



-
毛竹(Phyllostachys edulis (Carr.) H. de Lehaie)属于禾本科(Gramineae)竹亚科(Bambusoideae)刚竹属(Phyllostachys Sieb. Et Zucc.),是一种多年生木本散生竹,广泛分布在46° N以南地区,是我国种植面积最广的竹种之一,具有重要的经济、生态和文化价值[1-2]。毛竹开花后产生典型颖果类果实,薄而干的膜质果皮与种皮紧密贴合,在种子生物学和苗木培育上毛竹颖果通常被称为种子[3]。毛竹种子萌发是实生苗造林的基础,通过毛竹种子萌发进行实生苗造林能降低运输成本,便于移栽,有利于提高造林整齐度、成活率和遗传多样性,延长竹林高产时间,且实生苗竹林萌笋多、竹鞭生长较快、竹材利用率更高[4-5],因此,开展毛竹种子萌发研究对于竹类资源的培育和应用等具有重要意义。
随着气候变化的加剧,干旱和由土壤初级或次级盐碱化造成的盐胁迫已经成为全球性问题,并对种子萌发、植物生长等造成严重影响[6-7]。干旱和盐胁迫抑制了毛竹种子的萌发,进而影响了毛竹实生苗的生长,限制了通过毛竹种子萌发进行实生苗培育和应用[8]。目前,研究者逐步开展了干旱或盐胁迫对毛竹种子萌发率、种子活力、种子寿命和萌发后幼苗生长等生理方面的研究[9-12],但相关的分子调控机制鲜有报道。
MicroRNA(miRNA)是一类长度约为21~24 nt的小分子非编码RNA,能够通过互补配对抑制或降解靶基因进行转录后调控[13]。miRNA广泛参与了植物的生长发育[13-15],在种子萌发[16-18]和非生物胁迫响应[19]中具有重要的调控作用。在竹类植物中,研究者已经在组学水平开展了关于毛竹叶片[20]、花发育[21]、雄蕊发育[22]、茎秆快速生长[23]和竹笋发育[24]等的miRNA研究,但毛竹种子中miRNA的表达及其在干旱和盐胁迫下萌发时的调控仍是未知的,因此,本研究通过small RNA测序对毛竹种子露白阶段的miRNA进行系统分析,并探究其在不同PEG或NaCl胁迫下萌发时的表达模式,挖掘毛竹种子萌发阶段调控干旱或盐胁迫抗性潜在的关键miRNA,以期为后续的研究提供基础和参考。
-
试验所用毛竹种子于2020年9月收集自广西壮族自治区桂林市灵川县,剥除外稃后挑选大小均一、颗粒饱满、色泽明亮的完整种子备用(图1a)。种子的千粒质量为21.85 g ± 0.04 g,含水量为12.05% ± 0.35%。种子表面消毒后在培养皿中使用纸床法进行萌发[11]。聚乙二醇(PEG6000)是一种亲水性大分子物质,是模拟植物干旱胁迫的常用试剂之一,本研究使用PEG6000模拟干旱胁迫环境,同时使用氯化钠(NaCl)模拟盐胁迫。使用完全随机实验设计,将种子分为5组(A~E):A组为对照组,B组、C组模拟干旱胁迫,D组、E组模拟盐胁迫。参考前期预实验的结果,分别在培养皿中加入H2O(A)、10% PEG(B)、15% PEG(C)、50 mmol·L−1 NaCl(D)和100 mmol·L−1 NaCl(E),每组萌发200粒种子,并重复3次。每天观察并记录种子萌发状态,在第4天选择生长状态一致且均到达种皮破裂阶段的种子(图1b)。对于每个处理,在3个重复中各取5粒种子并将15粒种子混合取样后用于建库。取样后使用液氮速冻并置于−80 ℃保存。
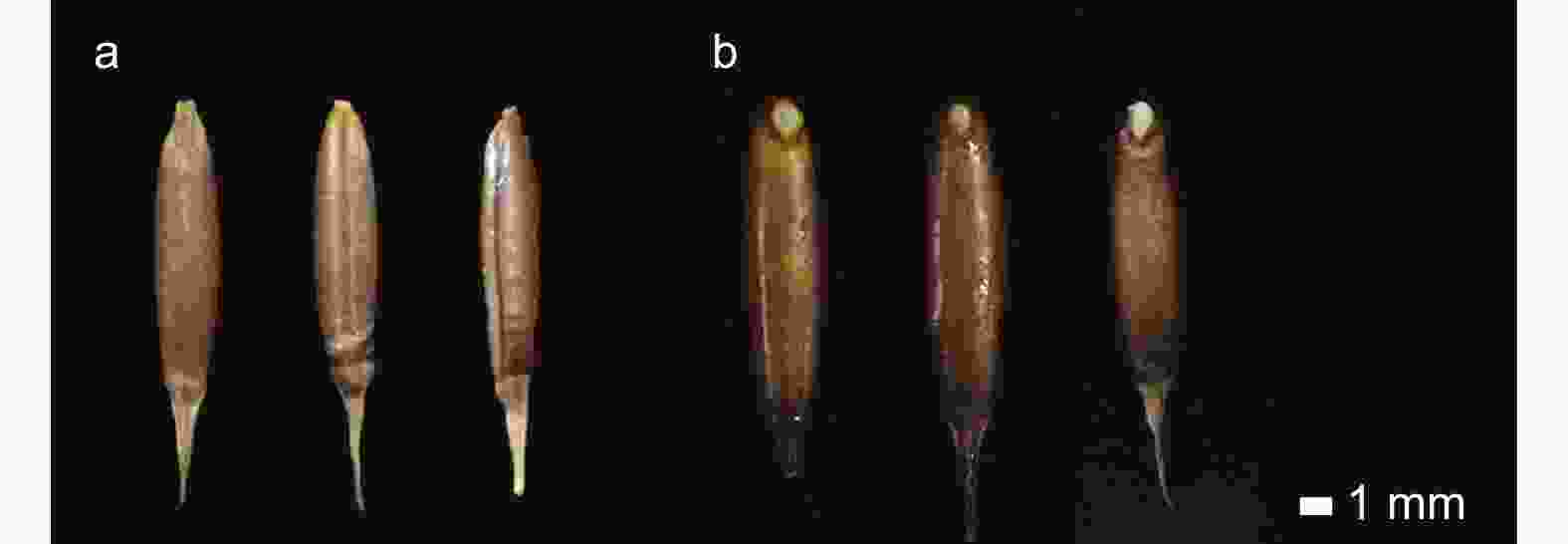
图 1 毛竹干种子(a)及种皮破裂阶段取样的萌发种子(b)
Figure 1. Dry seeds (a) and sampled seeds germinated at seed coat rupture stage (b) of Moso bamboo (Phyllostachys edulis)
-
使用植物多糖多酚RNA提取试剂盒(DP441,TIANGEN)提取样品总RNA。在建库时首先根据small RNA的结构特点将总RNA中small RNA加接头并进行纯化,随后进行反转录、PCR扩增和聚丙烯酰胺凝胶电泳,切胶回收目的条带即得到small RNA的cDNA文库。文库有效浓度合格后进行测序,使用Illumina高通量测序平台NovaSeq 6000进行单端测序,测序长度为50 bp。对获得的原始数据首先进行过滤,去除接头序列、低质量序列、含N比例>10% 的序列、5’接头污染的序列、没有3’接头序列和插入片段的序列及含连续的/T/G/C的序列;以及长度异常的序列。
-
首先将过滤后的数据比对到miRbase数据库(V22)鉴定已知miRNA[25];将未比对至miRBase的数据比对至Rfam数据库中的ncRNA 序列(rRNA、tRNA、snRNA、snoRNA) 和参考基因组重复序列[26];随后对于上述均未比对中的clean reads,使用mirEvo和miRdeep2进行新miRNA预测[27-28]。对于已知和新的miRNA序列,将其前体序列比对至miRBase数据库进行microRNA家族的鉴定[25]。
-
为了探究毛竹种子萌发露白阶段miRNA的表达,首先统计各样本中已知和新miRNA的readscount,进行TPM(Transcripts per million)归一化转化(TPM = readscount × 107/library size)。随后使用edgeR软件进行差异表达分析,以鉴定露白阶段毛竹种子响应干旱或盐胁迫的miRNA[29],差异表达miRNA筛选条件为|log2foldchange|≥1且p<0.05。
-
为了探究毛竹萌发种子中miRNA的潜在功能,对本研究中已知和新miRNA的靶基因进行预测,并进行靶基因的KEGG和GO功能富集;同时通过对差异表达miRNA靶基因的功能富集,进一步分析毛竹萌发种子中miRNA响应干旱或盐胁迫的潜在通路或途径。使用软件TargetFinder和psRNAtarget进行miRNA靶基因的预测,并对预测结果取交集[30-31]。使用OmicShare在线平台((
https://www.omicshare.com/tools ))进行靶基因KEGG和GO功能富集,富集显著性阈值为p<0.05[32-33]。 -
总RNA提取方法同1.2,反转录使用miRcute增强型miRNA cDNA第一链合成试剂盒(KR211,TIANGEN),荧光定量检测使用增强型荧光定量试剂盒(FP411,TIANGEN)。荧光定量正向引物见表1,反向引物使用试剂盒通用引物,内参引物为U6[34]。荧光定量检测仪器为qTOWER3.0 (Analytik jene),每个基因进行3次生物学重复和3次技术重复,采用2−△△CT方法计算相对表达水平[35]。
表 1 差异表达miRNA荧光定量验证引物序列
Table 1. Primers used for quantitative real time PCR of Differential expressed miRNA
miRNA编号
miRNA ID引物方向
Primer direction引物序列
Sequence (5′-3′)phe-miR3630-3p F UGUGGGAAUCUCUCUGAUGCUU phe-miR159a.1 F UUUGGAUUGAAGGGAGCUCUG phe-miR168a-5p F UCGCUUGGUGCAGAUCGGGAC phe-miR171e-5p F UGUUGGCUCGGCUCACUCAGA phe-miR396e-5p F UCCACAGGCUUUCUUGAACUG phe-miR894 F CGUUUCACGUCGGGUUCACC phe-miR6478 F CCGACCUUAGCUCAGUUGGUG novel_14 F CUGAGAGGUAGAGAACGGGAUG novel_311 F UCGUGUGCUGAACUUUAUUGA novel_5 F AUCUCAGCUGUUGAUUCCAUGAUC u6 F GGACATCCGATAAAATTGGAACGATACAG -
small RNA测序结果表明:每个文库产出数据约0.9~1.8 GB,过滤后每个样本中分别有12 984 751条(A,H2O)、14 982 423条(B,10% PEG)、13 401 719条(C,15% PEG)、13 995 450条(D,50 mmol·L−1NaCl)和11 452 796条(E,100 mmol·L−1NaCl)Clean Reads(表2)。Clean Reads的Q20值约为98%,Q30值均≥93%,与毛竹参考基因组的比对率≥88.42%,表明测序数据质量较好。具体数据产出及比对情况见表2。
表 2 small RNA数据产出信息
Table 2. Statistics of sequence data in small RNA libraries
样品名称
Sample原始读序
Raw ReadsQ20值
Q20/%Q30值
Q30/%GC含量
GC Content/%过滤后读序数
Clean Reads比对sRNA数量
Mapped sRNAA 29 379 983 98 94 54 12 984 751 11 504 336 (88.60%) B 37 101 403 98 93 54 14 982 423 13 326 206 (88.95%) C 32 426 286 98 94 54 13 401 719 11 915 649 (88.91%) D 33 041 222 98 94 54 13 995 450 12 374 157 (88.42%) E 19 605 421 98 93 53 11 452 796 10 198 110 (89.04%) -
对small RNA数据库中miRNA进行比对和预测,结果显示:每个样品中有0.08%~0.11%的序列被注释为已知miRNA,0.10%~0.11%的序列被预测为novel miRNA。对miRNA长度进行统计发现本研究成熟miRNA长度主要分布在21 nt和24 nt(图2)。本研究共鉴定miRNA成熟体序列508条,前体序列845条,其中,已知miRNA成熟体序列数量为246,前体序列数量为574,鉴定出novel miRNA成熟体数量共有262条,前体序列数量为271。
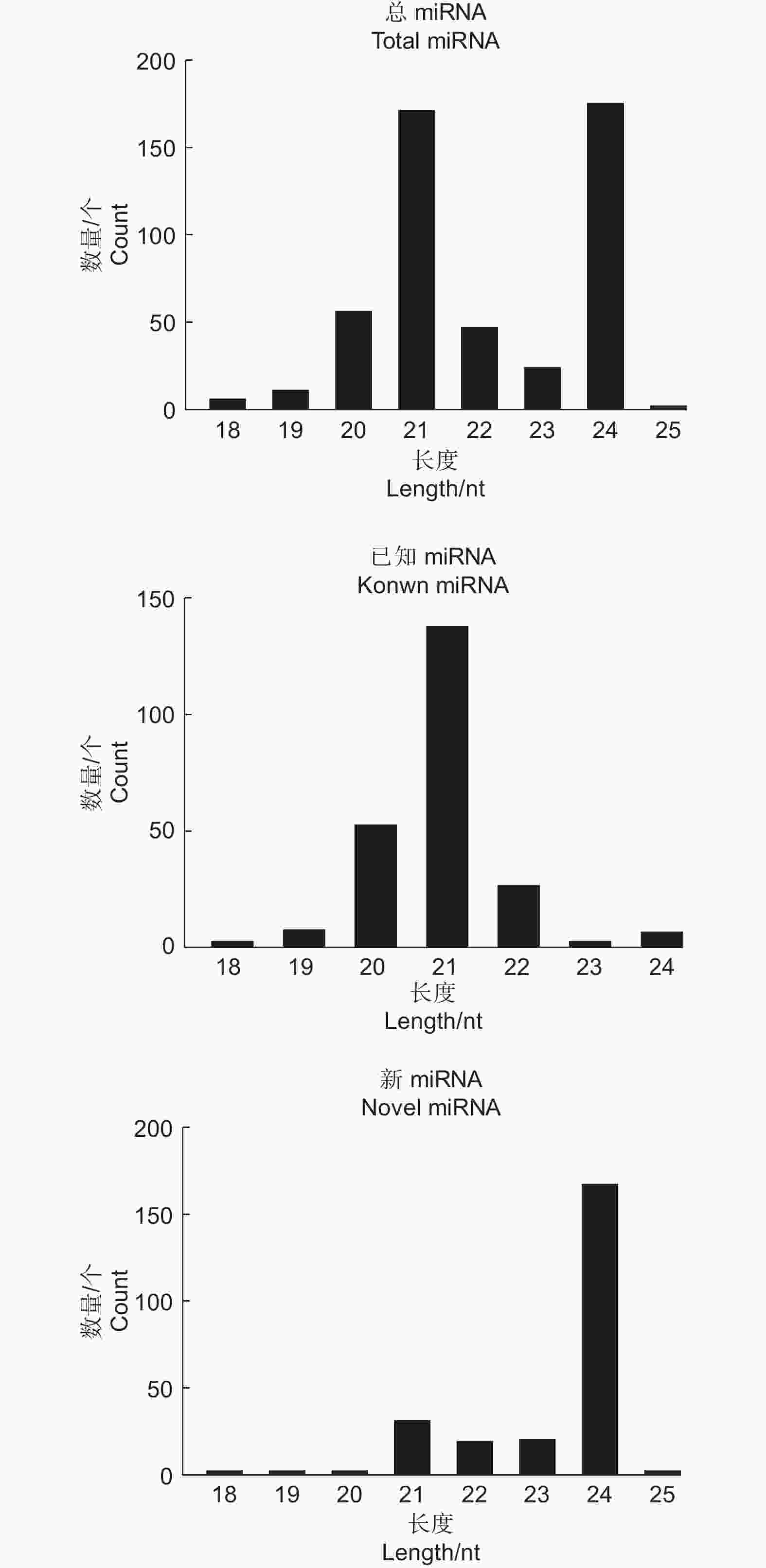
图 2 已知和新预测miRNA长度统计
Figure 2. Length distribution of known and novel miRNA
对miRNA前体序列进行家族分析,共鉴定到包含514条前体序列的45个家族,包括MIR159、MIR166、MIR156、MIR408、MIR399、MIR530等,其中,有10条新miRNA前体序列也被鉴定属于已知家族,如novel_57属于MIR408,novel_14和novel_66属于MIR169_2。统计每个家族包含的前体成员数目表明,本研究毛竹萌发种子中最大的miRNA家族为MIR159,包含54个成员,其次为MIR166(48)、MIR156(46)、MIR167_1(37)、MIR396(32)等(详细数据未列出)。
-
毛竹萌发种子中表达量前5的已知miRNA分别为phe-miR166a-3p、phe-miR159a.1、phe-miR319a-3p.2-3p、phe-miR6478、phe-miR156a-5p,表达量前5的新miRNA分别为novel_1、novel_199、novel_311、novel_241、novel_2,其中,novel_1在5个样本所有miRNA中表达量均为最高。表达量前10的新miRNA折叠最小自由能值为−187.0~−13.8 KJ·mol−1(表3、4)。
表 3 Small RNA文库中预测出前10位的保守miRNA及其序列表达量及同源性
Table 3. Sequences abundance and homologues of top ten predicted conserved miRNA in five libraries
同源序列名称
Name序列
Sequence表达量 (TPM Counts) A B C D E phe-miR166a-3p UCGGACCAGGCUUCAUUCCCC 74 864 57 593 64 907 73 677 66 116 phe-miR159a.1 UUUGGAUUGAAGGGAGCUCUG 54 736 42 178 41 405 45 018 31 525 phr-miR319a-3p.2-3p UUGGACUGAAGGGUGCUCCC 27 992 25 959 28 454 25 962 27 126 phe-miR6478 CCGACCUUAGCUCAGUUGGUG 25 651 48 135 24 454 21 842 28 992 phe-miR156a-5p UGACAGAAGAGAGUGAGCAC 17 694 16 099 15 274 23 739 28 492 phe-miR167d-5p UGAAGCUGCCAGCAUGAUCUG 17 288 28 696 19 426 18 345 16 262 phe-miR396e-5p CGUUUCACGUCGGGUUCACC 12 919 15 012 12 456 10 017 18 662 phe-miR894 UCCACAGGCUUUCUUGAACUG 11 796 17 105 14 627 10 106 5 332 phe-miR166k-3p UCGGACCAGGCUUCAAUCCCU 8 925 6 641 5 866 11 706 14 463 phe-miR319a UUGGACUGAAGGGAGCUCCCU 8 707 6 761 6 437 7 528 7 898 表 4 Small RNA文库中预测出前10位新miRNA序列及其表达量
Table 4. Sequences abundance and homologues of top ten predicted novel miRNA in five libraries
名称
Name序列
Sequence表达量 (TPM counts) 最小折叠自由能
Hairpin mfe/(KJ·mol−1)A B C D E novel_1 CUUCACUCUUCAGACUUUGAG 174 786 172 858 173 542 135 291 189 416 −46.6 novel_199 CGAUUAGUCGGCAAGUCGGGCGACU 17 413 23 705 16 760 12 951 15 796 −44.7 novel_311 UCGUGUGCUGAACUUUAUUGA 15 135 31 513 36 072 22 702 16 496 −13.4 novel_241 UUACCGGUCUUUAAACUUUUUAU 14 792 17 990 26 054 17 486 11 764 −175.9 novel_2 CCGGUCUAUAGAUUUUCAAAGUG 8 332 10 464 14 170 13 544 9 664 −115.7 novel_3 UCUCGGAUCAGGCUUCGUUCC 7 084 5 594 5 180 5 424 10 664 −39.4 novel_287 AUCGCGAUCUUGACAACCUUGACU 5 586 6 600 8 761 9 543 7 165 −27.5 novel_5 AUCUCAGCUGUUGAUUCCAUGAUC 4 993 3 381 4 723 6 698 2 299 −32.8 novel_9 CCUUCGAUCGUAUGAUGUGUCACG 4 494 4 306 4 457 4 742 2 799 −98.6 novel_87 UGUAGGAUCUCUGAUGACGGU 3 776 5 031 5 371 3 082 6 765 −187.0 对每个样本中表达水平前10的已知miRNA和新miRNA的进行TPM值统计,结果表明:排名前10的已知miRNA在每个样本总miRNA中的读序丰度(total miRNA reads)占45%~51.8%,排名前10的新miRNA在每个样本总miRNA中的读序丰度中占23.1%~29.5%。PEG和NaCl胁迫下这些miRNA在毛竹露白阶段种子表达水平均较高,推测其在毛竹种子萌发的调控中可能具有保守的重要作用。
-
通过差异表达分析,本研究共鉴定181个差异miRNA,其中,有84个上调表达,97个下调表达(图3 a)。与对照组(A)相比,10% PEG(B)、15% PEG(C)、50 mmol·L−1 NaCl(D)、100 mmol·L−1 NaCl(E)4种胁迫下分别有20、26、41和24个差异表达miRNA(DEmiRNA),此外在B-C和D-E比较组分别有16和54个DEmiRNA(图3 b)。

图 3 差异表达miRNA统计
Figure 3. Statistic of DEmiRNAs
依据miRNA的差异表达情况和表达水平,对本研究具有高丰度且显著差异表达的miRNA进行聚焦。与对照组相比,10% PEG、15% PEG、50 mmol·L−1 NaCl、100 mmol·L−1 NaCl中表达水平最高的DEmiRNA分别为novel_14、novel_311、novel_14、phe-miR159a.1,表达水平最高的已知DEmiRNA分别为phe-miR171e-5p、phe-miR3630-3p、phe-miR171e-5p和phe-miR159a.1,这些miRNA在毛竹种子萌发露白阶段中大量积累,同时在PEG或NaCl胁迫下差异表达,可能参与了种子萌发露白阶段miRNA对干旱或盐胁迫的调控。
-
通过Targetfinder预测,共获得505个miRNA的31 729个靶基因,psRNAtarget预测结果中501个miRNA可以靶向22 215个基因。对2个软件的预测结果取交集后共有17 666对miRNA-靶基因关系,包含489个miRNA 和8 812个靶基因。预测的miRNA-靶基因关系中,novel_153的靶基因数目最多,有188个靶基因,其次是phe-miR396b(167个)、phe-miR396e-5p(161个)、phe-miR396h(141个)、phe-miR164b(138个)。对同一miRNA家族成员的靶基因数目进行统计,靶基因数目前十的家族中MIR396家族靶基因数目最多。此外,根据预测结果8 812个靶基因中每个基因能够受到1~28个miRNA靶向,表明在毛竹种子萌发过程存在复杂的miRNA调控网络。具有miRNA-gene关系最多的基因中,排名前20的基因有7个属于GAMYB(Gibberellin MYB)家族,12个属于SPL(Squamosa promoter-binding-like)家族,推测GAMYB和SPL基因家族在毛竹萌发种子miRNA调控网络中可能具有重要意义。
-
根据靶基因预测结果,A-B、A-C、A-D、A-E、B-C、D-E 6个比较组差异表达miRNA分别有609、548、1 011、812、460和1 866个靶基因。GO富集结果显示:6个组合差异表达miRNA的靶基因分别显著富集在520(A-B)、516(A-C)、620(A-D)、446(A-E)、483(B-C)、610(D-E)个条目中,总计1 989个涉及细胞成分、生物学过程和分子功能的条目,推测miRNA可以通过这些靶基因广泛地参与不同的生物学过程或分子功能等调控。表5为不同比较组GO富集前10的生物学过程条目,与对照组相比,10% PEG(B)、15% PEG(C)、50 mmol·L−1 NaCl(D)、100 mmol·L−1 NaCl(E)胁迫下DEmiRNA靶基因富集最显著的生物学过程分别为跨膜转运(GO:0055085)、DNA复制正向调控(GO:0045740)、发育过程(GO:0032502)、海藻糖生物合成(GO:0005992),推测这些生物学过程对于miRNA调控PEG和NaCl胁迫下的种子萌发具有重要作用。
表 5 不同比较组差异表达miRNA靶基因GO显著富集性前10条目
Table 5. Top 10 significantly enriched GO terms for DEmiRNA target genes in different comparision groups
比较组
Comparison GroupsGO编号
GO IDGO条目
GO Term基因数量
Gene NumberP值
P-ValueA-B GO:0055085 跨膜转运Transmembrane transport 233 2.49E-11 A-B GO:0098656 阴离子跨膜转运Anion transmembrane transport 49 1.12E-07 A-B GO:0006820 阴离子转运 Anion transport 75 1.48E-07 A-B GO:1903825 有机酸跨膜转运 Organic acid transmembrane transport 41 1.83E-07 A-B GO:1905039 羧酸跨膜转运 Carboxylic acid transmembrane transport 41 1.83E-07 A-B GO:0003333 氨基酸跨膜转运 Amino acid transmembrane transport 38 2.49E-07 A-B GO:0006865 氨基酸转运 Amino acid transport 38 6.65E-07 A-B GO:0006811 离子转运 Ion transport 169 6.90E-07 A-B GO:0006885 pH调节 Regulation of pH 25 1.14E-06 A-B GO:0055067 单价无机阳离子稳态 Monovalent inorganic cation homeostasis 26 1.21E-06 A-C GO:0045740 DNA复制的正调控 Positive regulation of DNA replication 7 4.04E-07 A-C GO:0034644 细胞对紫外线的反应 Cellular response to UV 6 1.23E-06 A-C GO:0019985 跨损伤合成 Translesion synthesis 7 1.40E-06 A-C GO:0000731 参与DNA修复的DNA合成 DNA synthesis involved in DNA repair 8 1.57E-06 A-C GO:0051052 DNA代谢过程的调节 Regulation of DNA metabolic process 14 1.99E-06 A-C GO:0072593 活性氧代谢过程 Reactive oxygen species metabolic process 36 2.97E-06 A-C GO:0055074 钙离子稳态 Calcium ion homeostasis 5 3.06E-06 A-C GO:0071482 细胞对光刺激的反应 Cellular response to light stimulus 13 3.24E-06 A-C GO:0006979 对氧化应激的反应 Response to oxidative stress 41 5.55E-06 A-C GO:0051054 DNA代谢的正调控 Positive regulation of DNA metabolic process 7 6.17E-06 A-D GO:0032502 发育过程 Developmental process 288 9.35E-09 A-D GO:0080022 初生根发育 Primary root development 11 6.87E-08 A-D GO:0044767 单一有机体发育过程 Single-organism developmental process 266 1.59E-06 A-D GO:0016567 蛋白质泛素化 Protein ubiquitination 104 2.24E-06 A-D GO:0009657 质体组织 Plastid organization 46 2.80E-06 A-D GO:0070647 通过小蛋白结合或去除来修饰蛋白质
Protein modification by small protein conjugation or removal128 3.51E-06 A-D GO:0046271 苯丙烷代谢生物过程 Phenylpropanoid catabolic process 21 6.65E-06 A-D GO:0046274 木质素分解代谢过程 Lignin catabolic process 21 6.65E-06 A-E GO:0005992 海藻糖生物合成过程 Trehalose biosynthetic process 19 1.87E-06 A-E GO:0071478 细胞对辐射的响应 Cellular response to radiation 22 2.64E-06 A-E GO:0071214 细胞对非生物胁迫的响应 Cellular response to abiotic stimulus 22 4.18E-06 A-E GO:0044767 单一有机体发育过程 Single-organism developmental process 214 8.24E-06 A-E GO:0032502 发育过程 Developmental process 220 9.83E-06 B-C GO:0055085 跨膜转运 Transmembrane transport 230 1.21E-14 B-C GO:0006811 离子转运 Ion transport 173 1.54E-10 B-C GO:0006820 阴离子转运 Anion transport 78 2.58E-10 B-C GO:1903825 有机酸跨膜转运 Organic acid transmembrane transport 42 4.73E-09 B-C GO:1905039 羧酸跨膜转运 Carboxylic acid transmembrane transport 42 4.73E-09 B-C GO:0003333 氨基酸跨膜转运 Amino acid transmembrane transport 39 7.16E-09 B-C GO:0006865 氨基酸转运 Amino acid transport 39 2.11E-08 B-C GO:0006810 转运 Transport 424 2.33E-08 B-C GO:0051234 定位的建成 Establishment of localization 427 5.56E-08 B-C GO:0051179 定位 Localization 436 9.61E-08 D-E GO:0080022 初生根发育 Primary root development 12 1.52E-09 D-E GO:0006642 甘油三酯动员 Triglyceride mobilization 7 7.18E-06 D-E GO:0009658 叶绿体组织 Chloroplast organization 35 7.27E-06 D-E GO:0048506 分生组织相变时间调控 Regulation of timing of meristematic phase transition 9 8.33E-06 KEGG富集结果(表6)表明:苯丙烷生物合成途径在A-B、A-C、A-E 3个组合中被显著富集,推测苯丙烷生物合成途径对于毛竹种子萌发露白阶段miRNA响应PEG和NaCl胁迫可能具有重要意义,但毛竹种子萌发过程miRNA与苯丙烷合成途径关键基因的靶向关系及其是否共同调控干旱或盐胁迫下的种子萌发需要进一步的研究和验证。此外,果糖和甘露糖代谢、玉米素生物合成途径仅在盐胁迫下(A-D和A-E)显著富集,油菜素内酯生物合成和脂肪酸降解途径仅在A-B中显著富集,表明在毛竹种子露白阶段miRNA响应10% PEG、15% PEG、50 mmol·L−1 NaCl、100 mmol·L−1 NaCl胁迫的调控通路也存在差异。
表 6 差异表达miRNA靶基因KEGG显著富集结果
Table 6. Significantly enriched KEGG pathways for DEmiRNA target genes in different comparison groups
比较组
Comparision GroupsKEGG途径
KEGG Pathway途径ID
Pathway ID基因数量
Gene NumberP值
P-ValueA-B 糖基磷脂酰肌醇(GPI)-锚定生物合成
Glycosylphosphatidylinositol(GPI)-anchor biosynthesisko00563 10 0.000 709 3 脂肪酸延伸 Fatty acid elongation ko00062 18 0.001 981 2 2-羰基甲酸代谢 2-Oxocarboxylic acid metabolism ko01210 22 0.002 790 1 氨基酸生物合成 Biosynthesis of amino acids ko01230 69 0.003 417 7 精氨酸生物合成 Arginine biosynthesis ko00220 14 0.004 281 4 单环菌素生物合成 Monobactam biosynthesis ko00261 6 0.007 604 3 亚油酸代谢 Linoleic acid metabolism ko00591 7 0.007 743 9 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 48 0.010 722 2 黄酮和黄酮醇生物合成 Flavone and flavonol biosynthesis ko00944 3 0.013 592 9 真核生物中的核糖体生物发生 Ribosome biogenesis in eukaryotes ko03008 28 0.015 049 1 植物昼夜节律 Circadian rhythm - plant ko04712 14 0.020 296 1 油菜素内酯生物合成 Brassinosteroid biosynthesis ko00905 5 0.031 256 2 脂肪酸降解 Fatty acid degradation ko00071 13 0.048 351 3 A-C 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 38 7.98E-06 蛋白质输出 Protein export ko03060 16 3.14E-04 植物昼夜节律 Circadian rhythm - plant ko04712 11 1.99E-03 DNA复制 DNA replication ko03030 14 3.38E-03 RNA聚合酶 RNA polymerase ko03020 8 2.56E-02 A-D 磷酸肌醇代谢 Inositol phosphate metabolism ko00562 39 1.93E-06 异喹啉生物碱的生物合成 Isoquinoline alkaloid biosynthesis ko00950 16 1.59E-04 果糖和甘露糖代谢 Fructose and mannose metabolism ko00051 33 3.64E-03 自噬-其他真核生物 Autophagy - other eukaryotes ko04136 15 4.63E-03 类胡萝卜素生物合成 Carotenoid biosynthesis ko00906 16 9.62E-03 酮体的合成与降解 Synthesis and degradation of ketone bodies ko00072 5 2.43E-02 磷脂酰肌醇信号系统 Phosphatidylinositol signaling system ko04070 28 2.50E-02 A-D 维生素B6代谢 Vitamin B6 metabolism ko00750 8 2.85E-02 赖氨酸降解 Lysine degradation ko00310 17 2.89E-02 玉米素生物合成 Zeatin biosynthesis ko00908 10 3.33E-02 醚脂质代谢 Ether lipid metabolism ko00565 13 4.62E-02 A-E 醚脂质代谢 Ether lipid metabolism ko00565 16 0.000 205 9 非同源末端连接 Non-homologous end-joining ko03450 7 0.000 579 5 淀粉和蔗糖代谢 Starch and sucrose metabolism ko00500 51 0.000 857 3 类胡萝卜素生物合成 Carotenoid biosynthesis ko00906 15 0.001 705 1 甘油磷脂代谢 Glycerophospholipid metabolism ko00564 35 0.003 838 6 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 55 0.004 486 4 玉米素生物合成 Zeatin biosynthesis ko00908 9 0.016 233 1 果糖和甘露糖代谢 Fructose and mannose metabolism ko00051 23 0.036 076 3 B-C 脂肪酸延伸 Fatty acid elongation ko00062 18 0.000 833 6 黄酮合黄酮醇生物合成 Flavone and flavonol biosynthesis ko00944 3 0.010 997 1 亚油酸代谢 Linoleic acid metabolism ko00591 6 0.019 886 3 其他聚糖降解 Other glycan degradation ko00511 7 0.048 730 9 2-羰基甲酸代谢 2-Oxocarboxylic acid metabolism ko01210 17 0.041 135 3 D-E 磷酸肌醇代谢 Inositol phosphate metabolism ko00562 38 1.79E-06 磷脂酰肌醇信号系统 Phosphatidylinositol signaling system ko04070 31 2.35E-03 异喹啉生物碱的生物合成 Isoquinoline alkaloid biosynthesis ko00950 12 9.65E-03 膦酸酯和次膦酸酯代谢 Phosphonate and phosphinate metabolism ko00440 6 8.31E-03 鞘糖脂生物合成-乳糖和新乳糖系列
Glycosphingolipid biosynthesis - lacto and neolacto seriesko00601 2 1.27E-02 RNA转运 RNA transport ko03013 59 1.57E-02 组氨酸代谢 Histidine metabolism ko00340 9 2.64E-02 氮代谢 Nitrogen metabolism ko00910 15 2.09E-02 β-丙氨酸代谢 Beta-Alanine metabolism ko00410 17 2.67E-02 叶酸合成 Folate biosynthesis ko00790 9 4.69E-02 -
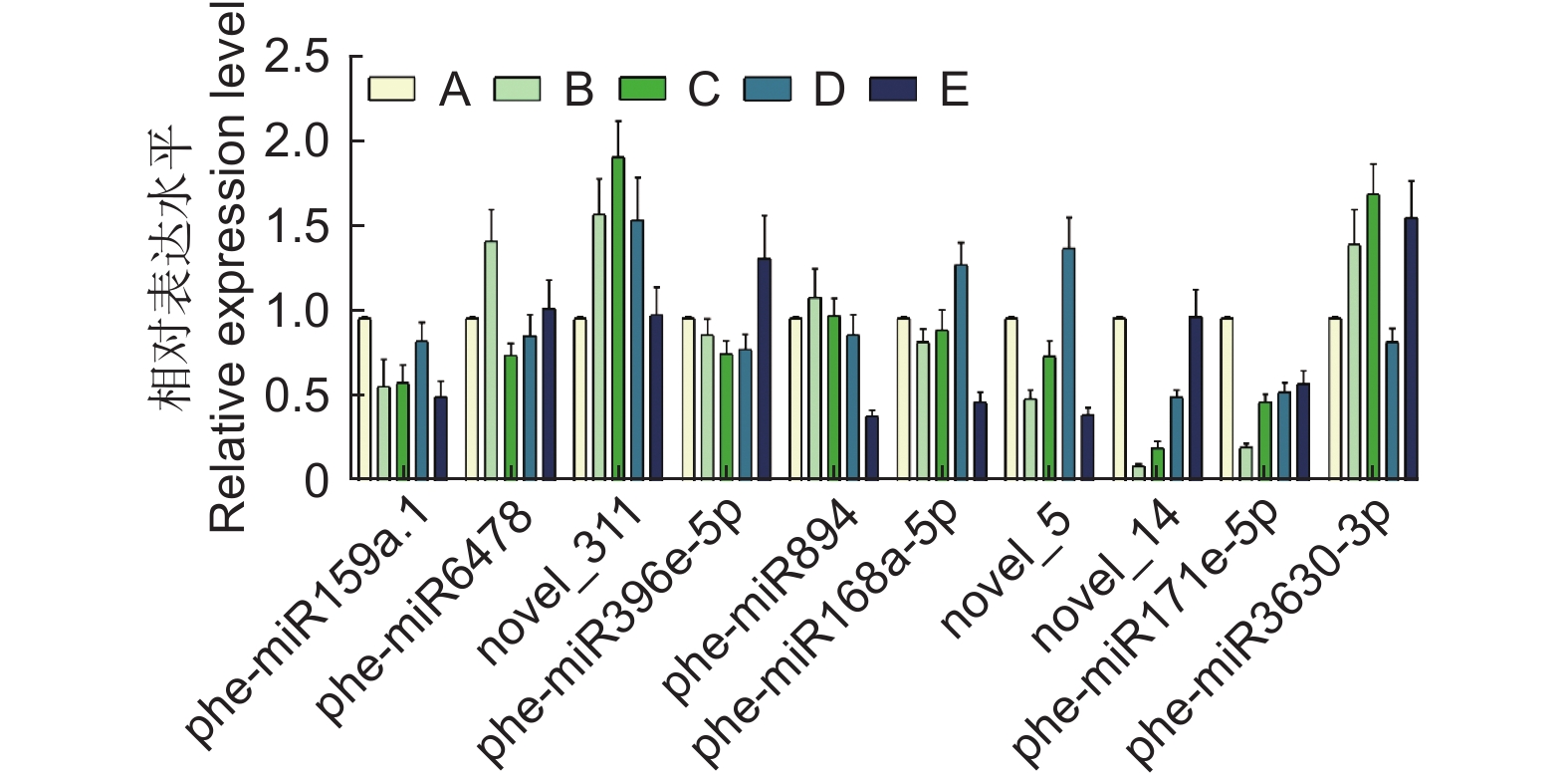
随机选取10个在不同样本间存在差异表达的miRNA进行qPCR验证,miRNA的表达量在不同处理组之间被上调或下调,如phe-miR6478在10% PEG胁迫下比对照组上调1.45倍,phe-miR3630-3p在100 mmol·L−1 NaCl胁迫下上调1.33倍,phe-miR171e-5p在10% PEG、15% PEG、50 mmol·L−1 NaCl、100 mmol·L−1 NaCl胁迫下分别下调0.23、0.50、0.56和0.61倍(图4)。miRNA荧光定量结果和small RNA测序数据在表达趋势上整体一致,表明测序数据的可靠性。
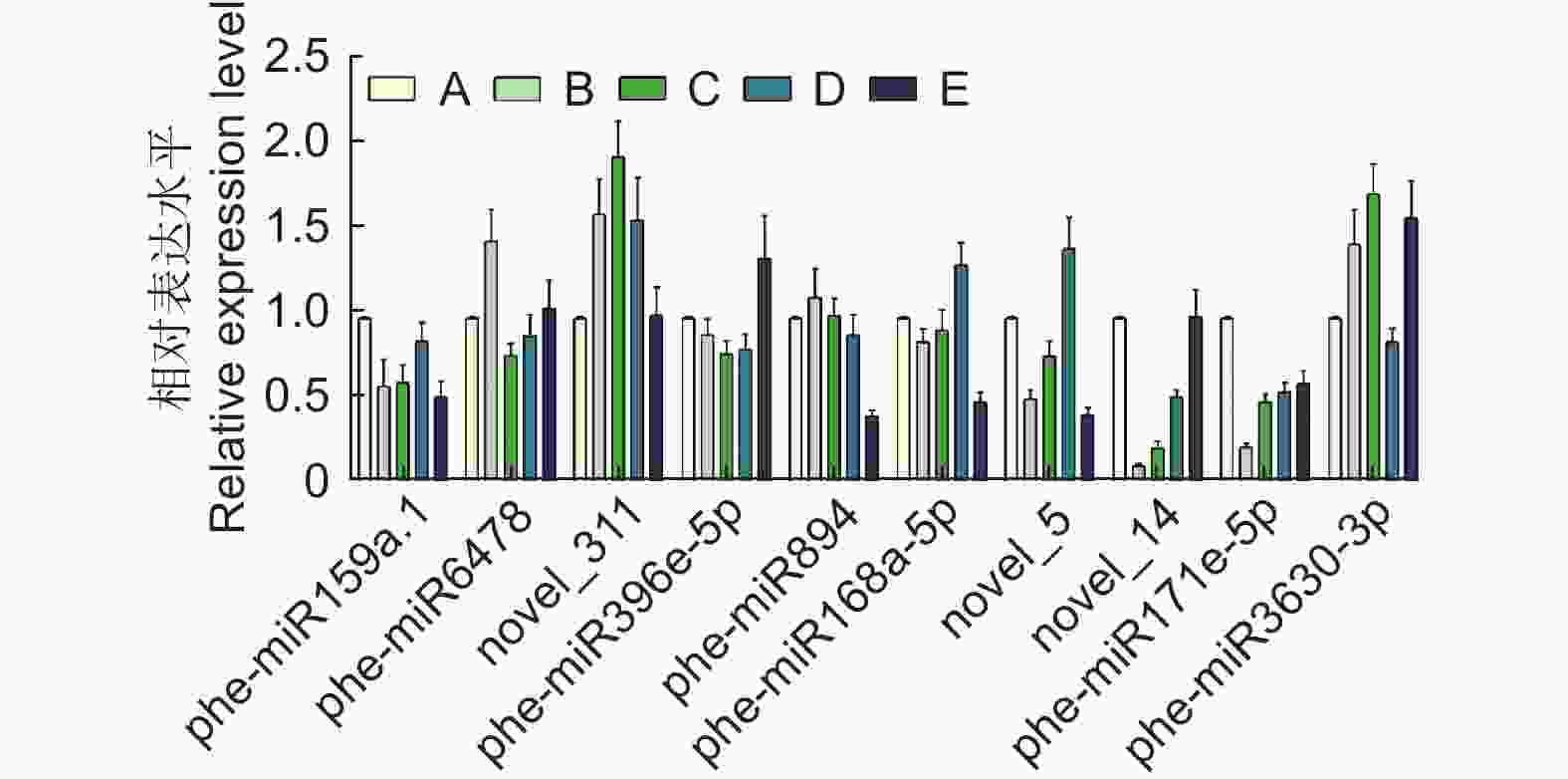
图 4 差异表达miRNA的qPCR验证
Figure 4. qPCR validation for DEmiRNAs
-
在本研究中,miR166、miR159、miR319、miR156、miR396、miR167、miR168、miR894、miR160等在对照组和不同浓度的PEG和NaCl胁迫条件下均具有较高表达水平,推测这些miRNA可能在毛竹种子萌发中有重要作用,其中,miR159[36]、miR156[37]、miR396[38]、miR160[17]已经被证明参与了种子萌发调控。在大麦(Hordeum vulgare L.)萌发种子中,miR156、miR166、miR167、miR168等也具有高表达水平,同时miR5071被大量积累,而在本研究中未检测到miR5071的表达,表明不同物种间miRNA对种子萌发的调控具有特异性[39]。在已有研究中,miR402[40]、miR163[41]和miR417[42]被证明能够参与种子萌发期的干旱或盐胁迫响应,而在本研究对照和处理组中均未检测到miR402、miR163和miR417的表达,推测这些miRNA在本研究中可能不是响应干旱或盐胁迫的关键miRNA,其分布和表达可能与物种有关。
与对照组相比,本研究中miRNA主要在单一胁迫下显著差异表达,仅有2个miRNA能够同时响应2种胁迫,表明miRNA在10% PEG、15% PEG、50 mmol·L−1 NaCl、100 mmol·L−1 NaCl 4种胁迫下的调控存在差异。本研究丰度较高且显著差异表达的miRNA中,phe-miR171e-5p能够响应10% PEG和50 mmol·L−1 NaCl胁迫,phe-miR3630-3p在15% PEG胁迫下显著上调表达,phe-miR159a.1在100 mmol·L−1 NaCl胁迫下差异表达,推测这3个miRNA对于毛竹种子萌发响应PEG或NaCl胁迫可能具有重要意义。
前人的研究中,PEG胁迫下甘蓝型油菜(Brassica napus L.)萌发种子中的miR171比对照组显著下调表达[43];此外,在拟南芥(Arabidopsis thaliana (L.) Heynh.)中过表达桑树(Morus alba L.)mno-miR171可提高转基因植株在NaCl和甘露醇胁迫下的种子发芽率[44],这与本研究phe-miR171e-5p能够响应PEG和NaCl胁迫是一致的。miR171主要通过靶向GRAS家族基因影响GA和ABA信号通路,参与胚胎发生潜能的维持[45-47]、花药发育、芽分枝和复叶形态的调控[48]、顶端优势的调控[49]等,其是否通过GRAS家族参与毛竹种子萌发过程中对干旱和盐胁迫的调控需要进一步研究。在苜蓿(Medicago sativa L.)中,miR3630被鉴定为响应干旱胁迫的miRNA[50],本研究中phe-miR3630-3p在15% PEG胁迫下显著上调表达,但目前关于miR3630的研究较少,本研究为miR3630响应干旱胁迫提供了新的证据。此外,phe-miR159a.1在A-E比较组显著下调。拟南芥中miR159通过介导GAMYB家族MYB101和MYB33的转录本切割,参与种子萌发过程糊粉层细胞的细胞程序性死亡[18],推测miR159对于100 mmol·L−1 NaCl胁迫下毛竹种子的萌发也可能具有重要的调控作用。
-
本研究首次系统鉴定了毛竹萌发种子中的miRNA,并对其在PEG和NaCl胁迫下毛竹萌发露白阶段种子中的表达模式进行研究,探究了响应不同PEG和NaCl胁迫的差异表达miRNA。下一步将对本研究中涉及的重要miRNA进行靶基因的验证,并对其调控机制和调控功能进行深入研究和探讨。
PEG和NaCl胁迫下毛竹萌发种子的MicroRNAs表达谱分析
Identification of MicroRNAs during Seed Germination and Its Response to PEG and NaCl Stresses
-
摘要:
目的 鉴定和分析不同干旱和盐胁迫下毛竹种子在露白时期的microRNAs(miRNAs)及其表达模式。 方法 分别使用聚乙二醇(PEG6000)和氯化钠(NaCl)模拟干旱和盐胁迫,构建H2O、10% PEG、15% PEG、50 mmol·L−1 NaCl和100 mmol·L−1 NaCl处理下毛竹露白时期种子的small RNA文库,通过高通量测序和生物信息学分析揭示样本中的miRNA及其表达谱。 结果 通过small RNA测序共鉴定了246条已知miRNAs的成熟体序列,并预测得到262条novel miRNAs的成熟体序列;在毛竹种皮破裂阶段的种子中,丰度最高的已知miRNA为miR166,其次是miR159、miR6478、miR319等;根据miRNA靶基因预测结果,靶基因数量最多的已知miRNA属于MIR396家族,PH02Gene13935(GAMYB)被预测受到MIR159、MIR319、MIR396家族共28个miRNAs的调控;在6个比较组中共鉴定了181个差异表达miRNA,与对照组相比,10% PEG、15% PEG、50 mmol·L−1 NaCl和100 mmol·L−1 NaCl胁迫下表达水平最高且显著差异表达的已知miRNA分别为phe-miR171e-5p、phe-miR3630-3p、phe-miR171e-5p和phe-miR159a.1;差异表达miRNA靶基因能够显著富集在不同的GO和KEGG途径中;对10个差异表达miRNAs进行qRT-PCR验证,荧光定量结果的整体趋势和测序数据一致。 结论 毛竹种皮破裂阶段种子中积累了大量的miR159、miR6478、miR319等已知miRNA,且在对照和胁迫处理组中均具有高表达水平,可能在毛竹种子萌发中具有保守调控作用;与对照组相比,phe-miR171e-5p、phe-miR3630-3p、phe-miR171e-5p和phe-miR159a.1在 10% PEG、15% PEG、50 mmol·L−1 NaCl、100 mmol·L−1 NaCl胁迫下分别显著差异表达,能够在毛竹种子露白阶段响应PEG或NaCl胁迫。 Abstract:Objective To identify microRNAs (miRNAs) and reveal its expressional pattern in seed coat rupture stage of Moso bamboo seeds (Phyllostachys edulis) under different drought and salt stresses. Methods Polyethylene glycol (PEG6000) and NaCl were used to simulate drought and salinity stress, respectively. Small RNA libraries were separately built for Moso bamboo seeds germinated under H2O, 10% PEG, 15% PEG, 50 mmol·L−1 NaCl and 100 mmol·L−1 NaCl, and the seeds were all sampled at seed coat rupture stage. High throughput sequencing and bioinformatics analysis were used to explore the expressional pattern of miRNA. Results A total of 246 known miRNAs and 262 novel mature miRNAs were identified in this study. The most abundant miRNAs in seed coat rupture stage of Moso bamboo was miR166, followed by miR159, miR6478, miR319, etc. According to miRNA target prediction, MIR396 family owned the largest number of target genes, and ph02gene13935 (GAMYB) could to be regulated by 28 miRNAs of MIR159, MIR319 or MIR396; A total of 181 differentially expressed miRNAs (DEmiRNA) were identified in six comparison groups; Compared with control group, in 10% PEG, 15% PEG, 50 mmol·L−1 NaCl and 100 mmol·L−1 NaCl treatments, phe-miR171e-5p, phe-miR3630-3p, phe-miR171e-5p and phe-miR159a were differentially expressed respectively with highest expressional level in known miRNAs; The target genes of DEmiRNA were significantly enriched in different GO and KEGG pathways; Ten DEmiRNAs were verified by qPCR, and the overall trend of qPCR results was consistent with the sequencing data. Conclusion In seed coat rupture stage of moso bamboo, there exhibit high accumulation of known miRNAs such as miR159, miR6478, miR319 in all control and four treatment groups, which may play a conservative regulatory role in Moso bamboo seed germination. Compared with the control group, phe-miR171e-5p, phe-miR3630-3p, phe-miR171e-5p and phe-miR159a 1 are differentially expressed in 10% PEG, 15% PEG, 50 mmol·L−1 NaCl and 100 mmol·L−1 NaCl, respectively, which can respond to PEG or NaCl stress during seed coat rupture stage of Moso bamboo. -
Key words:
- Moso bamboo
- / germination
- / PEG
- / NaCl
- / miRNA
-
图 1 毛竹干种子(a)及种皮破裂阶段取样的萌发种子(b)
Figure 1. Dry seeds (a) and sampled seeds germinated at seed coat rupture stage (b) of Moso bamboo (Phyllostachys edulis)
表 1 差异表达miRNA荧光定量验证引物序列
Table 1. Primers used for quantitative real time PCR of Differential expressed miRNA
miRNA编号
miRNA ID引物方向
Primer direction引物序列
Sequence (5′-3′)phe-miR3630-3p F UGUGGGAAUCUCUCUGAUGCUU phe-miR159a.1 F UUUGGAUUGAAGGGAGCUCUG phe-miR168a-5p F UCGCUUGGUGCAGAUCGGGAC phe-miR171e-5p F UGUUGGCUCGGCUCACUCAGA phe-miR396e-5p F UCCACAGGCUUUCUUGAACUG phe-miR894 F CGUUUCACGUCGGGUUCACC phe-miR6478 F CCGACCUUAGCUCAGUUGGUG novel_14 F CUGAGAGGUAGAGAACGGGAUG novel_311 F UCGUGUGCUGAACUUUAUUGA novel_5 F AUCUCAGCUGUUGAUUCCAUGAUC u6 F GGACATCCGATAAAATTGGAACGATACAG  下载: 导出CSV
下载: 导出CSV
表 2 small RNA数据产出信息
Table 2. Statistics of sequence data in small RNA libraries
样品名称
Sample原始读序
Raw ReadsQ20值
Q20/%Q30值
Q30/%GC含量
GC Content/%过滤后读序数
Clean Reads比对sRNA数量
Mapped sRNAA 29 379 983 98 94 54 12 984 751 11 504 336 (88.60%) B 37 101 403 98 93 54 14 982 423 13 326 206 (88.95%) C 32 426 286 98 94 54 13 401 719 11 915 649 (88.91%) D 33 041 222 98 94 54 13 995 450 12 374 157 (88.42%) E 19 605 421 98 93 53 11 452 796 10 198 110 (89.04%)
下载: 导出CSV
表 3 Small RNA文库中预测出前10位的保守miRNA及其序列表达量及同源性
Table 3. Sequences abundance and homologues of top ten predicted conserved miRNA in five libraries
同源序列名称
Name序列
Sequence表达量 (TPM Counts) A B C D E phe-miR166a-3p UCGGACCAGGCUUCAUUCCCC 74 864 57 593 64 907 73 677 66 116 phe-miR159a.1 UUUGGAUUGAAGGGAGCUCUG 54 736 42 178 41 405 45 018 31 525 phr-miR319a-3p.2-3p UUGGACUGAAGGGUGCUCCC 27 992 25 959 28 454 25 962 27 126 phe-miR6478 CCGACCUUAGCUCAGUUGGUG 25 651 48 135 24 454 21 842 28 992 phe-miR156a-5p UGACAGAAGAGAGUGAGCAC 17 694 16 099 15 274 23 739 28 492 phe-miR167d-5p UGAAGCUGCCAGCAUGAUCUG 17 288 28 696 19 426 18 345 16 262 phe-miR396e-5p CGUUUCACGUCGGGUUCACC 12 919 15 012 12 456 10 017 18 662 phe-miR894 UCCACAGGCUUUCUUGAACUG 11 796 17 105 14 627 10 106 5 332 phe-miR166k-3p UCGGACCAGGCUUCAAUCCCU 8 925 6 641 5 866 11 706 14 463 phe-miR319a UUGGACUGAAGGGAGCUCCCU 8 707 6 761 6 437 7 528 7 898
下载: 导出CSV
表 4 Small RNA文库中预测出前10位新miRNA序列及其表达量
Table 4. Sequences abundance and homologues of top ten predicted novel miRNA in five libraries
名称
Name序列
Sequence表达量 (TPM counts) 最小折叠自由能
Hairpin mfe/(KJ·mol−1)A B C D E novel_1 CUUCACUCUUCAGACUUUGAG 174 786 172 858 173 542 135 291 189 416 −46.6 novel_199 CGAUUAGUCGGCAAGUCGGGCGACU 17 413 23 705 16 760 12 951 15 796 −44.7 novel_311 UCGUGUGCUGAACUUUAUUGA 15 135 31 513 36 072 22 702 16 496 −13.4 novel_241 UUACCGGUCUUUAAACUUUUUAU 14 792 17 990 26 054 17 486 11 764 −175.9 novel_2 CCGGUCUAUAGAUUUUCAAAGUG 8 332 10 464 14 170 13 544 9 664 −115.7 novel_3 UCUCGGAUCAGGCUUCGUUCC 7 084 5 594 5 180 5 424 10 664 −39.4 novel_287 AUCGCGAUCUUGACAACCUUGACU 5 586 6 600 8 761 9 543 7 165 −27.5 novel_5 AUCUCAGCUGUUGAUUCCAUGAUC 4 993 3 381 4 723 6 698 2 299 −32.8 novel_9 CCUUCGAUCGUAUGAUGUGUCACG 4 494 4 306 4 457 4 742 2 799 −98.6 novel_87 UGUAGGAUCUCUGAUGACGGU 3 776 5 031 5 371 3 082 6 765 −187.0
下载: 导出CSV
表 5 不同比较组差异表达miRNA靶基因GO显著富集性前10条目
Table 5. Top 10 significantly enriched GO terms for DEmiRNA target genes in different comparision groups
比较组
Comparison GroupsGO编号
GO IDGO条目
GO Term基因数量
Gene NumberP值
P-ValueA-B GO:0055085 跨膜转运Transmembrane transport 233 2.49E-11 A-B GO:0098656 阴离子跨膜转运Anion transmembrane transport 49 1.12E-07 A-B GO:0006820 阴离子转运 Anion transport 75 1.48E-07 A-B GO:1903825 有机酸跨膜转运 Organic acid transmembrane transport 41 1.83E-07 A-B GO:1905039 羧酸跨膜转运 Carboxylic acid transmembrane transport 41 1.83E-07 A-B GO:0003333 氨基酸跨膜转运 Amino acid transmembrane transport 38 2.49E-07 A-B GO:0006865 氨基酸转运 Amino acid transport 38 6.65E-07 A-B GO:0006811 离子转运 Ion transport 169 6.90E-07 A-B GO:0006885 pH调节 Regulation of pH 25 1.14E-06 A-B GO:0055067 单价无机阳离子稳态 Monovalent inorganic cation homeostasis 26 1.21E-06 A-C GO:0045740 DNA复制的正调控 Positive regulation of DNA replication 7 4.04E-07 A-C GO:0034644 细胞对紫外线的反应 Cellular response to UV 6 1.23E-06 A-C GO:0019985 跨损伤合成 Translesion synthesis 7 1.40E-06 A-C GO:0000731 参与DNA修复的DNA合成 DNA synthesis involved in DNA repair 8 1.57E-06 A-C GO:0051052 DNA代谢过程的调节 Regulation of DNA metabolic process 14 1.99E-06 A-C GO:0072593 活性氧代谢过程 Reactive oxygen species metabolic process 36 2.97E-06 A-C GO:0055074 钙离子稳态 Calcium ion homeostasis 5 3.06E-06 A-C GO:0071482 细胞对光刺激的反应 Cellular response to light stimulus 13 3.24E-06 A-C GO:0006979 对氧化应激的反应 Response to oxidative stress 41 5.55E-06 A-C GO:0051054 DNA代谢的正调控 Positive regulation of DNA metabolic process 7 6.17E-06 A-D GO:0032502 发育过程 Developmental process 288 9.35E-09 A-D GO:0080022 初生根发育 Primary root development 11 6.87E-08 A-D GO:0044767 单一有机体发育过程 Single-organism developmental process 266 1.59E-06 A-D GO:0016567 蛋白质泛素化 Protein ubiquitination 104 2.24E-06 A-D GO:0009657 质体组织 Plastid organization 46 2.80E-06 A-D GO:0070647 通过小蛋白结合或去除来修饰蛋白质
Protein modification by small protein conjugation or removal128 3.51E-06 A-D GO:0046271 苯丙烷代谢生物过程 Phenylpropanoid catabolic process 21 6.65E-06 A-D GO:0046274 木质素分解代谢过程 Lignin catabolic process 21 6.65E-06 A-E GO:0005992 海藻糖生物合成过程 Trehalose biosynthetic process 19 1.87E-06 A-E GO:0071478 细胞对辐射的响应 Cellular response to radiation 22 2.64E-06 A-E GO:0071214 细胞对非生物胁迫的响应 Cellular response to abiotic stimulus 22 4.18E-06 A-E GO:0044767 单一有机体发育过程 Single-organism developmental process 214 8.24E-06 A-E GO:0032502 发育过程 Developmental process 220 9.83E-06 B-C GO:0055085 跨膜转运 Transmembrane transport 230 1.21E-14 B-C GO:0006811 离子转运 Ion transport 173 1.54E-10 B-C GO:0006820 阴离子转运 Anion transport 78 2.58E-10 B-C GO:1903825 有机酸跨膜转运 Organic acid transmembrane transport 42 4.73E-09 B-C GO:1905039 羧酸跨膜转运 Carboxylic acid transmembrane transport 42 4.73E-09 B-C GO:0003333 氨基酸跨膜转运 Amino acid transmembrane transport 39 7.16E-09 B-C GO:0006865 氨基酸转运 Amino acid transport 39 2.11E-08 B-C GO:0006810 转运 Transport 424 2.33E-08 B-C GO:0051234 定位的建成 Establishment of localization 427 5.56E-08 B-C GO:0051179 定位 Localization 436 9.61E-08 D-E GO:0080022 初生根发育 Primary root development 12 1.52E-09 D-E GO:0006642 甘油三酯动员 Triglyceride mobilization 7 7.18E-06 D-E GO:0009658 叶绿体组织 Chloroplast organization 35 7.27E-06 D-E GO:0048506 分生组织相变时间调控 Regulation of timing of meristematic phase transition 9 8.33E-06
下载: 导出CSV
表 6 差异表达miRNA靶基因KEGG显著富集结果
Table 6. Significantly enriched KEGG pathways for DEmiRNA target genes in different comparison groups
比较组
Comparision GroupsKEGG途径
KEGG Pathway途径ID
Pathway ID基因数量
Gene NumberP值
P-ValueA-B 糖基磷脂酰肌醇(GPI)-锚定生物合成
Glycosylphosphatidylinositol(GPI)-anchor biosynthesisko00563 10 0.000 709 3 脂肪酸延伸 Fatty acid elongation ko00062 18 0.001 981 2 2-羰基甲酸代谢 2-Oxocarboxylic acid metabolism ko01210 22 0.002 790 1 氨基酸生物合成 Biosynthesis of amino acids ko01230 69 0.003 417 7 精氨酸生物合成 Arginine biosynthesis ko00220 14 0.004 281 4 单环菌素生物合成 Monobactam biosynthesis ko00261 6 0.007 604 3 亚油酸代谢 Linoleic acid metabolism ko00591 7 0.007 743 9 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 48 0.010 722 2 黄酮和黄酮醇生物合成 Flavone and flavonol biosynthesis ko00944 3 0.013 592 9 真核生物中的核糖体生物发生 Ribosome biogenesis in eukaryotes ko03008 28 0.015 049 1 植物昼夜节律 Circadian rhythm - plant ko04712 14 0.020 296 1 油菜素内酯生物合成 Brassinosteroid biosynthesis ko00905 5 0.031 256 2 脂肪酸降解 Fatty acid degradation ko00071 13 0.048 351 3 A-C 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 38 7.98E-06 蛋白质输出 Protein export ko03060 16 3.14E-04 植物昼夜节律 Circadian rhythm - plant ko04712 11 1.99E-03 DNA复制 DNA replication ko03030 14 3.38E-03 RNA聚合酶 RNA polymerase ko03020 8 2.56E-02 A-D 磷酸肌醇代谢 Inositol phosphate metabolism ko00562 39 1.93E-06 异喹啉生物碱的生物合成 Isoquinoline alkaloid biosynthesis ko00950 16 1.59E-04 果糖和甘露糖代谢 Fructose and mannose metabolism ko00051 33 3.64E-03 自噬-其他真核生物 Autophagy - other eukaryotes ko04136 15 4.63E-03 类胡萝卜素生物合成 Carotenoid biosynthesis ko00906 16 9.62E-03 酮体的合成与降解 Synthesis and degradation of ketone bodies ko00072 5 2.43E-02 磷脂酰肌醇信号系统 Phosphatidylinositol signaling system ko04070 28 2.50E-02 A-D 维生素B6代谢 Vitamin B6 metabolism ko00750 8 2.85E-02 赖氨酸降解 Lysine degradation ko00310 17 2.89E-02 玉米素生物合成 Zeatin biosynthesis ko00908 10 3.33E-02 醚脂质代谢 Ether lipid metabolism ko00565 13 4.62E-02 A-E 醚脂质代谢 Ether lipid metabolism ko00565 16 0.000 205 9 非同源末端连接 Non-homologous end-joining ko03450 7 0.000 579 5 淀粉和蔗糖代谢 Starch and sucrose metabolism ko00500 51 0.000 857 3 类胡萝卜素生物合成 Carotenoid biosynthesis ko00906 15 0.001 705 1 甘油磷脂代谢 Glycerophospholipid metabolism ko00564 35 0.003 838 6 苯丙烷生物合成 Phenylpropanoid biosynthesis ko00940 55 0.004 486 4 玉米素生物合成 Zeatin biosynthesis ko00908 9 0.016 233 1 果糖和甘露糖代谢 Fructose and mannose metabolism ko00051 23 0.036 076 3 B-C 脂肪酸延伸 Fatty acid elongation ko00062 18 0.000 833 6 黄酮合黄酮醇生物合成 Flavone and flavonol biosynthesis ko00944 3 0.010 997 1 亚油酸代谢 Linoleic acid metabolism ko00591 6 0.019 886 3 其他聚糖降解 Other glycan degradation ko00511 7 0.048 730 9 2-羰基甲酸代谢 2-Oxocarboxylic acid metabolism ko01210 17 0.041 135 3 D-E 磷酸肌醇代谢 Inositol phosphate metabolism ko00562 38 1.79E-06 磷脂酰肌醇信号系统 Phosphatidylinositol signaling system ko04070 31 2.35E-03 异喹啉生物碱的生物合成 Isoquinoline alkaloid biosynthesis ko00950 12 9.65E-03 膦酸酯和次膦酸酯代谢 Phosphonate and phosphinate metabolism ko00440 6 8.31E-03 鞘糖脂生物合成-乳糖和新乳糖系列
Glycosphingolipid biosynthesis - lacto and neolacto seriesko00601 2 1.27E-02 RNA转运 RNA transport ko03013 59 1.57E-02 组氨酸代谢 Histidine metabolism ko00340 9 2.64E-02 氮代谢 Nitrogen metabolism ko00910 15 2.09E-02 β-丙氨酸代谢 Beta-Alanine metabolism ko00410 17 2.67E-02 叶酸合成 Folate biosynthesis ko00790 9 4.69E-02
下载: 导出CSV
-
[1] 江泽慧. 世界竹藤[M]. 沈阳: 辽宁科学技术出版社, 2002: 221-352. [2] 汪奎宏, 黄伯惠. 中国毛竹[M]. 杭州: 浙江科学技术出版社, 1996: 1-29. [3] 谭宏超, 陶现灵, 谭汝强. 13种竹种子低温储存条件及种子育苗环境研究[J]. 世界竹藤通讯, 2018, 16(1):16-20. [4] 姚文静, 王 茹, 王 星, 等. 毛竹实生苗生长发育规律及其模型拟合研究[J]. 西部林业科学, 2020, 49(3):14-20,28. [5] 张金菊, 张国敏, 贾碧玉, 等. 毛竹种子育苗技术初探[J]. 现代园艺, 2015(12):26. doi: 10.3969/j.issn.1006-4958.2015.12.021 [6] ACOSTA-MOTOS J, ORTUÑO M, BERNAL-VICENTE A, et al. Plant responses to salt stress: adaptive mechanisms[J]. Agronomy, 2017, 7(1): 18. doi: 10.3390/agronomy7010018 [7] SINGH M, KUMAR J, SINGH S, et al. Roles of osmoprotectants in improving salinity and drought tolerance in plants: a review[J]. Reviews in Environmental Science and Bio/Technology, 2015, 14(3): 407-426. doi: 10.1007/s11157-015-9372-8 [8] 张乃华, 汪爱君, 黄超钢, 等. 毛竹种子育苗造林技术[J]. 世界竹藤通讯, 2012(2):32-34. doi: 10.3969/j.issn.1672-0431.2012.02.010 [9] 黄业伟, 杨 丽, 张智俊. NaCl胁迫对毛竹种子萌发及幼苗生长的影响[J]. 种子, 2009(10):16-18. [10] 宋沁春, 魏 开, 漆冬梅, 等. 盐胁迫下超声波处理对毛竹种子萌发及幼苗生长的影响[J]. 种子, 2018(3):83-85. doi: 10.16590/j.cnki.1001-4705.2018.03.083 [11] 杨振亚, 周本智, 周 燕, 等. PEG模拟干旱对毛竹种子萌发及生长生理特性的影响[J]. 林业科学研究, 2018, 31(6):47-54. doi: 10.13275/j.cnki.lykxyj.2018.06.007 [12] 蔡春菊, 范少辉, 曹帮华, 等. PEG和GA_3引发处理对老化毛竹种子理化特性的影响[J]. 南京林业大学学报(自然科学版), 2018, 42(2):40-46. [13] ZHANG B H, PAN X P, COBB G P, et al. Plant microRNA: a small regulatory molecule with big impact[J]. Developmental Biology, 2006, 289(1): 3-16. doi: 10.1016/j.ydbio.2005.10.036 [14] DONG Q K, HU B B, ZHANG C. MicroRNAs and their roles in plant development[J]. Frontiers in Plant Science, 2022, 13: 824240. doi: 10.3389/fpls.2022.824240 [15] ISLAM W, ADNAN M, HUANG Z Q, et al. Small RNAs from seed to mature plant[J]. Critical Reviews in Plant Sciences, 2019, 38(2): 117-139. doi: 10.1080/07352689.2019.1608404 [16] DAS S S, KARMAKAR P, NANDI A K, et al. Small RNA mediated regulation of seed germination[J]. Frontiers in Plant Science, 2015, 6: 828. [17] LIU P, MONTGOMERY T A, FAHLGREN N, et al. Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages[J]. Plant Journal, 2007, 52(1): 133-146. doi: 10.1111/j.1365-313X.2007.03218.x [18] REYES J L, CHUA N. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination[J]. Plant Journal, 2007, 49(4): 592-606. doi: 10.1111/j.1365-313X.2006.02980.x [19] ZHANG B. MicroRNA: a new target for improving plant tolerance to abiotic stress[J]. Journal of Experimental Botany, 2015, 66(7): 1749-1761. doi: 10.1093/jxb/erv013 [20] ZHAO H, WANG L, DONG L, et al. Discovery and comparative profiling of microRNAs in representative monopodial bamboo (Phyllostachys edulis) and sympodial bamboo (Dendrocalamus latiflorus)[J]. PloS ONE, 2014, 9(7): e102375. doi: 10.1371/journal.pone.0102375 [21] GE W, ZHANG Y, CHENG Z, et al. Main regulatory pathways, key genes and microRNAs involved in flower formation and development of Moso Bamboo (Phyllostachys edulis)[J]. Plant Biotechnology Journal, 2017, 15(1): 82-96. doi: 10.1111/pbi.12593 [22] CHENG Z, HOU D, GE W, et al. Integrated mRNA, microRNA transcriptome and degradome analyses provide insights into stamen development in Moso Bamboo[J]. Plant and Cell Physiology, 2020, 61(1): 76-87. doi: 10.1093/pcp/pcz179 [23] WANG K L, ZHANG Y, ZHANG H M, et al. MicroRNAs play important roles in regulating the rapid growth of the Phyllostachys edulis culm internode[J]. New Phytologist, 2021, 231(6): 2215-2230. doi: 10.1111/nph.17542 [24] JIN Q, PENG H, LIN E, et al. Identification and characterization of differentially expressed miRNAs between bamboo shoot and rhizome shoot[J]. Journal of Plant Biology, 2016, 59(4): 322-335. doi: 10.1007/s12374-015-0581-z [25] KOZOMARA A, GRIFFITHS-JONES S. MiRBase: annotating high confidence microRNAs using deep sequencing data[J]. Nucleic Acids Research, 2013, 42(D1): D68-D73. [26] KALVARI I, NAWROCKI E P, ONTIVEROS-PALACIOS N, et al. Rfam 14: expanded coverage of metagenomic, viral and microRNA families[J]. Nucleic Acids Research, 2021, 49(D1): D192-D200. doi: 10.1093/nar/gkaa1047 [27] FRIEDLNDER M R, MACKOWIAK S D, NA L, et al. MiRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades[J]. Nucleic Acids Research, 2012(1): 37-52. [28] WEN M, SHEN Y, SHI S, et al. MiREvo: an Integrative microRNA evolutionary analysis platform for next-generation sequencing experiments[J]. BMC Bioinformatics, 2012, 13(1): 140. doi: 10.1186/1471-2105-13-140 [29] ROBINSON M D, MCCARTHY D J, SMYTH G K. EdgeR: a bioconductor package for differential expression analysis of digital gene expression data[J]. Bioinformatics, 2009, 26(1): 139-140. [30] DAI X, ZHAO P X. PsRNATarget: a plant small rna target analysis server[J]. Nucleic Acids Research, 2011, 39: W155-W159. doi: 10.1093/nar/gkr319 [31] FAHLGREN N, CARRINGTON J C. MiRNA target prediction in plants[J]. Methods in Molecular Biology (Clifton, N. J. ), 2010, 592: 51-57. [32] KANEHISA M. The KEGG resource for deciphering the genome[J]. Nucleic Acids Research, 2004, 32(90001): 277D-280D. doi: 10.1093/nar/gkh063 [33] YOUNG M D, WAKEFIELD M J, SMYTH G K, et al. Gene ontology analysis for RNA-seq: accounting for selection bias[J]. Genome Biology, 2010, 11(2): R14. doi: 10.1186/gb-2010-11-2-r14 [34] 黄志明, 李晨曦, 陈 磊, 等. 毛竹4个miRNA在胚芽萌发过程的表达响应研究[J]. 热带作物学报, 2017, 38(11):2112-2118. doi: 10.3969/j.issn.1000-2561.2017.11.019 [35] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method[J]. Methods, 2001, 25(4): 402-408. doi: 10.1006/meth.2001.1262 [36] ALEX D, BACH T J, CHYE M L. Expression of Brassica juncea 3-hydroxy-3-methylglutaryl CoA synthase is developmentally regulated and stress-Responsive[J]. Plant Journal, 2000, 22(5): 415-426. doi: 10.1046/j.1365-313X.2000.00751.x [37] NONOGAKI H. MicroRNA gene regulation cascades during early stages of plant development[J]. Plant and Cell Physiology, 2010, 51(11): 1840-1846. doi: 10.1093/pcp/pcq154 [38] LU Y, FENG Z, MENG Y, et al. SLENDER RICE1 and Oryza sativa INDETERMINATE DOMAIN2 regulating osmiR396 are involved in stem elongation[J]. Plant Physiology, 2020, 182(4): 2213-2227. doi: 10.1104/pp.19.01008 [39] BAI B, SHI B, HOU N, et al. MicroRNAs participate in gene expression regulation and phytohormone cross-talk in barley embryo during seed development and germination[J]. BMC Plant Biology, 2017, 17(1): 150. doi: 10.1186/s12870-017-1095-2 [40] KIM J Y, KWAK K J, JUNG H J, et al. MicroRNA402 affects seed germination of Arabidopsis thaliana under stress conditions via targeting Demeter-Like Protein3 mRNA[J]. Plant and Cell Physiology, 2010, 51(6): 1079-1083. doi: 10.1093/pcp/pcq072 [41] CHUNG P J, PARK B S, WANG H, et al. Light-inducible miR163 targets PXMT1 transcripts to promote seed germination and primary root elongation in Arabidopsis[J]. Plant Physiology, 2016, 170(3): 1772-1782. doi: 10.1104/pp.15.01188 [42] JUNG H J, KANG H. Expression and functional analyses of microRNA417 in Arabidopsis thaliana under stress conditions[J]. Plant Physiology and Biochemistry, 2007, 45(10-11): 805-811. doi: 10.1016/j.plaphy.2007.07.015 [43] JIAN H, WANG J, WANG T, et al. Identification of rapeseed microRNAs involved in early stage seed germination under salt and drought stresses[J]. Frontiers in Plant Science, 2016, 7: 658. [44] SUN Z, KUMAR R M S, LI J, et al. In silico search and biological validation of microR171 family related to abiotic stress response in mulberry (Morus alba)[J]. Horticultural Plant Journal, 2022, 8(2): 184-194. doi: 10.1016/j.hpj.2021.11.003 [45] ZANG Q, ZHANG Y, HAN S, et al. Transcriptional and post-transcriptional regulation of the miR171-LaSCL6 module during somatic embryogenesis in Larix kaempferi[J]. Trees, 2021, 35(1): 145-154. doi: 10.1007/s00468-020-02026-2 [46] LI H, ZHANG J, YANG Y, et al. MiR171 and its target gene SCL6 contribute to embryogenic callus induction and torpedo-shaped embryo formation during somatic embryogenesis in two lily species[J]. Plant Cell, Tissue and Organ Culture (PCTOC), 2017, 130(3): 591-600. doi: 10.1007/s11240-017-1249-9 [47] SHI Q, LONG J, YIN Z, et al. MiR171 modulates induction of somatic embryogenesis in Citrus callus[J]. Plant Cell Reports, 2022, 41(6): 1403-1415. doi: 10.1007/s00299-022-02865-y [48] KRAVCHIK M, STAV R, BELAUSOV E, et al. Functional characterization of microRNA171 family in tomato[J]. Plants, 2019, 8(1): 10. doi: 10.3390/plants8010010 [49] JIANG S, CHEN Q, ZHANG Q, et al. Pyr-miR171f-targeted PyrSCL6 and PyrSCL22 genes regulate shoot growth by responding to IAA signaling in pear[J]. Tree Genetics & Genomes, 2018, 14(2): 20. [50] LI Y, WAN L, BI S, et al. Identification of drought-responsive microRNAs from roots and leaves of alfalfa by high-throughput sequencing[J]. Genes, 2017, 8(4): 119. doi: 10.3390/genes8040119 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5158
- HTML全文浏览量: 2076
- PDF下载量: 59
- 被引次数: 0
