

-
栓皮栎(Quercus variabilis BI.)和蒙古栎(Q. mongolica Fisch. ex Ledeb.)属壳斗科栎属植物,是中国北方温带落叶阔叶林的主要建群树种,其形成的相应生境也成为分布区域内昆虫多样性的保藏、繁衍与发展的主要场所[1-3]。以往我国在栎林昆虫方面的研究主要集中于重要害虫,如栓皮栎波尺蛾(Larerannis filipjevi Wehrli)、栎空腔瘿蜂(Trichagalma acutissimae Monzen)、栎粉舟蛾(Fentonia ocypete Bremer)、栗山天牛(Massicus raddei Blessiq)等生物学和防治技术等研究[4-9];以及不同区域害虫种类调查[10-12];但关于栎林生态系统内昆虫群落整体组成、多样性格局及变化规律研究极少,对生物灾害形成与制约的重要机制—生物多样性维系缺乏了解与认知,影响了整体生态系统的健康管理。
昆虫群落构成及多样性是森林生态系统结构与稳定性的重要表征,也是害虫种群发生与制约的重要条件,其变化受到多种因素的共同影响。白栎(Q. alba L.)年龄变化与昆虫多样性关系研究发现,白栎中植食性昆虫的丰富度、均匀度和群落结构受其森林年龄、森林结构、植物种类相对丰度和非生物条件的影响[13];在评估不同的栎属植物生境对直翅目昆虫群落多样性的影响时,发现其生境内植被的高度均匀性使其昆虫群落结构相似,多样性差异小[14];而栎属植物的遗传多样性明显使其生境内的昆虫多样性增加[15]。群落多样性不仅受寄主的影响,还受到环境中的其他生物因子和非生物因子影响。一种麦蛾科Pseudotelphusa sp.幼虫使白栎叶片黏连形成的遮蔽空间,增加了同样在白栎叶片上取食的其它昆虫多样性[16];栎属植物上的植食性昆虫类群与气候的潜在关系研究表明,与气候(温度、降水量)相比,昆虫群落与栎属植物物种相关性更强,气候因子的影响不显著[17]。
国内对栎林昆虫群落多样性尚无针对性研究,较多研究纳入在不同区域昆虫的多样性调查,如黑里河自然保护区(蒙古栎林林缘与其他林型)的昆虫群落多样性比较[18],山西历山国家级自然保护区中不同演化阶段林型(红桦木、辽东栎等)蛾类群落多样性研究[19],抚顺市龙岗山自然保护区内的步甲科昆虫类群抽样调查等[20],均未明确栎林生境昆虫组成与生物多样性差异及影响,尤其缺乏不同区域间差异比较。
鳞翅目蛾类成虫具有趋光特性,容易采集,常被作为可以评价整个生态系统结构稳定性的指示生物,是了解不同生态区昆虫多样性动态的重要手段[21-23]。本研究采用灯诱法调查我国北方两种主要天然栎林生态系统中蛾类群落结构组成及多样性,揭示其生境与时序变化特征,为未来蛾类生物多样性基础调查合理布局,及栎林蛾类害虫监测与控制提供依据。
-
选取了3个不同栎林生境类型的样地,分别位于北京市西郊门头沟区九龙山自然保护区森林保护站的栓皮栎林A (115°59′ E, 39°54′ N)、河北省滦平县拉海岭林场的蒙古栎林B (117°29′ E, 40°85′ N)和内蒙古赤峰市克什克腾旗大局子林场的蒙古栎林C (113°37′ E, 42°40′ N)。可分别满足华北平原到内蒙高原山地过渡边缘区不同类型栎林比较(A栓皮栎林、B蒙古栎林),及处于不同生态交错区相同类型栎林比较(B平原山地交错区蒙古栎林、C林草交错区蒙古栎林)。
北京市西郊门头沟区九龙山自然保护区属典型暖温带石质山地次生落叶阔叶林生态系统的自然保护区(A),属太行山低山丘陵区,海拔在100 ~ 997 m之间;林间植被较稀疏且以阔叶林为主,少量针叶林分布,其中栓皮栎林大部分为幼龄林;该地气候条件为暖温带半湿润季风气候[24]。河北省滦平县拉海岭林场(B),属高原与山地的过渡带,海拔在396 ~ 1 132 m之间,地形地貌主要为中低山地。生境中主要植被类型有油松人工林、蒙古栎天然次生林、槲树天然次生林、侧柏天然次生林等[25],林间植被组成类型复杂多样,其中蒙古栎林的林龄以近熟林分布较广。内蒙古赤峰市克什克腾旗大局子林场(C),属内蒙古高原与大兴安岭南段山的林草交汇地带。海拔高度1 200 ~ 1 700 m,属半干旱大陆性季风气候。复杂的地貌类型,多样的生物气候条件,有着良好的生物多样性[26]。树种资源异常丰富,天然生长着几十种乔木,阔叶林和针叶林分布较广,其中蒙古栎林以近熟林为主。
-
2019年6—9月分别在3样地灯诱采集蛾类标本。诱捕点均设在距林缘0.05 km以上林内空旷地。采用500 W的高压汞灯(欧司朗GYZ250自镇流汞灯)和40 W黑光灯(GOLDVISS GS-803B)组成协同诱虫灯源。采集间隔时间为20 d。开灯时间20:00—24:00。内蒙古蒙古栎林样地8月设置了20:00—次日5:00开灯诱集试验。采集标本经制作后,以形态特征进行种类鉴定和统计[27-30]。标本均保存在中国林业科学研究院生物标本馆。
-
利用Excel对蛾类物种数及个体数分别进行数据统计并汇总,利用EstimateS[31]评估抽样数据。
多样性分析:采用Shannon-Wiener多样性指数(H)、Simpson优势度集中性指数(D)、Margalef丰富度指数(R)和Pielou均匀度指数(J),计算不同生境间的物种数和每种物种的个体数[32],公式如下:
Shannon-Wiener多样性指数(H):
$ H=-\sum _{i=1}^{s}{P}_{i}\mathrm{l}\mathrm{n}{P}_{i} $
Simpson优势度集中性指数(D):
$ D=\sum {P}_{i}^{2} $
Margalef丰富度指数(R):
$ R=\left(S-1\right)/\mathrm{l}\mathrm{n}N $
Pielou均匀度指数(J):
$ J=H/\mathrm{l}\mathrm{n}S $
其中S为物种数,
$ {N}_{i} $ 为样地第i个物种的个体数;N为样地内所有物种的个体数;$ {P}_{i} $ 为第i种个体数占总个体数N的比例,即$ {{P}}_{{i}}={{N}}_{{i}}{/}{N} $ 。不同生境之间蛾类昆虫群落相似性采用Jaccard指数计算[33]:
$ I=c/(a + b-c) $
其中a、b分别为样地A和样地B的群落物种数;c为两个样地共有的物种数。
-
由于时序、地理、空间变化等众多因素影响生物多样性,在分析比较时选取在一致截面下进行。对不同生境蛾类群落组成差异和相似性分析,分别选取3样地所有调查时段的数据累计结果比较;对不同时间尺度变化的蛾类群落组成差异比较:选取同一生境不同时段的数据进行比较。
-
研究共采集蛾类2 521只,隶属21科293种(表1)。从科水平来看,夜蛾科和尺蛾科物种数及个体数明显多于其他科,占绝对优势,均为常见类群;而部分科的蛾类种类较少,为各地区的稀有类群,虎蛾科、网蛾科和鹿蛾科均仅在样地A中出现,且均为1种;凤蛾科和蚕蛾科仅在样地B栎中出现;羽蛾科和巢蛾科仅在样地C中出现。卷蛾科、枯叶蛾科、钩蛾科、凤蛾科、羽蛾科、蚕蛾科和巢蛾科在样地A中未见;虎蛾科、网蛾科、鹿蛾科、羽蛾科和巢蛾科在样地B中未见;刺蛾科、木蠹蛾科和大蚕蛾科在样地C中未见。
表 1 不同样地栎林蛾类昆虫群落组成比较
Table 1. Comparison of moth community composition in three plots
科 Family A B C G S N G S N G S N 夜蛾科 Noctuidae 19 22 56 10 12 32 44 52 365 尺蛾科 Geometridae 13 13 40 17 19 50 42 49 226 螟蛾科 Pyralididae 16 19 41 2 2 6 16 19 70 舟蛾科 Notodontidae 7 8 17 12 14 964 8 8 21 天蛾科 Sphingidae 12 13 37 9 10 36 5 6 16 灯蛾科 Arctiidae 6 7 14 3 3 7 3 4 235 毒蛾科 Lymantriidae 2 3 11 5 6 10 4 6 15 卷蛾科 Tortricidae - - - 1 1 1 8 8 19 刺蛾科 Limacodidae 5 6 23 2 3 3 - - - 枯叶蛾科 Lasiocampidae - - - 1 1 1 4 5 123 波纹蛾科 Thyatiridae 1 2 14 1 2 4 1 2 7 大蚕蛾科 Saturniidae 2 2 3 1 1 1 - - - 钩蛾科 Drepanidae - - - 1 1 1 1 1 8 木蠹蛾科 Cossidae 1 1 1 1 2 3 - - - 虎蛾科 Agaristidae 1 1 2 - - - - - - 鹿蛾科 Ctenuchidae 1 1 29 - - - - - - 凤蛾科 Epicopeiidae - - - 1 1 1 - - - 羽蛾科 Pterophoridae - - - - - - 1 1 2 网蛾科 Thyrididae 1 1 2 - - - - - - 蚕蛾科 Bombycidae - - - 1 1 1 - - - 巢蛾科 Yponomeutidae - - - - - - 1 1 3 总计 Total 87 99 290 68 79 1 121 138 162 1 110 注:G:属数;S:种数;N:个体数;“-”指在该生境内未采集到
Note: G: Genus number; S: Species number; N: Individual number; - no found北京栓皮栎林采集鉴定14科87属99种,河北蒙古栎林16科68属79种,内蒙古蒙古栎林13科138属162种,不同样地共有种数量差异(表2):AB(24种)>BC(20种)>AC(10种)>ABC(4种),4个共有种分别是榆绿天蛾(Callambulyx tatarinovi Bremeret & Grey)、角顶尺蛾(Phthonandria emaria Bremer)、宽胫夜蛾(Protoschinia scutosa Schiffermüller)和尖锥额野螟(Loxostege verticalis Linnaeus)。
表 2 3个不同样地蛾类共有种数(对角线上)及相似性系数(对角线下)
Table 2. The number of common moth species in three plots (on the diagonal line) and similarity coefficient (under the diagonal line)
样地 Plot A B C A - 24 10 B 0.155 8 - 20 C 0.040 0 0.090 9 - 群落相似性分析结果表明(表2),3种生境的相似性系数均小于0.25,各生境间蛾类组成处于极不相似水平;北京栓皮栎林和河北蒙古栎林生境类型相似性相对较高,为0.155 8;北京栓皮栎林和内蒙古蒙古栎林的生境类型之间相似度最低,为0.04。
-
北京栓皮栎林样地内的蛾类成虫各科的物种数量及个体数随月份变化如图1所示,6月采集鉴定7科21种;7月份10科68种;8月份共采集11科29种。蛾类昆虫群落结构组成特征在不同月份中存在差异,物种数及个体数量均在7月份达到最大。
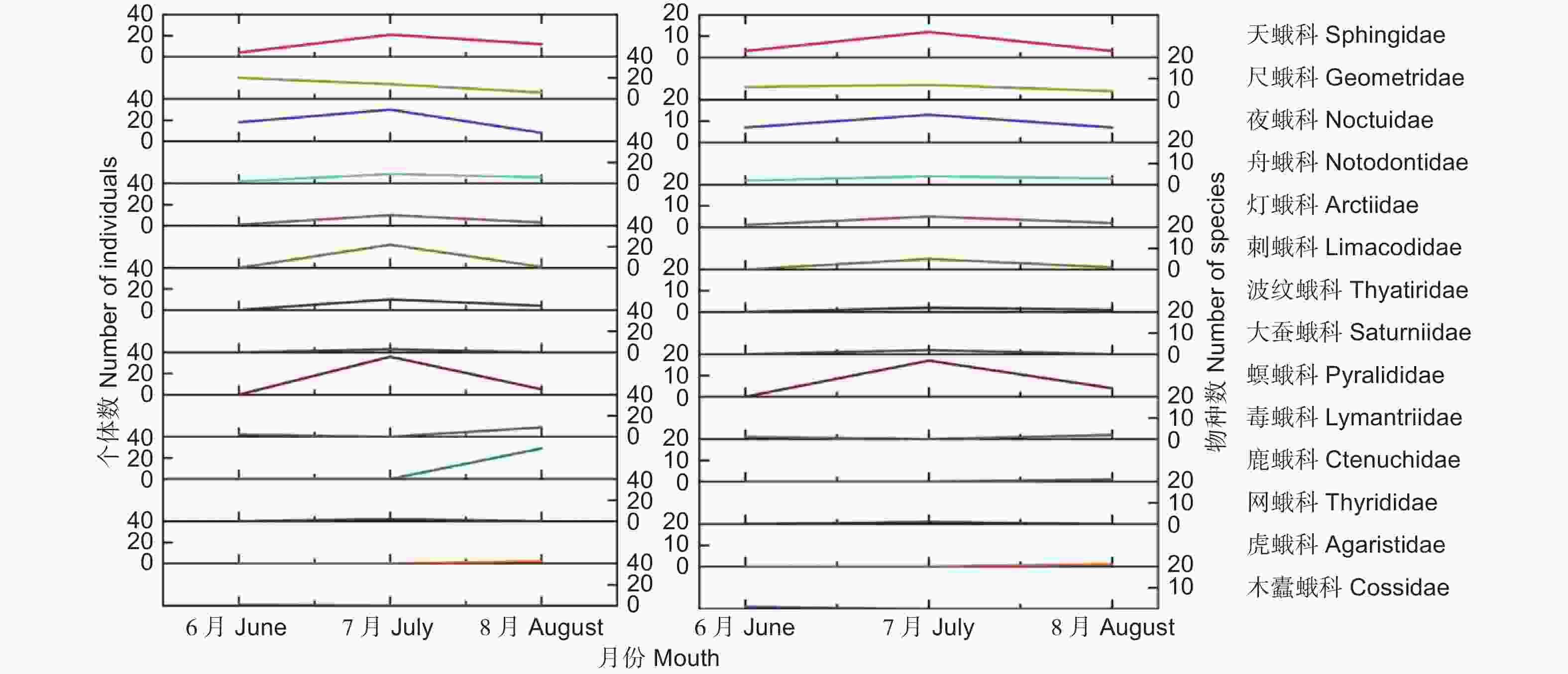
图 1 样地A不同月份蛾类各科种类与个体数量变化
Figure 1. Monthly variation of species and individual numbers in moth Families in plot A
-
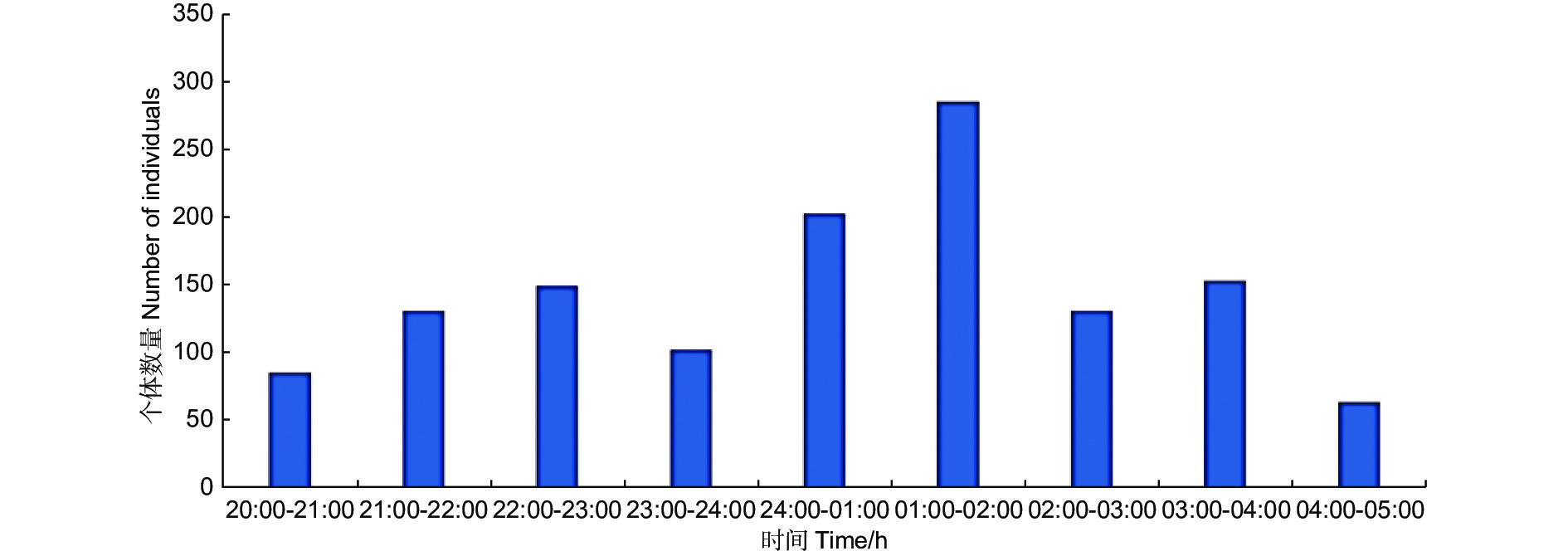
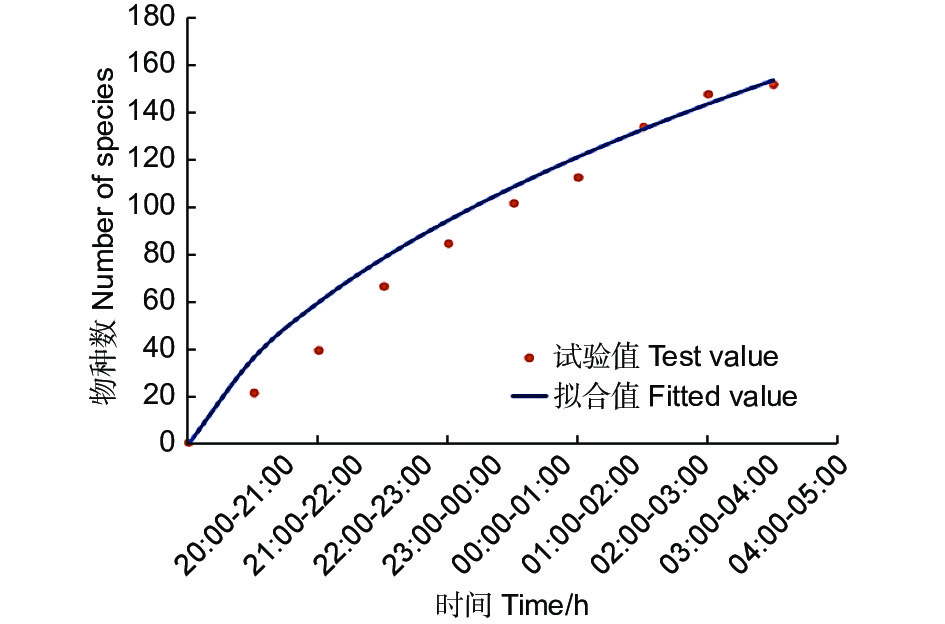
C样地按小时从8:00到次日凌晨5:00连续灯诱,诱集个体数量变化见图2,显示蛾类夜间不同时间段均可趋光飞行,但0:00—2:00间达到数量高峰。以小时计算的物种采集累积饱和曲线(图3)也表明,2:00以后的曲线趋向饱和,能够采集到新物种的几率降低。
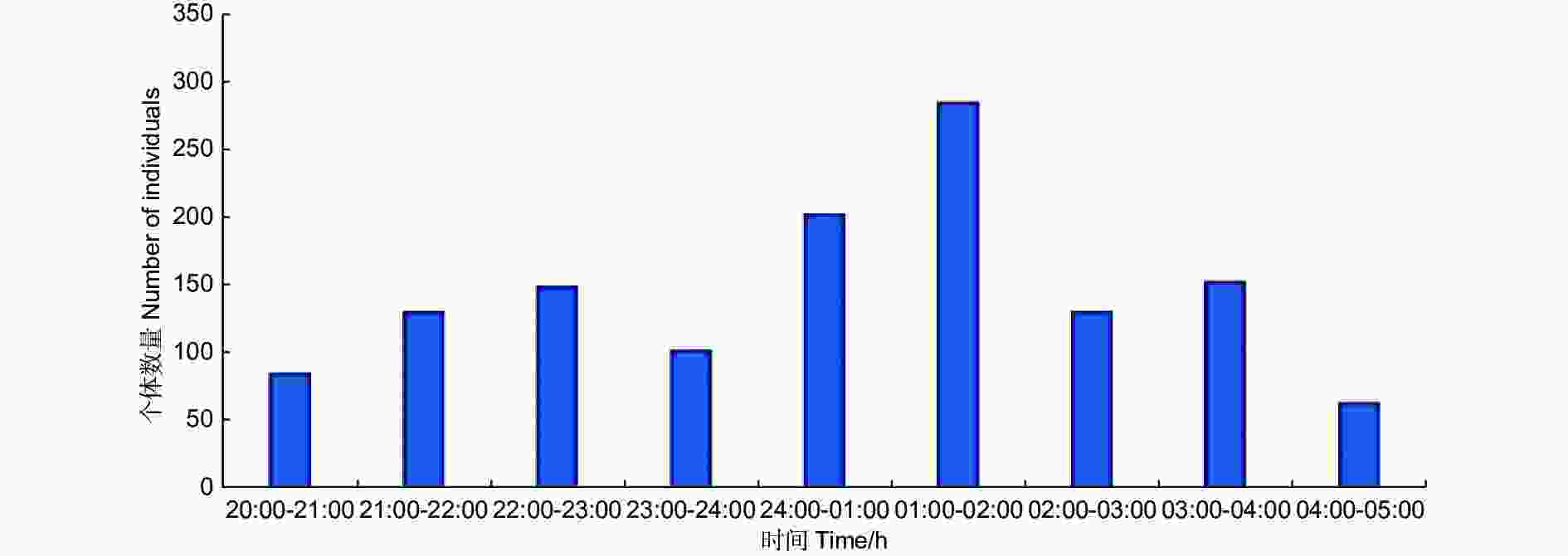
图 2 样地C不同开灯时段蛾类诱集个体数量变化
Figure 2. Hourly variation of individual numbers collected in plot C
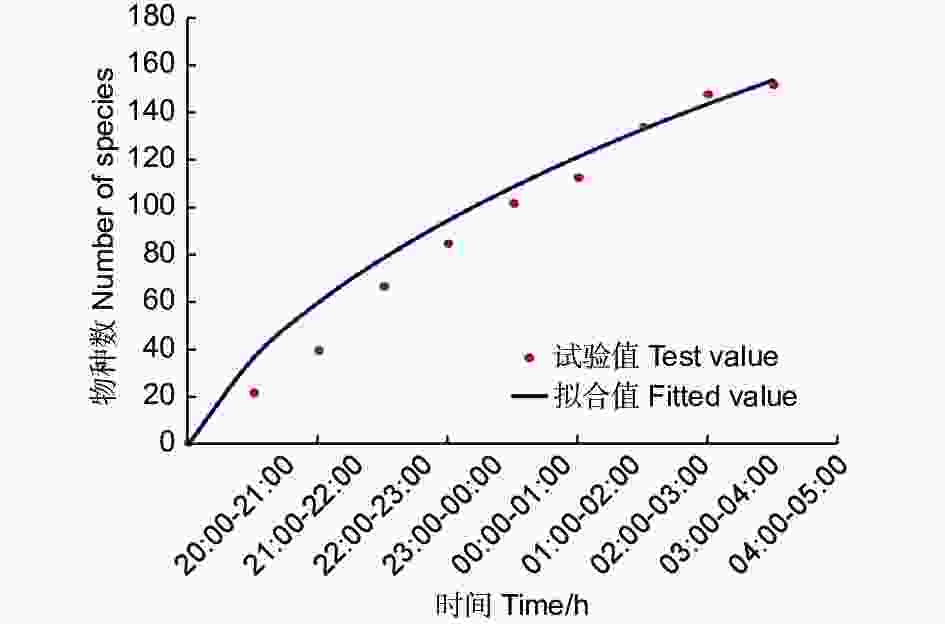
图 3 样地C内蛾类物种累积曲线
Figure 3. Cumulative curve of moth species in plot C
-
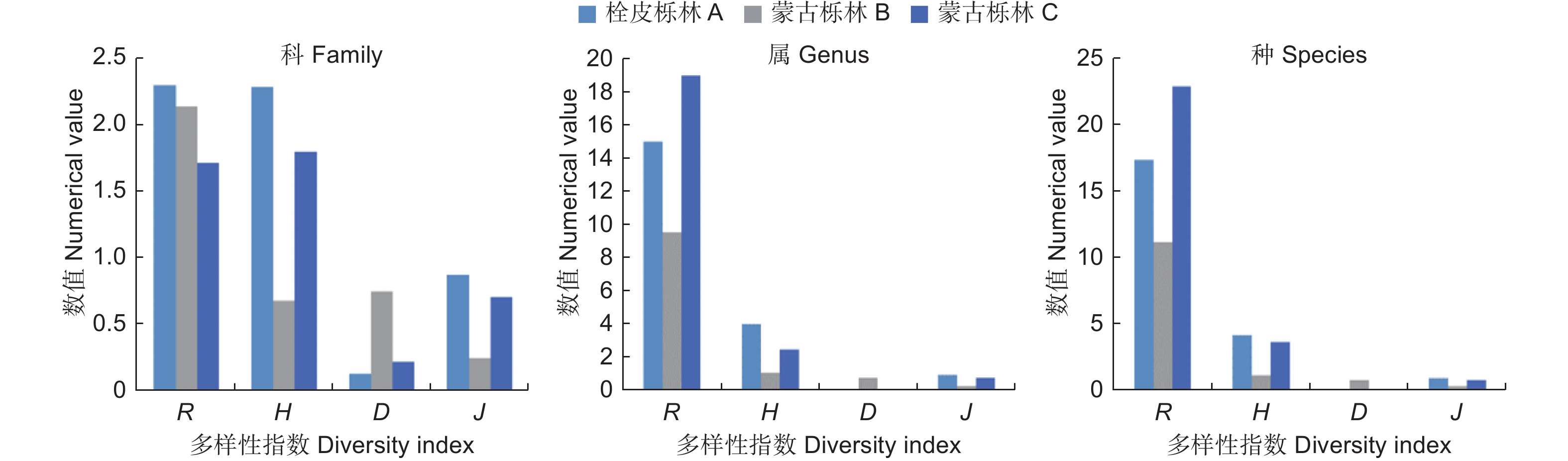
3个栎林生境内蛾类群落多样性特征分析结果表明,北京栓皮栎林和内蒙古蒙古栎林生境内的蛾类群落多样性指数均高于河北蒙古栎林生境。基于不同分类阶元的多样性分析结果表明(图4),无论在科级还是属种级水平,栓皮栎林的多样性指数和均匀度指数均高于蒙古栎林。3样地科级的Shannon多样性指数H、Simpson优势度指数D、Pielou均匀度指数J均与属、种各指数变化的一致,但Marglef丰富度指数R则显示出明显差异。在属种分析中表现最高的内蒙古蒙古栎林样地在科级丰富度指数却表现为最低,表明该区域种类较为集中在某些科。

图 4 栓皮栎林和蒙古栎林蛾类群落不同分类阶元多样性指数比较
Figure 4. Diversity index of moths in different taxonomic order in Q. variabilis and Q. mongolica forest
-
本研究对3个典型北方栎林样地内的蛾类昆虫进行了系统性调查,共获得21科293种2521头标本。3种生境中,以内蒙古蒙古栎林内的蛾类种数和个体数最高,河北蒙古栎林中最少,且蛾类类群以夜蛾科和尺蛾科为主。
蛾类群落组成相似性分析,3种栎林生境内的蛾类昆虫群落结构存在显著的差异,而生态地理区相近的北京栓皮栎林和河北蒙古栎林的相似性高于同为蒙古栎林生境的河北和内蒙古间的相似性,表现空间差异性影响明显。相关研究也指出昆虫群落组成结构、优势种组成以及个体数量变化与其生境内的植物组成类型、海拔、气候温度等综合作用均有关联[34-36]。北京栓皮栎林和河北蒙古栎林样地相较与内蒙古蒙古栎林样地的空间距离较近,且前二者均处于从平原向山地高原的过渡带,其环境气候及地被植物存在一定程度的相似性。同为蒙古栎林的B、C样地,不仅其林间混交树种及林下植被差异巨大,其空间距离和海拔也相差较远,随海拔增加的生物物种形成不适应机制[37-42],综合因素均决定了该差异的形成。
3种栎林生境内的蛾类昆虫出现具有明显的时序特征。月序变化揭示,7月达到物种个体及种类数量的高峰。这应与我国北方大部分省区7月份为温度最高及降水较多相关,长期进化的生态适应性使大多数蛾类选择在这一期间大量成虫羽化产卵,高温高湿环境有利于后代卵的孵化及幼虫迅速生长发育。夜间诱集的时间序列也存在蛾类在0:00至2:00时段个体大量集中出现,分析产生这一现象的原因一是蛾类长期形成的固有节律,在某个特定时段容易飞行活动,二是可能温湿度等环境条件在这个时刻达到了最佳,这需要今后通过精确环境记录与更多次不同时段诱集进行揭示。
3种不同栎林生境内蛾类群落的多样性指数显现趋势具有一致性。北京栓皮栎林和内蒙古蒙古栎林内的蛾类群落多样性指数和均匀度指数均比河北蒙古栎林高。不同生境内昆虫群落多样性的高低在很大程度上受到了环境气候条件、植物群落结构、人为干扰等[43-44]影响,栎粉舟蛾在河北样地的种群暴发应是造成这一结果的根本原因。
以不同分类阶元进行多样性指数分析,北京栓皮栎林生境蛾类群落在科、属级水平的多样性指数和均匀度指数,均比蒙古栎林内的蛾类群落表现更丰富多样且分布均匀。内蒙古蒙古栎林丰富度指数显示出科级与属、种级多样性特征的变化存在明显差异,显示出不同类群的进化与地理格局分化对丰富度指数的强烈影响,为今后应用于区域不同类群的分布与演化分析奠定了基础。
-
通过3个样地蛾类群落种类组成与生物多样性指数差异比较分析,栎林蛾类昆虫群落组成与多样性呈现明显的时间和空间异质性格局,环境因素直接影响蛾类生物多样性,不同阶元丰富度指标可判别类群地理分布与分化程度。
今后蛾类监测调查可作为定量指标,显示不同类型和区域栎林生物与非生物环境因素差异及健康程度评价,但应用于不同尺度的定量评价中,需要充分考虑对环境、气候因子设计梯度监测布局和调查时序周期设计,使结果准确可靠。
致谢:感谢中国林业科学研究院华北林业实验中心、河北滦平县拉海岭林场、内蒙古克什克腾旗大局子林场的调查协助;感谢中国林业科学研究院标本馆张培毅老师蛾类鉴定帮助。
北方蒙古栎和栓皮栎林蛾类多样性比较分析
Analysis of Moth Diversity in Oak Forests, Quercus variabilis and Q. mongolica in Northern China
-
摘要:
目的 调查比较我国北方两种主要天然栎林生态系统-栓皮栎和蒙古栎林蛾类群落结构组成及多样性,揭示其生境与时序变化特征,为栎林蛾类害虫监测与控制提供依据。 方法 选择北京门头沟区九龙山栓皮栎林、河北滦平县蒙古栎林和内蒙古克什克腾旗蒙古栎林3个典型样地,2019年6—9月利用灯诱法进行定期采集,比较不同样地和时间内蛾类组成与多样性差异。 结果 3个样地共获得蛾类成虫21科293种,夜蛾科和尺蛾科数量及种类最丰富;蛾类群落种类组成空间与时间差异明显;河北蒙古栎林蛾类组成结构与北京栓皮栎林相似性高于同为蒙古栎林的内蒙古样地;蛾类成虫出现期时序变化显著,种类和个体数量均在7月份达到最高。多样性结果表明北京栓皮栎林和内蒙古蒙古栎林生境内的蛾类群落Marglef丰富度指数、Shannon多样性指数和Pielou均匀度指数均高于河北蒙古栎林,群落表现稳定。其中Marglef丰富度指数在科级指数变化与属、种级指数变化表现明显差异,而其它指数变化呈一致性。 结论 栎林蛾类昆虫群落组成与多样性呈现明显的时空异质性格局,环境因素直接影响蛾类生物多样性,不同阶元丰富度指标可判别类群地理分布与分化程度。 Abstract:Objective To provide the basis for monitoring and controlling moth pests in oak forests, the structure and diversity of moth communities were investigated in two main natural oak forest ecosystems, Quercus variabilis, and Q. mongolica, in northern China for revealing the potential changes caused by temporal and habitat. Methods The composition and diversity of moths in temporal and spatial differences were compared through collecting the specimen of moth using light trap during summer from June to September 2019 in three typical plots, including Q. variabilis forest in Jiulong Mountain, Mentougou District, Beijing; Q. mongolica forest in Luanping County, Hebei and in Keshiketeng, Inner Mongolia. Result A total of 293 moth species in 21 families were obtained from the plots. The largest number of moths and the most species were concentrated in Noctuidae and Geometridae. However, the species composition of the community differed significantly in terms of space and time. The moth species composition and structural similarity in the Q. mongolica forest in Hebei were more similar with the Q. variabilis in Beijing than those in Q. mongolica forest in Inner Mongolia. In addition, the emergence period of adult moths exhibited obvious temporal changes, and the greatest number of species and individuals were found in July at each plot. Diversity results showed that the Marglef richness index, Shannon diversity index, and Pielou evenness index of moths in Beijing and Inner Mongolia plots were higher than those in Hebei plot, and the community performance was stable. Among them, the Marglef richness index showed obvious differences in the diversity characteristics of family, genus, and species, while the changes of other indexes were consistent. Conclusion The composition and diversity of moth communities in oak forests show obvious spatio-temporal heterogeneity. Environmental factors can significantly affect moth biodiversity indicators. Richness indicator differences at various taxonomic levels can be used for distinguishing the geographical distribution and degree of certain taxa. -
Key words:
- moth community
- / Quercus variabilis
- / Quercus mongolica
- / diversity index
-
图 1 样地A不同月份蛾类各科种类与个体数量变化
Figure 1. Monthly variation of species and individual numbers in moth Families in plot A
图 2 样地C不同开灯时段蛾类诱集个体数量变化
Figure 2. Hourly variation of individual numbers collected in plot C
图 4 栓皮栎林和蒙古栎林蛾类群落不同分类阶元多样性指数比较
Figure 4. Diversity index of moths in different taxonomic order in Q. variabilis and Q. mongolica forest
表 1 不同样地栎林蛾类昆虫群落组成比较
Table 1. Comparison of moth community composition in three plots
科 Family A B C G S N G S N G S N 夜蛾科 Noctuidae 19 22 56 10 12 32 44 52 365 尺蛾科 Geometridae 13 13 40 17 19 50 42 49 226 螟蛾科 Pyralididae 16 19 41 2 2 6 16 19 70 舟蛾科 Notodontidae 7 8 17 12 14 964 8 8 21 天蛾科 Sphingidae 12 13 37 9 10 36 5 6 16 灯蛾科 Arctiidae 6 7 14 3 3 7 3 4 235 毒蛾科 Lymantriidae 2 3 11 5 6 10 4 6 15 卷蛾科 Tortricidae - - - 1 1 1 8 8 19 刺蛾科 Limacodidae 5 6 23 2 3 3 - - - 枯叶蛾科 Lasiocampidae - - - 1 1 1 4 5 123 波纹蛾科 Thyatiridae 1 2 14 1 2 4 1 2 7 大蚕蛾科 Saturniidae 2 2 3 1 1 1 - - - 钩蛾科 Drepanidae - - - 1 1 1 1 1 8 木蠹蛾科 Cossidae 1 1 1 1 2 3 - - - 虎蛾科 Agaristidae 1 1 2 - - - - - - 鹿蛾科 Ctenuchidae 1 1 29 - - - - - - 凤蛾科 Epicopeiidae - - - 1 1 1 - - - 羽蛾科 Pterophoridae - - - - - - 1 1 2 网蛾科 Thyrididae 1 1 2 - - - - - - 蚕蛾科 Bombycidae - - - 1 1 1 - - - 巢蛾科 Yponomeutidae - - - - - - 1 1 3 总计 Total 87 99 290 68 79 1 121 138 162 1 110 注:G:属数;S:种数;N:个体数;“-”指在该生境内未采集到
Note: G: Genus number; S: Species number; N: Individual number; - no found 下载: 导出CSV
下载: 导出CSV
表 2 3个不同样地蛾类共有种数(对角线上)及相似性系数(对角线下)
Table 2. The number of common moth species in three plots (on the diagonal line) and similarity coefficient (under the diagonal line)
样地 Plot A B C A - 24 10 B 0.155 8 - 20 C 0.040 0 0.090 9 -
下载: 导出CSV
-
[1] Pons X, Lumbierres B, Starý P. Expansion of the aphid Myzocallis (Lineomyzocallis) walshii (Monell) on the red oak Quercus rubra, and adaptation of local parasitoids in the northeastern Iberian Peninsula (Hom., Aphididae, Calaphidinae; Hym., Braconidae, Aphidiinae)[J]. Journal of Pest Science, 2006, 79(1): 17-21. doi: 10.1007/s10340-005-0105-6 [2] Semalde A A M, Mendioro M S, Delomen M L C, et al. DNA barcoding and morphometric analyses of oriental fruit fly species Bactrocera occipitalis (Bezzi) and B. philippinensis (Drew and Hancock) (Diptera: Tephritidae) from Guimaras, Philippines[J]. Philippine Entomologist, 2013, 27(1): 58-74. [3] Yun Y H, Suh D Y, Yoo H D, et al. Yeast Associated with the Ambrosia Beetle, Platypus koryoensis, the Pest of Oak Trees in Korea[J]. Mycobiology, 2015, 43(4): 458-466. doi: 10.5941/MYCO.2015.43.4.458 [4] 姜 静, 杨忠岐, 唐艳龙, 等. 专用黑光灯对栗山天牛的诱杀技术研究[J]. 环境昆虫学报, 2010(3):369-374. doi: 10.3969/j.issn.1674-0858.2010.03.013 [5] 王小艺, 杨忠岐, 唐艳龙, 等. 白蜡吉丁肿腿蜂对栗山天牛低龄幼虫的寄生作用[J]. 昆虫学报, 2010, 53(6):675-682. doi: 10.16380/j.kcxb.2010.06.012 [6] 李方平, 孙新杰, 高 梅. 栓皮栎波尺蛾幼虫林间分布型观察研究[J]. 河南林业科技, 2010(3):42-43. doi: 10.3969/j.issn.1003-2630.2010.03.016 [7] 唐艳龙, 杨忠岐, 王小艺, 等. 释放花绒寄甲成虫和卵防治栗山天牛[J]. 林业科学, 2012, 48(7):186-191. doi: 10.11707/j.1001-7488.20120729 [8] 王景顺, 王相宏, 武三安. 栎空腔瘿蜂自然种群生命表的组建与分析[J]. 中国农学通报, 2014, 30(28):29-33. doi: 10.11924/j.issn.1000-6850.2014-0558 [9] 李秀梅, 闫立军, 李 兴. 承德市栎纷舟蛾危害现状及防治技术探讨[J]. 河北林业科技, 2020(2):55-57. doi: 10.16449/j.cnki.issn1002-3356.2020.02.016 [10] 刘玉娟, 衣岩敏, 詹雅丽, 等. 蒙古栎主要害虫调查[J]. 内蒙古林业调查设计, 2011(2):72-116. [11] 曹亮明, 魏 可, 李雪薇, 等. 我国栎类植物蛀干蛀果害虫及其天敌多样性研究进展[J]. 植物保护学报, 2019, 46(6):15-26. [12] 李雪薇, 王小艺, 曹亮明, 等. 中国典型地区栎类害虫种类调查及其危害情况研究[J]. 林业与生态科学, 2020, 35(4):387-392. [13] Jeffries J M, Marquis R J, Forkner R E. Forest age influences oak insect herbivore community structure, richness, and density[J]. Ecological Applications, 2006, 16(3): 901-912. doi: 10.1890/1051-0761(2006)016[0901:FAIOIH]2.0.CO;2 [14] Moyano L, Cardenas A M, Gallardo P, et al. Short-term effects of a revegetation program on the Orthopteran diversity in oak forests of the Southern Iberian Peninsula[J]. Journal of Insect Science, 2014, 14(1): 290. [15] Bastien C, L Lélia, Brice G, et al. Genetic diversity increases insect herbivory on oak saplings[J]. PLoS ONE, 2012, 7(8): e44247. doi: 10.1371/journal.pone.0044247 [16] Lill J T, Marquis R J. Ecosystem engineering by caterpillars increases insect herbivore diversity on white Oak[J]. Ecology, 2003, 84(3): 682-690. doi: 10.1890/0012-9658(2003)084[0682:EEBCII]2.0.CO;2 [17] Leckey E H, Smith D M. Nufio, C R, et al. Oak-insect herbivore interactions along a temperature and precipitation gradient[J]. Acta Oecologica, 2014, 61: 1-8. doi: 10.1016/j.actao.2014.08.001 [18] 郭 成, 袁树先, 刘 斐, 等. 黑里河国家级自然保护区昆虫多样性研究[J]. 吉林农业大学学报, 2012, 34(1):31-36. doi: 10.13327/j.jjlau.2012.01.003 [19] 侯沁文, 白海艳, 铁 军, 等. 山西历山国家级自然保护区不同演化阶段林型蛾类多样性[J]. 应用昆虫学报, 2014(5):1310-1319. doi: 10.7679/j.issn.2095-1353.2014.05.021 [20] 张旭臣, 刘佩旋, 辛 蓓, 等. 抚顺市龙岗山自然保护区步甲科昆虫种类的调查初报[J]. 辽宁农业科学, 2017(5):6-10. doi: 10.3969/j.issn.1002-1728.2017.05.002 [21] Usher M B, Keiller S. The macrolepidoptera of farm woodlands: Determinants of diversity and community structure[J]. Biodiversity and Conservation, 1998, 7(6): 725-748. doi: 10.1023/A:1008836302193 [22] Ricketts T H, Daily G C, Ehrlich P R. Does butterfly diversity predict moth diversity? Testing a popular indicator taxon at local scales[J]. Biological Conservation, 2001, 103(3): 361-370. [23] Summerville K S, Ritter L M, Crist T O. Forest moth taxa as indicators of lepidopteran richness and habitat disturbance: a preliminary assessment[J]. Biological Conservation, 2004, 116(1): 9-18. doi: 10.1016/S0006-3207(03)00168-X [24] 胡淑萍, 刘鹏举, 高开通, 等. 北京九龙山自然保护区植物群落物种多样性分析[J]. 水土保持研究, 2013, 20(4):6. [25] 李建忠, 王 洋. 燕山山地造林树种筛选试验初报[J]. 山东林业科技, 2020, 50(2):78-80. doi: 10.3969/j.issn.1002-2724.2020.02.015 [26] 巧 云, 包玉海, 金来全, 等. 内蒙古克什克腾旗镇郊林场森林空间分布研究[J]. 内蒙古师范大学学报(自然科学汉文版, 2012, 41(3):293-296. [27] 陈一心. 中国经济昆虫志: 夜蛾科[M]. 北京: 科学出版社, 1985. [28] 李喜升, 李树英, 秦 利, 等, 中国柞树害虫原色图鉴[M]. 沈阳: 辽宁科学技术出版社, 2016. . [29] 中国科学院动物研究所. 中国蛾类图鉴[M]. 北京: 科学出版社, 1919. [30] 中国科学院中国动物志委员会. 中国经济昆虫志[M]. 北京: 科学出版社, 1980. [31] 李 巧. 物种累积曲线及其应用[J]. 应用昆虫学报, 2011, 48(6):7. [32] 李志刚, 张碧胜, 翟 欣, 等. 广州不同生境类型区域昆虫多样性[J]. 生态学杂志, 2010, 29(2):357-362. doi: 10.13292/j.1000-4890.2010.0012 [33] 刘 棋, 成文章, 拜如霞, 等. 云南普洱紫花三叉白及昆虫群落多样性研究[J]. 环境昆虫学报, 2018, 40(5):1036-1050. [34] Hawes J, Motta, C D, Overal W L, et al. Diversity and composition of Amazonian moths in primary, secondary and plantation forests[J]. Journal of Tropical Ecology, 2009, 25: 281-300. doi: 10.1017/S0266467409006038 [35] 葛 洋, 郭 苗, 曹玉言, 等. 长江中下游菜子湖湿地不同生境昆虫群落多样性[J]. 生态学杂志, 2014, 33(8):2084-2090. doi: 10.13292/j.1000-4890.2014.0190 [36] 吴龙飞, 姜文虎, 刘 洋, 等. 不同林分类型对昆虫群落多样性的影响[J]. 生态学报, 2017, 37(7):2217-2224. [37] 刘生冬, 孟庆繁, 高文韬. 长白山自然保护区北坡蛾类的多样性[J]. 东北林业大学学报, 2007(10):51-53. doi: 10.3969/j.issn.1000-5382.2007.10.019 [38] 李德贤, 施金铎, 李昊杰, 等. 陕西化龙山国家级自然保护区鳞翅目昆虫多样性与垂直分布研究[J]. 西北林学院学报, 2022, 37(1):177-182. doi: 10.3969/j.issn.1001-7461.2022.01.26 [39] 顾 伟, 马 玲, 丁新华, 等. 扎龙湿地不同生境的昆虫多样性[J]. 应用生态学报, 2011, 22(9):2405-2412. doi: 10.13287/j.1001-9332.2011.0350 [40] 何云川, 杨贵军, 王新谱. 鸣翠湖湿地昆虫群落功能团组成及其多样性[J]. 生态学杂志, 2018, 37(10):2968-2975. doi: 10.13292/j.1000-4890.201810.022 [41] 徐华潮, 郝晓东, 黄俊浩. 浙江凤阳山昆虫物种多样性[J]. 浙江农林大学学报, 2011, 28(1):1-6. doi: 10.3969/j.issn.2095-0756.2011.01.001 [42] 韩争伟, 马 玲, 曹传旺, 等. 太湖湿地昆虫群落结构及多样性[J]. 生态学报, 2013, 33(14):4387-4397. [43] Zhou X, Zou Y, Bi S, et al. Seasonal dynamics of arthropod community diversity, evenness, and relative stability in peach orchards[J]. Chinese Journal of Applied Ecology, 2004, 15(8): 1427-1430. [44] 朱 慧, 彭媛媛, 王德利. 植物对昆虫多样性的影响[J]. 生态学杂志, 2008, 27(12):2215-2221. doi: 10.13292/j.1000-4890.2008.0033 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3605
- HTML全文浏览量: 2573
- PDF下载量: 69
- 被引次数: 0








