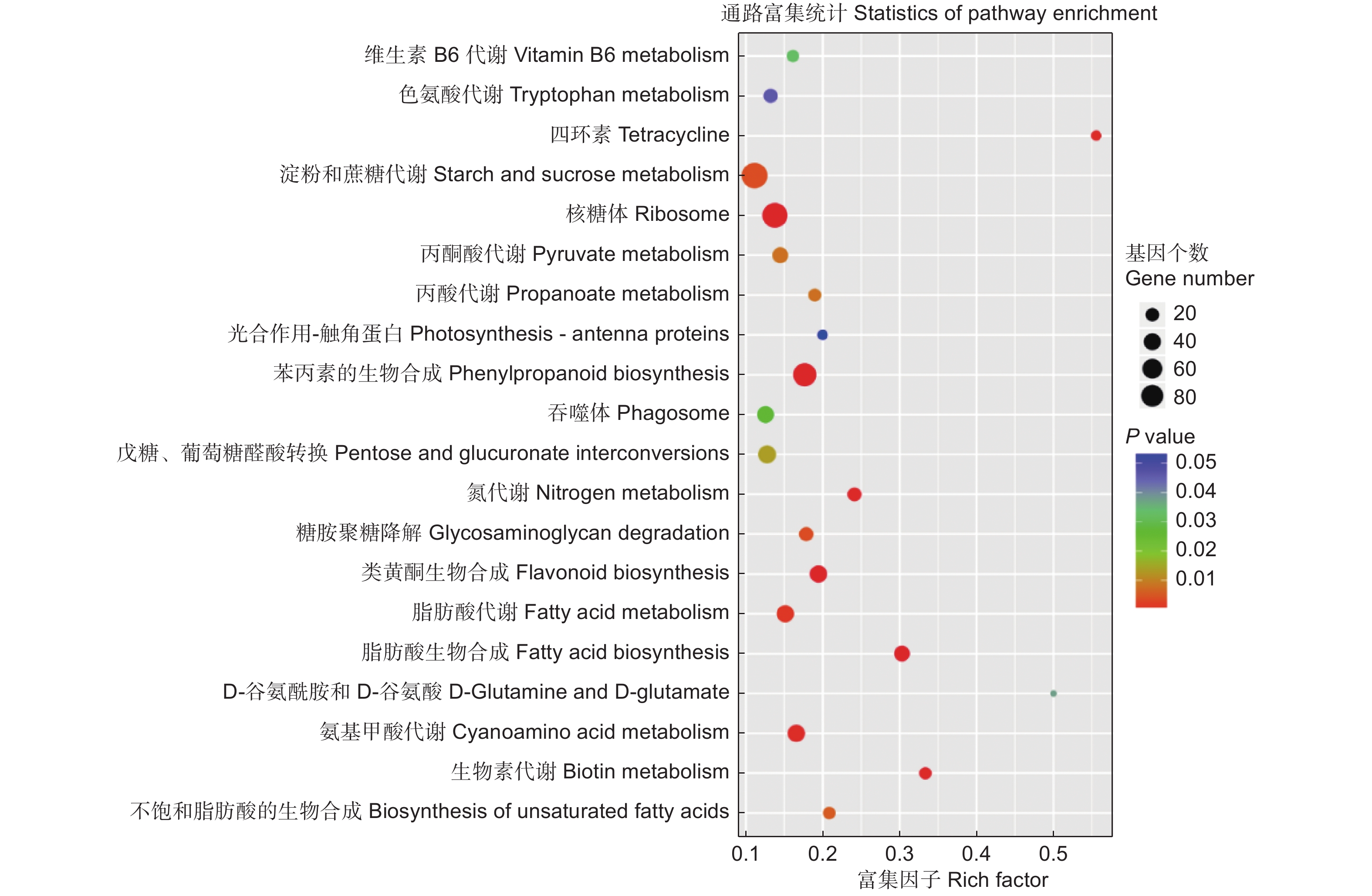
为进一步了解上述差异表达基因的功能,对识别出的差异表达基因进行KEGG通路分析。共获得1 192个差异表达基因,涉及到20条KEGG通路(图4)。其中,有9条与代谢相关,并且大多与氮代谢和氨基酸代谢相关,有4条与生物合成相关。KEGG分析从功能的角度锁定基因,直观的显示了杨树根尖响应不同氮形态差异基因的代谢过程,有助于更好地研究杨树根尖响应不同氮形态的分子机制。
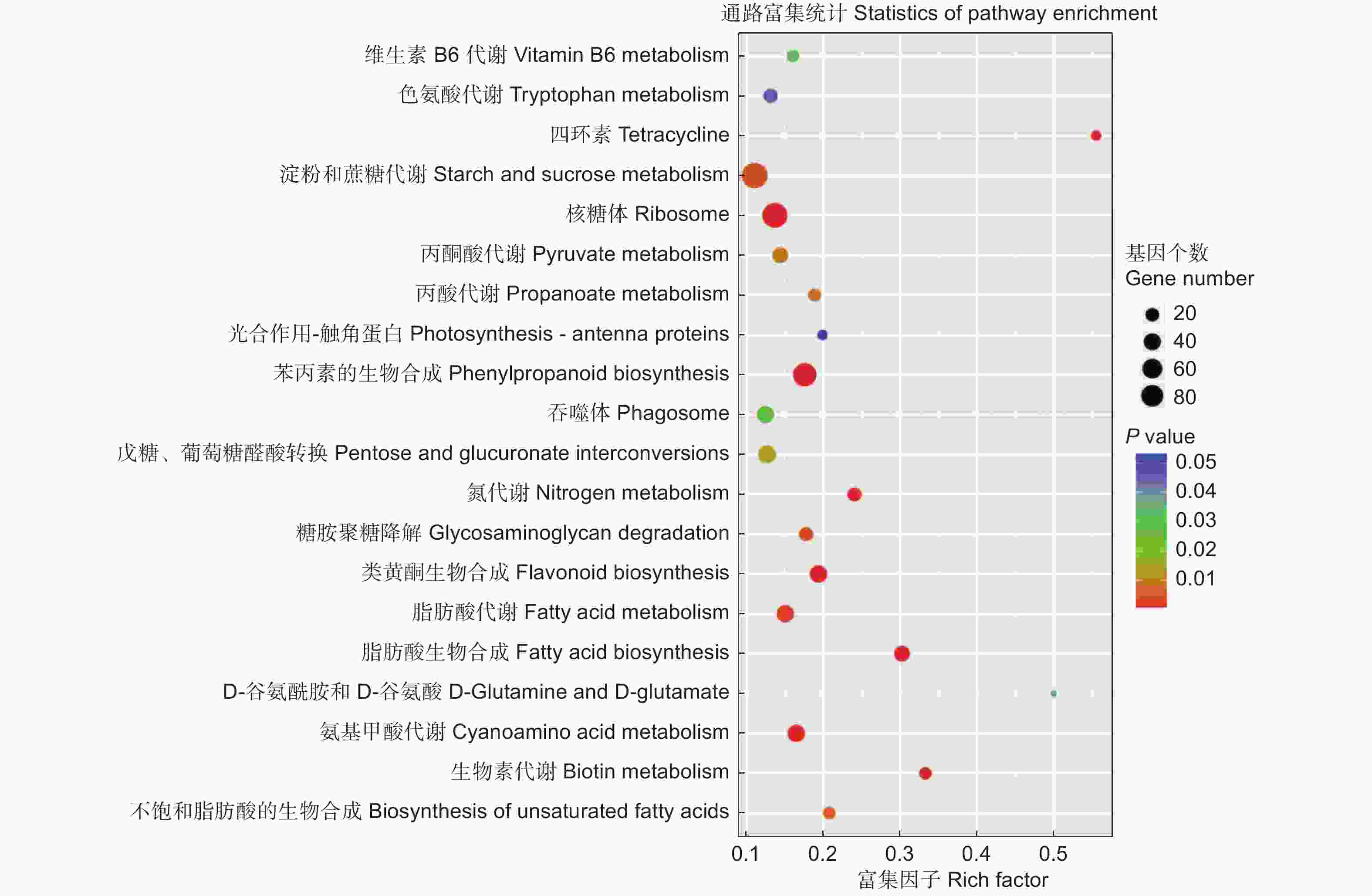
图 4 KEGG通路分析鉴定差异表达基因
Figure 4. KEGG pathway analysis of significantly differentially expressed target genes
-
利用MapMan软件,筛选出36个与氮代谢通路过程、各类氨基酸的生物合成以及代谢过程相关的差异表达基因(表3)。其中,有8个基因参与到氮代谢过程,26个基因参与到氨基酸代谢与合成过程,还有2个基因参与到硝态氮转运过程。已有研究表明,硝态氮处理能够改变NRTs的表达水平,进一步影响植物根尖生长发育过程[18-19]。
表 3 氮代谢相关差异表达基因
Table 3. Differentially expressed genes related to nitrogen metabolism
基因名字 Gene name 调节类型 Regulation 功能注释 Description log2(fc) p value Potri.005G172400 up nitrate reductase family protein 4.18 0.0049 Potri.002G088600 up nitrate reductase family protein 3.84 0.0037 Potri.004G140800 up Ferredoxin--nitrite reductase family protein 3.03 0.0117 Potri.008G200100 up glutamine synthetase family protein 1.78 0.0174 Potri.012G043900 up glutamine synthetase family protein 1.02 0.0153 Potri.015G111000 down glutamate dehydrogenase [NAD(P)+] −1.82 0.0194 Potri.012G113500 down glutamate dehydrogenase [NAD(P)+] −2.68 0.0036 Potri.002G146100 down nitroreductase family protein −1.47 0.0469 Potri.014G143300 up aspartate aminotransferase 1.11 0.0335 Potri.001G162800 down alanine aminotransferas family protein −1.34 0.0344 Potri.002G106200 up argininosuccinatelyase 1.03 0.0231 Potri.006G123200 up S-adenosylmethionine synthetase family protein 1.78 0.0000 Potri.014G114700 up S-adenosylmethionine synthetase 1 family protein 1.60 0.0011 Potri.004G190900 up methionine synthase 1.18 0.0031 Potri.005G213100 up selenocysteine methyltransferase family protein 1.43 0.0222 Potri.010G083600 down Homocysteine S-methyltransferase 3 family protein −1.57 0.0370 Potri.013G099500 up aspartate kinase family protein 1.38 0.0252 Potri.002G113600 down branched-chain-amino-acid transaminase −1.71 0.0218 Potri.014G022800 up D-3-phosphoglycerate dehydrogenase 1.38 0.0163 Potri.008G009500 up phosphoglycerate dehydrogenase 1.30 0.0033 Potri.T069600 up hypothetical protein 1.89 0.0311 Potri.019G038900 up hypothetical protein 1.41 0.0111 Potri.005G048200 up O-acetylserine(thiol)lyase family protein 1.13 0.0020 Potri.011G004700 up prephenate dehydratase family protein 1.57 0.0172 Potri.013G066500 up C2 domain-containing protein 1.02 0.0190 Potri.001G348600 down L-aspartate oxidase −1.07 0.0390 Potri.017G123300 up methionyl-tRNAformyltransferase 1.52 0.0405 Potri.003G187100 down methionine gamma-lyase −1.52 0.0001 Potri.001G005400 up protein lysine decarboxylase-like protein 1.43 0.0494 Potri.009G010800 down protein carboxy-lyase −1.25 0.0282 Potri.019G018500 down L-allo-threonine aldolase-related family protein −2.50 0.0386 Potri.001G374100 down homogentisate 1,2-dioxygenase −1.10 0.0203 Potri.002G057700 up enoyl-CoA hydratase/isomerase family protein 1.73 0.0103 Potri.001G327000 down NA −1.12 0.0044 Potri.014G179400 down nitrate transmembrane transporter 1.5 −1.69 0.0462 Potri.003G088800 down nitrate transmembrane transporter 1.5 −3.71 0.0094 -
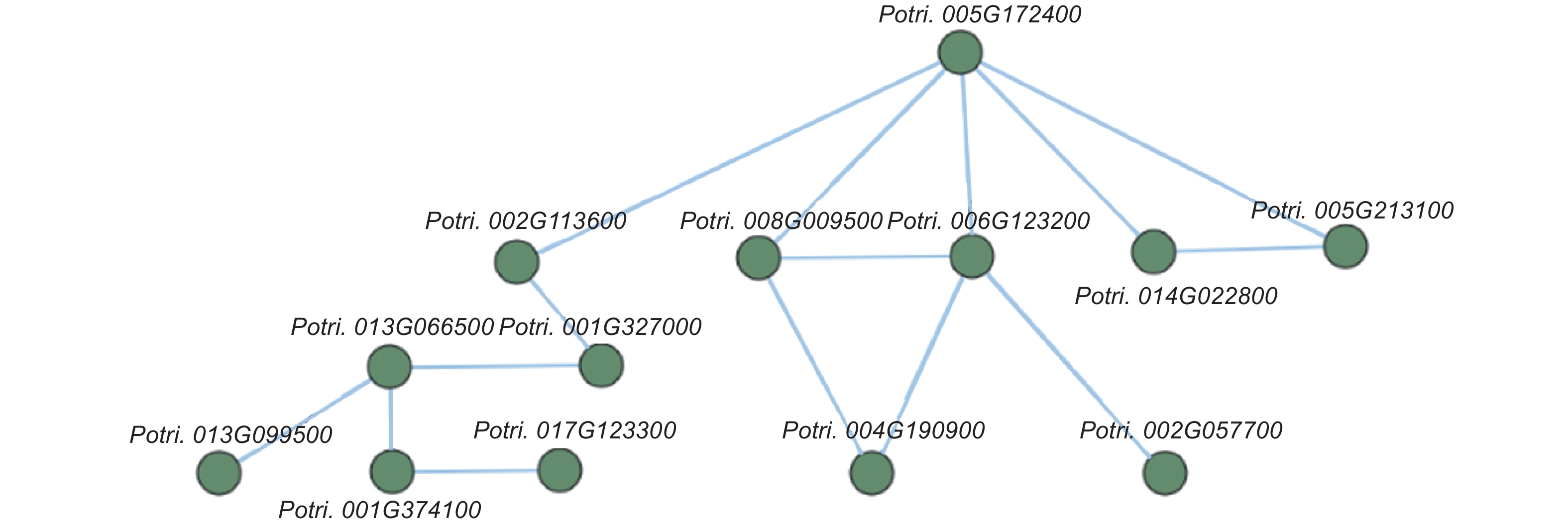
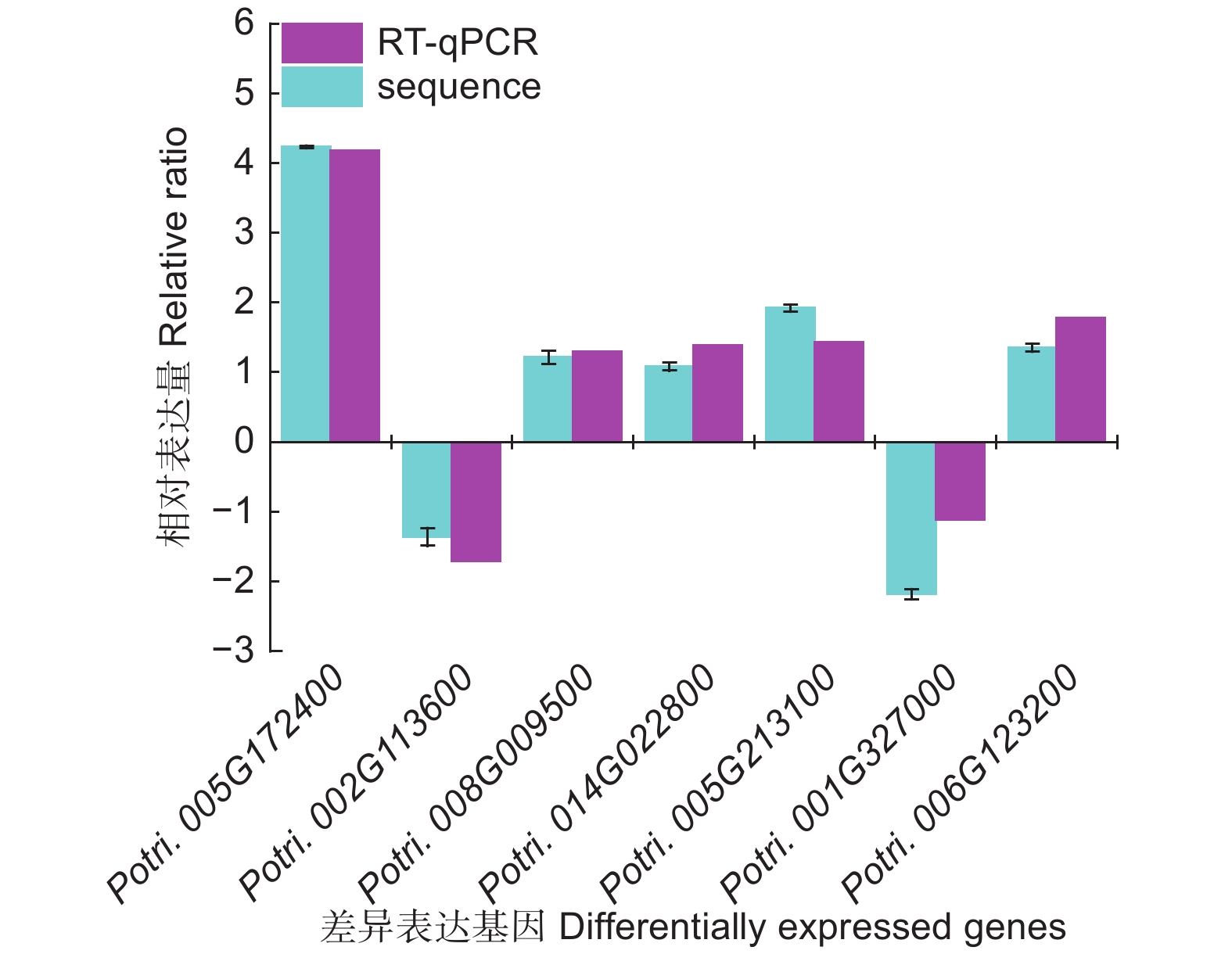
为了进一步研究这些基因的功能,利用Popgenie数据库以及Cytoscape程序,构建了灰杨根尖对不同氮形态响应过程的互作调控网络。在基因作用网络中,检测到一组相互作用的与氮代谢过程相关的基因簇,共包含13个基因(图5)。其中,硝酸还原酶(Potri.005G172400)基因作为硝态氮同化过程中的关键酶,占据了网络的核心位置。已有研究表明,硝酸还原酶作为植物氮素同化的关键酶之一,能够调控根系生长素水平,从而影响根尖生长发育过程[20-21]。进一步,挑选了调控网络中7个显著差异表达基因,选取不同氮形态处理10天的灰杨根尖材料进行RT-qPCR实验。结果显示:这7个基因在RT-qPCR的结果与转录组分析的趋势一致(图6)。
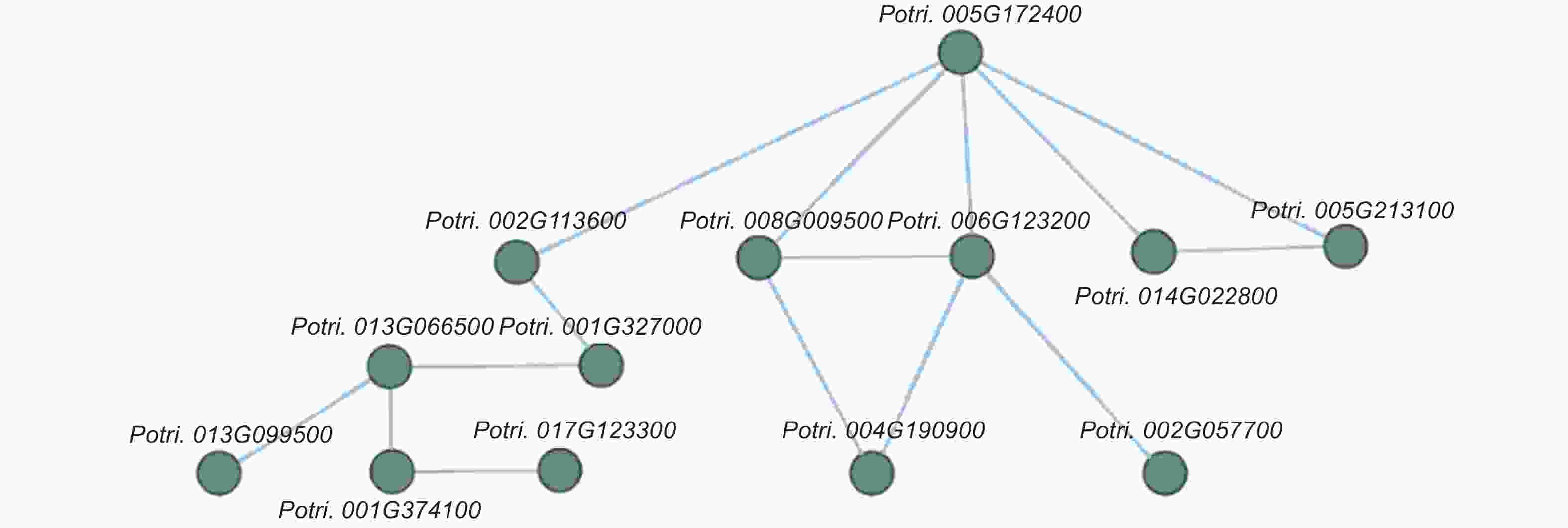
图 5 不同氮形态处理条件下,灰杨根尖差异表达基因的互作网络
Figure 5. The interaction network of differentially expressed genes in root tips of P. × canescens under different nitrogen forms treatments
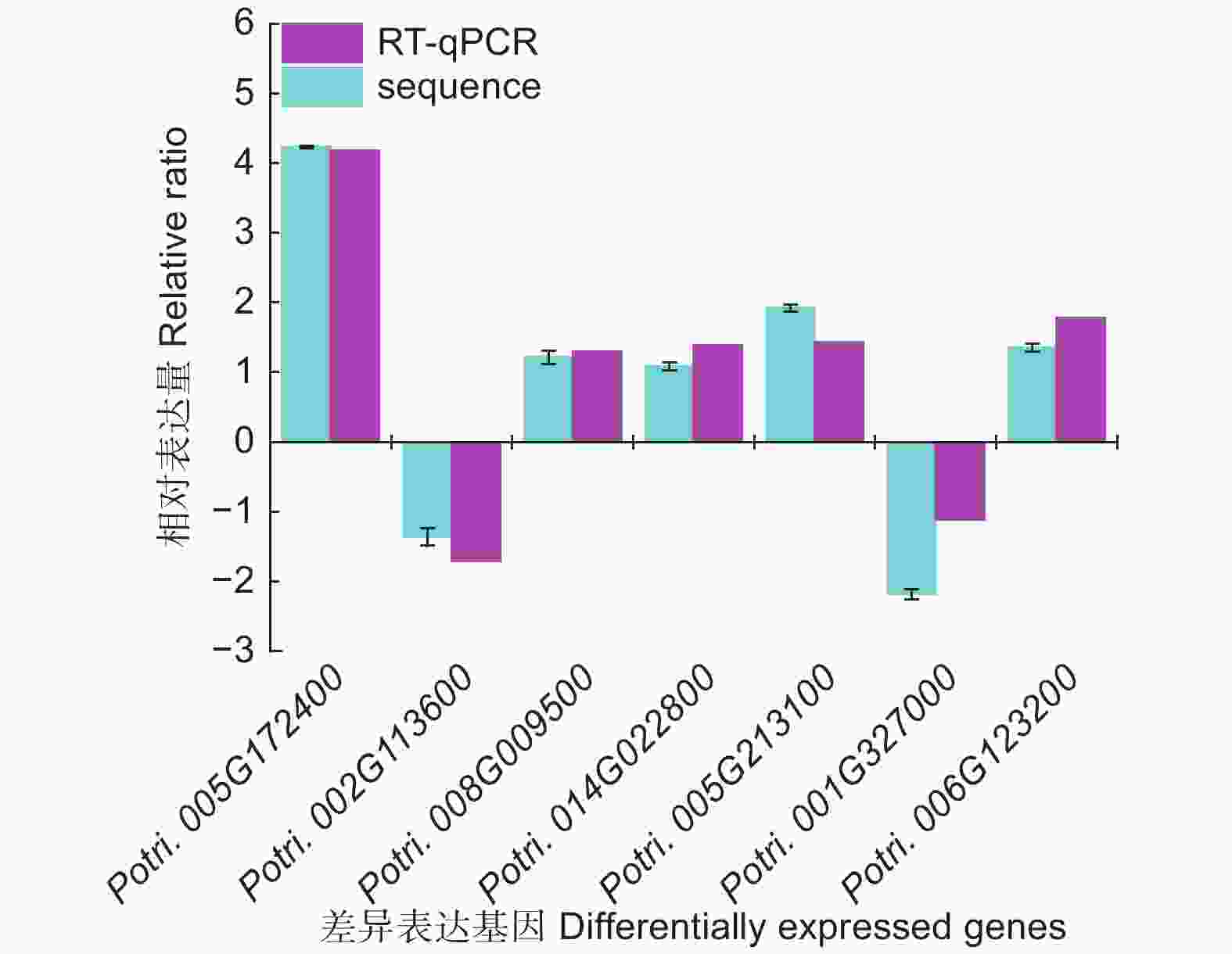
图 6 不同氮形态处理条件下,灰杨根尖差异基因表达分析
Figure 6. Validation of significantly differentially expressed genes under different nitrogen forms treatments by sRNA-seq and RT-qPCR
-
氮素是植物生长发育所必需的大量矿质营养元素之一,是蛋白质、核酸、叶绿素等重要物质的组成成分[22]。土壤中的氮素都需要在适宜的温度、水分和通气条件下,在土壤微生物和酶的作用下,将其水解或氧化为硝态氮或铵态氮,才能够被植物根系直接吸收利用[23-24]。而土壤中的不同氮形态可能导致杨树根尖氮代谢相关基因表达模式发生改变,从而影响植物根尖生长和发育过程[25]。本研究在不同氮形态处理10 d后,对杨树根尖进行观察发现,硝态氮处理下的主根长度几乎是铵态氮处理条件下的一倍。类似的现象在小黑杨根系中(P. simonii × P. nigra)也有报道[6]。为了进一步了解木本植物根尖对不同氮形态响应的过程,本研究利用转录组测序技术,对不同氮形态处理条件下,杨树根尖差异表达基因进行了分析。
在不同氮形态处理10 d后,经过高通量测序分析,获得了2207个差异表达基因。其中,Potri.013G102100基因编码一个II型过氧化物酶(peroxiredoxin type 2),其下调倍数最为显著,前人研究表明,II型过氧化物酶是植物中分布最广的一类非典型2-半胱氨酸过氧化物酶(2-Cysteine Peroxiredoxin),其在对抗氧化能力和共生固氮中发挥着重要作用[26]。另有研究表明,2-半胱氨酸过氧化物酶可以与氮同化过程中亚硝酸还原酶(nitrite reductase)和谷氨酸合成酶(glutamate synthase)相互作用,从而影响氮代谢过程[27]。Potri.004G138100基因上调倍数最显著,但该基因功能未知,推测其可能参与杨树根尖对不同氮形态的响应,从而影响杨树根尖生长发育过程。同时,在本实验中,部分差异表达转录因子通过响应不同氮形态,影响杨树根尖生长发育过程,这与在草本植物中的研究结果相似[6, 8, 28]。例如,在水稻(Oryza sativa L.)和小麦(Triticum aestivum)中,过表达NFYAs能够调控NRTs的表达[18, 29]。同时,增加了转基因小麦根尖对硝态氮的吸收,促进了侧根生长[18]。在本研究中,相比较于铵态氮处理,硝态氮处理导致灰杨根尖PcNFYA7(Potri.011G101000)下调表达,同时促进杨树主根伸长。在拟南芥(Arabidopsis thaliana)根中柱鞘细胞中,5mM 硝态氮可以促进ARF6/8的表达水平,促进了拟南芥侧根起始和随后萌发的过程[17, 30]。在本研究中,相比较于铵态氮处理,硝态氮处理导致灰杨根尖中PcARF8/16(Potri.004G078200和Potri.010G223200)上调表达,同时促进了主根伸长。上述研究表明,灰杨根尖部分差异表达的基因,能够通过响应不同氮形态而参与调控根尖生长发育过程。
对差异基因进行GO和KEGG通路分析。结果表明,一些差异表达基因参与氮代谢和氨基酸代谢途径。例如,氮代谢、色氨酸代谢、丙酮酸代谢和氨基甲酸代谢途径等。同时,利用MapMan筛选出36个氮代谢相关差异表达基因。其中,值得关注的是两个硝态氮转运体基因PcNRT1.5(Potri.014G179400和Potri.003G088800)。相比较于铵态氮处理,硝态氮处理导致灰杨根尖中PcNRT1.5显著下调表达。在拟南芥中,AtNRT1.1作为第一个被鉴定出的NRT在根中有转运和感应硝态氮两个作用[31]。同时,在小麦和水稻中,硝态氮处理均能改变NRT基因的表达水平,从而影响植物根尖生长发育过程[18-19]。该结果说明PcNRT1.5在杨树根尖响应不同氮形态过程中发挥着重要的调控作用。进一步,通过Popgenie数据库以及Cytoscape程序,构建了灰杨根尖响应不同氮形态的互作调控网络。在基因作用网络中,硝酸还原酶(Potri.005G172400)作为硝态氮同化过程中第一步的关键酶,在杨树根尖对不同氮形态响应过程中发挥着关键作用。这与在草本植物中的研究结果类似[20]。在拟南芥根中,硝酸还原酶作为氮素同化和根系构型重塑的关键酶之一,能够调控根系生长素水平,从而影响根尖生长发育过程[20]。同时,在水稻中,硝酸还原酶生成的一氧化氮,通过诱导水稻侧根形成和无机氮的吸收,提高水稻对氮的吸收能力[21]。而杨树作为多年生木本植物,其根系比浅根系草本植物发达得多,且构型更为复杂[9]。同时,木本植物往往遭受季节性、物候性土壤氮素反复变化,面临的土壤氮素环境复杂程度远远大于草本植物[4, 32-33]。因此,杨树根系中硝酸还原酶可能存在着更为复杂的氮适应性变化的作用机制,值得深入研究。同时,通过基因互作网络的分析,可以逐步了解参与互作网络的基因在响应不同氮形态过程中的作用,其相互作用关系也有待进一步确认。
-
本研究在不同氮形态处理10 d后,对杨树根尖进行观察发现,硝态氮处理下的主根长度几乎是铵态氮处理条件下的一倍。同时,通过转录组测序分析,筛选出2 207个差异表达基因。通过GO功能和KEGG通路分析,理清了差异表达基因富集的分子功能与代谢通路。利用MapMan筛选出36个氮代谢相关差异表达基因,并构建了这些基因的互作网络,得到了关键基因。综上所述,本研究为挖掘和寻找杨树根尖响应不同氮形态关键调控基因提供了参考,并阐述了这些关键基因可能通过响应不同氮形态,影响杨树根尖生长发育过程,这为进一步培育出高效吸收利用氮素的杨树新种质奠定了基础。
不同氮形态处理条件下杨树根尖差异表达基因的特征分析
Characterization of Differentially Expressed Genes in Root Tips of Poplar Under Different Nitrogen Forms
-
摘要:
目的 利用高通量转录组测序技术,在硝态氮或铵态氮处理条件下,对杨树根尖差异表达基因进行了筛选和研究,同时分析和描述了差异表达基因对杨树根尖生长发育的影响,为后续开发高氮素吸收利用效率的杨树新种质提供科学依据。 方法 以灰杨幼苗根尖为材料,用0.5 mmol·L−1硝态氮(NO3−)和0.5 mmol·L−1铵态氮(NH4+)对幼苗处理10 d,并对植株根尖进行转录组测序以及生物信息学分析。 结果 硝态氮处理下的主根长度几乎是铵态氮处理条件下的一倍。从两种不同氮形态处理杨树根尖转录组文库中,筛选到2207个差异表达基因。通过差异基因GO和KEGG功能聚类分析,分别获得50个GO功能聚类和20条KEGG通路。进一步利用MapMan分析,筛选出36个氮代谢通路过程、各类氨基酸的生物合成以及代谢过程相关的差异表达基因。对这些差异基因互作调控网络分析发现,硝酸还原酶(Potri.005G172400)基因通过响应不同氮形态,在影响杨树根尖生长发育过程中发挥了重要作用。 结论 本研究获得了不同氮形态处理条件下杨树根尖差异表达基因,并对差异表达基因功能进行分析,有助于理解杨树通过响应不同氮形态影响根尖生长发育过程的分子机制。 Abstract:Objective To understand the molecular regulation mechanism of Populus × canescens root tips in response to different nitrogen forms. Method High-throughput transcriptome sequencing technology was used to screen and analyze the differentially expressed genes in poplar root tips under nitrate or ammonium treatments. At the same time, the effects of differently expressed genes on the growth and development process of poplar root tips were analyzed and described, which provides a scientific basis for the subsequent development of new poplar germplasm with high nitrogen absorption and utilization. The root tips of P. × canescens were treated with 0.5 mmol·L−1nitrate (NO3−) and 0.5 mmol·L−1 ammonium (NH4 + ) for 10 days, and transcriptome sequencing and bioinformatics analysis were performed on the root tips of the plants. Result The root length under nitrate treatment was almost twice that under ammonium treatment. 2207 differentially expressed genes were identified from the root tip transcriptome library of poplar treated with two nitrogen forms. Fifty GO functional clusters and 20 KEGG pathways were obtained by means of differential gene GO and KEGG functional clustering analysis, respectively. Furthermore, 36 differentially expressed genes related to nitrogen metabolism pathway, biosynthesis of various amino acids and metabolic process were screened by MapMan analysis. Analysis of the regulatory network of these differential gene interactions showed that nitrate reductase (Potri.005G172400) gene played an important role in affecting root tip growth and development by responding to different nitrogen forms. Conclusion Differentially expressed genes of poplar root tips under different nitrogen form treatments were obtained. The functions of the differentially expressed genes were analyzed, which may help to understand the molecular mechanism of poplar root tip growth and development process by responding to different nitrogen forms. -
Key words:
- nitrate
- / ammonium
- / poplar root tip
- / transcriptome
- / differentially expressed gene
-
图 1 不同氮形态处理后,灰杨根系表型
Figure 1. Morphological parameters of P. × canescens roots with different nitrogen forms
图 2 不同氮形态处理条件下,杨树根尖显著差异表达的基因
Figure 2. Significantly differentially expressed mRNAs in root tips of P. × canescens under different nitrogen form treatments
图 3 不同氮形态处理条件下,灰杨根尖差异表达基因的GO功能聚类注释
Figure 3. Gene Ontology classification annotation of differentially expressed genes in root tips of P. × canescens under different nitrogen form treatments
图 4 KEGG通路分析鉴定差异表达基因
Figure 4. KEGG pathway analysis of significantly differentially expressed target genes
图 5 不同氮形态处理条件下,灰杨根尖差异表达基因的互作网络
Figure 5. The interaction network of differentially expressed genes in root tips of P. × canescens under different nitrogen forms treatments
图 6 不同氮形态处理条件下,灰杨根尖差异基因表达分析
Figure 6. Validation of significantly differentially expressed genes under different nitrogen forms treatments by sRNA-seq and RT-qPCR
表 1 RT-qPCR引物序列
Table 1. Primers used for RT-qPCR
基因 ID gene ID 正向引物 Forward primer 反向引物 Reverse primer Potri.007G102400 ATTCCTGACATGGGCACTGA AGAAGTGATCCAGTCCCTGC Potri.001G010700 TACCATGGGAAAGCTGCTGA AGGCCTTGGAGCAGATCTTT Potri.009G045100 ACAATACCTGCCTCTCCACC GTGGTGCCGGCTTTTCTAAA Potri.003G152300 ATATCCACGAACCGACCTCC TATCGTTTTGACGCTCCTGC Potri.004G116500 CCCGTTATTGCAGCCTCATC AATGAATCCCAGAGCCAGCT Potri.001G391900 GCCTGGCAGTTATACAAGGC AAGTGCTCCTCTCCGAAACA Potri.006G123200 TGGTGACCAAGGCCATATGT AGAATCGTGTGAACCCGGAT Potri.019G131500 AATGGAGAATGTGGGGAGGG TTCCCCTAGCAGTTCCAAGG Potri.005G232600 GTCAATCTACGTGCCGGAAC CCATTGCTGCCATGTCAAGT Potri.007G045300 GGAGTGGACAAGTAGCCCTT TATCACCAGCAGAGCAACCA Potri.005G172400 CTGCTGACAACTGGATCGAA TTTTAACCAGACCGCAAACC Potri.002G113600 GGATGAGTTTCATCGGGCTA CCCGCTTGTAGCAGGAGTAG Potri.008G009500 AGGTTGTAGGAAGGGCTGGT GCCAACGTCTTCCCAACTAA Potri.014G022800 AATTTGGGAGACACCTGTGC CAGCCATTCTGCAGTTCGTA Potri.005G213100 TCCTCTCTGGAGTGCCAAGT ATATCCCTTGCCTCACATGC Potri.001G327000 CGACCAAGATGGGAAGAAAA TTGCAGTGGAGGTCTGTGAG Actin CCCATTGAGCACGGTATTGT TACGACCACTGGCATACAGG  下载: 导出CSV
下载: 导出CSV
表 2 转录组文库测序数据统计
Table 2. Distribution of RNA-seq in different categories
样品
Sample原始数据
Raw data有效数据
Valid data有效比例
Valid ratio(reads) /%比对碱基数
Mapped readsQ20% Q30% GC含量
GC content/%NO3− 57407539 57001409 99.29 46521558(81.59%) 99.51 95.89 45 NH4+ 57945941 57517359 99.26 47517203(82.60%) 99.44 95.95 45
下载: 导出CSV
表 3 氮代谢相关差异表达基因
Table 3. Differentially expressed genes related to nitrogen metabolism
基因名字 Gene name 调节类型 Regulation 功能注释 Description log2(fc) p value Potri.005G172400 up nitrate reductase family protein 4.18 0.0049 Potri.002G088600 up nitrate reductase family protein 3.84 0.0037 Potri.004G140800 up Ferredoxin--nitrite reductase family protein 3.03 0.0117 Potri.008G200100 up glutamine synthetase family protein 1.78 0.0174 Potri.012G043900 up glutamine synthetase family protein 1.02 0.0153 Potri.015G111000 down glutamate dehydrogenase [NAD(P)+] −1.82 0.0194 Potri.012G113500 down glutamate dehydrogenase [NAD(P)+] −2.68 0.0036 Potri.002G146100 down nitroreductase family protein −1.47 0.0469 Potri.014G143300 up aspartate aminotransferase 1.11 0.0335 Potri.001G162800 down alanine aminotransferas family protein −1.34 0.0344 Potri.002G106200 up argininosuccinatelyase 1.03 0.0231 Potri.006G123200 up S-adenosylmethionine synthetase family protein 1.78 0.0000 Potri.014G114700 up S-adenosylmethionine synthetase 1 family protein 1.60 0.0011 Potri.004G190900 up methionine synthase 1.18 0.0031 Potri.005G213100 up selenocysteine methyltransferase family protein 1.43 0.0222 Potri.010G083600 down Homocysteine S-methyltransferase 3 family protein −1.57 0.0370 Potri.013G099500 up aspartate kinase family protein 1.38 0.0252 Potri.002G113600 down branched-chain-amino-acid transaminase −1.71 0.0218 Potri.014G022800 up D-3-phosphoglycerate dehydrogenase 1.38 0.0163 Potri.008G009500 up phosphoglycerate dehydrogenase 1.30 0.0033 Potri.T069600 up hypothetical protein 1.89 0.0311 Potri.019G038900 up hypothetical protein 1.41 0.0111 Potri.005G048200 up O-acetylserine(thiol)lyase family protein 1.13 0.0020 Potri.011G004700 up prephenate dehydratase family protein 1.57 0.0172 Potri.013G066500 up C2 domain-containing protein 1.02 0.0190 Potri.001G348600 down L-aspartate oxidase −1.07 0.0390 Potri.017G123300 up methionyl-tRNAformyltransferase 1.52 0.0405 Potri.003G187100 down methionine gamma-lyase −1.52 0.0001 Potri.001G005400 up protein lysine decarboxylase-like protein 1.43 0.0494 Potri.009G010800 down protein carboxy-lyase −1.25 0.0282 Potri.019G018500 down L-allo-threonine aldolase-related family protein −2.50 0.0386 Potri.001G374100 down homogentisate 1,2-dioxygenase −1.10 0.0203 Potri.002G057700 up enoyl-CoA hydratase/isomerase family protein 1.73 0.0103 Potri.001G327000 down NA −1.12 0.0044 Potri.014G179400 down nitrate transmembrane transporter 1.5 −1.69 0.0462 Potri.003G088800 down nitrate transmembrane transporter 1.5 −3.71 0.0094
下载: 导出CSV
-
[1] Kant S. Understanding nitrate uptake, signaling and remobilisation for improving plant nitrogen use efficiency[J]. Semin Cell Dev Biol, 2018, 74: 89-96. doi: 10.1016/j.semcdb.2017.08.034 [2] Krisztina Ötvös, Marco Marconi, Andrea Vega, et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport[J]. EMBO J, 2021, 40(3): e106862. [3] Luo J, Zhou J-J, et al. Growth performance, photosynthesis, and root characteristics are associated with nitrogen use efficiency in six poplar species[J]. Environmental and Experimental Botany, 2019, 164: 40-51. doi: 10.1016/j.envexpbot.2019.04.013 [4] Rennenberg H, Wildhagen H, Ehlting B. Nitrogen nutrition of poplar trees[J]. Plant Biology, 2010, 12(2): 275-291. doi: 10.1111/j.1438-8677.2009.00309.x [5] 卢孟柱, 胡建军, 我国转基因杨树的研究及应用现状[J]. 林业科技开发, 2007, 20(6): 1-4. [6] Qu CP, Xu ZR, Hu YB, et al. RNA-SEQ reveals transcriptional level changes of poplar roots in different forms of nitrogen treatments[J]. Front Plant Sci, 2016, 7: 51. [7] Zhou J, Lu Y, Shi WG, et al. Physiological characteristics and RNA sequencing in two root zones with contrasting nitrate assimilation of Populus × canescens[J]. Tree Physiol, 2020, 40(10): 1392-1404. Zhang Y, Fang L, Jing P. Analysis of broad leaved forest carbon sinks changes and forest economics and management in China[J]. Environmental Science and Pollution Research, 2019, 27(12): 12922-12931. [8] Rewald B, Kunze ME, Godbold DL. NH4 : NO3 nutrition influence on biomass productivity and root respiration of poplar and willow clones[J]. GCB Bioenergy, 2016, 8(1): 51-58. Castro-Rodríguez V, García-Gutiérrez A, Canales J, et al. Poplar trees for phytoremediation of high levels of nitrate and applications in bioenergy[J]. Plant Biotechnology Journal, 2016, 14(1): 299-312. [9] Wei H, Yordanov YS, Georgieva T, et al. Nitrogen deprivation promotes Populus root growth through global transcriptome reprogramming and activation of hierarchical genetic networks[J]. New Phytol, 2013, 200(2): 483-497. doi: 10.1111/nph.12375 [10] Dash M, Yordanov YS, Georgieva T, et al. A network of genes associated with poplar root development in response to low nitrogen[J]. Plant Signal Behav, 2016, 11(8): e1214792. doi: 10.1080/15592324.2016.1214792 [11] Zhou X, Jacobs TB, Xue LJ, et al. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial populus reveals 4-coumarate: CoA ligase specificity and redundancy[J]. New Phytol, 2015, 208(2): 298-301. doi: 10.1111/nph.13470 [12] Mortazavi A, Williams BA, McCue K, et al. Mapping and quantifying mammalian transcriptomes by RNA-Seq[J]. Nat Methods, 2008, 5(7): 621-628. doi: 10.1038/nmeth.1226 [13] Conesa A, Götz S, García-Gómez JM, et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research[J]. Bioinformatics, 2005, 21(18): 3674-3676. doi: 10.1093/bioinformatics/bti610 [14] Kanehisa M, Araki M, Goto S, et al. KEGG for linking genomes to life and the environment[J]. Nucleic Acids Res, 2008, 36(Database issue): D480-484. [15] Lu Y, Deng S, Li Z, et al. Competing endogenous RNA networks underlying anatomical and physiological characteristics of poplar wood in acclimation to low nitrogen availability[J]. Plant Cell Physiology, 2019, 60(11): 2478-2495. doi: 10.1093/pcp/pcz146 [16] Sorin C, Declerck M, Christ A, et al. A miR169 isoform regulates specific NF-YA targets and root architecture in Arabidopsis[J]. New Phytologist, 2014, 202(4): 1197-1211. doi: 10.1111/nph.12735 [17] Gutiérrez RA. Systems biology for enhanced plant nitrogen nutrition[J]. Science, 2012, 336(6089): 1673-1675. doi: 10.1126/science.1217620 [18] Qu B, He X, Wang J, et al. A wheat CCAAT box-binding transcription factor increases the grain yield of wheat with less fertilizer input[J]. Plant Physiology, 2015, 167(2): 411-423. doi: 10.1104/pp.114.246959 [19] Bouguyon E, Brun F, Meynard D, et al. Multiple mechanisms of nitrate sensing by Arabidopsis nitrate transceptor NRT1.1[J]. Nature Plants, 2015, 1(3): 15015. doi: 10.1038/nplants.2015.15 [20] Fu YF, Zhang ZW, Yang XY, et al. Nitrate reductase is a key enzyme responsible for nitrogen-regulated auxin accumulation in Arabidopsis roots[J]. Biochem Biophys Res Commun, 2020, 532(4): 633-639. doi: 10.1016/j.bbrc.2020.08.057 [21] Sun H, Li J, Song W, Tao J, et al. Nitric oxide generated by nitrate reductase increases nitrogen uptake capacity by inducing lateral root formation and inorganic nitrogen uptake under partial nitrate nutrition in rice[J]. J Exp Bot, 2015, 66(9): 2449-2459. doi: 10.1093/jxb/erv030 [22] Kung JT, Colognori D, Lee JT. Long noncoding RNAs: past, present, and future[J]. Genetics, 2013, 193(3): 651-669. doi: 10.1534/genetics.112.146704 [23] 索沛蘅, 杜大俊, 王玉哲, 等. 杉木连栽对土壤氮含量和氮转化酶活性的影响[J]. 森林与环境学报, 2019, 39(2):113-119. [24] Le Luo, Yali Zhang, Guohua Xu. How does nitrogen shape plant architecture?[J]. J Exp Bot, 2020, 71(15): 4415-4427. doi: 10.1093/jxb/eraa187 [25] Oldroyd GED, Leyser O. A plant’s diet, surviving in a variable nutrient environment[J]. Science, 2020, 368(6486): eaba0196. doi: 10.1126/science.aba0196 [26] Wang S, Lu T, Xue Q, et al. Antioxidation and symbiotic nitrogen fixation function of prxA gene in Mesorhizobium huakuii[J]. Microbiologyopen, 2019, 8(10): e889. [27] Müller SM, Wang S, Telman W, et al. The redox-sensitive module of cyclophilin 20-3, 2-cysteine peroxiredoxin and cysteine synthase integrates sulfur metabolism and oxylipin signaling in the high light acclimation response[J]. Plant J, 2017, 91(6): 995-1014. doi: 10.1111/tpj.13622 [28] Li H, Hu B, Wang W, et al. Identification of microRNAs in rice root in response to nitrate and ammonium[J]. J Genet Genomics, 2016, 43(11): 651-661. doi: 10.1016/j.jgg.2015.12.002 [29] Yu C, Chen Y, Cao Y, et al. Overexpression of miR169o, an overlapping microRNA in response to both nitrogen limitation and bacterial infection, promotes nitrogen use efficiency and susceptibility to bacterial blight in rice[J]. Plant and Cell Physiology, 2018, 59(6): 1234-1247. doi: 10.1093/pcp/pcy060 [30] Gifford ML, Dean A, Gutierrez RA, et al. Cell-specific nitrogen responses mediate developmental plasticity[J]. PNAS, 2008, 105(2): 803-808. doi: 10.1073/pnas.0709559105 [31] Ho CH, Lin SH, Hu HC, et al. CHL1 functions as a nitrate sensor in plants[J]. Cell, 2009, 138(6): 1184-94. doi: 10.1016/j.cell.2009.07.004 [32] Luo J, Zhou J, Li H, et al. Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess[J]. Tree Physioloy, 2015, 35(12): 1283-1302. doi: 10.1093/treephys/tpv091 [33] Luo J, Qin J, He F, et al. Net fluxes of ammonium and nitrate in association with H + fluxes in fine roots of Populus popularis[J]. Planta, 2013, 237(4): 919-931. doi: 10.1007/s00425-012-1807-7 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4697
- HTML全文浏览量: 2772
- PDF下载量: 62
- 被引次数: 0
