

 下载:
下载:

-
红豆杉(Taxus spp.)是举世公认的珍贵林木,在全世界一共有11种,我国存在4种和1变种,即东北红豆杉(T. cuspidata Sieb. et Zucc.)、西藏红豆杉(T. wallichiana Zucc.)、云南红豆杉(T. yunnanensis Cheng et L. K. Fu)、中国红豆杉(T. chinensis (Pilger) Rehd.)以及南方红豆杉(T. chinensis var. mairei (Lemeee et Levl) Cheng et L. K. Fu)[1]。曼地亚红豆杉(Taxus × media Rehder)为一种天然杂交种,于上世纪90年代由美国或加拿大引种于我国。
NAC (NAM/ATAF/CUC)是植物中最常见的转录因子家族之一,在植物生长发育过程中起到重要的调控作用,如响应非生物胁迫、调控器官生长发育以及参与激素信号转导等[2-4]。研究发现,水稻OsNAC2蛋白能够通过影响植物生长素和细胞分裂素(cytokinin)反应基因来调控根的发育[5], 而拟南芥ANAC092可以与生长素应答因子ARF8和PIN4的启动子相结合,通过控制生长素信号途径来影响拟南芥根系的发育[6-8]。此外,在拟南芥中异源表达BnNAC14、GmNAC109、CiNAC3和CiNAC4等其他物种的NAC基因,也可以对侧根的形成起到促进作用[8-11]。这些研究结果表明,NAC转录因子在植物根系的形成过程中扮演重要角色。
近年来,人们为了缓解红豆杉开发与利用的矛盾,在红豆杉人工种植研究上取得了巨大进展,但仍存在红豆杉扦插繁殖生根较慢(大约需要45~60 d)的技术瓶颈。并且,目前关于红豆杉生根的分子机制研究也较为少见。本研究从曼地亚红豆杉三代全长转录组数据中鉴定出了44个NAC转录因子基因,并对该基因家族进行了系统生物信息学及组织表达分析,为探索NAC转录因子调控红豆杉根系的形成奠定理论基础,有助于今后利用分子手段调控其根系生长、缩短扦插育苗时间。
-
通过隐马可夫模型HMM(PF02365) (TBtools v1.064[12])从课题组曼地亚红豆杉三代全长转录组数据(未发表)中获取NAC蛋白序列。利用在线工具CD-HIT Suite进行去冗余[13],利用NCBI CD Search工具进行结构域预测,并筛选结构域完整的序列进行后续分析。
-
利用ExPASy在线工具ProtParam(
https://web.expasy.org/protparam/ )分析TmNAC蛋白理化特性。根据进化树结果,每个亚家族选取一个TmNAC蛋白作为代表,利用phyre2在线网站(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index )进行三级结构建模。 -
拟南芥NAC蛋白序列下载于Plant TFDB (
http://planttfdb.cbi.pku.edu.cn/ )数据库。从138个拟南芥NAC蛋白质序列中随机选取6个序列,用ClustalX2软件将其与44个TmNAC序列进行多序列比对,比对结果利用EsPript 3.0 (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi )进行美化。利用MEGA 7软件中的NJ(Neighbor-Joining)法构建进化树,重复1 000次,其他参数为默认值。iTOL(https://itol.embl.de/ )在线网站美化进化树。利用 MEME在线网站对TmNAC蛋白进行保守基序(Motif)分析[14],最大motif检索数为10,其余为默认参数,将结果通过TBtoolsv1.064软件进行可视化处理[12]。
-
利用云南红豆杉植株根、茎和叶的RNA-seq数据(SRP127697)和TBtools v1.064软件绘制TmNACs基因在根、茎和叶组织中的表达热图,对NAC成员基因表达量FPKM值使用log2 (FPKM + 1)对数转化处理[12]。
-
本研究共获得44条NAC蛋白序列,命名为TmNAC1~TmNAC44。对获得的44个TmNAC蛋白序列进行理化性质分析,发现TmNAC38序列最长,含有1010个氨基酸残基,而TmNAC9序列最短,含有154个氨基酸残基;这些TmNAC蛋白等电点为 4.70~9.60,相对分子质量为18.13~113.57 kDa,GRAVY值均为负值,表明TmNAC蛋白均为亲水蛋白。
-
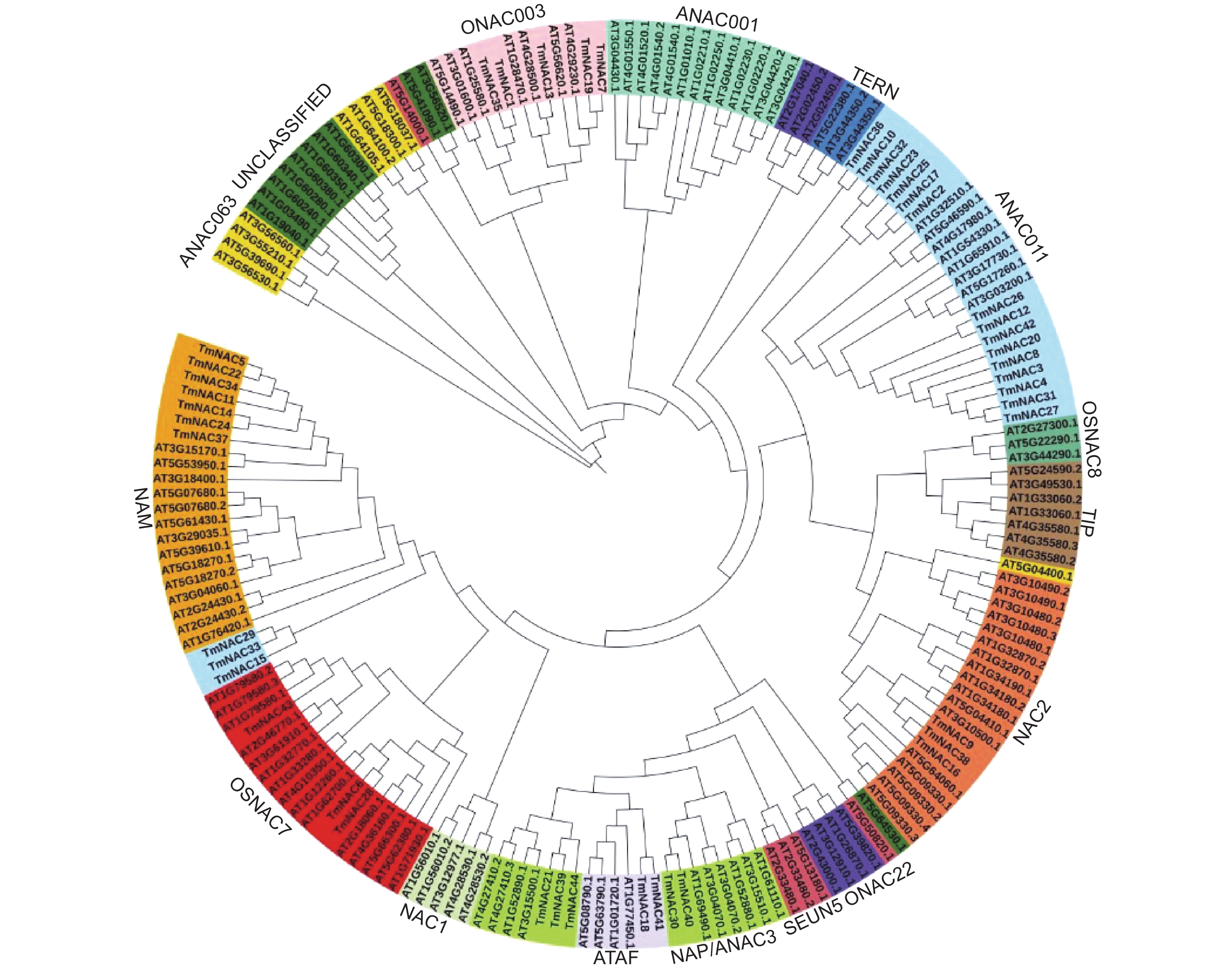
系统进化树结果显示,44个TmNAC 蛋白被分为NAM等7个亚家族,其他NAC1、ONAC22和SEUN5等拟南芥9个NAC亚家族中无TmNACs蛋白分布(图1)。
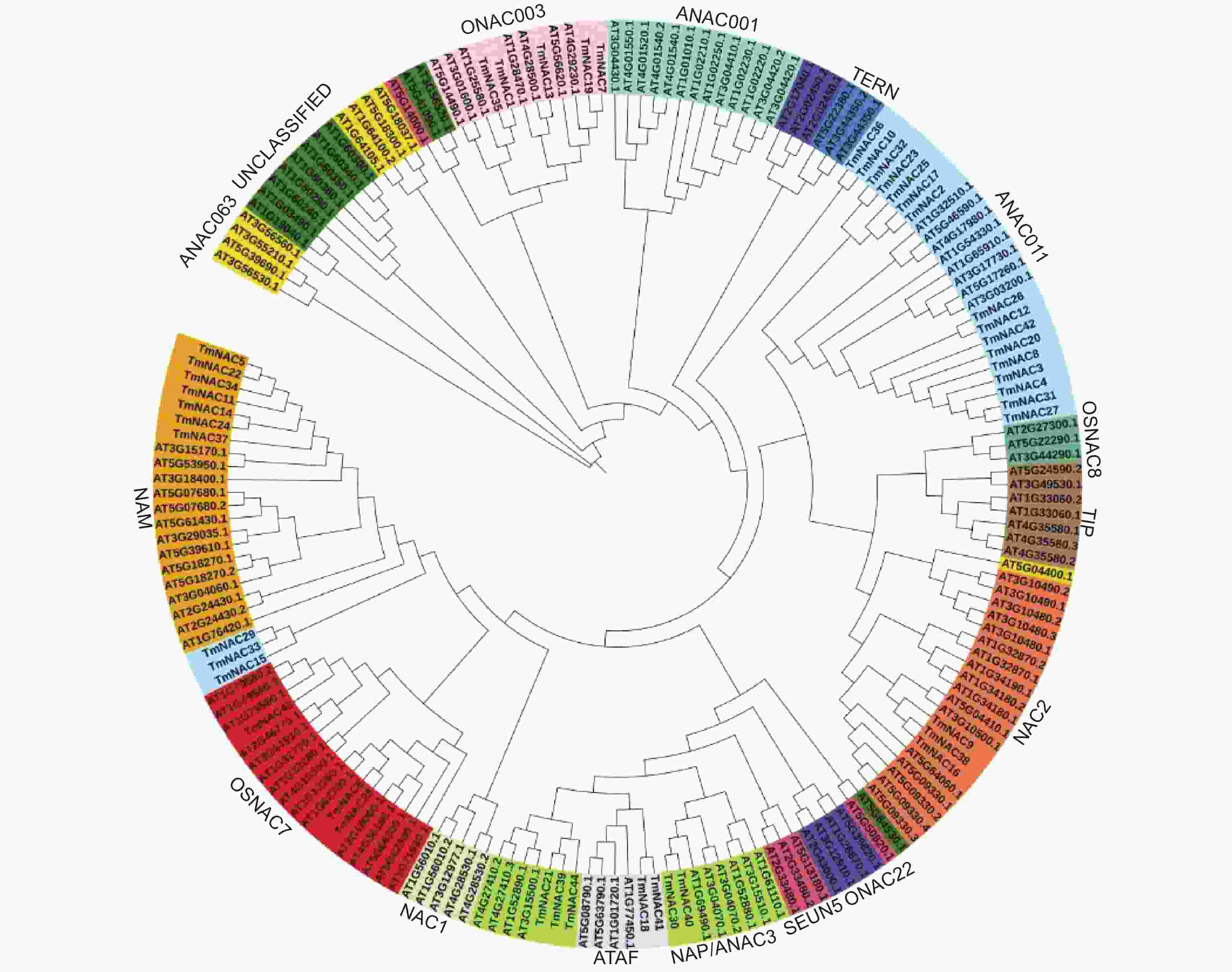
图 1 曼地亚红豆杉与拟南芥NAC家族进化树分析
Figure 1. Phylogenetic relationship among NAC superfamily genes of of Taxus × media and Arabidopsis thaliana
-
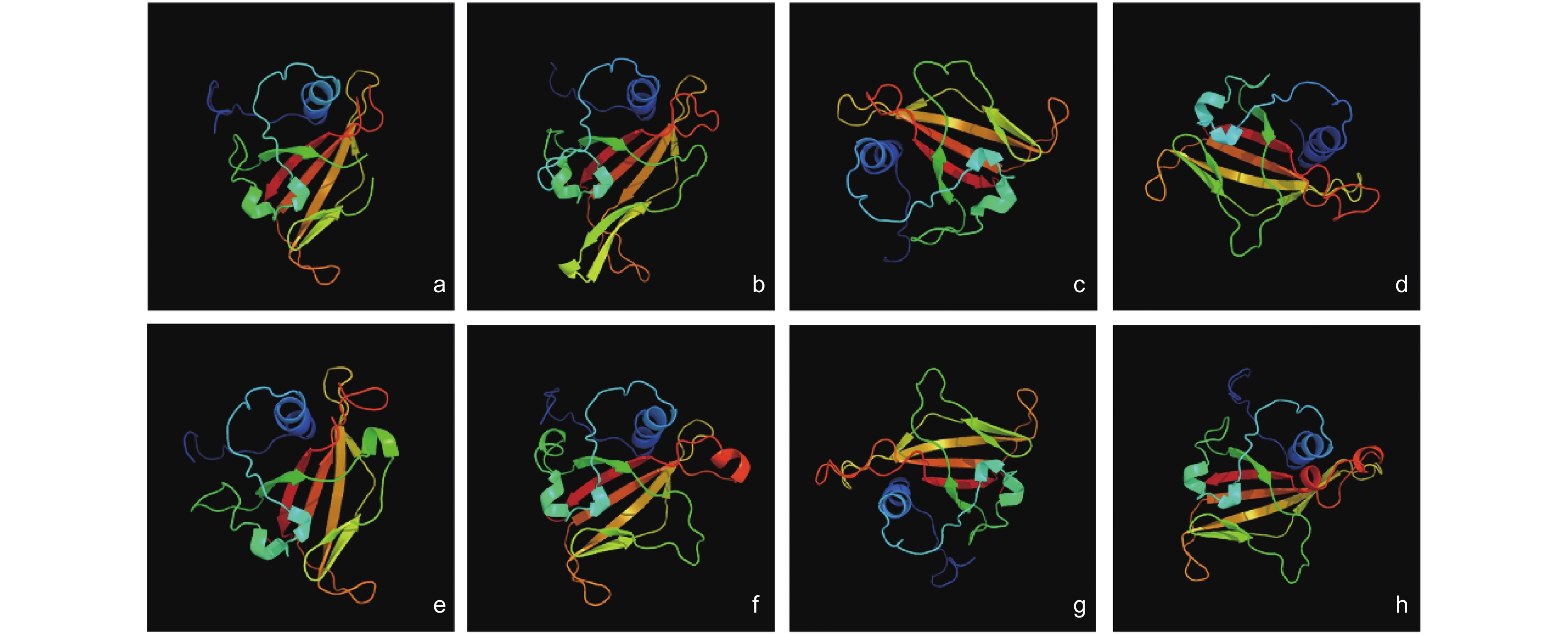
从图1中可以看出,各TmNAC成员的三级结构十分相似,但也存在些许差异,如TmNAC1蛋白比其他蛋白多了一个β-折叠,TmNAC16和TmNAC21蛋白比其他蛋白多一个α螺旋(图2)。
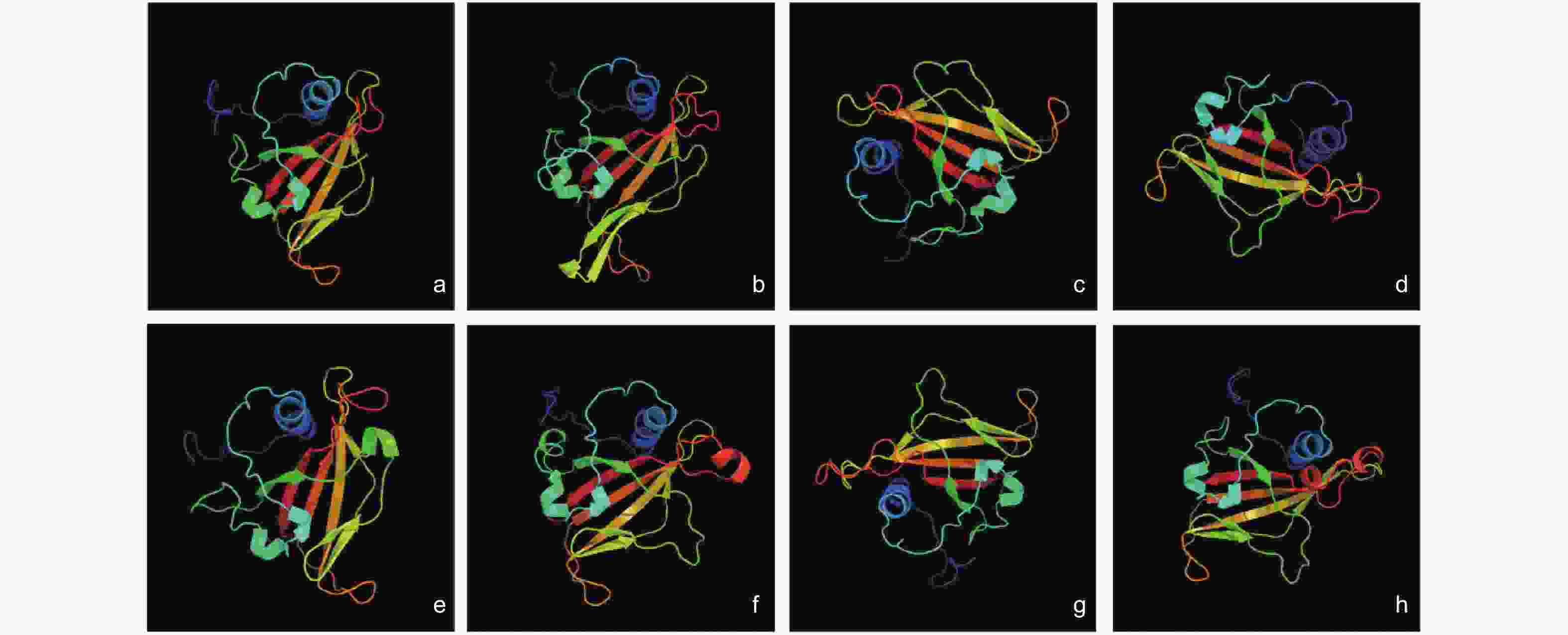
图 2 TmNAC蛋白的三级结构分析
Figure 2. 3D structures of TmNAC proteins
-
多序列比对结果发现,所有的NAC蛋白在N端均具有保守的NAC结构域(图3),其结果与拟南芥和水稻NAC转录因子家族分析结果相似[15]。此外,TmNAC7和TmNAC42蛋白具有不完整的亚结构域D与E (图3),其余蛋白均具有5个保守的亚结构域(A~E)。

图 3 曼地亚红豆杉和拟南芥NAC蛋白保守结构域分析
Figure 3. Analysis of conserved domains of the NAC proteins of Taxus × media and Arabidopsis thaliana
-
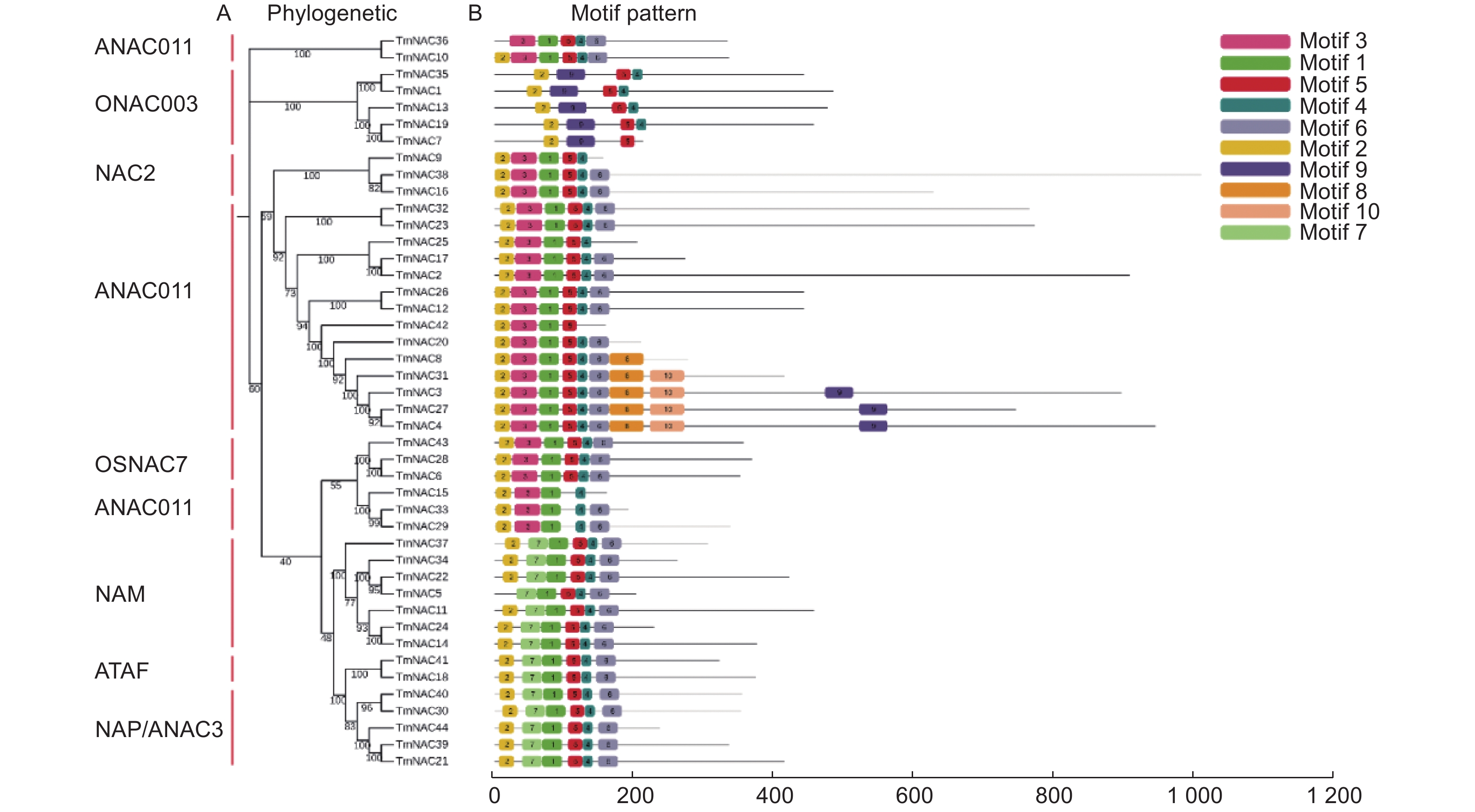
基序分析结果发现motif 2、4、5几乎存在于所有的NAC蛋白成员中。另外,有的亚家族具有或缺失某些蛋白基序,如motif 8、10仅存在于ANAC011亚家族中,motif 7仅存在于NAM与ATAF亚家族中;而ANAC011亚家族中的TmNAC36和TmNAC15则分别缺少motif 2和motif 5 (图4)。
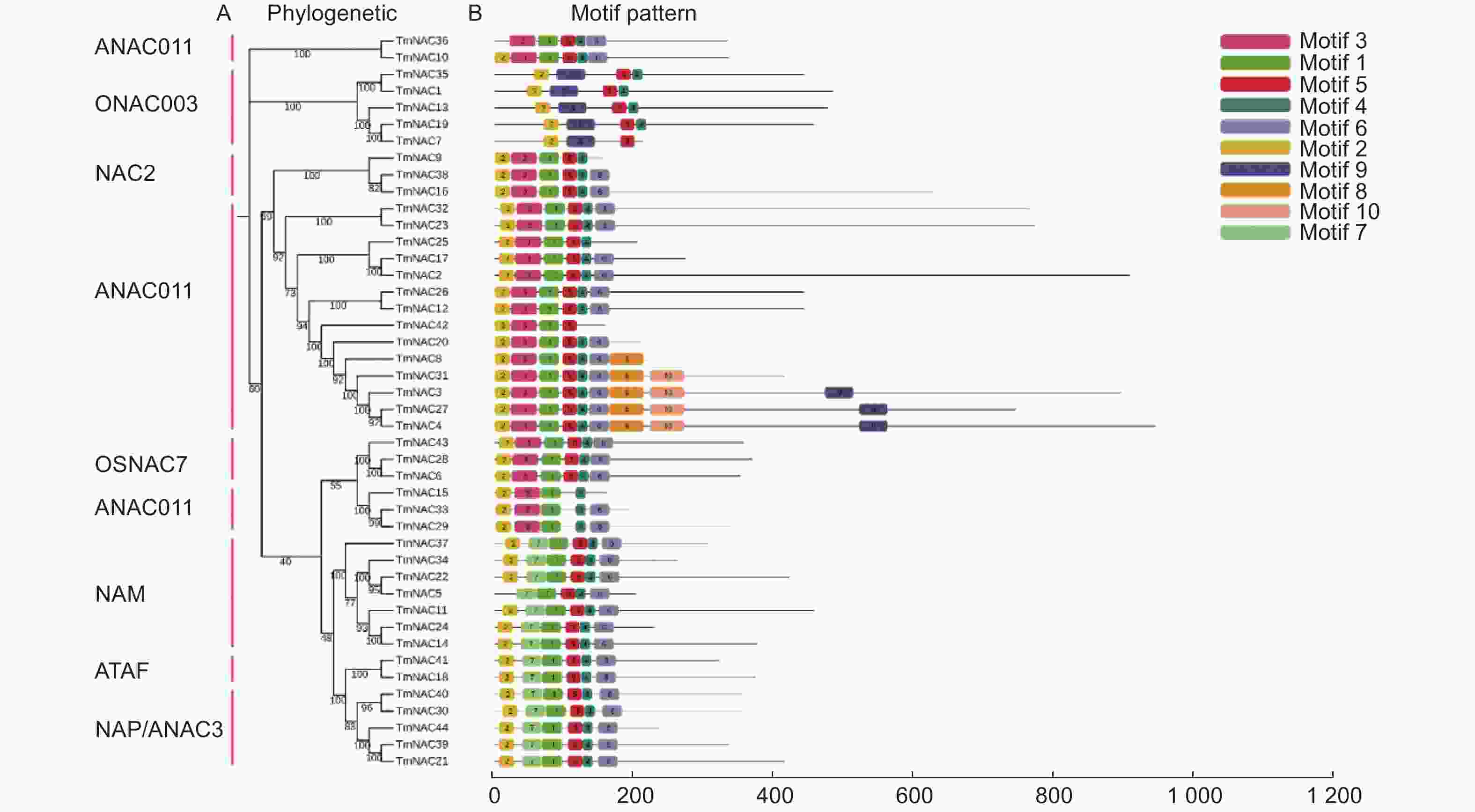
图 4 曼地亚红豆杉TmNAC蛋白motif分析
Figure 4. Motif analysis of TmNAC proteins of Taxus × media
-
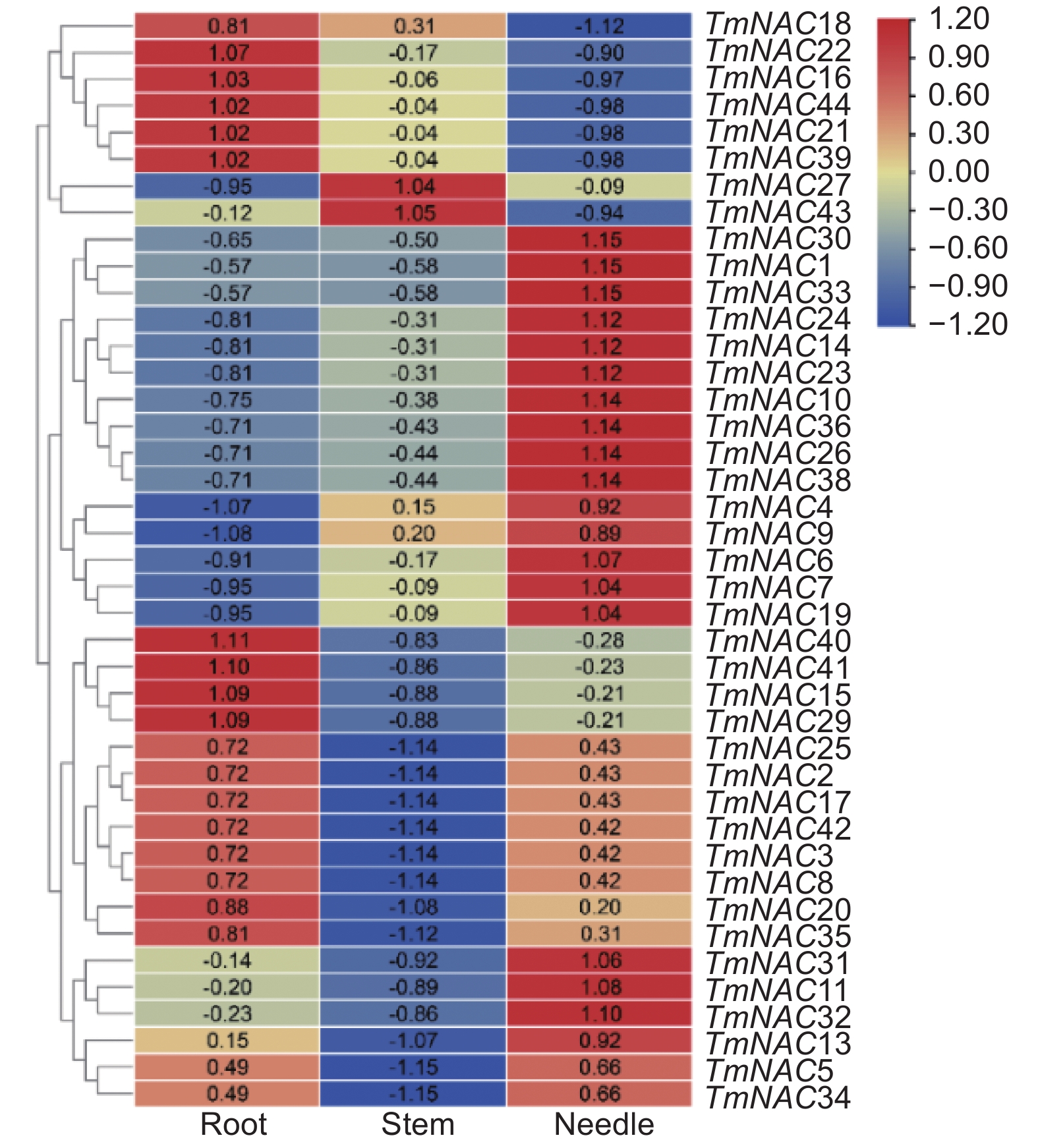
组织表达分析结果显示,TmNAC15、16、18、21、22、29、39、40、41和44在根中均有较高的表达量,而在叶和茎中的表达量都相对较低。另外,TmNAC2、3、8、17、25和42在根与叶中表达量相近,而TmNAC27和TmNAC43在茎中表达量最高,在叶和根中的表达量均较低(图5)。
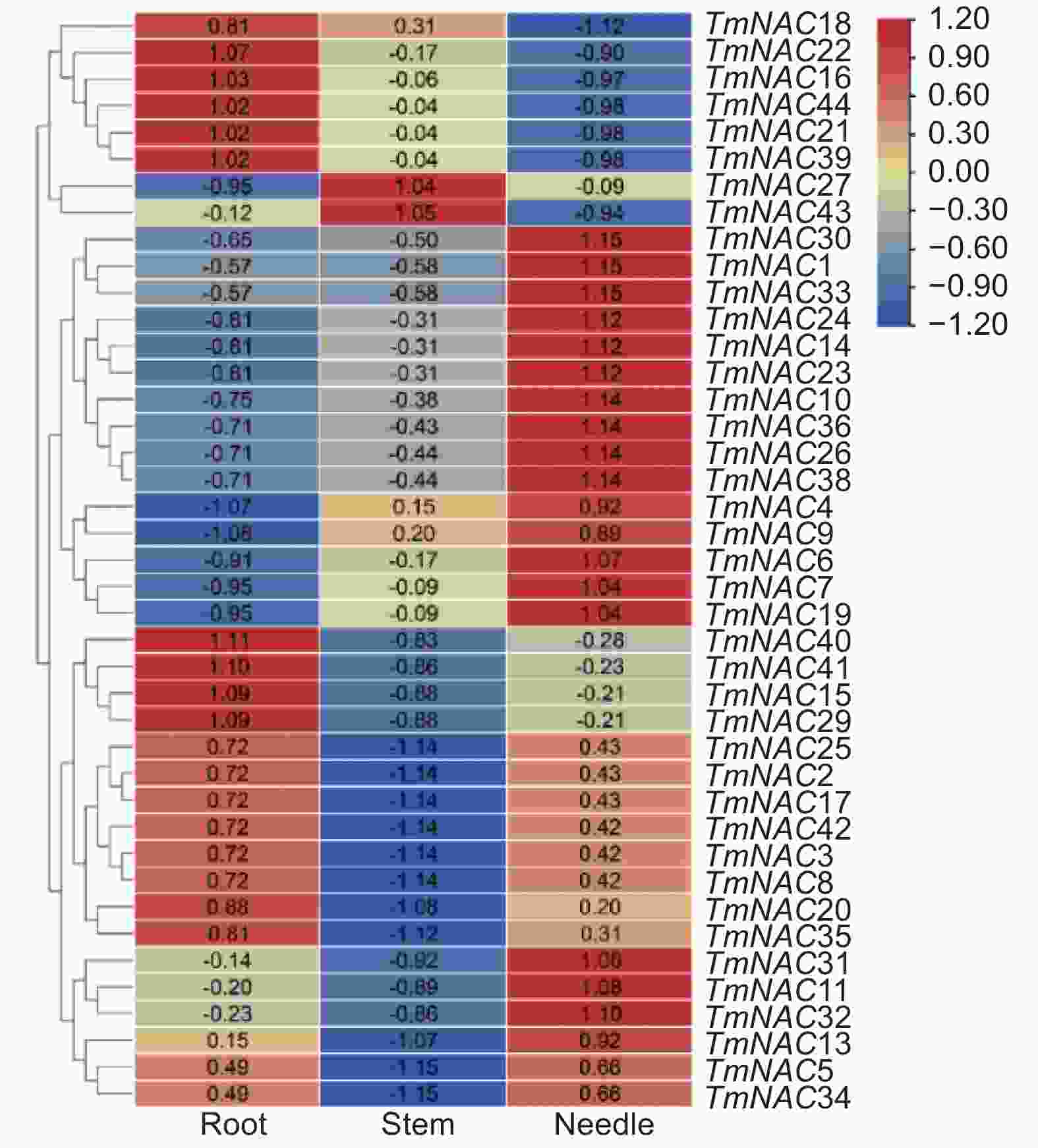
图 5 TmNAC基因在根、茎和叶中表达模式
Figure 5. The expression patterns of TmNAC genes in roots, leaves and stems
-
NAC基因家族具有保守的结构域和功能域,对植物的生长发育具有重要的作用[15-17]。本研究从曼地亚红豆杉转录组数据中鉴定出 44个NAC 转录因子,数目明显少于拟南芥和水稻基因组中的138个和140个[18-19]。究其原因,很有可能是由于本研究的NAC来源于三代全长转录组测序数据,也有可能是因为物种不同造成。目前,已经有西藏红豆杉和南方红豆杉基因组发表[20-21],未来可以利用这些基因组信息对NAC基因家族进一步进行分析,红豆杉NAC家族的完整性可以得到进一步完善和确定。
研究表明NAC蛋白具有典型结构特征,均含有NAC功能域,且NAC区域包含 A~E共5个NAC亚结构域。本研究蛋白多序列比对结果显示,44个TmNAC蛋白在N端均含有 NAC结构域,但TmNAC7和TmNAC42蛋白具有不完整的D和E亚结构域。这5个亚结构域的保守性顺序为A>C>D>B>E,亚结构域A与C对蛋白质的稳定发挥着重要作用。此外,亚结构域 E是NAC 蛋白中一个重要的 DNA 结合区域,可能参与了发育时期调控,并且前人也推测存在由亚结构域D和E组成的DNA结合区[22]。蛋白的功能与其结构密切相关,由此可以推测,TmNAC家族有很大可能与其他植物的NAC蛋白功能相似,而TmNAC7和TmNAC42蛋白功能则极其可能与其他TmNAC蛋白功能有所不同。
进化树分析显示红豆杉TmNAC成员与大多数拟南芥NAC家族各亚家族成员具有较高同源性,表明NAC家族在物种间进化保守。因此,可以通过其他物种NAC蛋白功能来预测红豆杉NAC同源蛋白的功能。此外,拟南芥中NAC1、ONAC22和SEUN5等9个亚家族并不包含红豆杉NAC同源蛋白,这些情况可能是由于曼地亚红豆杉的NAC家族序列是从转录组数据中获得,并未全部获取到基因组中其他亚家族NAC蛋白序列所造成。
多序列比对、motif及蛋白质三级结构分析均发现,44个曼地亚红豆杉NAC蛋白具有较高的相似性和保守性,该结果与桃树、蓖麻、茶树、番茄中的报道相类似,说明其在进化上非常保守[23-26]。值得注意的是,TmNAC1、TmNAC16和TmNAC21三者所在的亚家族与其他亚家族成员结构上存在显著差异,且不同亚家族或同一亚家族不同成员之间motif类型和数目也存在差异,这些结构差异是否导致它们功能发生变异尚不清楚,值得研究人员进一步深入研究。
研究表明,不同的物种间同源基因在进化过程中会保留相同或相似的功能,而基因的组织表达模式往往与其功能密切相关[27]。拟南芥和水稻的一些NAC转录因子的功能已经得到验证,这为预测红豆杉TmNAC转录因子的功能提供了线索。TmNAC家族TmNAC15、16、18、21、22、29、39、40、41和44基因在红豆杉根中表达量较高,同时TmNAC21、30、39、40和44蛋白与拟南芥AtNAC2/AT3G15510蛋白在系统发育树中聚为一支,而AtNAC2恰恰参与侧根的形成与发育[28];TmNAC5、11、14、22、24、34和37蛋白与拟南芥AtNAC1/AT3G15170蛋白在系统发育树中聚为一支,AtNAC1介导生长素信号以促进侧根的形成[29]。综合上述,推测红豆杉TmNAC21、39、40、44(NAP/ANAC3亚家族)和TmNAC22(NAM亚家族)可能在红豆杉根系生长发育中起到重要调控作用。
-
本研究从红豆杉转录组中鉴定出44个NAC成员,发现红豆杉NAC基因编码的44个蛋白在N端均具有保守的NAC结构域,TmNAC家族有很大可能与其他植物的NAC蛋白功能相似。蛋白功能结构域分析结果预示着红豆杉NAC家族成员可能在亚家族间以及亚家族内部具有功能的特异性。表达量比较分析显示,TmNAC基因在红豆杉不同组织中的表达量各不相同,TmNAC21、22、39、40和44可能与红豆杉根的生长发育有关,为进一步探索红豆杉NAC转录因子调控红豆杉根系生长提供理论基础。
曼地亚红豆杉NAC基因家族鉴定及表达分析
Identification and Expression Analysis of NAC Gene Family in Taxus × media Rehder
-
摘要:
目的 筛选红豆杉NAC转录因子家族中影响根系形成的关键基因,探索红豆杉根系生长发育分子机理。 方法 利用曼地亚红豆杉全长转录组数据,通过生物信息学方法鉴定NAC 转录因子,并对筛选出的NAC 转录因子进行蛋白结构及基因组织表达谱等分析。 结果 共鉴定出 44个NAC 转录因子,其在N端均具有典型的保守NAC结构域,分别聚类到拟南芥的7个亚家族中,蛋白三级结构较为相似,且成员大多含有5个保守亚结构域。组织表达谱显示TmNAC15、16、18、21、22、29、39、40、41和44在根中表达水平高于茎和叶。 结论 从曼地亚红豆杉中共鉴定出44个NAC 转录因子,其结构较为保守,且聚类为7个亚家族,其中成员TmNAC21、22、39、40和44极有可能参与红豆杉根系生长发育。 Abstract:Objective To obtain the key NAC genes involving in regulating the root growth and development of yew trees. Method Basing on the full-length transcriptome data of Taxus × media, bioinformatics methods were used to identify NAC transcription factors. The protein structure and gene tissue expression profile of the selected NAC genes were also analyzed. Result A total of 44 NAC transcription factors were identified, which have typical conserved NAC domains at the N-terminal, and most of the members contain 5 conserved structural functional subdomains. Phylogenetic tree analysis showed that 44 T. × media NAC (TmNAC) proteins were clustered into 7 subfamilies of Arabidopsis. Sequence structure analysis indicated that the tertiary structures of the TmNAC proteins were similar, but the structures of the representative member of different subfamily also showed obvious differences. The expression results showed that TmNAC15, 16, 18, 21, 22, 29, 39, 40, 41, and 44 had higher expression level in roots than stems and needles. Conclusion In this study, 44 TmNAC transcription factors were identified from T. × media and they were clustered into 7 subfamilies. Among them, TmNAC21, 22, 39, 40, and 44 may be involved in the root growth and development progresses of yew trees. -
Key words:
- Taxus spp.
- / NAC gene
- / gene identification
- / expression analysis
-
图 1 曼地亚红豆杉与拟南芥NAC家族进化树分析
Figure 1. Phylogenetic relationship among NAC superfamily genes of of Taxus × media and Arabidopsis thaliana
图 3 曼地亚红豆杉和拟南芥NAC蛋白保守结构域分析
Figure 3. Analysis of conserved domains of the NAC proteins of Taxus × media and Arabidopsis thaliana
-
[1] 王亚飞, 王 强, 阮 晓, 等. 红豆杉属植物资源的研究现状与开发利用对策[J]. 林业科学, 2012, 48(5):116-125. doi: 10.11707/j.1001-7488.20120518 [2] 赵 灿, 刘任重, 柳展基, 等. 亚洲棉全基因组中NAC类转录因子基因的鉴定与分析[J]. 石河子大学学报:自科版, 2015, 33(4):397-404. [3] Yang Y F, Zhu K, Wu J, et al. Identification and characterization of a novel NAC-like gene in chrysanthemum (Dendranthema lavandulifolium)[J]. Plant Cell Reports, 2016, 35(8): 1783-98. doi: 10.1007/s00299-016-1996-9 [4] 王立国, 傅明川, 李 浩, 等. 陆地棉NAC转录因子基因GhSNAC1的克隆及其抗旱耐盐分析[J]. 农业生物技术学报, 2019, 27(4):571-580. [5] Mao C, He J, Liu L, et al. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development[J]. Plant Biotechnology Journal, 2020, 18(2): 429-442. doi: 10.1111/pbi.13209 [6] Xi D, Chen X, Wang Y, et al. Arabidopsis ANAC092 regulates auxin-mediated root development by binding to the ARF8 and PIN4 promoters[J]. Journal of Integrative Plant Biology, 2019, 61(9): 1015-1031. doi: 10.1111/jipb.12735 [7] Chen D, Chai S, McIntyre C L, et al. Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance[J]. Plant Cell Reports, 2018, 37(2): 225-237. doi: 10.1007/s00299-017-2224-y [8] Hao Y J, Wei W, Song Q X, et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants[J]. Plant Journal, 2011, 68(2): 302-13. doi: 10.1111/j.1365-313X.2011.04687.x [9] Han X, Feng Z, Xing D, et al. Two NAC transcription factors from Caragana intermedia altered salt tolerance of the transgenic Arabidopsis[J]. BMC Plant Biology, 2015, 15: 208. doi: 10.1186/s12870-015-0591-5 [10] Yang X, Kim M Y, Ha J, et al. Overexpression of the soybean NAC gene GmNAC109 increases lateral root formation and abiotic stress tolerance in transgenic Arabidopsis plants[J]. Frontiers in Plant Science, 2019, 10: 1036. doi: 10.3389/fpls.2019.01036 [11] Hegedus D, Yu M, Baldwin D, et al. Molecular characterization of Brassica napus NAC domain transcriptional activators induced in response to biotic and abiotic stress[J]. Plant Molecular Biology, 2003, 53(3): 383-97. doi: 10.1023/B:PLAN.0000006944.61384.11 [12] Chen C, Chen H, Zhang Y, et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13(8): 1194-1202. doi: 10.1016/j.molp.2020.06.009 [13] Huang Y, Niu B, Gao Y, et al. CD-HIT Suite: a web server for clustering and comparing biological sequences[J]. Bioinformatics, 2010, 26(5): 680-2. doi: 10.1093/bioinformatics/btq003 [14] Bailey T L, Boden M, Buske F A, et al. MEME SUITE: tools for motif discovery and searching[J]. Nucleic Acids Research, 2009, 37 (Web Server issue): W202-8. [15] Sun H, Hu M, Li J, et al. Comprehensive analysis of NAC transcription factors uncovers their roles during fiber development and stress response in cotton[J]. BMC Plant Biology, 2018, 18(1): 150. doi: 10.1186/s12870-018-1367-5 [16] Zhuo X, Zheng T, Zhang Z, et al. Genome-wide analysis of the NAC transcription factor gene family reveals differential expression patterns and cold-stress responses in the woody plant Prunus mume[J]. Genes (Basel), 2018, 9(10): 494. doi: 10.3390/genes9100494 [17] Gong X, Zhao L, Song X, et al. Genome-wide analyses and expression patterns under abiotic stress of NAC transcription factors in white pear (Pyrus bretschneideri)[J]. BMC Plant Biology, 2019, 19(1): 161. doi: 10.1186/s12870-019-1760-8 [18] Ooka H, Satoh K, Doi K, et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana[J]. DNA Research, 2003, 10(6): 239-47. doi: 10.1093/dnares/10.6.239 [19] Fang Y, You J, Xie K, et al. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice[J]. Molecular Genetics and Genomics, 2008, 280(6): 547-63. doi: 10.1007/s00438-008-0386-6 [20] Cheng J, Wang X, Liu X, et al. Chromosome-level genome of Himalayan yew provides insights into the origin and evolution of the paclitaxel biosynthetic pathway[J]. Molecular Plant, 2021, S1674-2052(21): 00160-X. [21] Xiong X, Gou J, Liao Q, et al. The Taxus genome provides insights into paclitaxel biosynthesis[J]. BioRxiv, 2021: 2021.04. 29.441981. [22] Ernst H A, Olsen A N, Larsen S, et al. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors[J]. EMBO Reports, 2004, 5(3): 297-303. doi: 10.1038/sj.embor.7400093 [23] 张春华, 上官凌飞, 俞明亮, 等. 桃NAC基因家族生物信息学分析[J]. 江苏农业学报, 2012, 28(2):406-414. doi: 10.3969/j.issn.1000-4440.2012.02.031 [24] 代梦媛, 高 梅, 李文昌. 蓖麻NAC转录因子家族的鉴定及生物信息学分析[J]. 分子植物育种, 2020, 18(6):1808-1817. [25] 韩雅彭, 程 琳, 杨凌霄. 茶树NAC转录因子家族的鉴定及生物信息学分析[J]. 河南大学学报:自然版, 2017(47):301-309. [26] 姜秀明, 牛义岭, 许向阳. 番茄NAC基因家族的系统进化及表达分析[J]. 分子植物育种, 2016, 14(8):1948-1964. [27] 王 震, 米要磊, 孟祥霄, 等. 中药火麻仁基原植物大麻LBD基因家族成员的鉴定与表达分析[J]. 中国中药杂志, 2020, 45(22):175-184. [28] He X J, Mu R L, Cao W H, et al. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development[J]. Plant Journal, 2005, 44(6): 903-16. doi: 10.1111/j.1365-313X.2005.02575.x [29] Xie Q, Frugis G, Colgan D, et al. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development[J]. Genes & Development, 2000, 14(23): 3024-3036. -

 点击查看大图
点击查看大图
图(5)
计量
- 文章访问数: 4947
- HTML全文浏览量: 2132
- PDF下载量: 52
- 被引次数: 0
