



-
生态位是现代生态学的研究热点之一,Johnson于1910年率先提出了生态位概念[1]。1917年,Grinnell将生态位正式定义为种的最后分布单元,且生态位实际上是空间生态位[2-3];Elton于1927年对Grinnell生态位概念做了进一步完善,强调生态位是指物种在生物群落中的作用和地位[4];Hutchinson在1958年提出了现代的生态位概念,他从空间和资源利用等方面考虑,提出“多维超体积生态位”,把生态位看成种群所能生存条件的总和[2,5]。之后,Levins,Pianka和Schoener,Hurlbert等人对生态位的定义更加严格化和定量化,并提出了生态位宽度和生态位重叠两种测度方法,至今仍被广泛使用[6-9]。上世纪80年代开始,我国学者王刚[10]、刘建国和马世骏[11]、张桂莲和张金屯[12]等开始对生态位进行了深入研究,从不同的角度给出了生态位的定义。
生态位宽度反映物种对环境的适应性,在资源量非常有限的情况下,物种生态位宽度一般增加,以使种群得到足够的资源[2,13]。生态位重叠值大小反映物种之间利用资源的相似程度,当两个物种共同占有同一资源(食物、营养成分、空间等)时,就会出现生态位重叠现象。生态位重叠包括完全重叠和部分重叠,完全重叠是指两个物种具有完全一样的生态位,这种重叠几率较小,大多数情况下生态位之间只发生部分重叠,即一部分资源是被共同利用的,其他部分则是被各自所占据[2]。Hutchinson认为两个物种发生竞争的前提条件是生态位重叠,他假设环境在充分饱和的情况下,任何两个物种生态位重叠都必然发生竞争排斥作用[2,14]。但事实上,任何两个物种生态位重叠并不一定能导致竞争,除非共用的空间资源不足[2]。
近年来,我国有不少学者就不同地区植物种群生态位展开了大量研究,并得出了丰硕的成果,但系统地对火烧迹地植被群落(尤其是对不同年代火烧迹地灌木群落生态位的研究)种群的生态位研究较少,只有闫东锋和金山等人对火烧迹地草本群落生态位进行过相应的研究[15-17]。本研究以青藏高原东北边缘云-冷杉林不同年限火烧迹地灌木群落和未过火林地林下灌木层为研究对象,探讨灌木种群生态位宽度、种间生态位重叠,并划分群落泛化种和特化种,揭示青藏高原东北边缘云-冷杉林火烧迹地灌木群落种群生态位特征,为当地林草部门对火烧迹地植被的恢复和演替进程中进行合理的干预、引种和造林等提供合理的依据。
-
研究区位于青藏高原东北边缘(103°12′~103°21′ E,33°49′~34°10′ N),该区域地形复杂,地域辽阔,白龙江穿越其东南部,黄河又在其西南部折流,是长江流域和黄河流域的重要分水岭[18-19]。属于甘南高寒湿润区,处于大陆性气候与季风气候的过渡带上,年平均气温8~11℃,无霜期147 d,平均降水量635 mm,平均海拔3 200 m,地形相对高差约1 200 m,平均坡度30°~50°,平均土层深度约70 cm,土壤总体属棕壤草甸土区,以山地棕壤、山地灰棕壤、亚高山草甸土为主。历年来,该区域曾多年发生过森林火灾,次数频繁,森林生态系统遭受了严重毁坏。研究区森林火灾分别发生于2005年4月19日和2016年3月2日,均为重度森林火灾,过火后其森林生态系统遭到毁灭性破坏,火烧迹地上大部分生物多样性丧失,现均以灌木植物为建群种[18-19]。目前恢复5 a火烧迹地主要有菰帽悬钩子(Rubus pileatus Focke)、秀丽莓(R. amabilis Focke)等;恢复15 a火烧迹地主要有唐古特忍冬(Lonicera tangutica Maxim.)、扁刺蔷薇(Rosa sweginzowii Koehne)、黑水柳(Salix heishuiensis N. Chao)、山梅花(Philadelphus incanus Koehne)、峨眉蔷薇(R. omeiensis Rolfe)等[18-19]。
-
通过前期对火烧迹地资料收集和有效信息的筛选,2019年5月和8月实地勘察和样地初选,2020年8月和2021年7月至8月,分别在5 a、15 a云-冷杉林火烧迹地设置20 m × 20 m的样地,并在未过火云-冷杉林中设置20 m × 20 m对照样地,共计22个。在每个样地中设置5个5 m × 5 m灌木样方,共设置110个灌木样方,5 a、15 a火烧迹地各30个样方,共计60个;未过火林地50个样方。在每个样方中测定灌木种类、地径、平均树高、盖度、株(丛)数、冠幅等[18-19]。
-
用Levins和Shannon Wiener生态位宽度来度量[2,20-21],公式如下:
(1)Levins生态位宽度:
$ {B_{i1}} = \frac{1}{{\sum\limits_{j = 1}^r {{{({P_{ij}})}^2}} }} $
(2)Shannon Wiener生态位宽度:
$ {B_{i2}} = - \sum\limits_{j = 1}^r {({P_{ij}}\ln {P_{ij}}} ) $
其中Bi为第i种的生态位宽度,j表示样方,r表示样方的数量。
-
用Pianka和Schoener生态位重叠指数来度量[2,20-21],公式如下:
(1)Pianka生态位重叠
$ O_{ik1} = \frac{{\sum\limits_{j = 1}^r {P_{ij}P_{kj}} }}{{\sqrt {{{\left( {\sum\limits_{j = 1}^r {P_{ij}} } \right)}^2}{{\left( {\sum\limits_{j = 1}^r {P_{kj}} } \right)}^2}} }} $
(2)Schoener生态位重叠
$ {O_{ik2}} = 1 - \frac{1}{2}\sum\limits_{j = 1}^r {|{P_{ij}} - {P_{kj}}} | $
其中,Oik为种i和种k的生态位重叠系数,Pij和Pkj分别为种i和种k在第j个样方的多度,r为总的样方数。
-
采用Shannon Wiener和Levins生态位宽度两种方法,通过EcolUtils包置换方法对物种的出现频率进行999次随机化重排,并将生态位宽度指数超过95%置信区间上限的物种定义为泛化种,低于零分布的95%置信区间下限的物种定义为特化种[22-23]。
基于Excel 2016进行基础数据整理,所有数据的计算和作图都在R 4.0.5中实现,其中生态位宽度和生态位重叠指数分别在spaa包中完成。泛化种和特化种的划分在EcolUtils包中完成。
-
采用Levins和Shannon Wiener生态位宽度两种方法研究不同年限火烧迹地灌木群落和未过火林地林下灌木层物种的生态位宽度(表1)。结果显示,两种方法所得到的各物种生态位宽度大小变化趋势基本相似,相同物种在不同年限火烧迹地和未过火林地生态位宽度大小存在差异。
表 1 火烧迹地灌木群落和未过火林地林下灌木层物种生态位宽度
Table 1. Niche width of shrub communities in burned and unburned areas
物种 Species 5 a 15 a 未过火林地 unburned area Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener 唐古特忍冬 Lonicera tangutica 8.32 2.24 19.19 3.10 20.20 3.18 菰帽悬钩子 Rubus pileatus 12.92 2.74 — — 16.01 2.90 扁刺蔷薇 Rosa sweginzowii 7.14 2.16 11.85 2.60 13.54 2.78 箭竹 Fargesia spathacea 5.56 1.95 15.06 2.90 13.39 2.96 山梅花 Philadelphus incanus 15.01 2.94 17.61 3.01 10.91 2.57 大刺茶藨子 Ribes alpestre 11.10 2.57 11.71 2.82 10.25 2.42 美丽茶藨子 R. pulchellum 13.27 2.87 — — 8.38 2.31 天山茶藨子 R. meyeri — — — — 7.31 2.15 峨眉蔷薇 R. omeiensis — — 22.44 3.19 7.25 2.07 刺果茶藨子 R. burejense — — — — 6.81 2.16 毛花忍冬 L. trichosantha 4.00 1.39 8.43 2.34 6.78 2.26 小叶蔷薇 R. willmottiae 1.00 0.00 4.12 1.59 6.57 2.07 蓝靛果忍冬 L. caerulea 1.00 0.00 5.26 1.72 6.13 1.95 甘肃小檗 Berberis kansuensis 1.00 0.00 2.00 0.69 6.00 1.79 陕甘花楸 Sorbus koehneana — — — — 5.00 1.61 灰栒子 Cotoneaster acutifolius 2.78 1.05 1.00 0.00 4.50 1.72 川西锦鸡儿 Caragana sinica — — — — 4.24 1.55 秀丽莓 R. amabilis — — — — 3.82 1.66 刺五加 Eleutherococcus senticosus 6.41 2.22 — — 3.32 1.32 东陵绣球 Hydrangea bretschneideri 4.67 1.73 — — 3.27 1.28 葱皮忍冬 L. ferdinandi — — — — 3.17 1.30 珍珠梅 Sorbaria sorbifolia 11.78 2.59 — — 3.07 1.28 黑果菝葜 Smilax glaucochina — — — — 3.00 1.50 托柄菝葜 S. discotis 2.70 1.40 — — 2.79 1.16 楤木 Aralia elata 4.03 1.89 — — 2.00 0.87 杯腺柳 Salix cupularis — — — — 2.00 0.69 卫矛 Euonymus alatus — — — — 2.00 0.69 金银忍冬 L. maackii — — — — 1.80 0.64 小叶忍冬 L. microphylla 1.00 0.00 — — 1.69 0.60 红脉忍冬 L. nervosa 2.00 0.69 3.60 1.33 1.68 0.74 六道木 Zabelia biflora — — 1.00 0.00 1.47 0.50 长果茶藨子 R. stenocarpum — — — — 1.17 0.27 荚蒾 Viburnum dilatatum 1.60 0.56 — — 1.00 0.00 黑水柳 S. heishuiensis 10.81 2.62 10.91 2.66 1.00 0.00 秦岭蔷薇 R. tsinglingensis — — 3.63 1.33 1.00 0.00 藤山柳 Clematoclethra scandens 3.38 1.30 2.99 1.29 — — 尖叶栒子 C. acuminatus — — 1.00 0.00 — — 接骨木 Sambucus williamsii 5.40 1.83 — — — — 圆锥绣球 Hydrangea paniculata 8.50 2.33 — — — — 5 a火烧迹地灌木物种Levins和Shannon Wiener生态位宽度排名前20%的是山梅花(P. incanus Koehne)、秀丽莓(R. amabilis Focke)、菰帽悬钩子(R. pileatus Focke)、珍珠梅(Sorbaria sorbifolia (L.) A. Braun)和大刺茶藨子(R. alpestre Wall. ex Decne.),两种方法计算结果均表明山梅花生态位宽度值最大,Levins和Shannon Wiener生态位宽度分别为15.01和2.94,其后依次是秀丽莓、菰帽悬钩子、珍珠梅和大刺茶藨子,Levins和Shannon Wiener生态位宽度值分别是13.27、12.92、11.78、11.10和2.87、2.74、2.59、2.57。生态位宽度排名后20%的物种是荚蒾(Viburnum dilatatum Thunb.)、甘肃小檗(Viburnum dilatatum C. K. Schneid.)、蓝靛果忍冬(L. caerulea L.)、小叶蔷薇(R. willmottiae Hemsl.)和小叶忍冬(L. microphylla Willd. ex Roem. & Schult.),其中荚蒾Levins和Shannon Wiener生态位宽度值分别是1.60和0.56,甘肃小檗、蓝靛果忍冬、小叶蔷薇和小叶忍冬Levins和Shannon Wiener生态位宽度值最小,均为1.00和0.00。Levins和Shannon Wiener生态位宽度值排名处于20%~80%的物种生态位处于中等地位。
15 a火烧迹地灌木群落物种Levins和Shannon Wiener生态位宽度排名前20%的是峨眉蔷薇(R. omeiensis Rolfe)、唐古特忍冬(L. tangutica Maxim.)、山梅花,两种方法计算结果均指示峨眉蔷薇生态位宽度值最大,Levins和Shannon Wiener生态位宽度分别为22.44和3.19,其后依次是唐古特忍冬、山梅花,Levins和Shannon Wiener生态位宽度值分别是19.19、17.61和3.10、3.01。生态位宽度排名后20%的物种是灰栒子(Cotoneaster acutifolius Turcz.)、尖叶栒子(C. acuminatus Lindl.)和六道木(Zabelia biflora (Turcz.) Makino),其Levins和Shannon Wiener生态位宽度值均为1.00和0.00。
未过火林地林下灌木层物种Levins和Shannon Wiener生态位宽度排名前20%的是唐古特忍冬、菰帽悬钩子、扁刺蔷薇、箭竹、山梅花、大刺茶藨子和美丽茶藨子(R. pulchellum Turcz.),两种方法计算结果均表明唐古特忍冬生态位宽度值最大,Levins和Shannon Wiener生态位宽度值分别为20.20和3.18,其后依次是菰帽悬钩子、扁刺蔷薇、箭竹、山梅花、大刺茶藨子(Ribes alpestre Wall. ex Decne.)和美丽茶藨子,Levins和Shannon Wiener生态位宽度值分别是16.01、13.54、13.39、10.91、10.25、8.38和2.90、2.78、2.96、2.57、2.42、2.31。生态位宽度排名后20%的物种是小叶忍冬、红脉忍冬(L. nervosa Maxim.)、六道木、长果茶藨子(R. stenocarpum Maxim.)、荚蒾、黑水柳和秦岭蔷薇,其中荚蒾、黑水柳和秦岭蔷薇Levins和Shannon Wiener生态位宽度值最小,均为1.00和0.00,小叶忍冬、红脉忍冬、六道木和长果茶藨子Levins和Shannon Wiener生态位宽度值分别为1.69、1.68、1.47、1.17和0.60、0.74、0.50、0.27。
对物种的出现频率进行999次随机化重排后(表2),5 a火烧迹地灌木群落有3个特化种,分别是托柄菝葜(S. discotis Warb.)、楤木(Aralia elata (Miq.) Seem.)和小叶蔷薇(R. willmottiae Hemsl.),Levins和Shannon Wiener生态位宽度指数的实际计算值分别为2.70、4.03、1.00和1.40、1.89、0.00,随机化重排999次模拟后Levins和Shannon Wiener生态位宽度指数零分布均值分别12.41、9.08、7.27和2.72、2.38、2.13。
表 2 火烧迹地灌木群落和未过火林地林下灌木层物种特化种与泛化种
Table 2. Specialized species and generalized species of shrub communities in burned and unburned areas
群落
Communities物种
Species生态位宽度指数
实际值
Observed重排模拟后生态位宽度
指数零分布均值
Mean simulated95%置信区间下限
LowCI95%置信区间上限
UppCI划分结果
SignLevins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener 5 a火烧迹地 托柄菝葜 2.70 1.40 12.41 2.72 8.73 2.48 15.53 2.91 特化种 特化种 楤木 4.03 1.89 9.08 2.38 5.72 2.04 12.59 2.69 特化种 特化种 小叶蔷薇 1.00 0.00 7.27 2.13 4.21 1.72 10.45 2.46 特化种 特化种 15 a火烧
迹地秦岭蔷薇 3.63 1.33 6.37 1.95 3.76 1.53 9.80 2.34 特化种 特化种 唐古特忍冬 19.19 3.10 13.32 2.75 9.62 2.50 17.01 2.97 泛化种 泛化种 藤山柳 2.99 1.29 9.54 2.40 6.02 2.04 13.11 2.69 特化种 特化种 未过火林地 长果茶藨子 1.17 0.27 4.08 1.49 2.09 0.91 6.76 1.99 特化种 特化种 15 a火烧迹地灌木群落有2个特化种和1个泛化种。特化种分别是秦岭蔷薇(R. tsinglingensis Pax & K. Hoffm.)和藤山柳,Levins和Shannon Wiener生态位宽度指数的实际计算值分别为3.63、2.99和1.33、1.29,随机化重排999次模拟后Levins和Shannon Wiener生态位宽度指数零分布均值分别6.37、9.54和1.95、2.40;泛化种是唐古特忍冬,Levins和Shannon Wiener生态位宽度指数的实际计算值分别为19.19和3.10,随机化重排999次模拟后Shannon Wiener和Levins生态位宽度指数零分布均值分别13.32和2.75。
未过火林地林下灌木层有1个特化种,为长果茶藨子,Levins和Shannon Wiener生态位指数的实际计算值分别为1.17和0.27,随机化重排999次模拟后Levins和Shannon Wiener生态位宽度指数零分布均值分别为4.08和1.49。
火烧迹地特化种物种数最多,5 a火烧迹地有3种灌木,5 a火烧迹地有2种灌木,未过火林地最少,仅1种。说明随着火烧迹地植被演替的正常进行,植被朝向多样化发展。
-
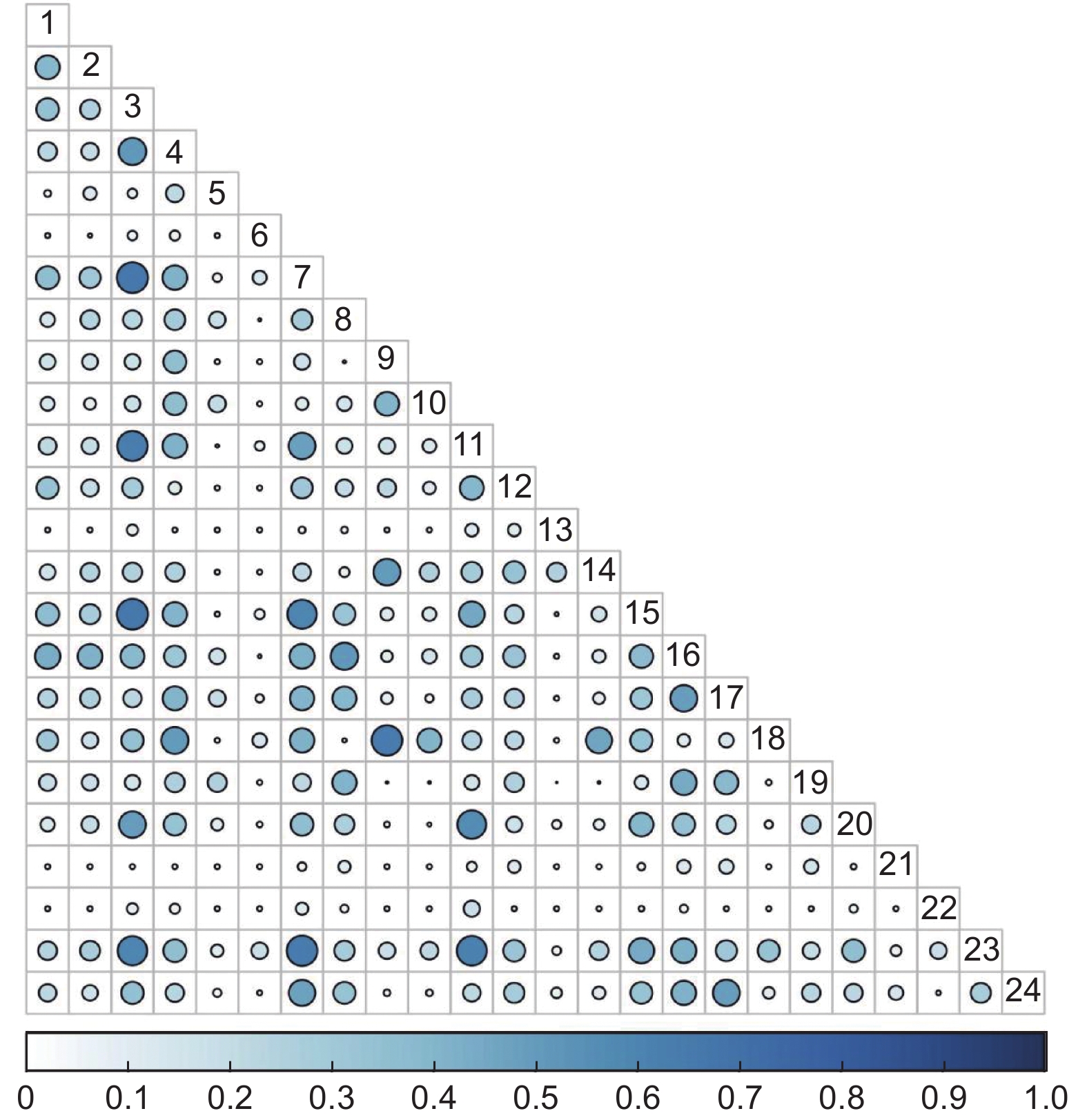
5 a火烧迹地灌木群落共有276个种对,Pianka生态位重叠指数值在0.57≤P≤1的种对有26对,占总对数的9.42%;在0.27≤P<0.57的种对有98对,占总对数的35.51%;在0≤P<0.27的种对有152对,占总对数的55.07%(图1)。
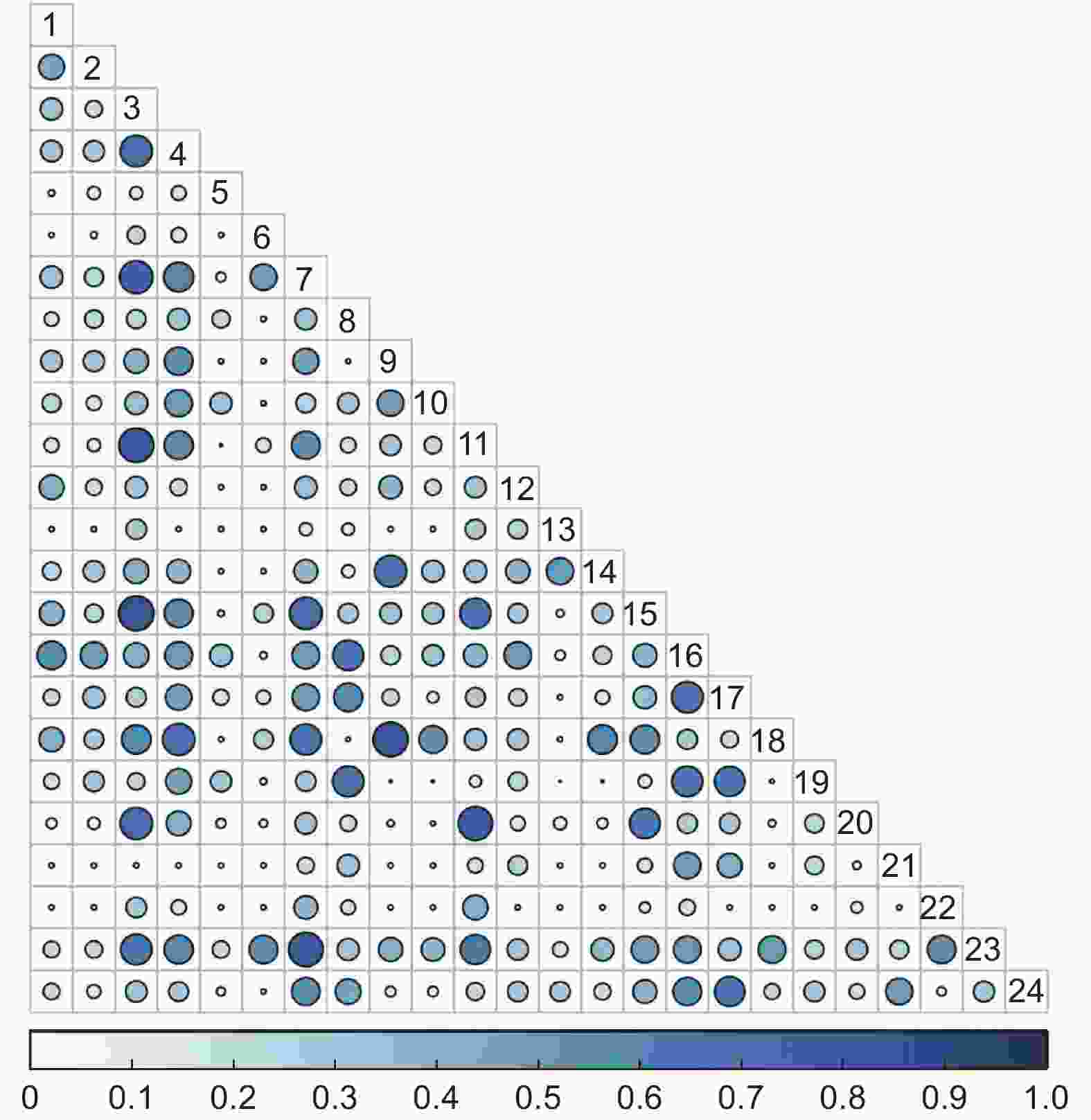
图 1 5 a火烧迹地灌木群落Pianka生态位重叠
Figure 1. Overlap of Pianka niche of shrub community in 5 a burned area
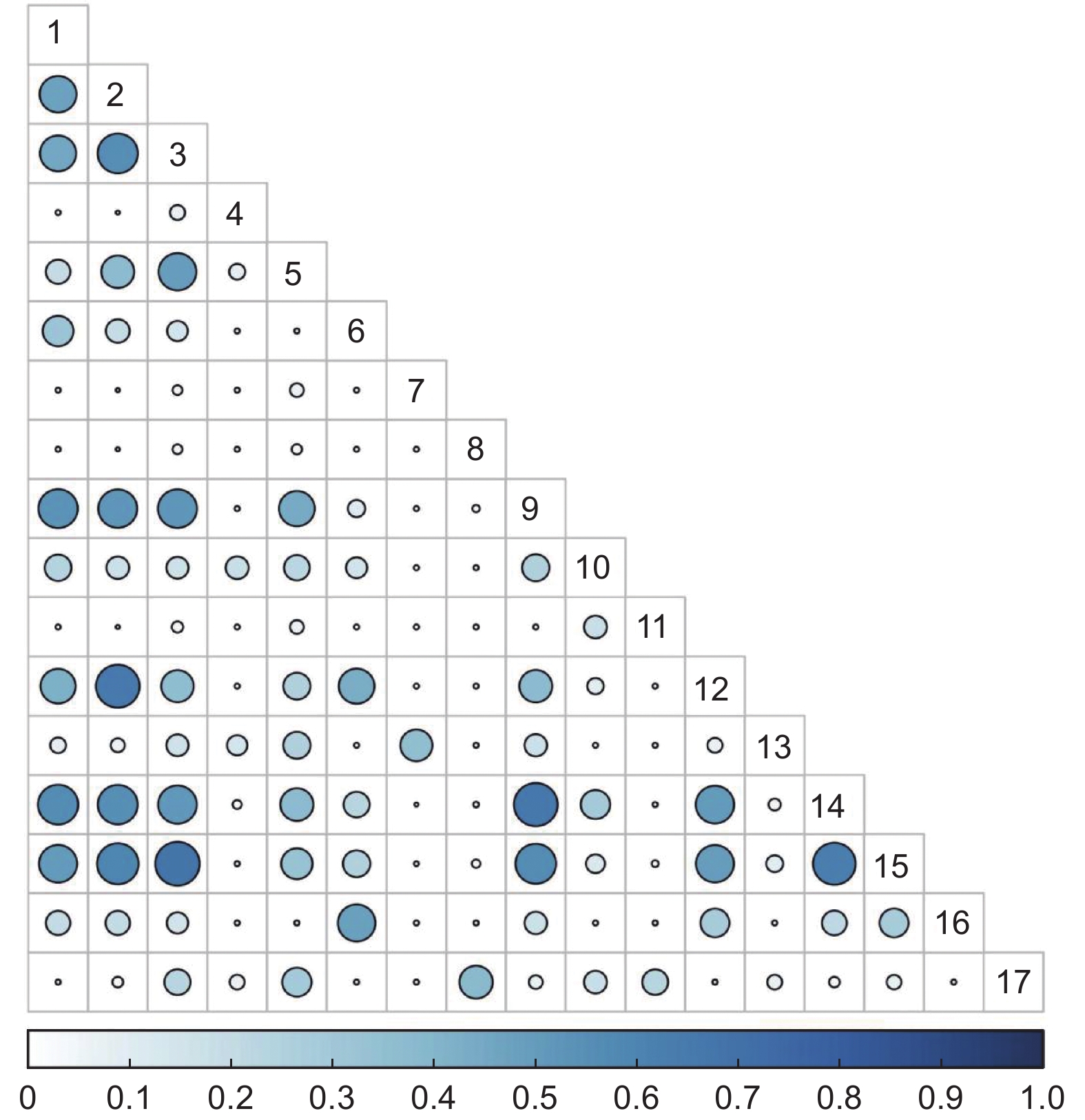
15 a火烧迹地灌木群落共有136个种对,Pianka生态位重叠指数值在0.57≤P≤1的种对有17对,占总对数的12.50%;在0.27≤P<0.57的种对有37对,占总对数的27.21%;在0≤P<0.27的种对有82对,占总对数的60.29%(图2)。
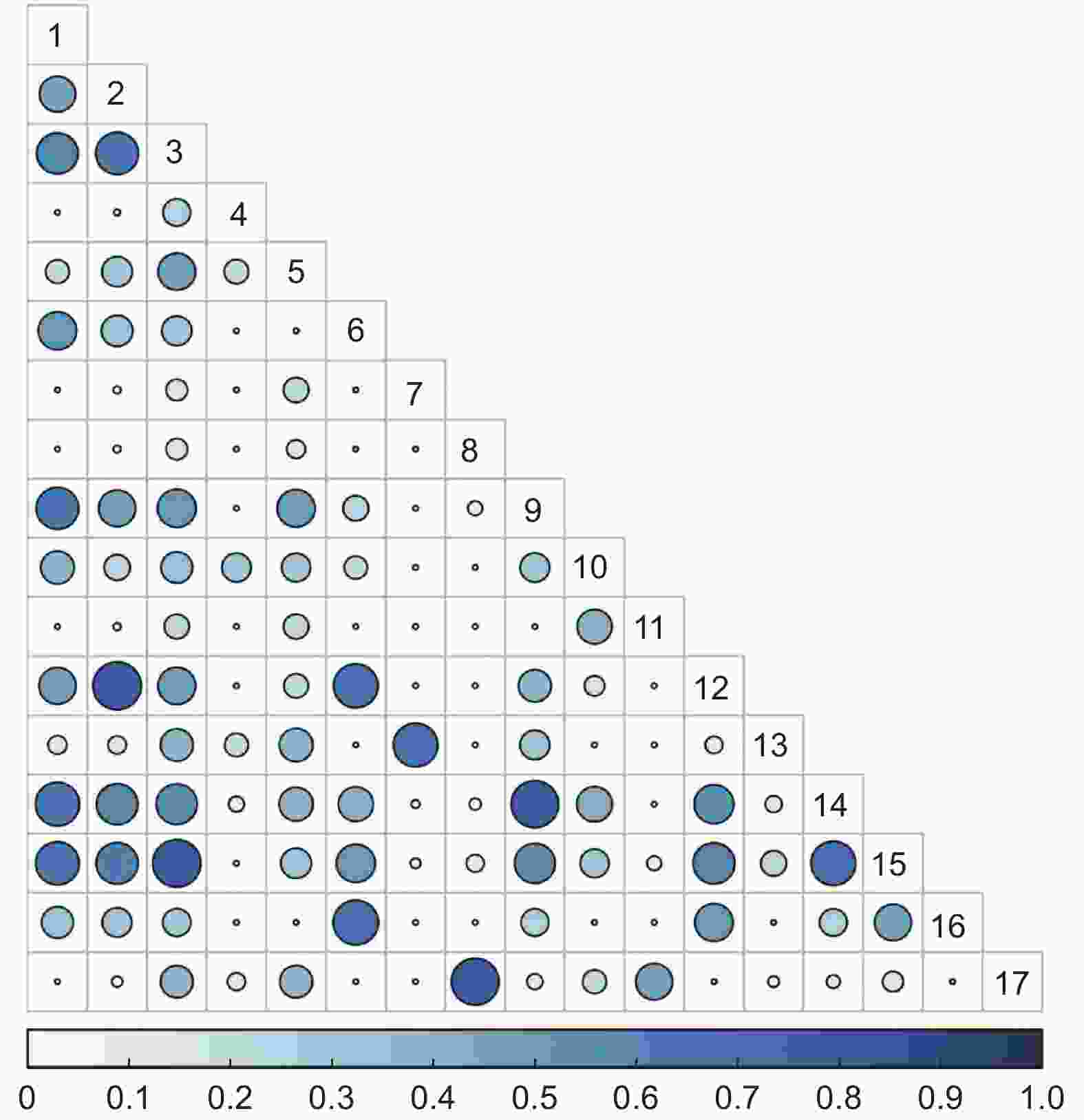
图 2 15 a火烧迹地灌木群落Pianka生态位重叠
Figure 2. Overlap of Pianka niche of shrub community in 15 a burned area
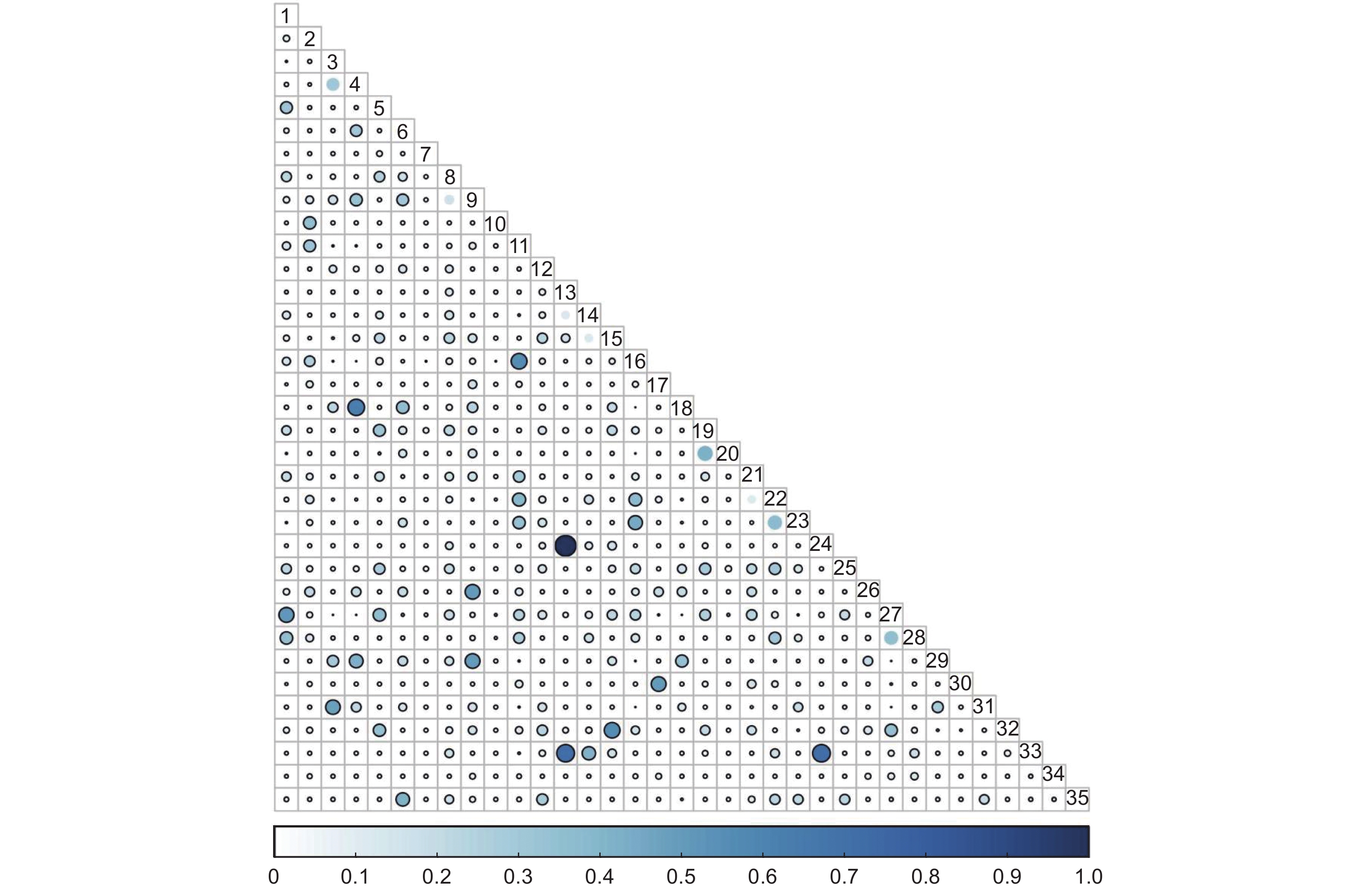
未过火林地林下灌木层共有595个种对,Pianka生态位重叠指数值在0.57≤P≤1的种对有13对,占总对数的2.19%;在0.27≤P<0.57的种对有64对,占总对数的10.76%;在0.27≤P<0.57的种对有518对,占总对数的87.06%(图3)。
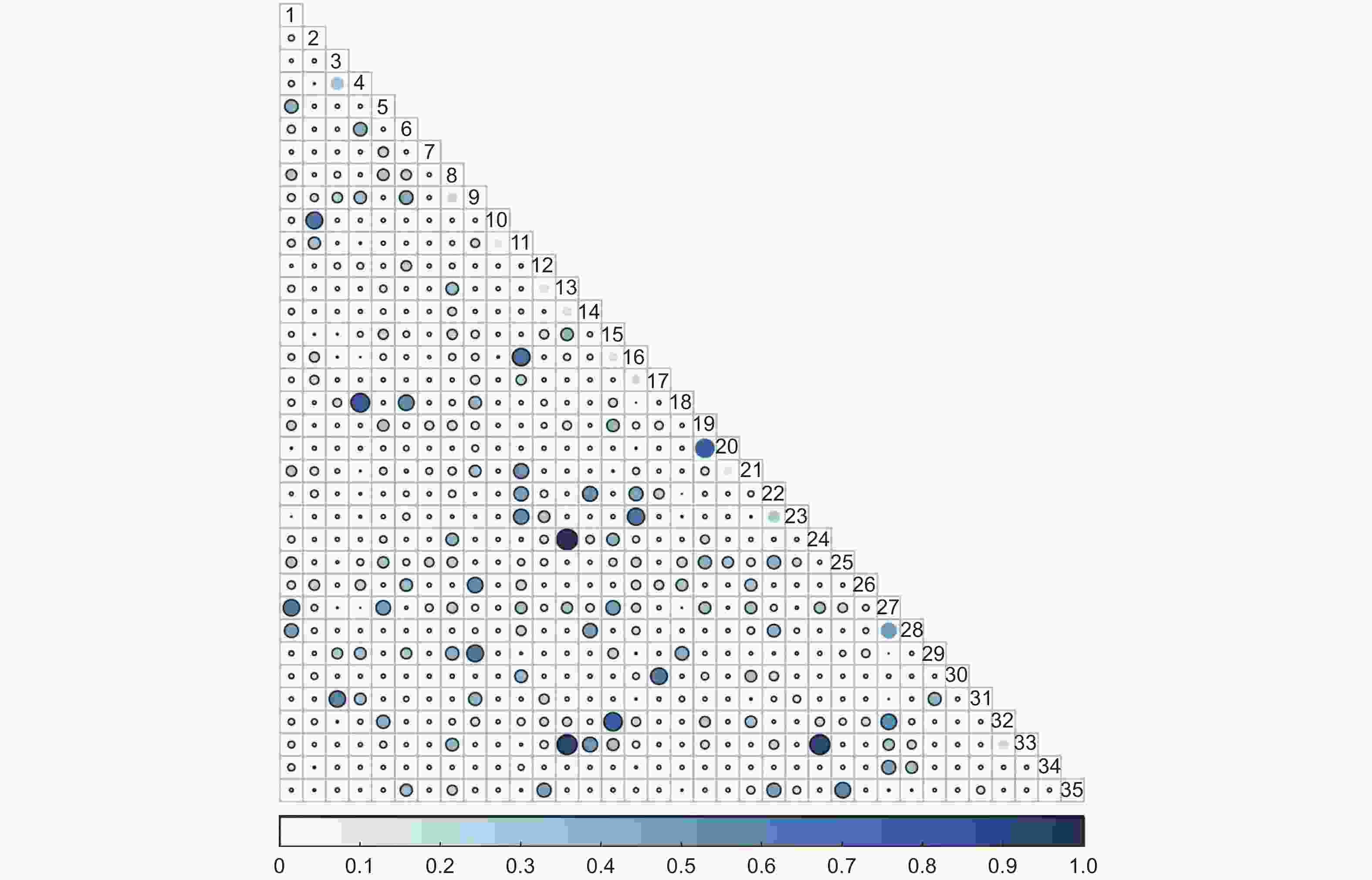
图 3 未过火林地林下灌木层Pianka生态位重叠
Figure 3. Overlap of Pianka niche of shrub community in unburned area
-
5 a火烧迹地灌木群落Schoener生态位重叠指数值在0.65≤S≤1的种对有2对,占总对数的0.73%;在0.22≤S<0.65的种对有107对,占总对数的38.77%;在0≤S<0.22的种对有167对,占总对数的60.51%(图4)。

图 4 5 a火烧迹地灌木群落Schoener生态位重叠
Figure 4. Overlap of Schoener niche of shrub community in 5 a burned area
15 a火烧迹地灌木群落Schoener生态位重叠指数值在0.65≤S≤1的种对有3对,占总对数的2.21%;在0.22≤S<0.65的种对有41对,占总对数的30.15%;在0≤S<0.22的种对有92对,占总对数的67.65%(图5)。

图 5 15 a火烧迹地灌木群落Schoener生态位重叠
Figure 5. Overlap of Schoener niche of shrub community in 15 a burned area
未过火林地林下灌木层Schoener生态位重叠指数值在0.65≤S≤1的种对有3对,占总对数的0.50%;在0.22≤S<0.65的种对有60对,占总对数的10.08%;在0≤S<0.22的种对有532对,占总对数的89.41%(图6)。
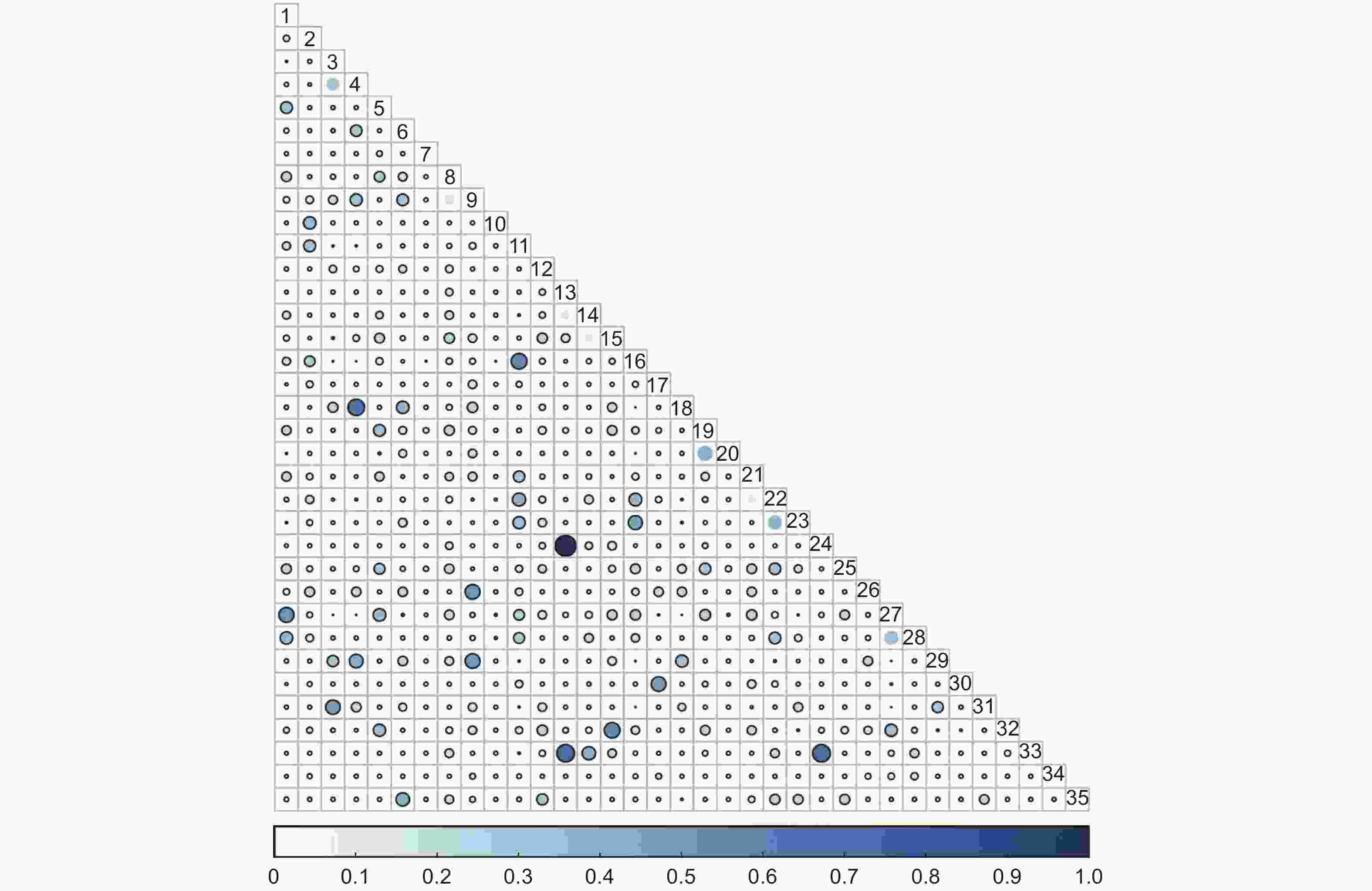
图 6 未过火林地林下灌木层Schoener生态位重叠
Figure 6. Overlap of Schoener niche of shrub community in unburned area
-
生态位宽度度量物种对各类资源环境综合利用状况,也反映了其对环境的适应能力[24]。采用Levins和Shannon Wiener生态位宽度两种方法来研究火烧迹地和未过火林地所有灌木物种生态位宽度所得到的结果基本一致,本研究同一物种在不同年限火烧迹地灌木群落和未过火林地林下灌木层的生态位宽度不一样,这与冯湘等和姚珊等的研究结果相似[25-26]。物种生态位宽度的变化与竞争、环境等因素息息相关,在不同演替阶段,随着火烧迹地的资源和环境得到进一步改善,使物种对环境资源利用和环境适应性发生改变,从而导致物种的生态位宽度改变,生态位发生分离[27-28]。
相关研究表明,一个种的生态位越宽,该物种的特化程度就越小,也就是说它更倾向于一个泛化种,泛化种的生态位宽,具有较强的竞争能力;物种的生态位越窄,该种的特化程度就越强,它更倾向于是一个特化种,特化种生态位窄,在资源竞争中处于劣势[22-23,29-31]。本研究通过对火烧迹地灌木群落和未过火林地林下灌木层中物种的出现频率进行999次随机化重排后,托柄菝葜、楤木、小叶蔷薇在5 a火烧迹地被指示为特化种;秦岭蔷薇和藤山柳在15 a火烧迹地被指示为特化种,唐古特忍冬被指示为泛化种;长果茶藨子在未过火林地被指示为特化种。相较于火烧迹地,未过火林地长果茶藨子属于特化种,生态位窄,对环境资源的竞争能力较小,容易被淘汰[12]。但在本研究的火烧迹地中,生态位窄的物种不一定生存能力和竞争能力小,如楤木树体较大,在5 a火烧迹地中分布较集中,在小生境范围内占据一定的生态位,影响和制约着其他植物的生存,很有可能今后成为火烧迹地主要优势种。个别物种也有可能由于特化严重而被淘汰,当火烧迹地植被恢复到一定阶段之后,由于各植物种群对相同资源的竞争和利用加剧,导致本身生态位较窄的物种被淘汰,如托柄菝葜在火烧迹地中数量虽多,但分布范围较窄,个体较小,只在5 a火烧迹地局部分布,很容易被淘汰。
两个物种占据相同空间和环境资源时造成种间生态位重叠,当重叠值越大时其生态学或生物学相似性可能就越高[28,32-33]。当环境资源有限,生态位重叠较大引起物种对资源性竞争,在这过程中不断会有物种的更新、淘汰、相互适应及协同进化[34-35]。本研究通过采用Pianka和Schoener生态位重叠两种方法研究火烧迹地和未过火林地所有灌木物种的生态位重叠得到的结果基本一致,种间生态位重叠严重的种对数占比为15 a火烧迹地>5 a火烧迹地>未过火林地,生态位重叠一般的种对数占比为5 a火烧迹地>15 a火烧迹地>未过火林地,生态位重叠较小的种对数占比为未过火林地>15 a火烧迹地>5 a火烧迹地;形成这种生态位格局,主要有两个原因:一是在火烧迹地初期(火烧迹地恢复至5 a),在局部范围内资源比较充足,先锋物种菰帽悬钩子、秀丽莓等迅速侵入并在火烧迹地大量繁殖占据着重要生态位,其他物种对资源的利用处于劣势,生态位重叠严重的种对相对于15 a火烧迹地占比较低;二是在小生境范围内,环境资源充足,各灌木种群对利用同一空间资源的竞争较小,群落中生态位重叠严重的种对相对于15 a火烧迹地占比较低[36-37],但目前已经演替至第5 a,灌木群落已经郁闭成林,大部分种群数量不断增加,所以处于中等生态位重叠的种对数占比最大。随着正向演替的进行(火烧迹地恢复至15 a),各灌木种群个体数量和耐阴性植物的增加,种间对相同资源利用的竞争加剧,群落中生态位重叠大的种对占比增加[38],但在火烧迹地中可能由于红桦先锋种的存在,对灌木群落各种群的空间分布产生了一定的影响,导致群落的部分物种生态位重叠中等种对数占比趋向于减少;而未过火林地林下灌木层生态位重叠严重的种对数占比最少,生态位重叠小的种对数占比最多,说明种间对相同的空间资源利用较小,这有可能是由于环境空间异质性和乔木层的共同作用,维持灌木群落物种的生态位重叠在一个较低的状态,此时群落的稳定性较高[28]。
-
Levins和Shannon Wiener生态位宽度两种方法研究青藏高原东北边缘云-冷杉林不同年限火烧迹地火烧迹地和未过火林地所有灌木物种生态位宽度所得到的结果基本一致,同一物种在火烧迹地和未过火林地的生态位宽度不一样,5 a火烧迹地特化种为托柄菝葜、楤木和小叶蔷薇;15 a火烧迹地特化种秦岭蔷薇和藤山柳,泛化种为唐古特忍冬;未过火林地特化种为长果茶藨子。
5 a、15 a火烧迹地灌木群落和未过火林地林下灌木层Pianka和Schoener生态位重叠指数分别在0.57≤P≤1和0.65≤S≤1的种对数占总种对数比例为15 a火烧迹地>5 a火烧迹地>未过火林地;在0.27≤P<0.57和0.22≤S<0.65的种对数占总种对数比例为5 a火烧迹地>15 a火烧迹地>未过火林地;在0≤P<0.27和0≤S<0.22的种对数占总种对数比例为未过火林地>15 a火烧迹地>5 a火烧迹地。由此可见,青藏高原东北边缘云-冷杉林经过重度火干扰后,不同年限火烧迹地灌木群落种间生态位重叠程度正在不断的发生着变化,而未过火林地林下灌木层种间生态位重叠则保持一个相对较低的状态,火烧迹地灌木群落正处于正向演替阶段。
青藏高原东北边缘云-冷杉林火烧迹地灌木群落种群生态位特征
Niche Characteristics of Population in Shrub Communities in Spruce-fir
-
摘要:
目的 揭示青藏高原东北边缘云-冷杉林不同年限火烧迹地灌木群落种群生态位特征。 方法 以空间代替时间方法,采用Levins和Shannon Wiener生态位宽度指数、Pianka和Schoener生态位重叠指数计算种群生态位宽度和生态位重叠程度,并划分群落特化种和泛化种。 结果 火烧迹地和未过火林地灌木物种Levins和Shannon Wiener生态位宽度指数结果基本一致,同一物种在火烧迹地和未过火林地的生态位宽度存在差异。5 a火烧迹地有3个特化种;15 a火烧迹地有2个特化种和1个泛化种;未过火林地有1个特化种。种间生态位重叠严重的种对数占比为15 a火烧迹地>5 a火烧迹地>未过火林地;生态位重叠一般的种对数占比为5 a火烧迹地>15 a火烧迹地>未过火林地;生态位重叠较小的种对数占比为未过火林地>15 a火烧迹地>5 a火烧迹地。 结论 青藏高原东北边缘云-冷杉林经过重度火干扰后,不同年限火烧迹地灌木群落种间生态位重叠程度正在不断地发生变化,而未过火林地林下灌木层种间生态位重叠程度则保持一个相对较低的状态,火烧迹地灌木群落正处于正向演替阶段。 Abstract:Objective To reveal the niche characteristics of shrub community population in the burned areas of Spruce-fir forest in different years on the northeast edge of Qinghai-Tibetan Plateau. Method Levins and Shannon Wiener niche breadth index, Pianka and Schoener niche overlap index were used to calculate the niche breadth and niche overlap degree of population, and the specialized species and generalized species of community were divided. Result The results of Levins and Shannon Wiener niche breadth index of shrub species in burned and unburned forest land were basically the same, and there were differences in the niche breadth of the same species in burned and unburned forest land. There are three specialized species in the 5-years burned area, two specialized species and 1 generalized species in the 15-years burned area and one specialized species in unburned forest land. The proportion of species logarithms with severe niche overlap among species was 15-year-burned land>5-year-burned land>unfired forest land. The species logarithm proportion of general niche overlap was 5-years-burned land>15-yearly-burned land>unfired Woodland. And the proportion of species logarithm with less niche overlap was unburned forest land > 15 years burned area > 5 years burned area. Conclusion After severe fire disturbance in the Spruce-fir forest on the northeast edge of Qinghai-Tibetan Plateau, the degree of niche overlap among species of shrub communities in burned areas of different years is constantly changing, while the degree of niche overlap among species in undergrowth of unburned forest land remains relatively low, and the community is in a positive succession stage. -
图 1 5 a火烧迹地灌木群落Pianka生态位重叠
Figure 1. Overlap of Pianka niche of shrub community in 5 a burned area
图 2 15 a火烧迹地灌木群落Pianka生态位重叠
Figure 2. Overlap of Pianka niche of shrub community in 15 a burned area
图 3 未过火林地林下灌木层Pianka生态位重叠
Figure 3. Overlap of Pianka niche of shrub community in unburned area
图 4 5 a火烧迹地灌木群落Schoener生态位重叠
Figure 4. Overlap of Schoener niche of shrub community in 5 a burned area
图 5 15 a火烧迹地灌木群落Schoener生态位重叠
Figure 5. Overlap of Schoener niche of shrub community in 15 a burned area
图 6 未过火林地林下灌木层Schoener生态位重叠
Figure 6. Overlap of Schoener niche of shrub community in unburned area
表 1 火烧迹地灌木群落和未过火林地林下灌木层物种生态位宽度
Table 1. Niche width of shrub communities in burned and unburned areas
物种 Species 5 a 15 a 未过火林地 unburned area Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener 唐古特忍冬 Lonicera tangutica 8.32 2.24 19.19 3.10 20.20 3.18 菰帽悬钩子 Rubus pileatus 12.92 2.74 — — 16.01 2.90 扁刺蔷薇 Rosa sweginzowii 7.14 2.16 11.85 2.60 13.54 2.78 箭竹 Fargesia spathacea 5.56 1.95 15.06 2.90 13.39 2.96 山梅花 Philadelphus incanus 15.01 2.94 17.61 3.01 10.91 2.57 大刺茶藨子 Ribes alpestre 11.10 2.57 11.71 2.82 10.25 2.42 美丽茶藨子 R. pulchellum 13.27 2.87 — — 8.38 2.31 天山茶藨子 R. meyeri — — — — 7.31 2.15 峨眉蔷薇 R. omeiensis — — 22.44 3.19 7.25 2.07 刺果茶藨子 R. burejense — — — — 6.81 2.16 毛花忍冬 L. trichosantha 4.00 1.39 8.43 2.34 6.78 2.26 小叶蔷薇 R. willmottiae 1.00 0.00 4.12 1.59 6.57 2.07 蓝靛果忍冬 L. caerulea 1.00 0.00 5.26 1.72 6.13 1.95 甘肃小檗 Berberis kansuensis 1.00 0.00 2.00 0.69 6.00 1.79 陕甘花楸 Sorbus koehneana — — — — 5.00 1.61 灰栒子 Cotoneaster acutifolius 2.78 1.05 1.00 0.00 4.50 1.72 川西锦鸡儿 Caragana sinica — — — — 4.24 1.55 秀丽莓 R. amabilis — — — — 3.82 1.66 刺五加 Eleutherococcus senticosus 6.41 2.22 — — 3.32 1.32 东陵绣球 Hydrangea bretschneideri 4.67 1.73 — — 3.27 1.28 葱皮忍冬 L. ferdinandi — — — — 3.17 1.30 珍珠梅 Sorbaria sorbifolia 11.78 2.59 — — 3.07 1.28 黑果菝葜 Smilax glaucochina — — — — 3.00 1.50 托柄菝葜 S. discotis 2.70 1.40 — — 2.79 1.16 楤木 Aralia elata 4.03 1.89 — — 2.00 0.87 杯腺柳 Salix cupularis — — — — 2.00 0.69 卫矛 Euonymus alatus — — — — 2.00 0.69 金银忍冬 L. maackii — — — — 1.80 0.64 小叶忍冬 L. microphylla 1.00 0.00 — — 1.69 0.60 红脉忍冬 L. nervosa 2.00 0.69 3.60 1.33 1.68 0.74 六道木 Zabelia biflora — — 1.00 0.00 1.47 0.50 长果茶藨子 R. stenocarpum — — — — 1.17 0.27 荚蒾 Viburnum dilatatum 1.60 0.56 — — 1.00 0.00 黑水柳 S. heishuiensis 10.81 2.62 10.91 2.66 1.00 0.00 秦岭蔷薇 R. tsinglingensis — — 3.63 1.33 1.00 0.00 藤山柳 Clematoclethra scandens 3.38 1.30 2.99 1.29 — — 尖叶栒子 C. acuminatus — — 1.00 0.00 — — 接骨木 Sambucus williamsii 5.40 1.83 — — — — 圆锥绣球 Hydrangea paniculata 8.50 2.33 — — — —  下载: 导出CSV
下载: 导出CSV
表 2 火烧迹地灌木群落和未过火林地林下灌木层物种特化种与泛化种
Table 2. Specialized species and generalized species of shrub communities in burned and unburned areas
群落
Communities物种
Species生态位宽度指数
实际值
Observed重排模拟后生态位宽度
指数零分布均值
Mean simulated95%置信区间下限
LowCI95%置信区间上限
UppCI划分结果
SignLevins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener Levins Shannon Wiener 5 a火烧迹地 托柄菝葜 2.70 1.40 12.41 2.72 8.73 2.48 15.53 2.91 特化种 特化种 楤木 4.03 1.89 9.08 2.38 5.72 2.04 12.59 2.69 特化种 特化种 小叶蔷薇 1.00 0.00 7.27 2.13 4.21 1.72 10.45 2.46 特化种 特化种 15 a火烧
迹地秦岭蔷薇 3.63 1.33 6.37 1.95 3.76 1.53 9.80 2.34 特化种 特化种 唐古特忍冬 19.19 3.10 13.32 2.75 9.62 2.50 17.01 2.97 泛化种 泛化种 藤山柳 2.99 1.29 9.54 2.40 6.02 2.04 13.11 2.69 特化种 特化种 未过火林地 长果茶藨子 1.17 0.27 4.08 1.49 2.09 0.91 6.76 1.99 特化种 特化种
下载: 导出CSV
-
[1] 牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17(6):579-593. [2] 张金屯. 数量生态学[M]. 第3版. 北京: 科学出版社, 2018. [3] Grinnell J. The Niche-Relationships of the California Thrasher[J]. The Auk:Ornithological Advances, 1917, 34(4): 427-433. [4] Elton C S. Animal Ecology[M]. London: Sedgwick and Jackson, 1927. [5] Holt R D. Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106: 19659-19665. doi: 10.1073/pnas.0905137106 [6] Levins R. Evolution in Changing Environments[M]. Princeton: Princeton University Press, 1968. [7] Pianka E R. The structure of lizard communities[J]. Annual Review of Ecology & Systematics, 1973, 4: 53-74. [8] Schoener T W. Some methods for calculating competition coefficients from resource-tiliation spectra[J]. The American Naturalist, 1974, 108(961): 332-340. doi: 10.1086/282911 [9] Hurlbert S H. The measurement of niche overlap and some relatives[J]. Ecology, 1978, 59(1): 67-77. doi: 10.2307/1936632 [10] 王 刚, 赵松岭, 张鹏云, 等. 关于生态位定义的探讨及生态位重叠计测公式改进的研究[J]. 生态学报, 1984, 4(2):119-127. [11] 刘建国, 马世骏. 扩展的生态位理论[M]. 北京: 科学出版社, 1990. [12] 张桂莲, 张金屯. 关帝山神尾沟优势种生态位分析[J]. 武汉植物学研究, 2002, 20(3):203-208. [13] 张林静, 岳 明, 张远东, 等. 新疆阜康绿洲荒漠过渡带主要植物种的生态位分析[J]. 生态学报, 2002, 22(6):969-973. doi: 10.3321/j.issn:1000-0933.2002.06.026 [14] Hutchinson G E. Concluding remarks[J]. Cold Spring Harbor Symposium of Quantitative Biology, 1957, 22(1507): 239. [15] 金 山, 武帅楷. 太岳山油松林火烧迹地恢复初期植物群落结构特征[J]. 生态学报, 2021, 41(10):4182-4193. [16] 金 山, 武帅楷. 太行山南段油松林火烧迹地优势草本生态位及种间关系[J]. 北京林业大学学报, 2021, 43(4):35-46. [17] 闫东锋, 郭丹丹, 杨喜田, 等. 中度火干扰对具茨山典型植被草本植物早期恢复特征的影响[J]. 华中农业大学学报, 2015, 34(5):31-36. doi: 10.13300/j.cnki.hnlkxb.2015.05.006 [18] 赵 安, 周晓雷, 田 青, 等. 迭山北坡云冷杉林火烧迹地灌木群落特征和物种多样性研究[J]. 西部林业科学, 2021, 50(5):90-100. doi: 10.16473/j.cnki.xblykx1972.2021.05.013 [19] 赵 安, 田 青, 周晓雷, 等. 迭山北坡云冷杉林火烧迹地灌木群落种间关联性[J]. 林业科学研究, 2022, 35(2):163-170. [20] 何忠伟, 胡仁传, 黄日波, 等. 广西银杉林主要树种种群生态位分析[J]. 林业科学研究, 2012, 25(6):761-766. doi: 10.3969/j.issn.1001-1498.2012.06.014 [21] 李 菁, 骆有庆, 石 娟, 等. 阿尔山地区兴安落叶松林下植物种群生态位[J]. 林业科学研究, 2011, 24(5):651-658. doi: 10.13275/j.cnki.lykxyj.2011.05.013 [22] Wu W, Logares R, Huang B, et al. Abundant and rare picoeukaryotic subcommunities present contrasting patterns in the epipelagic waters of marginal seas in the northwestern Pacific Ocean[J]. Environmental Microbiology, 2017, 19: 287-300. doi: 10.1111/1462-2920.13606 [23] Zhang J, Zhang B, Liu Y, et al. Distinct large-scale biogeographic patterns of fungal communities in bulk soil and soybean rhizosphere in China[J]. Science of the Total Environment, 2018, 644: 791-800. doi: 10.1016/j.scitotenv.2018.07.016 [24] 杨利民, 周广胜, 王国宏. 草地群落物种多样性维持机制的研究Ⅱ物种实现生态位[J]. 植物生态学报, 2001, 25(5):634-638. doi: 10.3321/j.issn:1005-264X.2001.05.020 [25] 冯 湘, 刘宝军, 刘建康, 等. 半干旱区人工封育草地植被动态研究——以宁夏盐池县为例[J]. 生态环境学报, 2020, 29(10):1961-1970. doi: 10.16258/j.cnki.1674-5906.2020.10.005 [26] 姚 珊, 郭其强, 盘金文, 等. 马尾松人工林发育过程中林下灌草群落优势种生态位动态分析[J]. 西北植物学报, 2021, 41(8):1417-1424. [27] 徐治国, 何 岩, 闫百兴, 等. 三江平原典型沼泽湿地植物种群的生态位[J]. 应用生态学报, 2007, 18(4):783-787. doi: 10.3321/j.issn:1001-9332.2007.04.013 [28] 刘加珍, 陈亚宁, 张元明. 塔里木河中游植物种群在四种环境梯度上的生态位特征[J]. 应用生态学报, 2004, 15(4):549-555. doi: 10.3321/j.issn:1001-9332.2004.04.002 [29] 吕世杰, 刘红梅, 吴艳玲, 等. 放牧对荒漠草原建群种和优势种生态位的影响[J]. 生态环境学报, 2014, 23(10):1600-1609. doi: 10.3969/j.issn.1674-5906.2014.10.006 [30] 董 雪, 李永华, 张正国, 等. 甘肃酒泉荒漠戈壁灌木群落优势物种生态位特征[J]. 中国沙漠, 2020, 40(4):138-145. [31] 马一明, 李秋华, 潘少朴, 等. 贵州高原花溪水库浮游植物优势种生态位及种间联结性动态分析[J]. 湖泊科学, 2021, 33(3):785-796. doi: 10.18307/2021.0314 [32] 夏 霆, 陈 静, 曹方意, 等. 镇江通江城市河道浮游植物优势种群生态位分析[J]. 长江流域资源与环境, 2014, 23(3):344-350. doi: 10.11870/cjlyzyyhj201403006 [33] 杨新东, 陈流保, 曾献兴, 等. 广东翁源青云山自然保护区森林群落优势树种的生态位特征[J]. 热带亚热带植物学报, 2020, 28(6):565-573. doi: 10.11926/jtsb.4214 [34] 张 峰, 上官铁梁. 翅果油树群落优势种群生态位分析[J]. 西北植物学报, 2004, 24(1):70-74. doi: 10.3321/j.issn:1000-4025.2004.01.013 [35] 赵永华, 雷瑞德, 何兴元, 等. 秦岭锐齿栎林种群生态位特征研究[J]. 应用生态学报, 2004, 15(6):913-918. doi: 10.3321/j.issn:1001-9332.2004.06.001 [36] 丁 松, 欧阳杰, 吕 丹, 等. 井冈山国家自然保护区栲树林优势种群生态位特征[J]. 中南林业科技大学学报, 2014, 34(7):99-103. doi: 10.3969/j.issn.1673-923X.2014.07.019 [37] 陈 俊, 艾训儒, 姚 兰, 等. 木林子川陕鹅耳枥 + 青冈-水马桑群落优势种生态位与种间关联[J]. 湖北民族学院学报(自然科学版), 2017, 35(3):255-260. [38] 白晓航, 张金屯. 小五台山森林群落优势种的生态位分析[J]. 应用生态学报, 2017, 28(12):3815-3826. doi: 10.13287/j.1001-9332.201712.002 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4081
- HTML全文浏览量: 2397
- PDF下载量: 101
- 被引次数: 0




