

-
油茶(Camellia oleifera Abel.)隶属山茶科(Theaceae)山茶属(Camellia),为常绿小乔木或灌木,是我国特有的木本油料树种,与油橄榄(Olea europaea L.)、油棕(Elaeis guineensis Jacq.)和椰子(Cocos nucifera L.)并称世界四大木本油料植物[1]。油茶果极具经济价值,茶油富含不饱和脂肪酸,是优质保健食用植物油[2]。近年来,我国植物油自给率逐年下降,对油料和植物油的进口均居世界前列[3]。为缓解我国油料供需矛盾,国务院批准实施《全国油茶产业发展规划》(2009—2020),2020年我国油茶种植面积超过462万hm2[4]。新形势下,油茶产业发展前景良好,种植面积逐渐增大。
磷(P)是植物细胞内的重要组成部分,如磷酸、磷脂和蛋白质等,约占植物干质量的0.05%~0.5%,是植物生长过程中需要的大量营养元素之一。它不仅参与植物多种生理过程如光合作用、酶活性调节等,还是调节能量代谢、信号传导过程中关键因子[5-6]。植物缺磷会影响细胞分裂,从而导致植株出现矮化、叶片枯黄、枝干细弱等症状[7]。油茶林的栽培对土壤肥力要求很高、需肥量大,而红壤中可利用磷的含量不能满足油茶林生长发育的需要,磷是限制红壤区油茶生长发育的主要营养元素之一[8]。近年来,油茶集约化经营过程中常配施大量的磷肥,然而施入的磷肥极易被土壤中的金属离子固定,导致植物所吸收的磷素仅占施磷量的25%左右[9]。而过量的磷流失到土壤中会造成土壤退化、板结和水体富营养化等环境问题,从而降低油茶林的生产力[10]。因此,如何提高油茶林地磷素有效性,降低磷肥使用和减轻磷肥对土壤环境的压力已成为经济作物的热点研究之一。
丛枝菌根(arbuscular mycorrhiza,AM)真菌,隶属球囊菌门(Glomeromycota),能与大约80%的陆生植物根系形成共生关系[11]。AM真菌从植物获取碳源,同时向植物转运多种营养物质,具有促进植物生长、改善植物营养状况、提高植物抵抗生物和非生物逆境的功能[11-12]。大量研究表明丛枝菌根真菌在植物利用磷中起着重要作用,一方面,丛枝菌根真菌侵染植株后,其根尖表皮变厚,细胞层数增多,根冠比增加,根系长度变长,根系与土壤接触面积增大,覆盖范围可超过根系的700倍,进一步提高宿主植物对土壤磷的吸收作用[13-14]。另一方面,丛枝菌根真菌的根外菌丝能够分泌一些物质,包括磷酸酶、糖类和有机酸等,而这些酶可以促进土壤中难溶性的磷进行溶解,从而提高植物的磷吸收效率。与此同时,大量研究表明,在低磷条件下,植物对磷的吸收主要依靠菌丝来完成[15-16]。AM真菌可为植物提供高达100%的磷,金鑫等[17]研究表明根外菌丝对局部P养分获取受环境调控,在环境磷养分较低而局部磷养分高于环境磷养分时,较多的菌丝会进入局部区域获取磷。通过单个真菌菌丝营养吸收的数学模型发现菌丝吸收P的速率比根的吸收速率高出一个数量级[18],也有研究表明接种AM真菌可能会完全抑制植物根对P的吸收[19]。目前研究推测AM真菌对根吸收的限制可能是两者竞争所致,如P转运蛋白的减少[16],在高磷条件下,AM真菌生物量显著降低[20],研究表明随着P含量的增加,未接种的豆科植物地上部磷含量增量要远高于未接种处理,而地下部无显著差异,即在高磷条件下AM真菌可能会限制P向地上部的传递[21]。本课题组前期研究表明:油茶是菌根植物,施有机磷可以改变菌根化油茶根系的形态结构,AM真菌通过提高油茶对矿质元素的吸收来增强叶片的光合作用,改善根系的形态结构,进而促进油茶生长和生物量的增加[22]。然而AM真菌对植物无机磷的吸收和培养土壤磷组分转化的影响还未见报道。因此,本研究以油茶1年生实生苗为试验材料,采用盆栽试验探究不同无机磷(KH2PO4)水平下,接种幼套近明球囊酶(Claroideogolmus etuicatum)对油茶无机磷形态、植株磷浓度和磷转运蛋白基因的影响,为油茶集约化经营中通过菌根生物技术途径提升土壤磷素利用效率提供科学依据。
-
选用油茶赣无2高产品种的1年生实生苗为供试植物材料,由江西省林业科学院提供。
-
供试幼套近明球囊霉(Claroideogolmus etuicatum)菌剂购自长江大学根系生物学研究所,并通过纯沙(灭菌)栽培玉米和三叶草(Trifolium pretense)扩繁菌剂,菌剂包括菌丝、孢子(密度为34个·g−1)、宿主根及基质。
-
试验土壤采自江西农业大学红壤土,土壤基本理化特征为:pH 5.8,有机质41.25 g·kg−1,有效磷2.5 mg·kg−1,铵态氮14.24 mg·kg−1,硝态氮2.26 mg·kg−1,过2 mm筛,经过高温灭菌(121 ℃,2 h)处理,以消除土著微生物的干扰。同时取过2 mm筛的河沙,用自来水冲洗干净,晾干后干热灭菌(180 ℃,4 h),供试基质为灭菌土和灭菌沙(V∶V=1∶1)混合。
-
采用双因素完全随机区组试验设计:(1)2个AM真菌(C. etuicatum)处理,即接种(+AM)和不接种(−AM);(2)4个无机磷处理水平,即0、10、50、100 KH2PO4 mg·kg−1。有8个处理,每处理10盆,重复3次,每处理累计30盆,共240盆。
将灭菌供试基质装入花盆(上口直径17 cm、下底直径12 cm、高15 cm)中,每盆装2 kg基质,并加入70 g菌剂,对照中加入等量灭活菌剂(121 ℃,灭菌2 h)。每盆种植1棵油茶苗,确保根系与菌剂接触。试验于2019年5月开始,将盆栽放在江西农业大学基地大棚内管理,温度25~28 ℃,相对湿度70%~75%。磷处理前,浇灌无磷的Hoagland营养液,即(236 Ca(NO3)2·4H2O、246.5 MgSO4·7H2O、2.86 H3BO3、1.81 MnCl2·4H2O、0.22 ZnSO4·7H2O、0.08 CuSO4·5H2O、0.02 H2MnO4·H2O、7.354 C10H12O8N2NaFe、74.5 KCl) g·L−1,1个月浇灌1次,每次每盆250 mL营养液。3个月后,开始施磷处理,每隔一天分别浇灌200 mL浓度为0、10、50、100 KH2PO4 mg·kg−1溶液,持续处理30 d,再培养2个月后收获。
-
测定油茶植株地上部和地下部全磷含量,以及土壤全磷、有效磷、有机磷含量。具体测定方法参照《土壤农业化学分析方法》[23]。
-
测定土壤中无机磷含量,土壤无机磷分级的具体测定方法参照雷宏军等[24]。
-
油茶根、叶总RNA的提取方法参照天根生物工程(TIANGEN)公司RNA prep Pure多糖多酚植物总RNA提取试剂盒(DP441)所提供的方法,所得RNA溶液放置−80 ℃备用,引物序列参考周俊琴等[25] 。
-
使用Microsoft Excel 2016对数据进行整理,在IBM SPSS statistics 20.0中采用双因素方差分析(Two-way ANOVA)检测接种AM真菌处理、施无机磷处理及其交互作用对所有测定指标的影响。采用单因素方差分析(One-way ANOVA)中的Duncan’s检验检测不同处理间的差异(P<0.05)。通过皮尔森相关系数(Pearson correlation coefficient)分析油茶苗磷浓度与土壤不同磷形态含量的相关性。使用origin2019绘制柱状图。
-
双因素方差分析表明(表1),接种AM真菌、施无机磷及其两者交互作用对油茶地上部磷浓度含量和根系磷含量影响均为极显著(P<0.01)。
表 1 接种AM真菌、施磷处理及两者交互作用对油茶苗磷含量、培养土壤磷组分含量的影响
Table 1. Effects of AM fungi inoculation, phosphorus application and their interaction on the phosphorus concentration of Camellia oleifera and soil phosphorus concentration
指标
Index接种处理
AMF施磷处理
P接种处理 × 施磷处理
AMF × P地上磷含量 Aboveground phosphorus concentration 155.252** 390.720** 10.375** 根系磷含量 Root phosphorus concentration 501.600** 337.067** 45.867** 土壤全磷含量 Soil total phosphorus concentration 171.322** 117.561** 195.548** 土壤有效磷含量 Soil available phosphorus concentration 14.188** 150.772** 9.516** 土壤有机磷含量 Soil organic phosphorus concentration 3.302** 37.920** 21.514** 磷酸铝盐 Al-P 73.113** 396.468** 14.229** 磷酸铁盐 Fe-P 618.327** 263.515** 199.117** 磷酸钙盐 Ca-P 85.918** 74.857** 4.570** 闭蓄态磷 O-P 86.918* 74.859** 45.570NS 叶片Pht1;1相对表达量 Leaf Pht1;1 relative expression 14.882** 4.217* 19.919** 根系Pht1;1相对表达量 Root system Pht1;1 relative expression 19.900** 8.329** 2.718NS 注:*,在P<0.05水平有显著影响;**,在P<0.01水平有显著影响;NS,无显著影响。下同。
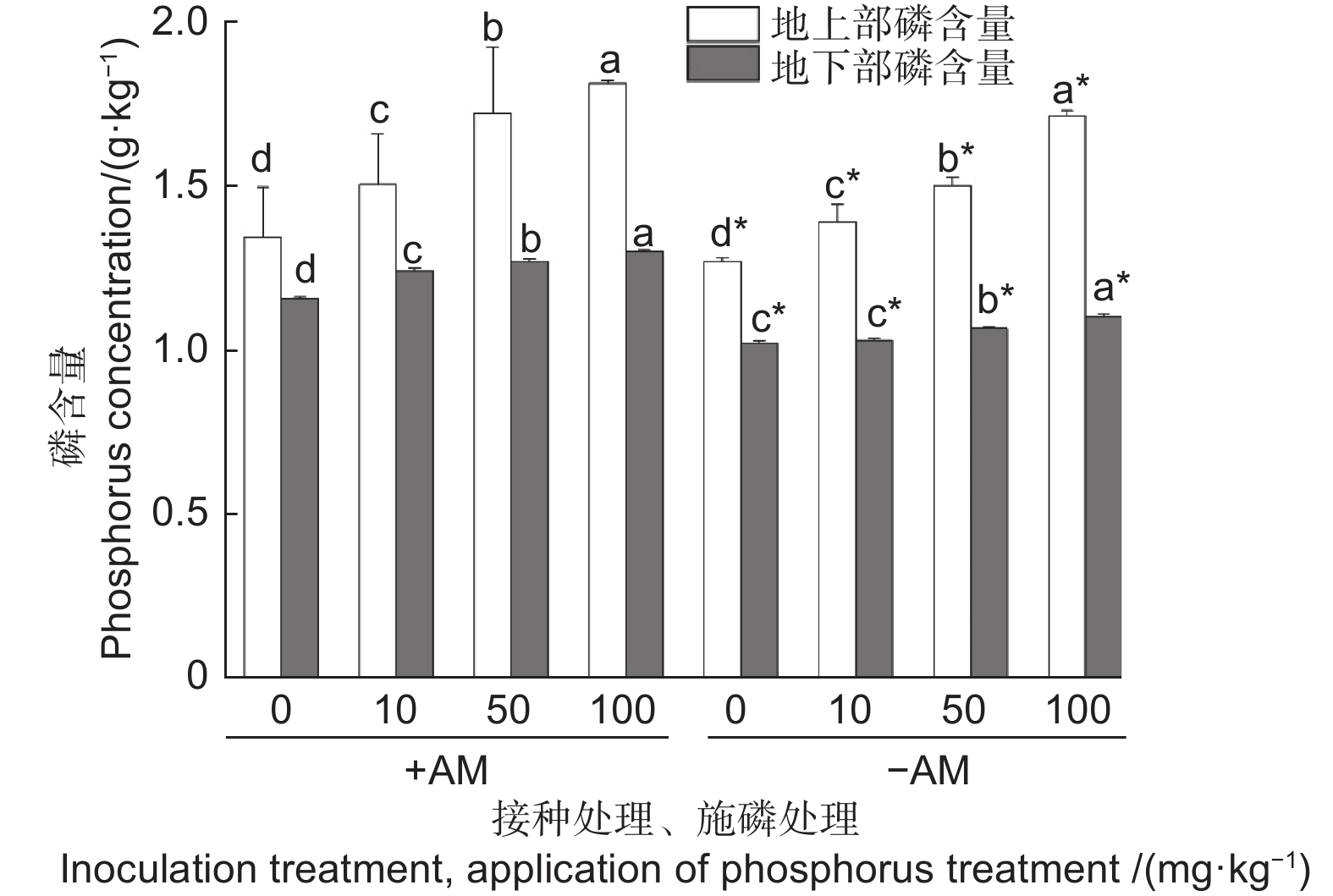
Note: *, significant differences at P<0.05; **, significant differences at P<0.01; NS, no significant differences. The same as below.图1可知,无论是否接种,油茶地上部磷含量随着KH2PO4浓度的增加均呈递增趋势。在不施磷(0 mg·kg−1)、低磷(10 mg·kg−1)、中磷(50 mg·kg−1)和高磷(100 mg·kg−1)条件下,菌根化油茶苗地上部磷含量均显著高于未菌根化的磷含量,分别提高了5.64%、8.14%、14.28%、5.79%。无论是否接种AM真菌,随着KH2PO4浓度的增加,油茶根系磷含量均呈逐步上升的趋势。在4个不同KH2PO4浓度下,接种AM真菌均显著提高油茶根系的磷含量,分别提高了12.85%、20.01%、19.63%、18.09%。

图 1 接种AM真菌及施磷对油茶植株地上部和根系磷含量的影响
Figure 1. Effect of inoculation of AM fungi and application of phosphorus on aboveground and root phosphorus concentration of Camellia oleifera
-
双因素方差分析表明(表1),接种AM真菌处理、施无机磷处理及其两者交互作用对土壤全磷含量、有效磷含量及有机磷含量的影响均极显著(P<0.01)。
表2可知,无论是否接种,随着KH2PO4浓度的增加,土壤全磷含量、有效磷含量和有机磷含量均呈逐步上升趋势。不施磷条件下,接种AM真菌显著降低油茶土壤全磷含量和有机磷含量,分别降低了3.32%、11.40%。在低磷(10 mg·kg−1)条件下,接种AM真菌显著增加了土壤全磷含量(6.17%)和有机磷含量(10.44%),而降低了有效磷含量(2.20%)。在中磷(50 mg·kg−1)条件下,接种AM真菌显著降低了油茶土壤全磷含量(9.43%)和有效磷含量(2.58%),而增加了有机磷含量(0.49%)。在高磷(100 mg·kg−1)条件下,接种AM真菌均显著降低了土壤全磷含量(9.09%)、有效磷含量(6.63%)和有机磷含量(4.44%)。
表 2 接种AM真菌及施磷对土壤磷组分含量的影响
Table 2. Effect of inoculation of AM fungi and application of soil phosphorus concentration of Camellia oleifera seedlings
接种处理
AMF施磷
KH2PO4 /(mg·kg−1)土壤全磷含量
Soil total phosphorus
concentration土壤有效磷含量
Soil available
phosphorus concentration土壤有机磷含量
Soil organic
phosphorus concentration+AM 0 133.20±0.84 d 10.25±0.05 d 14.91±0.52 d 10 155.03±1.75 c 11.25±0.05 c 23.75±0.25 c 50 158.75±0.74 b 14.93±0.07 b 35.68±0.62 b 100 186.82±1.09 a 19.13±0.73 a 45.33±1.04 a −AM 0 137.77±2.10 d* 9.94±0.01 d* 16.83±0.63 d* 10 146.02±1.41 c* 11.50±0.10 c* 21.50±0.25 c* 50 175.28±2.04 d* 15.33±0.13 b* 35.50±0.25 b* 100 205.51±0.73 a* 20.49±0.22 a* 47.43±0.16 a* 注: + AM:接种AM真菌;−AM:不接种AM真菌。数值为均值±标准差(n=3),不同小写字母表示相同接种处理间差异显著(P<0.05);星号表示相同磷水平处理间差异显著(P<0.05)。下同。
Note: + AM: AMF inoculation; −AM: non-inoculation. Values are presented as means ± SD (n=3),different letters indicate significant differences between the same inoculation treatments (P<0.05), asterisks indicate significant differences between treatments with the same phosphorus level (P<0.05). The same as below. -
双因素方差分析表明(表1),接种AM真菌处理对土壤Al-P含量、Fe-P含量、Ca-P含量和O-P含量的影响均极为显著(P<0.01)。施磷对土壤Al-P含量、Fe-P含量、Ca-P含量和O-P含量的影响均极显著(P<0.01)。接种AM真菌处理和施磷处理两者交互作用对土壤中Al-P、Fe-P和O-P 3种磷形态含量的影响均极为显著(P<0.01),而对土壤中Ca-P含量的影响不显著(P>0.05)。
表3可知,未接种条件下,Al-P、Fe-P、Ca-P、O-P 4种磷形态含量均显著高于接种处理,由低到高顺序为O-P<Ca-P<Al-P<Fe-P,其中Ca-P、O-P、Al-P和Fe-P最大值分别是27.99 mg·kg−1、28.50 mg·kg−1、39.46 mg·kg−1、47.21 mg·kg−1。
表 3 接种AM真菌及施磷对不同形态磷含量的影响
Table 3. Effect of inoculation of AM fungi and application on different forms of grown soil phosphorus content of Camellia oleifera seedlings
接种处理
AMF施磷
KH2PO4 mg·kg−1土壤Al-P
Soil Al-P content /(mg·kg−1)土壤Fe-P
Soil Fe-P content /(mg·kg−1)土壤Ca-P
Soil Ca-P Content /(mg·kg−1)土壤O-P
Soil O-P Content /(mg·kg−1)+AM 0 16.78±0.19 d 29.62±0.10 d 13.17±0.18 d 11.49±0.04 c 10 22.09±0.27 c 33.98±0.30 c 17.46±0.35 c 9.55±0.16 d 50 29.10±0.14 b 36.68±0.25 b 24.12±0.51 b 16.98±0.08 b 100 38.75±0.65 a 44.14±0.52 a 26.43±0.74 a 23.86±0.27 a −AM 0 18.71±0.23 d* 28.95±0.04 d* 15.67±0.32 d* 13.17±0.18 c* 10 21.78±0.17 c 36.25±0.61 c* 19.77±0.68 c* 12.81±0.13 c* 50 31.13±0.40 b* 45.51±0.29 b* 24.77±0.81 b 20.27±0.65 b 100 40.03±0.57 a* 47.71±0.25 a* 29.01±0.33 a* 28.86±0.86 a* 无论是否接种AM真菌,随着KH2PO4浓度的增加土壤Al-P、Fe-P和Ca-P含量均呈递增趋势,而土壤O-P含量随着KH2PO4浓度的增加均呈先下降后上升的趋势。不施磷条件下,接种AM真菌降低土壤Al-P含量(10.85%)、Fe-P含量(2.25%)、Ca-P含量(15.92%)和O-P含量(12.82%)。施磷量为10 mg·kg−1时,接种AM真菌与未接种处理间对土壤Al-P含量差异不显著,显著降低了土壤Fe-P含量(6.27%)、Ca-P含量(11.67%)和O-P含量(25.48%)。施磷量为50 mg·kg−1时,接种AM真菌显著降低了土壤Al-P含量(4.90%)和土壤Fe-P含量(19.40%),对Ca-P含量和O-P含量差异不显著。施磷量为100 mg·kg−1时,接种AM真菌与未接种AM真菌处理间土壤Al-P含量差异不显著,显著降低了Fe-P含量、 Ca-P含量和O-P含量,分别为7.48%、9.13%和17.30%。
-
相关性分析表明(表4),土壤有效磷、有机磷和Al-P含量均与油茶根系磷浓度呈显著正相关关系;土壤全磷、有效磷、有机磷、Al-P、Fe-P、Ca-P和O-P含量均与油茶地上磷含量呈正相关关系。土壤有效磷、有机磷、Al-P、Fe-P、Ca-P和O-P含量与土壤全P含量呈显著正相关关系;土壤有机磷、Al-P、Fe-P、Ca-P和O-P含量与土壤有效磷含量呈正相关关系;土壤Al-P、Fe-P、Ca-P和O-P含量与土壤有机磷含量呈正相关关系。
表 4 油茶植株磷含量与土壤不同形态磷含量之间的相关性
Table 4. Correlation between the phosphorus content in Camellia oleifera seedlings and the phosphorus content of different forms in soil
根系磷含量 地上磷含量 全磷 有效磷 有机磷 Al-P Fe-P Ca-P 地上磷含量 0.637** 全磷 0.272 0.843** 有效磷 0.323* 0.895** 0.954** 有机磷 0.371* 0.922** 0.953** 0.988** Al-P 0.331* 0.897** 0.959** 0.985** 0.990** Fe-P 0.110 0.752** 0.931** 0.920** 0.912** 0.918** Ca-P 0.228 0.859** 0.921** 0.959** 0.966** 0.959** 0.938** O-P 0.108 0.757** 0.903** 0.942** 0.922** 0.938** 0.868** 0.904** 注:*,在P<0.05水平有显著性差异;**,在P<0.01水平有显著性差异;NS,无显著性差异。下同。
Note: *, significant differences at P<0.05; **, significant differences at P<0.01; NS, no significant differences. The same as below. -
双因素方差分析表明(表1)接种AM真菌对油茶叶片和根系的Pht1;1基因表达影响均极显著(P<0.01)。施无机磷对油茶叶片Pht1;1基因表达的影响显著(P<0.05),对油茶根系Pht1;1基因表达的影响极显著(P<0.01)。接种处理和施磷处理两者交互作用对油茶叶片的Pht1;1基因表达的影响极显著(P<0.01),而对根系的Pht1;1基因表达的影响不显著(P>0.05)。
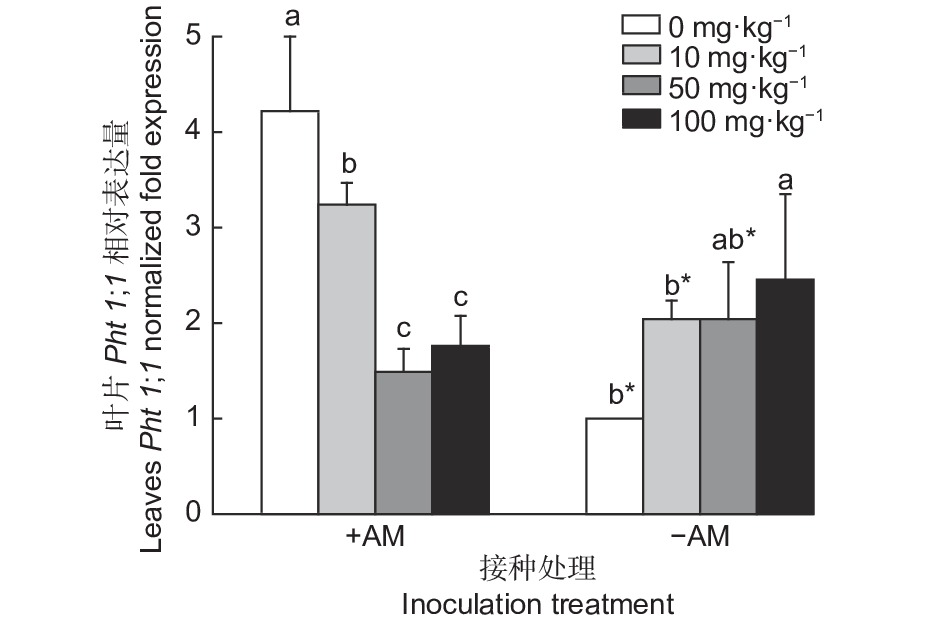
图2所示,接种AM真菌条件下,油茶叶片Pht1;1基因表达量随着供磷水平的增加呈先降后升的趋势;而不接种AM真菌条件下,油茶叶片Pht1;1基因表达量随着供磷水平的增加呈逐步上升的趋势。在不施磷和低磷(10 mg·kg−1)条件下,接种AM真菌均显著提高油茶叶片Pht1;1基因的相对表达量,分别提高了37.14%、39.58%。在中磷(50 mg·kg−1)和高磷(100 mg·kg−1)条件下,接种AM真菌与未接种AM真菌油茶叶片Pht1;1基因的相对表达量之间差异不显著。

图 2 接种AM真菌及施磷下油茶叶片Pht1;1相对表达量
Figure 2. Relative expression level of Pht1;1 in Camellia oleifera seedling leaves inoculated with AM fungi and phosphorus
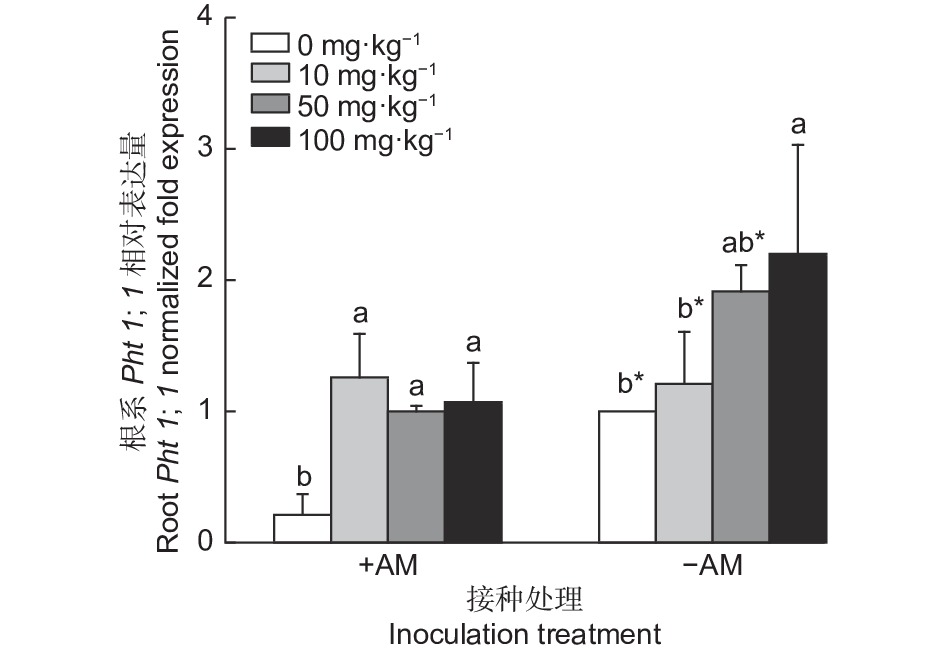
图3所示,接种AM真菌下油茶根系Pht1;1基因表达量随着KH2PO4浓度的增加呈先上升后下降的趋势;而不接种AM真菌下油茶叶片Pht1;1基因表达量随着KH2PO4浓度的增加呈逐步上升的趋势。不施磷条件下,接种AM真菌油茶根系Pht1;1基因表达量显著低于未接种,降低了78.74%。低磷条件下,接种AM真菌显著增加油茶根系Pht1;1基因表达量,增加了3.73%。中磷条件下,接种AM真菌显著降低了油茶根系Pht1;1基因表达量(47.59%)。高磷条件下,接种AM真菌显著降低油茶根系Pht1;1基因表达量(51.38%)。
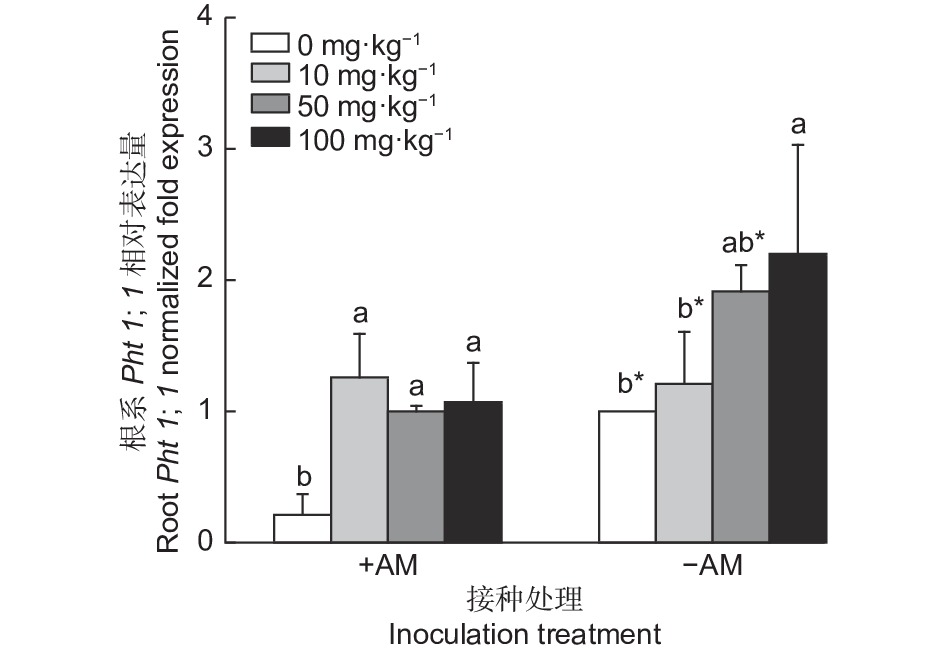
图 3 接种AM真菌及施磷下油茶根系Pht1;1相对表达量
Figure 3. Relative expression of Pht1;1 in the roots of Camellia oleifera seedling inoculated with AM fungi and phosphorus
-
磷素极易被固定于土壤中而使其有效性降低,土壤磷胁迫极大程度地限制植物的生长和产量[13,26]。本研究结果表明,低磷浓度下,接种AM真菌显著提高了油茶地上部磷含量和根系磷含量,且施加无机磷和AM真菌存在显著的交互作用。这可能是低磷环境下,油茶更依赖AM真菌吸收土壤中的养分和水分,同时根外菌丝分泌苹果酸等有机酸,溶解土壤中难溶性磷化合物,提高土壤磷素的活性,从而改善油茶植株的磷含量[27],与刘春艳[28] 、孙艳梅[29]、李芳[30]等试验结果一致。
土壤中磷素的形态和含量是其有效性的关键因子[26]。根据雷宏军等[24]提出的酸性土壤无机磷分级方法,可分成Al-P、Fe-P、Ca-P和O-P。其中Al-P、Fe-P、Ca-P是植物的第二有效磷源,而O-P是难以被植物吸收利用的潜在磷源[31] 。钟熊等[32]对菌丝室红壤中添加不同形态磷处理,发现接种丛枝菌根真菌一定程度上增加了Ca2-P、Al-P、O-Al、和Ca10-P的含量。此外,张丽[33]、张宇亭[34]分别在红壤和石灰性土壤研究发现,AM真菌主要活化土壤中潜在磷源(如Ca10-P、O-P),进而提高土壤有效性。本试验结果表明,接种AM真菌显著降低了土壤Al-P、Fe-P、Ca-P和O-P的含量,这与张宇亭[34]等的结论一致。这可能是由于AM真菌与油茶建立共生关系后,AM真菌自身分泌有机酸等或促进油茶根系分泌有机酸(如柠檬酸、葡萄酸等),促进了土壤磷从植物难以利用的形态向可利用形态转变[35]。因此接种C. etuicatum可有效的促进油茶土壤磷形态的变化以及对矿质养分的吸收,促进土壤磷素向着有利于宿主植物吸收的方向转化。
植物磷的吸收是跨表皮及皮层细胞质膜的逆浓度的主动运输过程,依赖于根系细胞膜上的高亲和磷转运蛋白家族。舒波[36]研究表明枳实生苗根系中的磷转运蛋白基因表达受AM真菌侵染和土壤磷水平的调控,菌根侵染使PtaPT1和PtaPT2的相对表达量降低了20%~80%,而PtaPT3和PtaPT7的相对表达量降低了10%~30%,且低磷胁迫提高了出PtaPT6之外的其它Pht1基因家族成员的表达量;刘芳[37]在玉米全基因组水平上鉴定出13个Pht1磷转运基因,其中8个为新成员,且低磷胁迫下6个新成员基因受AM真菌诱导表达。此外,有研究表明AM真菌不仅可以通过诱导磷转运蛋白基因的表达来提高宿主植物对磷素的吸收和利用,在低磷胁迫下也能够改变植物根系基因型的表达[38];周俊琴等[25]发现了油茶湘林4号(Camellia oleifera ‘Xianglin 4’)在低磷条件下诱导Pht1;1的表达。本试验结果表明,未接种AM真菌油茶叶片Pht1;1基因表达量随着KH2PO4浓度的升高而递增,这可能是随着施磷浓度的升高,转运至叶片中的磷量增加的结果;低磷条件下,接种C. etuicatum显著增加了油茶叶片和根系中Pht1;1基因的表达;中磷、高磷条件下,接种C. etuicatum降低了油茶叶片和根系中Pht1;1基因的表达,表明AM真菌可通过调控油茶体内PO43−的运输过程,从而影响油茶对外界不同磷浓度的响应。Pht1;1基因只是油茶Pht1基因家族成员中一员,其基因家族成员的具体数量及各成员之间相互调控方式和机制并未清楚,需要在进一步对其家族成员进行克隆、表达模式和调控方式等进行研究,探索转运子在油茶根系中如何进行表达。
-
本研究研究了接种AM真菌和施无机磷对油茶磷吸收和土壤磷组分的影响,结果表明,接种C. etuicatum可将土壤中难溶性磷转化成为植物可利用的磷形态,并提高油茶植株对土壤中有效磷的吸收效率。同时,在低磷条件下,接种C. etuicatum显著增加了油茶叶片和根系中Pht1;1基因的表达,在中磷、高磷条件下,接种C. etuicatum降低了油茶叶片和根系中Pht1;1基因的表达。
AM真菌和无机磷对油茶苗磷吸收和培养土壤磷组分的影响
Effects of AM Fungi and Inorganic Phosphorus on Phosphorus Uptake and Grown Soil Phosphorus Fraction of Camellia oleifera Seedlings
-
摘要:
目的 探究不同无机磷水平下丛枝菌根真菌对油茶苗磷素吸收的促进作用,为接种丛枝菌根真菌提高油茶磷效率的研究提供理论依据。 方法 以1年生油茶苗为宿主植物,接种幼套近明球囊霉(Claroideogolmus etuicatum)和施KH2PO4 (0、10、50、100 mg·kg−1),探究接种AM真菌和无机磷对油茶磷吸收和土壤磷组分的影响。 结果 在不施磷(0 mg·kg−1)、低磷(10 mg·kg−1)、中磷(50 mg·kg−1)和高磷(100 mg·kg−1)条件下,菌根化油茶苗磷含量均显著高于未菌根化,地上部分别提高了5.64%、8.14%、14.28%、5.79%,根系磷含量分别提高了12.85%、20.01%、19.63%、18.09%。接种AM真菌处理、施加无机磷处理及其两者交互作用下对土壤全磷含量、有效磷含量以及有机磷含量的影响均极显著。接种幼套近明球囊霉极显著降低了土壤中Al-P、Fe-P、Ca-P、O-P 4种磷形态含量。在低磷条件下,接种幼套近明球囊霉显著增加了油茶苗叶片和根系中Pht1;1基因的表达;在中磷、高磷条件下,接种幼套近明球囊霉则降低了油茶叶片和根系中Pht1;1基因的表达。 结论 AM真菌可改变油茶苗土壤中磷形态,促进植株对土壤中有效磷的吸收效率,并参与调控油茶植株中Pht1;1基因的表达。该试验结果为微生物菌肥提升油茶苗对土壤磷素吸收效率和减少磷肥施入量提供科学依据,同时促进我国油茶产业持续健康发展。 Abstract:Objective To investigate the promotion of phosphorus uptake in Camellia oleifera seedlings by mycorrhizal fungi at different inorganic phosphorus levels, and provide a theoretical basis for the study of inoculation with mycorrhizal fungi to enhance phosphorus efficiency of C. oleifera seedlings. Method One-year old C. oleifera was used as the host plant, and young sets of Claroideogolmus etuicatum were inoculated with KH2PO4 (0, 10, 50, 100 mg·kg−1). The effects of inoculation with AM fungi and inorganic phosphorus on phosphorus uptake and grown soil phosphorus fraction of C. oleifera seedlings were investigated. Result Under the conditions of no (0 mg·kg−1), low (10 mg·kg−1), medium (50 mg·kg−1) and high (100 mg·kg−1) phosphorus application, the phosphorus content of mycorrhization C. oleifera seedlings was significantly higher than that of non- mycorrhization. The above-ground phosphorus content increased by 5.64%, 8.14%, 14.28% and 5.79%, respectively, and the phosphorus content of seedling root system increased by 12.85%, 20.01%, 19.63% and 18.09%, respectively. The effects of inoculation with AM fungal treatment, application of inorganic phosphorus treatment and their interaction on soil total phosphorus content, effective phosphorus content and organic phosphorus content were highly significant. Inoculation with C. etuicatum significantly reduced the content of four phosphorus forms, Al-P, Fe-P, Ca-P and O-P in the soil. Under low phosphorus conditions, inoculation of C. etuicatum significantly increased the expression of Pht1;1 gene in leaves and root systems of the seedlings; under medium and high phosphorus conditions, inoculation of C. etuicatum decreased the gene expression of Pht1;1 gene in leaves and root of C. oleifera seedlings. Conclusion AM fungi can alter phosphorus morphology in C. oleifera grown soil, promote the uptake efficiency of effective phosphorus in soil by plants, and participate in regulating the expression of Pht1;1 gene in C. oleifera. The results of this study provide a scientific basis for microbial fertilizer to improve soil phosphorus uptake efficiency of C. oleifera seedlings and reduce phosphorus fertilizer application, which promotes the healthy and sustainable development of C. oleifera industry in China. -
图 1 接种AM真菌及施磷对油茶植株地上部和根系磷含量的影响
Figure 1. Effect of inoculation of AM fungi and application of phosphorus on aboveground and root phosphorus concentration of Camellia oleifera
图 2 接种AM真菌及施磷下油茶叶片Pht1;1相对表达量
Figure 2. Relative expression level of Pht1;1 in Camellia oleifera seedling leaves inoculated with AM fungi and phosphorus
图 3 接种AM真菌及施磷下油茶根系Pht1;1相对表达量
Figure 3. Relative expression of Pht1;1 in the roots of Camellia oleifera seedling inoculated with AM fungi and phosphorus
表 1 接种AM真菌、施磷处理及两者交互作用对油茶苗磷含量、培养土壤磷组分含量的影响
Table 1. Effects of AM fungi inoculation, phosphorus application and their interaction on the phosphorus concentration of Camellia oleifera and soil phosphorus concentration
指标
Index接种处理
AMF施磷处理
P接种处理 × 施磷处理
AMF × P地上磷含量 Aboveground phosphorus concentration 155.252** 390.720** 10.375** 根系磷含量 Root phosphorus concentration 501.600** 337.067** 45.867** 土壤全磷含量 Soil total phosphorus concentration 171.322** 117.561** 195.548** 土壤有效磷含量 Soil available phosphorus concentration 14.188** 150.772** 9.516** 土壤有机磷含量 Soil organic phosphorus concentration 3.302** 37.920** 21.514** 磷酸铝盐 Al-P 73.113** 396.468** 14.229** 磷酸铁盐 Fe-P 618.327** 263.515** 199.117** 磷酸钙盐 Ca-P 85.918** 74.857** 4.570** 闭蓄态磷 O-P 86.918* 74.859** 45.570NS 叶片Pht1;1相对表达量 Leaf Pht1;1 relative expression 14.882** 4.217* 19.919** 根系Pht1;1相对表达量 Root system Pht1;1 relative expression 19.900** 8.329** 2.718NS 注:*,在P<0.05水平有显著影响;**,在P<0.01水平有显著影响;NS,无显著影响。下同。
Note: *, significant differences at P<0.05; **, significant differences at P<0.01; NS, no significant differences. The same as below. 下载: 导出CSV
下载: 导出CSV
表 2 接种AM真菌及施磷对土壤磷组分含量的影响
Table 2. Effect of inoculation of AM fungi and application of soil phosphorus concentration of Camellia oleifera seedlings
接种处理
AMF施磷
KH2PO4 /(mg·kg−1)土壤全磷含量
Soil total phosphorus
concentration土壤有效磷含量
Soil available
phosphorus concentration土壤有机磷含量
Soil organic
phosphorus concentration+AM 0 133.20±0.84 d 10.25±0.05 d 14.91±0.52 d 10 155.03±1.75 c 11.25±0.05 c 23.75±0.25 c 50 158.75±0.74 b 14.93±0.07 b 35.68±0.62 b 100 186.82±1.09 a 19.13±0.73 a 45.33±1.04 a −AM 0 137.77±2.10 d* 9.94±0.01 d* 16.83±0.63 d* 10 146.02±1.41 c* 11.50±0.10 c* 21.50±0.25 c* 50 175.28±2.04 d* 15.33±0.13 b* 35.50±0.25 b* 100 205.51±0.73 a* 20.49±0.22 a* 47.43±0.16 a* 注: + AM:接种AM真菌;−AM:不接种AM真菌。数值为均值±标准差(n=3),不同小写字母表示相同接种处理间差异显著(P<0.05);星号表示相同磷水平处理间差异显著(P<0.05)。下同。
Note: + AM: AMF inoculation; −AM: non-inoculation. Values are presented as means ± SD (n=3),different letters indicate significant differences between the same inoculation treatments (P<0.05), asterisks indicate significant differences between treatments with the same phosphorus level (P<0.05). The same as below.
下载: 导出CSV
表 3 接种AM真菌及施磷对不同形态磷含量的影响
Table 3. Effect of inoculation of AM fungi and application on different forms of grown soil phosphorus content of Camellia oleifera seedlings
接种处理
AMF施磷
KH2PO4 mg·kg−1土壤Al-P
Soil Al-P content /(mg·kg−1)土壤Fe-P
Soil Fe-P content /(mg·kg−1)土壤Ca-P
Soil Ca-P Content /(mg·kg−1)土壤O-P
Soil O-P Content /(mg·kg−1)+AM 0 16.78±0.19 d 29.62±0.10 d 13.17±0.18 d 11.49±0.04 c 10 22.09±0.27 c 33.98±0.30 c 17.46±0.35 c 9.55±0.16 d 50 29.10±0.14 b 36.68±0.25 b 24.12±0.51 b 16.98±0.08 b 100 38.75±0.65 a 44.14±0.52 a 26.43±0.74 a 23.86±0.27 a −AM 0 18.71±0.23 d* 28.95±0.04 d* 15.67±0.32 d* 13.17±0.18 c* 10 21.78±0.17 c 36.25±0.61 c* 19.77±0.68 c* 12.81±0.13 c* 50 31.13±0.40 b* 45.51±0.29 b* 24.77±0.81 b 20.27±0.65 b 100 40.03±0.57 a* 47.71±0.25 a* 29.01±0.33 a* 28.86±0.86 a*
下载: 导出CSV
表 4 油茶植株磷含量与土壤不同形态磷含量之间的相关性
Table 4. Correlation between the phosphorus content in Camellia oleifera seedlings and the phosphorus content of different forms in soil
根系磷含量 地上磷含量 全磷 有效磷 有机磷 Al-P Fe-P Ca-P 地上磷含量 0.637** 全磷 0.272 0.843** 有效磷 0.323* 0.895** 0.954** 有机磷 0.371* 0.922** 0.953** 0.988** Al-P 0.331* 0.897** 0.959** 0.985** 0.990** Fe-P 0.110 0.752** 0.931** 0.920** 0.912** 0.918** Ca-P 0.228 0.859** 0.921** 0.959** 0.966** 0.959** 0.938** O-P 0.108 0.757** 0.903** 0.942** 0.922** 0.938** 0.868** 0.904** 注:*,在P<0.05水平有显著性差异;**,在P<0.01水平有显著性差异;NS,无显著性差异。下同。
Note: *, significant differences at P<0.05; **, significant differences at P<0.01; NS, no significant differences. The same as below.
下载: 导出CSV
-
[1] 罗 帅, 钟秋平, 葛晓宁, 等. 不同氮、磷、钾施肥配比对油茶花芽分化的影响[J]. 林业科学研究, 2019, 32(2):131-138. doi: 10.13275/j.cnki.lykxyj.2019.02.019 [2] 江 南, 谭晓风, 张 琳, 等. 基于RNA-Seq的油茶种子α-亚麻酸代谢途径及相关基因分析[J]. 林业科学, 2014, 50(8):69-75. [3] 王汉中, 殷 艳. 我国油料产业形势分析与发展对策建议[J]. 中国油料作物学报, 2014, 36(3):414-421. doi: 10.7505/j.issn.1007-9084.2014.03.020 [4] 秦声远, 戎 俊, 张文驹, 等. 油茶栽培历史与长江流域油茶遗传资源[J]. 生物多样性, 2018, 26(4):61-72. doi: 10.17520/biods.2017254 [5] Martin M H, Marschner H. Mineral nutrition of higher plants[J]. Journal of Ecology, 1988, 76(4): 1250. [6] Lambers H, Finnegan P M, Laliberte E, et al. Update on phosphorus nutrition in Proteaceae. Phosphorus nutrition of proteaceae in severely phosphorus-impoverished soils: are there lessons to be learned for future crops?[J]. Plant Physiology, 2011, 156(3): 58-66. [7] Stewart J D, Abidine A E, Bernier P Y. Stomatal and mesophyll limitations of photosynthesis in black spruce seedlings during multiple cycles of drought[J]. Tree Physiology, 1994, 15(1): 57-64. [8] 龚丽娜, 胡冬南, 张文元, 等. 土壤养分状况系统研究法在油茶林地养分管理上的应用[J]. 经济林研究, 2015, 107(4):71-75. [9] 陈家法, 陈隆升, 涂 佳, 等. 长期施肥对油茶林产果量及土壤地力可持续性的影响[J]. 中南林业科技大学学报, 2017, 37(7):59-65. doi: 10.14067/j.cnki.1673-923x.2017.07.009 [10] 罗汉东. 不同磷水平施肥对油茶生长及土壤环境动态影响[D]. 南昌: 江西农业大学, 2017. [11] Smith S E, Read D J. Mycorrhizal symbiosis[J]. Quarterly Review of Biology, 2008, 3(3): 273-281. [12] Xie X, Weng B, Cai B, et al. Effects of arbuscular mycorrhizal inoculation and phosphorus supply on the growth and nutrient uptake of Kandelia obovata (Sheue, Liu & Yong) seedlings in autoclaved soil[J]. Applied Soil Ecology, 2014, 75: 162-171. doi: 10.1016/j.apsoil.2013.11.009 [13] Rakshit A, Bhadoria P S. Influence of arbuscular mycorrhizal hyphal length on simulation of P influx with the mechanistic model[J]. African Journal of Microbiology Research, 2009, 3(1): 1-4. [14] Yao Q, Wang L R, Zhu H H, et al. Effect of arbuscular mycorrhizal fungal inoculation on root system architecture of trifoliate orange (Poncirus trifoliata L. Raf. ) seedlings[J]. Scientia Horticulturae, 2009, 121(4): 458-461. doi: 10.1016/j.scienta.2009.03.013 [15] Smith S E, Jakobsen S I. Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake[J]. New Phytologist, 2004, 162(2): 511-524. doi: 10.1111/j.1469-8137.2004.01039.x [16] 韦莉莉, 卢昌熠, 丁 晶, 等. 丛枝菌根真菌参与下植物-土壤系统的养分交流及调控[J]. 生态学报, 2016, 36(14):4233-4243. [17] 金 鑫, 曾新颖, 齐昌国, 等. 供磷水平对玉米丛枝菌根侵染及其对异质养分吸收的影响[J]. 植物营养与肥料学报, 2018, 118(1):167-173. [18] Schnepf A, Jones D, Roose T. Modelling nutrient uptake by individual hyphae of arbuscular mycorrhizal fungi: temporal and spatial scales for an experimental design[J]. Bull Math Biol, 2011, 73(9): 2175-2200. doi: 10.1007/s11538-010-9617-1 [19] Duan T, Facelli E, Smith S E, et al. Differential effects of soil disturbance and plant residue retention on function of arbuscular mycorrhizal (AM) symbiosis are not reflected in colonization of roots or hyphal development in soil[J]. Soil Biol Biochem, 2011, 43(3): 571-578. doi: 10.1016/j.soilbio.2010.11.024 [20] Balzergue C, Puech-Pagès V, Bécard G, et al. The regulation of arbuscular mycorrhizal symbiosis by phosphate in pea involves early and systemic signalling events[J]. Journal of Experimental Botany, 2011, 62(3): 1049-1060. doi: 10.1093/jxb/erq335 [21] Nazeri N K, Lambers H, Tibbett M, et al. Do arbuscular mycorrhizas or heterotrophic soil microbes contribute toward plant acquisition of a pulse of mineral phosphate[J]. Plant & Soil, 2013, 373(12): 699-710. [22] 林宇岚, 李正昀, 张林平, 等. 有机磷和AM真菌对油茶生长、根系形态和光合作用的影响[J]. 经济林研究, 2021, 39(1):121-128 + 210. doi: 10.14067/j.cnki.1003-8981.2021.01.014 [23] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科学技术出版社, 2000. [24] 雷宏军, 刘 鑫, 朱端卫. 酸性土壤磷分级新方法建立与生物学评价[J]. 土壤学报, 2007, 44(5):860-866. doi: 10.3321/j.issn:0564-3929.2007.05.013 [25] 周俊琴, 谭晓风, 袁 军, 等. 油茶Pht1;1基因克隆及其表达分析[J]. 植物遗传资源学报, 2013, 14(3):512-517. [26] 薛英龙, 李春越, 王苁蓉, 等. 丛枝菌根真菌促进植物摄取土壤磷的作用机制[J]. 水土保持学报, 2019, 33(6):10-20. doi: 10.13870/j.cnki.stbcxb.2019.06.002 [27] Cavagnaro T R. Impacts of compost application on the formation and functioning of arbuscular mycorrhizas[J]. Soil Biology and Biochemistry, 2014, 78: 38-44. doi: 10.1016/j.soilbio.2014.07.007 [28] 刘春艳, 吴强盛, 邹英宁. AM真菌对枳吸收磷和分泌磷酸酶的影响[J]. 菌物学报, 2017, 36(7):942-949. doi: 10.13346/j.mycosystema.170069 [29] 孙艳梅, 张前兵, 苗晓茸, 等. 解磷细菌和丛枝菌根真菌对紫花苜蓿生产性能及地下生物量的影响[J]. 中国农业科学, 2019, 52(13):2230-2242. doi: 10.3864/j.issn.0578-1752.2019.13.004 [30] 李 芳, 郝志鹏, 陈保冬. 菌根植物适应低磷胁迫的分子机制[J]. 植物营养与肥料学报, 2019, 25(11):1989-1997. doi: 10.11674/zwyf.18490 [31] Xiang W, Huang M, Li X. Progress on fractioning of soil phosphorous and availability of various phosphorous fractions to crops in soil[J]. Plant Nutr. Fert. Sci, 2004, 10: 663-670. [32] 钟 雄, 王 硕, 包 立, 等. 间作作物菌根菌丝对红壤磷形态的影响[J]. 中国生态农业学报, 2018, 26(11):1624-1633. [33] 张 丽, 柳 勇, 谷林静, 等. 外源磷与AMF对间作玉米种植红壤无机磷形态的影响[J]. 中国土壤与肥料, 2016(1):26-33. doi: 10.11838/sfsc.20160105 [34] 张宇亭, 朱 敏, 线岩相洼, 等. 接种AM真菌对玉米和油菜种间竞争及土壤无机磷组分的影响[J]. 生态学报, 2012, 32(22):11. [35] 刘润进. 菌根学[M]. 北京: 科学出版社, 2007. [36] 舒波. 丛枝菌根真菌促进枳(Poncirus trifoliata L. Raf)磷吸收效应及其机理研究[D]. 武汉: 华中农业大学, 2013. [37] 刘芳. 丛枝菌根真菌诱导ZmPHT1;9促进玉米对磷吸收的分子机制研究[D]. 合肥: 安徽农业大学, 2017. [38] 周德贵, 周少川, 王重荣, 等. 植物磷利用研究在水稻分子设计育种中的应用[J]. 分子植物育种, 2018, 16(16):236-246. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3618
- HTML全文浏览量: 1709
- PDF下载量: 99
- 被引次数: 0
