

-
碳水化合物(Carbohydrate)是植物形态构建和能量代谢的主要能量物质[1],根据其存在形式和功能的不同,一般分为结构性碳水化合物(Structural carbohydrate,SC)和非结构性碳水化合物(Non-structural carbohydrate,NSC)两大类。其中SC主要成分是木质素、纤维素和果胶等高分子化合物,主要参与植物体结构与形态构建,在植物体内无法移动;NSC主要成分是可溶性糖(如果糖、葡萄糖和蔗糖等)和淀粉,主要参与植物的代谢过程并为其提供能源物质,在植物体内可以移动、相互转化[2]。研究表明,NSC中的可溶性糖在植物的抗寒性能上起着重要作用[3-4],而淀粉则作为植物组织中长期的能量储存物质,是植物贮藏器官中最主要的碳水化合物存在形式之一。NSC含量的变化能较好地反映植物体对外界环境变化作出的响应与适应策略[5]。藏东南地区是我国面积最大的原生林区之一,急尖长苞冷杉(Abies georgei var. smithii (Viguié & Gaussen) W. C. Cheng & L. K. Fu
)系松科冷杉亚科冷杉属植物,是中国特有种之一,在藏东南原生林区海拔2 500~4 500 m的高山地带广泛分布,在低海拔常与其它树种混交成林,中海拔适生区单独成林,高海拔则是主要的林线树种之一[6-7],并在藏东南形成了全球海拔最高的林线之一[8]。本研究通过对不同季节、不同海拔、不同器官的急尖长苞冷杉取样和分析,旨在回答和探索如下问题:1)季节、海拔和器官对NSC及其组分时空分布特征的影响;2)植物C、N、P含量及其化学计量对NSC及其组分时空分布特征的影响;3)急尖长苞冷杉林线是否存在“碳平衡失调”[9]或是“生长抑制”[10]。 -
本研究地位于西藏工布自然保护区林芝市境内的色季拉山。色季拉山属念青唐古拉山向南延伸的余脉,地理坐标29°10′~30°15′ N,93°12′~95°35′ E,海拔2 200~5 400 m,为湿润山地暖温带、半湿润山地温带气候。在色季拉山,急尖长苞冷杉、林芝云杉(Picea linzhiensis (W. C. Cheng & L.K. Fu) Rushforth
)和川滇高山栎(Quercus aquifolioides Rehd. et Wils.)是主要乔木层组成树种,是构成群落外貌的主要建群种[11-12]。样地基本信息如表1所示。 表 1 急尖长苞冷杉样地基本信息
Table 1. Basic information of Abies georgei var. smithii forest sample points
海拔/m
Altitude经纬度
Latitude and
longitude土壤
类型
Soil type坡度/°
Slope坡向/°
Aspect坡位
Slope positions郁闭度
Crown density树高
Tree height/m胸径/cm
DBH冠幅(东西)/m
Canopy
(E-W)冠幅(南北)/m
Canopy
(S-N)年均温/℃
Annual average
temperature年降水/mm
Annual precipitation3 500
(3 530)94°43′31″E;
29°41′53″N酸性暗
棕壤22 25 下 0.90 21.3 ± 1.6 37.4 ± 17.3 6.0 ± 1.9 5.6 ± 1.6 4.67 826.6 3 900
(3 870)94°36′12″E;
29°35′34″N酸性暗
棕壤27 155 中 0.85 27.8 ± 1.3 46.0 ± 15.5 5.7 ± 1.9 5.8 ± 2.0 3.61 867.1 4 300
(4 330)94°41′42″E;
29°36′52″N酸性暗
棕壤25 55 上 0.70 10.7 ± 1.2 34.6 ± 13.3 4.8 ± 1.1 5.1 ± 1.0 0.56 966.3 注:年均温、年降水为样地自动气象站数据
Note: Annual average temperature and annual precipitation are the data of automatic weather stations in the sample area. -
以色季拉山建群种急尖长苞冷杉为研究对象,在其低海拔针阔叶混交林(3 500 m)、中海拔纯林(3 900 m)和高海拔林线过渡带(4 300 m)分别选取具有典型性、代表性的急尖长苞冷杉林,设置一个20 m × 20 m的样方,共计3个样方,测量样方内所有该树种的胸径和树高,并根据调查结果选择5株长势良好的平均标准木作为样树,于2019年7月初(生长季)和2019年11月底(非生长季)分别采取样树的叶、枝、干和根作为样本,测定各器官的NSC及其组分含量,同时测定其C、N、P含量。在生长季和非生长季各采集样品120份,共计样品240份,各器官样品采集方法如下:
(1)枝叶的采集:用高枝剪采取冠层中部东西南北方向的枝叶,现场摘取新枝和1~2年生树枝,所有枝叶混合后分选出枝和叶的样品。
(2)树干的采集:用内径5 mm生长锥在每株样树树干的胸高(离地面1.3 m)位置钻取深至髓心的树芯作为树干样品。
(3)树根的采集:采用挖掘法围绕样树四周在5~30 cm深度的土层获取树根,洗净后选取细根(直径<2 mm)作为树根样品。
将采集到的样品装进带有冰盒的保温箱里保存,及时运回实验室处理。样品参照《植物生理学实验指导》[13-14],在烘箱内105 ℃杀青15 min,然后80 ℃烘干至恒质量,粉碎后过筛,装在自封袋中,存放在带有干燥剂的箱子里待测。
-
采用蒽酮比色法[13-14]测定急尖长苞冷杉叶、枝、干和根等器官可溶性总糖和淀粉的含量,采用元素分析仪(vario EL cube,Germany)测定植物C、N含量,采用电感耦合等离子体发射光谱仪(Thermo iCAP 6300,USA)测定植物P含量[15]。
植物中NSC的成分有90%以上是可溶性糖和淀粉,所以本研究用可溶性糖和淀粉的含量之和来代表NSC含量。本研究的急尖长苞冷杉总体NSC、总体可溶性糖及总体淀粉含量为各器官含量之和的算术平均值。树木NSC的“源”主要指具有同化作用的器官—叶,“汇”主要指非同化器官—枝、干和根[16-17],汇含量用枝、干和根含量之和的平均值表示。本研究NSC及其组分含量用百分比干质量表示[18],1 mg·g−1 = 0.1%。
-
采用SPSS 24软件对数据进行分析,采用Origin 2022软件作图。
为了避免不同生长季节复杂环境因子对实验结果的干扰,通过方差分析比较海拔和器官及其交互作用对急尖长苞冷杉NSC及其组分含量的影响;单因素方差分析和最小差异显著法(LSD)分析不同海拔各器官NSC及其组分含量的差异显著性水平,显著水平为P = 0.05。用冗余分析(RDA)分析海拔、植物器官、C、N、P含量及化学计量比与NSC的相关性。
-
由表2可知,在不同生长季节,器官、海拔梯度及其交互作用都显著地影响急尖长苞冷杉(简称:冷杉)可溶性糖和NSC的含量(P<0.05)。在生长季,器官、海拔梯度及其交互作用显著影响地影响了淀粉的含量,但对可溶性糖和淀粉的比值影响不显著(P>0.05)。而在非生长季,淀粉的含量受到器官、器官和海拔梯度交互作用的显著影响;器官对可溶性糖和淀粉的比值影响显著。
表 2 不同生长季节急尖长苞冷杉NSC含量、可溶性糖和淀粉含量及其比值影响因子方差分析
Table 2. Variance analysis of non-structural carbohydrate content, soluble sugar and starch content and ratio of soluble sugar and starch in different growth seasons of Abies georgei var. smithii
因变量
Dependent variable变异来源
Sources of variation生长季
The growing season非生长季
The non-growing season自由度
dfF P 自由度
dfF P 可溶性糖
Soluble sugar器官
Organ3 118.98 0.000 3 204.666 0.000 海拔
Altitude2 5.882 0.005 2 3.594 0.035 器官 × 海拔
Organ × Altitude6 6.023 0.000 6 3.775 0.004 淀粉
Starch器官
Organ3 236.366 0.000 3 113.284 0.000 海拔
Altitude2 3.547 0.037 2 3.097 0.054 器官 × 海拔
Organ × Altitude6 3.011 0.014 6 4.013 0.002 NSC 器官
Organ3 210.267 0.000 3 86.804 0.000 海拔
Altitude2 4.851 0.012 2 4.359 0.018 器官 × 海拔
Organ × Altitude6 5.114 0.000 6 4.665 0.001 可溶性糖和淀粉的比值
Soluble sugar/ Starch器官
Organ3 1.657 0.189 3 72.161 0.000 海拔
Altitude2 3.163 0.051 2 1.231 0.301 器官 × 海拔
Organ × Altitude6 2.206 0.058 6 1.054 0.403 -
由表3可知,生长季冷杉NSC含量中海拔显著>高海拔、低海拔子集;非生长季冷杉NSC含量随海拔升高呈增加趋势,中海拔、高海拔子集显著>高海拔、低海拔子集。非生长季冷杉NSC、源、汇含量均显著高于生长季,非生长季高海拔、中海拔和低海拔NSC含量分别是生长季的3.23、2.89和3.00倍;源含量是生长季的2.32、2.26和2.07倍;汇含量分别是生长季的3.69、3.12和3.52倍。生长季冷杉在低海拔NSC源含量最高,汇含量最低;中海拔NSC源含量最低,汇含量显著高于低海拔与高海拔;不同生长季节冷杉NSC源含量在各个海拔之间差异不显著。不同生长季冷杉NSC源-汇比表现一致:低海拔>高海拔>中海拔;生长季低海拔、高海拔冷杉NSC源-汇比显著高于中海拔,而非生长季各海拔之间NSC源-汇比差异不显著。生长季冷杉NSC源-汇比均大于1;而非生长季源-汇比在海拔上差异不显著且均小于1,但源含量与汇含量之间无显著差异。
表 3 不同海拔急尖长苞冷杉NSC及源-汇含量
Table 3. Non-structural carbohydrates and source-sink content of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
AltitudeNSC含量/%
NSC concentrationNSC源/%
NSC sourceNSC汇/%
NSC sink源/汇
Source/Sink生长季
The growing season3 500 3.06 ± 0.34bB 4.42 ± 0.20aB 2.61 ± 0.07bB 1.69 ± 0.10aB 3 900 3.50 ± 043aB 3.95 ± 0.11aB 3.35 ± 0.17aB 1.18 ± 0.07bB 4 300 3.12 ± 0.37bB 4.19 ± 0.08aB 2.76 ± 0.14bB 1.52 ± 0.11aB 非生长季
The non-growing season3 500 9.17 ± 0.43bA 9.13 ± 0.48aA 9.18 ± 0.31bA 0.99 ± 0.05aA 3 900 10.08 ± 0.71aA 8.95 ± 0.36aA 10.46 ± 0.30aA 0.86 ± 0.05aA 4 300 10.07 ± 0.64aA 9.72 ± 0.23aA 10.18 ± 0.32aA 0.95 ± 0.04aA 注:不同小写字母表示不同海拔之间的差异显著(最小差异显著法(LSD),P<0.05),不同大写字母表示季节间的差异显著。
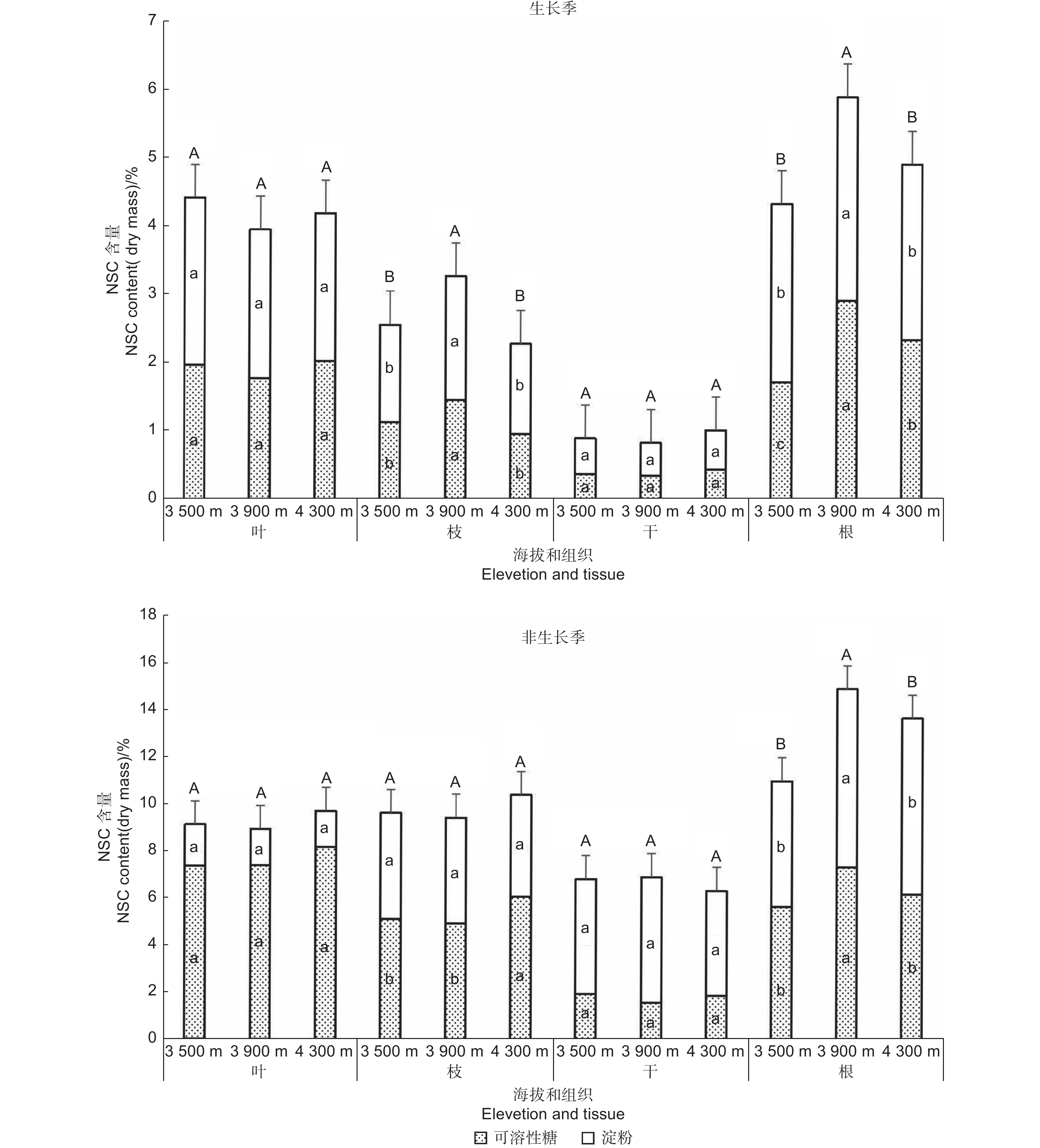
Note: Different lowercase letters indicate significant difference between different altitudes (LSD, P < 0.05), and different uppercase letters indicate significant difference between seasons.不同器官NSC、可溶性糖和淀粉的含量差异显著(表2)。从图1可以看出,冷杉叶的NSC含量不同季节在各海拔间均无显著差异;生长季:低海拔(4.42%)>高海拔(4.19%)>中海拔(3.95%);非生长季:高海拔(9.72%)>低海拔(9.13%)>中海拔(8.95%)。冷杉枝的NSC含量在生长季表现为中海拔显著高于高海拔:中海拔(3.28%)>低海拔(2.57%)>高海拔(2.29%);非生长季各海拔枝的NSC含量差异不显著,含量特征正好与生长季相反:高海拔(10.43%)>低海拔(9.65%)>中海拔(9.45%)。冷杉干的NSC含量不同季节在各海拔间均无显著差异,干与枝的NSC含量在海拔间的分布相反,生长季:高海拔(1.04%)>低海拔(0.91%)>中海拔(0.85%);非生长季:中海拔(6.95%)>低海拔(6.86%)>高海拔(6.37%)。冷杉根的NSC含量在生长季与非生长季均表现为:中海拔(5.92%,14.98%)>高海拔(4.93%,13.75%)>低海拔(4.36%,11.05%),生长季中海拔与高海拔和低海拔差异显著,非生长季中高海拔显著高于低海拔。

图 1 不同海拔急尖长苞冷杉各器官NSC及其组分含量。不同大写字母(表示NSC)和小写字母(上部字母代表淀粉,下部字母代表可溶性糖)表示同一器官不同海拔见差异显著(P<0.05,Duncan多重比较)
Figure 1. The content of non-structural carbohydrates (the sum of soluble sugar and starch content) in various organs of Abies georgei var. smithii at different altitudes. Different uppercase letters (indicating non-structural carbohydrates) and lowercase letters (upper letter representing starch, lower letter representing soluble sugar) indicate significant differences in the same tissue at different altitudes (P<0.05, Duncan test)
-
从表4可以看出,生长季冷杉的可溶性糖含量中海拔、高海拔子集显著>高海拔、低海拔子集;非生长季可溶性糖含量随海拔升高而增加,高海拔、中海拔子集显著>中海拔、低海拔子集。非生长季冷杉高、中、低海拔的可溶性糖含量分别是生长季的3.85、3.26和3.85倍。
表 4 不同海拔急尖长苞冷杉可溶性糖、淀粉含量及其比值
Table 4. The content and ratio of soluble sugar and starch of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
Altitude可溶性糖/%
Soluble sugar淀粉/%
Starch可溶性糖/淀粉
Soluble sugar/Starch生长季
The growing season3 500 1.31 ± 0.15bB 1.76 ± 0.20abB 0.76 ± 0.03bB 3 900 1.64 ± 0.22aB 1.86 ± 0.21aB 0.85 ± 0.04aB 4 300 1.46 ± 0.19abB 1.66 ± 0.19bB 0.85 ± 0.03aB 非生长季
The non-growing season3 500 5.03 ± 0.48bA 4.14 ± 0.35bA 1.70 ± 0.35aA 3 900 5.33 ± 0.56abA 4.75 ± 0.52aA 1.84 ± 0.43aA 4 300 5.61 ± 0.54aA 4.46 ± 0.52abA 2.17 ± 0.60aA 注:不同小写字母表示不同海拔之间的差异显著(最小差异显著法(LSD,P<0.05),不同大写字母表示季节间的差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference between different altitudes (LSD, P < 0.05), and different uppercase letters indicate significant difference between seasons (P < 0.05).由图1可知,生长季叶的可溶性糖含量:高海拔(2.03%)>低海拔(1.97%)>中海拔(1.78%),非生长季叶的可溶性糖含量随海拔升高而增加(7.35%,7.39%,8.17%),不同季节海拔间均差异均不显著。生长季枝的可溶性糖含量在中海拔(1.46%)显著高于低海拔(1.13%)和高海拔(0.97%);非生长季枝的可溶性糖含量高海拔(6.09%)显著高于低海拔(5.12%)和中海拔(4.94%)。生长季与非生长季干的可溶性糖含量都在中海拔最低(0.37%,1.61%),生长季高海拔最高(0.46%),而非生长季低海拔最高(1.95%),各海拔间差异不显著。生长季与非生长季根的可溶性糖含量均表现为:中海拔(2.94%,7.38%)>高海拔(2.36%,6.24%)>低海拔(1.74%,5.70%),生长季各海拔间差异显著,非生长季中海拔显著高于低海拔与高海拔。
-
由表4可知,生长季冷杉淀粉含量中海拔、低海拔子集显著>低海拔、高海拔子集;非生长季冷杉淀粉含量中海拔、高海拔子集显著>高海拔、低海拔子集,非生长季冷杉高、中、低海拔的淀粉含量分别是生长季的2.68、2.55和2.36倍。
从图1可以看出,叶的淀粉含量在生长季与非生长季均随海拔升高而减少,生长季低海拔叶的淀粉含量(2.45%)显著高于中高海拔(2.18%,2.17%),非生长季各海拔叶的淀粉含量(1.78%,1.56%,1.54%)之间差异不显著。枝的淀粉含量在非生长季随着海拔的升高而减少(4.52%,4.51%,4.34%),各海拔间差异不显著;生长季中海拔枝的淀粉含量显著高于低海拔和高海拔:中海拔(1.82%)>低海拔(1.43%)>高海拔(1.32%)。干的淀粉含量在生长季和非生长季相反,生长季:高海拔(0.58%)>低海拔(0.52%)>中海拔(0.48%);非生长季:干的淀粉含量特征与生长季相反,中海拔(5.35%)>低海拔(4.84%)>高海拔最低(4.46%);各海拔间均无显著差异。根的淀粉含量在生长季与非生长季都在中海拔最高,生长季:中海拔(2.98%)>低海拔(2.62%)>高海拔(2.58%),非生长季:中海拔(7.60%)>高海拔(7.50%)>低海拔(5.35%);生长季中海拔显著高于高海拔和低海拔,非生长季中海拔、高海拔显著高于低海拔。
-
生长季、非生长季冷杉可溶性糖和淀粉比值均随海拔上升呈增加趋势,海拔间差异不显著(表4)。从表5可以看出,不同生长季的叶、非生长季的枝、生长季的干可溶性糖和淀粉的比值都随海拔升高而增加;生长季枝可溶性糖和淀粉的比值:中海拔=低海拔>高海拔;非生长季不同海拔干的可溶性糖和淀粉的比值:高海拔>低海拔>中海拔。生长季不同海拔叶的可溶性糖和淀粉的比值之间差异不显著,而非生长季高海拔叶可溶性糖和淀粉的比值与低海拔差异显著;不同海拔冷杉枝和干的可溶性糖和淀粉的比值之间均无显著差异。生长季根可溶性糖和淀粉的比值在中高海拔显著高于低海拔,中海拔最高;非生长季根可溶性糖和淀粉的比值随着海拔升高而减少,但各海拔之间差异不显著。
表 5 不同海拔急尖长苞冷杉各器官可溶性糖与淀粉比值
Table 5. The ratio of soluble sugar and starch in various organs of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
Altitude叶
Leaf枝
Branch干
Trunk根
Root生长季
The growing season3 500 0.80 ± 0.05aB 0.79 ± 0.01aB 0.77 ± 0.05aA 0.67 ± 0.07b 3 900 0.83 ± 0.07aB 0.79 ± 0.07aB 0.80 ± 0.06aA 0.99 ± 0.03a 4 300 0.94 ± 0.06aB 0.73 ± 0.03aB 0.81 ± 0.04aA 0.91 ± 0.08a 非生长季The non-growing season 3 500 4.22 ± 0.31bA 1.14 ± 0.07aA 0.39 ± 0.05aB 1.07 ± 0.06a 3 900 4.90 ± 0.45abA 1.17 ± 0.19aA 0.30 ± 0.03aB 0.98 ± 0.05a 4 300 5.97 ± 1.37aA 1.41 ± 0.08aA 0.43 ± 0.06aB 0.86 ± 0.07a 注:不同小写字母表示不同海拔之间的差异显著(P<0.05),不同大写字母表示季节间的差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference between different altitudes (P < 0.05), and different uppercase letters indicate significant difference between seasons (P < 0.05). -
如表6所示,从海拔来看,冷杉C含量在生长季、非生长季海拔间差异均不显著;冷杉N含量不同季节均随海拔上升而增加,生长季各海拔间差异显著,非生长季高海拔、中海拔子集显著>低海拔;冷杉P含量生长季在海拔间差异不显著,非生长季随海拔上升先降低后升高,高海拔、低海拔子集显著>中海拔。C∶N比不同季节均随海拔上升而下降,生长季低海拔、中海拔子集显著>高海拔子集,非生长季低海拔、中海拔子集显著>中海拔、高海拔子集。C∶P比随海拔上升呈先增加后减少的趋势,生长季中海拔、低海拔子集显著>低海拔、高海拔子集,非生长季海拔间差异不显著。N∶P比生长季随海拔升高呈上升趋势,中海拔、高海拔子集显著>高海拔、低海拔子集,非生长季随海拔上升呈先增加后减少的趋势,海拔间差异显著。
表 6 不同海拔急尖长苞冷杉C、N、P含量及化学计量特征
Table 6. The content and stoichiometric characteristics of C, N and P in Abies georgei var. smithii at different altitudes
季节
Season器官
Organ海拔/m
AltitudeC/(g·kg−1) N/(g·kg−1) P/(g·kg−1) C:N C:P N:P 生长季
The growing
season叶
Leaf4 300 508.06 ± 3.07Ac 12.19 ± 0.93Ad 1.22 ± 0.14Ac 41.9 ± 3.45Aa 422.28 ± 53.81Aab 10.05 ± 0.46Ab 3 900 511.65 ± 4.16Ad 12.36 ± 0.84Ad 1.13 ± 0.1Ac 41.57 ± 3.01Aa 453.68 ± 40.29Aa 10.92 ± 0.63Bc 3 500 518.33 ± 4.38Bc 10.89 ± 0.28Bd 1.31 ± 0.12Ac 47.6 ± 1.25Ba 399.01 ± 34.25Aa 8.38 ± 0.69Cd 枝
Branch4 300 488.61 ± 7.78Ab 9.75 ± 1.18Ac 1.78 ± 0.48Ad 50.72 ± 6.17Aa 288.74 ± 64.77Aa 5.64 ± 0.79Aa 3 900 502.46 ± 7.83Bc 8.32 ± 1.21Ac 1.18 ± 0.16Ac 61.46 ± 9.21Ba 433.89 ± 68.47Ba 7.06 ± 0.36Aa 3 500 507.63 ± 7.83Bb 8.17 ± 0.73Ac 1.47 ± 0.25ABc 62.49 ± 4.54Ba 351.48 ± 52.3ABa 5.64 ± 0.85Bb 干
Trunk4 300 481.29 ± 2.62Aa 1.54 ± 0.27Aa 0.23 ± 0.06Ba 319.19 ± 47.81Ab 2 151.11 ± 477.56Bc 6.69 ± 0.69Aa 3 900 477.29 ± 4.64Aa 1.39 ± 0.42Aa 0.18 ± 0.06Ba 371.05 ± 118.38Ab 2 834.81 ± 876.68Bb 7.64 ± 0.72Aa 3 500 477.64 ± 2.06Aa 1.13 ± 0.22Aa 0.16 ± 0.03Ba 436.14 ± 94.91Ac 3 071.08 ± 726.3Bb 7.04 ± 0.56Ac 根
Root4 300 492.21 ± 5.88Bb 4.21 ± 0.66Ab 0.68 ± 0.13Bb 119.36 ± 19.54Ac 748.86 ± 150.39Ab 6.28 ± 0.83Aa 3 900 488.13 ± 5.99Bb 4.2 ± 0.42Ab 0.45 ± 0.05Ab 117.18 ± 12.9Aa 1 084.65 ± 123.2Ba 9.33 ± 1.36Bb 3 500 483.49 ± 7.29Ba 3.64 ± 0.44Ab 0.87 ± 0.23Bb 134.79 ± 19.68Ab 590.7 ± 162.69Aa 4.35 ± 0.78Ca 非生长季
The non-growing
season叶
Leaf4 300 510.94 ± 3.71Ac 13.91 ± 1.1Bd 1.2 ± 0.43Ac 36.92 ± 3.04Aa 501.81 ± 281.83Ba 13.43 ± 6.98Ab 3 900 513.82 ± 4.74ABd 13.18 ± 1.09Bd 1.17 ± 0.11Ac 39.19 ± 3.32Aa 444.23 ± 44.52Ba 11.33 ± 0.4Ac 3 500 514.62 ± 4.39Bd 12.12 ± 1.18Ad 1.21 ± 0.12Ac 42.77 ± 4.02Ba 427.03 ± 39Ba 10 ± 0.52Ac 枝
Branch4 300 493.16 ± 8.36Ab 8.68 ± 1.04Bc 1.76 ± 0.42Bd 57.53 ± 7.25Aa 291.76 ± 58.16Aa 5.04 ± 0.58Aa 3 900 503.07 ± 7.5Bc 8.18 ± 1.17ABc 1.27 ± 0.25Ac 62.71 ± 10.53ABa 411.54 ± 104.27Aa 6.51 ± 0.68Ba 3 500 500.59 ± 9.95Bc 6.38 ± 1.54Ac 1.12 ± 0.37Ac 83.49 ± 26.07Ba 526.34 ± 303.4Aa 6.01 ± 1.33Aa 干
Trunk4 300 473.25 ± 3.78Aa 1.71 ± 0.65Ba 0.25 ± 0.15Ba 302.03 ± 85.46Ac 2 322.86 ± 929.47Ab 7.46 ± 1.48Aa 3 900 471.58 ± 1.3Aa 1.16 ± 0.13Aa 0.14 ± 0.05Aa 411.09 ± 217.85Bc 3 551.93 ± 981.43Bc 8.51 ± 1.68Ab 3 500 475.4 ± 4.46Aa 1.04 ± 0.15Aa 0.14 ± 0.03Aa 466.29 ± 63.67Bc 3 527.29 ± 653.63Bc 7.77 ± 2.25Ab 根
Root4 300 483.01 ± 2.69Bb 5.29 ± 0.77Bb 0.83 ± 0.14Ab 92.85 ± 48.11Ab 595.52 ± 104.91Ba 6.52 ± 1.5Ba 3 900 485.41 ± 8.01Bb 4.26 ± 0.32ABb 0.6 ± 0.32Ab 114.37 ± 9.07Ab 945.23 ± 328.75Cb 8.21 ± 2.81Cb 3 500 481.92 ± 6.2Bb 4.21 ± 0.39Ab 1.41 ± 0.57Bb 115.15 ± 9.94Ab 383.73 ± 129.7Ab 3.35 ± 1.1Ab 从器官来看,冷杉C、N含量在生长季、非生长季均为叶>枝>根>干,器官间差异显著;P含量生长季枝、叶子集>叶、根子集>干子集,差异显著;非生长季枝>叶>根>干,差异显著。C∶N比不同季节均表现为干子集>根子集>枝、叶子集,差异显著;C∶P比生长季干子集显著>根、叶、枝子集,非生长季干子集>根子集>枝、叶子集,差异显著。N∶P比生长季叶子集显著>干、根、枝子集,非生长季叶子集>干子集>根、枝子集,差异显著。
-
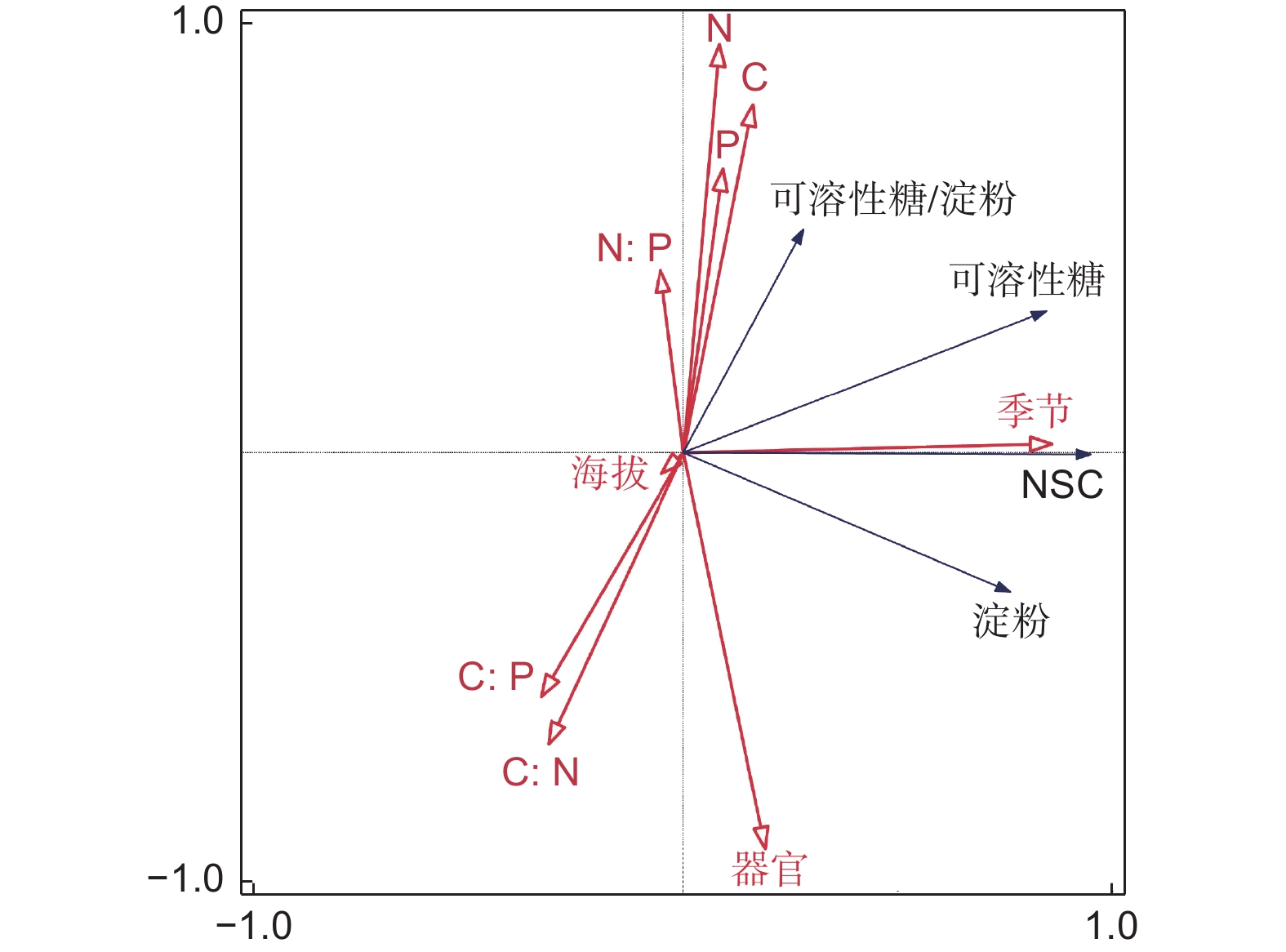
有研究表明[1,19-20],环境胁迫下树木的NSC含量往往高于正常情况,植物体内碳供应充足,本研究中非生长季树木总体NSC含量显著高于生长季,是其2.89~3.23倍,RDA分析结果(图2)也表明季节与冷杉NSC含量、可溶性糖含量和淀粉含量与季节呈正相关,且解释率最大(58.6%),这种非生长季NSC含量高的现象也表现在各个器官上,这是低温胁迫的结果,色季拉山非生长季温度低,有季节性冻土现象,“根际低温”可能会导致树木无法利用充足的光合产物生长和细胞分化[21],同时低温抑制了呼吸作用和植物生长速率,使得碳水化合物消耗减少,NSC在植物组织中积累 [22-23]。植物组织中NSC含量可用于表征光合作用的碳获得与生长呼吸过程的碳需求(即源与汇)之间的平衡[17]。本研究结果表明树木NSC源含量是汇的1.18~1.69倍,生长季各海拔冷杉未表现出碳供应不足;非生长季不同海拔的冷杉NSC源和汇显著高于生长季,且源和汇含量无显著差异,表明非生长季冷杉的源与汇也是平衡的,在进入非生长季前,植物自身储存了大量的NSC,这有利于树木在严酷环境下生存[24]。NSC并不是色季拉山急尖长苞冷杉生长上限的限制条件,更多的可能是低温导致充足的碳不能被生长所利用,从而致使树木新组织形成受到抑制,进而抑制植物生长(即“生长抑制”假说)。

图 2 植物养分与NSC含量RDA分析
Figure 2. RDA analysis of plant nutrients and NSC content
-
在海拔上生长季与非生长季急尖长苞冷杉NSC含量有相同的表现,中海拔显著>低海拔,高海拔非生长季同比增加显著,RDA分析结果表明海拔与NSC含量呈负相关,但影响较小,解释率最低(0.1%),类似情况在周永斌[19]、Piper等[25]的研究报告中有出现,他们认为植物组织的密度随海拔升高而增加,采用干物质作为计算单位会引起稀释效应,进而对结果造成一定影响。另外,中海拔是冷杉的适生区[26],植物生长过程中需要大量的NSC来参与生长过程,而非生长季高海拔NSC含量同比显著增加,也再次表明植物需要更高的NSC含量来应对严酷的生存环境。
-
从器官分布来看,急尖长苞冷杉器官NSC含量均表现为根最高,干最低,生长季叶高于枝,非生长季枝高于叶。RDA分析结果表明器官与NSC含量、淀粉含量呈正相关,对淀粉含量影响较大,解释率为12.0%。非生长季各器官NSC含量显著高于生长季,这是植物器官储备过冬“能源”的表现[27]。根作为植物生长养分来源,需要大量NSC参与以保障生理活动的正常开展,同时也是重要的NSC存储器官非生长季根的NSC含量增长最多,相对增加值达82.19 g·kg−1。枝是树体与叶片间重要的NSC运输通道[28],生长季各海拔急尖长苞冷杉源-汇比均大于1,不需要枝来存储过剩的NSC,主要为叶片等组织生长提供NSC来源保障,故生长季NSC含量枝小于叶;非生长季植物生长速率减弱,各海拔急尖长苞冷杉源-汇比均小于1,枝主要功能转变为贮存NSC。生长季叶是主要的同化器官,新组织的生长需要大量的NSC[29],非生长季叶的同化作用减弱,储存“能源”以应对冬季低温、强辐射等生存环境。干是主要的NSC存储器官,非生长季干NSC含量同比增长6.27~8.52倍,远高于叶的增长速率1.83~2.05倍,充足的NSC储量是来年植物枝、叶生长的重要碳源。
-
树木能长期生活在高山恶劣环境主要依赖于多年生器官中可溶性糖的积累,可溶性糖可以提高组织细胞液渗透压,从而防止细胞内结冰,植物从而可以顺利度过寒冬[30-31]。研究发现,急尖长苞冷杉生长季,非生长季可溶性糖均随海拔上升而增加,非生长季可溶性糖显著高于生长季,为2.96~3.49倍,为了保障树木顺利度过冬季漫长的寒冷,可溶性糖含量维持在一个高水平状态,这是植物的一种自我保护策略。这种保护策略也体现于植物组织的可溶性糖与淀粉比值,有研究表明:足够高的可溶性糖与淀粉比值与充足的碳水化合物是植物顺利过冬的保障[32],冷杉叶的可溶性糖与淀粉比值非生长季较生长季高4.73~5.35倍,这表明急尖长苞冷杉的叶片在非生长季具有大量的可溶性糖来抵御环境胁迫。
可溶性糖与淀粉的相互转化是一种有效的植物抗逆胁迫机制[2],RDA分析表明可溶性糖和淀粉呈显著正相关。而不同器官对可溶性糖和淀粉的存储规律不尽一致,在非生长季叶趋向于积累和存储更多的可溶性糖,较生长季高3.37~3.75倍,相对增加值为最大(49.73 g·kg−1);干趋向于存储更多的淀粉,较生长季高7.88~11.41倍,相对增加值也是最大(49.96 g·kg−1)。非生长季冷杉的枝、干和根,由于低温降低了呼吸作用,导致淀粉大量累积[33],同时也有较高的可溶性糖含量来保证植物能够存活,因此枝、干、根的NSC含量虽然都较高,但可溶性糖和淀粉的比值都不高,尤其是干仅为0.26~0.36。较高的可溶性糖、淀粉和可溶性糖与淀粉比值说明急尖长苞冷杉并没有因为低温等环境胁迫导致高山树木生存受限。
-
RDA分析发现,急尖长苞冷杉NSC含量、可溶性糖和可溶性糖/淀粉比值与季节、C、N、P含量、N∶P比呈正相关,其中N∶P比、N含量解释率分别为2.1%、0.9%;淀粉含量与器官、C∶N比呈正相关,C∶N比解释率为9.6%。植物叶片C、N、P含量及化学计量是评估植物生长是否受限的重要指标,反应了植物对环境的适应性[34]。Koerselman和Meuleman[35]提出植物N∶P<14和 N∶P>16时,植物可能收到N限制或P限制;而Güsewell [36]提出N∶P < 10和N∶P > 20更适用于判定N和P的养分限制类型。本研究发现,冷杉叶片的N含量和N∶P比随海拔上升呈现增加后减少的趋势,N含量低于全国平均(18.74±8.33 mg·g−1)和常绿针叶林平均水平(12.79±4.25 mg·g−1)[37],且N∶P比在各海拔均≤10,表明冷杉的生长主要受N元素限制。冷杉细根C含量略高于全国植物细根平均水平(473.9 mg·g−1),细根N、P含量均显著小于全国平均水平(9.2 mg·g−1、1.0 mg·g−1)[38],表明冷杉根系生长受到N、P元素的双重影响。
-
(1)色季拉山急尖长苞冷杉NSC含量非生长季显著大于生长季3倍左右,急尖长苞冷杉在生长季与非生长季均未出现碳供应不足现象,限制高山林线树木分布上限的更多的是低温造成的生长受限。
(2)随着海拔的升高,生长季与非生长季急尖长苞冷杉叶片可溶性糖和淀粉的比值都逐渐增大,表明植物组织的自我保护策略在越恶劣的环境下越是明显。
(3)植物器官在不同生长季节,为了应对环境的变化,对NSC的利用策略不同。生长季急尖长苞冷杉枝对NSC运输功能大于储藏功能,干的生长优于NSC的储备;非生长季叶趋向于存储更多的可溶性糖,干趋向于存储更多的淀粉,枝、干和根储存了大量的非结构性碳水化合物,用以保障可持续生存。
(4)植物C、N、P含量、季节与急尖长苞冷杉NSC含量、可溶性糖和可溶性糖/淀粉比值呈正相关,其中季节解释率为58.6%;器官与淀粉含量呈正相关,解释率为12%。急尖长苞冷杉叶片生长主要受N元素限制,冷杉根系生长受到N、P元素的双重影响。
藏东南色季拉山急尖长苞冷杉非结构性碳水化合物时空动态特征
Spatiotemporal Dynamic Characteristics of Non-structural Carbohydrates of Abies georgei var. smithii in Sygera Mountain
-
摘要:
目的 为了验证高山林线的形成机理,探究树木分布上限的限制因素。 方法 本研究以色季拉山急尖长苞冷杉为研究对象,在其低海拔针阔叶混交林(3 500 m)、中海拔适生区(3 900 m)和高海拔林线过渡带(4 300 m),分别于生长季(7月初)与非生长季(11月底)测定树木叶、枝、干和根的非结构性碳水化合物(NSC)及其组分含量,以及C、N、P含量。 结果 不同生长季节急尖长苞冷杉非结构性碳水化合物源与汇含量比较稳定,未出现碳供应不足现象;海拔对急尖长苞冷杉非结构性碳水化合物及其组分影响较小;器官间NSC含量差异显著,叶、枝、干和根在不同生长季节所侧重的生理活动和功能不同;非生长季急尖长苞冷杉非结构性碳水化合物及其组分显著高于生长季,非生长季较高的可溶性糖和NSC含量有利于抵御低温等恶劣环境;急尖长苞冷杉的NSC含量随海拔的上升呈现出递增的趋势,一定程度上支持了“生长抑制”假说。 结论 色季拉山急尖长苞冷杉的生长更多的受到了植物养分的限制,而非碳限制。 Abstract:Objective To verify the formation mechanism of the alpine treeline and explore the limiting factors of the upper limit of tree distribution. Methods In the low altitude coniferous and broad-leaved mixed forest (3500 m), suitable altitude area (3900 m) and high altitude treeline transition zone (4300 m) in Sygera Mountain, the contents of non-structural carbohydrates and C, N, and P in leaves, branches, stems and roots of Abies georgei var. smithii as well as were measured both in the growing season (early July) and non-growing season (end of November). Results The results showed that the non-structural carbohydrate sources and sinks of Abies georgei var. smithii were stable in different growing seasons, and there was no insufficient carbon supply. There were little effects of altitude on the non-structural carbohydrates and their components in Abies georgei var. smithii . There were significant differences in NSC content among organs, and the physiological activities and functions of leaves, branches, trunks, and roots were significantly different in different growing seasons. In the non-growing season, non-structural carbohydrates and their components were significantly higher than those in the growing season. In the non-growing season, higher soluble sugars and NSC contents enabled the vegetation to resist cold temperature and other harsh conditions. The content of NSC in Abies georgei var. smithii increased with the increase of altitude, which supported the hypothesis “growth limitation” . Conclusion The growth of Abies georgei var. smithii is more restricted by plant nutrients than by carbon. -
图 1 不同海拔急尖长苞冷杉各器官NSC及其组分含量。不同大写字母(表示NSC)和小写字母(上部字母代表淀粉,下部字母代表可溶性糖)表示同一器官不同海拔见差异显著(P<0.05,Duncan多重比较)
Figure 1. The content of non-structural carbohydrates (the sum of soluble sugar and starch content) in various organs of Abies georgei var. smithii at different altitudes. Different uppercase letters (indicating non-structural carbohydrates) and lowercase letters (upper letter representing starch, lower letter representing soluble sugar) indicate significant differences in the same tissue at different altitudes (P<0.05, Duncan test)
表 1 急尖长苞冷杉样地基本信息
Table 1. Basic information of Abies georgei var. smithii forest sample points
海拔/m
Altitude经纬度
Latitude and
longitude土壤
类型
Soil type坡度/°
Slope坡向/°
Aspect坡位
Slope positions郁闭度
Crown density树高
Tree height/m胸径/cm
DBH冠幅(东西)/m
Canopy
(E-W)冠幅(南北)/m
Canopy
(S-N)年均温/℃
Annual average
temperature年降水/mm
Annual precipitation3 500
(3 530)94°43′31″E;
29°41′53″N酸性暗
棕壤22 25 下 0.90 21.3 ± 1.6 37.4 ± 17.3 6.0 ± 1.9 5.6 ± 1.6 4.67 826.6 3 900
(3 870)94°36′12″E;
29°35′34″N酸性暗
棕壤27 155 中 0.85 27.8 ± 1.3 46.0 ± 15.5 5.7 ± 1.9 5.8 ± 2.0 3.61 867.1 4 300
(4 330)94°41′42″E;
29°36′52″N酸性暗
棕壤25 55 上 0.70 10.7 ± 1.2 34.6 ± 13.3 4.8 ± 1.1 5.1 ± 1.0 0.56 966.3 注:年均温、年降水为样地自动气象站数据
Note: Annual average temperature and annual precipitation are the data of automatic weather stations in the sample area. 下载: 导出CSV
下载: 导出CSV
表 2 不同生长季节急尖长苞冷杉NSC含量、可溶性糖和淀粉含量及其比值影响因子方差分析
Table 2. Variance analysis of non-structural carbohydrate content, soluble sugar and starch content and ratio of soluble sugar and starch in different growth seasons of Abies georgei var. smithii
因变量
Dependent variable变异来源
Sources of variation生长季
The growing season非生长季
The non-growing season自由度
dfF P 自由度
dfF P 可溶性糖
Soluble sugar器官
Organ3 118.98 0.000 3 204.666 0.000 海拔
Altitude2 5.882 0.005 2 3.594 0.035 器官 × 海拔
Organ × Altitude6 6.023 0.000 6 3.775 0.004 淀粉
Starch器官
Organ3 236.366 0.000 3 113.284 0.000 海拔
Altitude2 3.547 0.037 2 3.097 0.054 器官 × 海拔
Organ × Altitude6 3.011 0.014 6 4.013 0.002 NSC 器官
Organ3 210.267 0.000 3 86.804 0.000 海拔
Altitude2 4.851 0.012 2 4.359 0.018 器官 × 海拔
Organ × Altitude6 5.114 0.000 6 4.665 0.001 可溶性糖和淀粉的比值
Soluble sugar/ Starch器官
Organ3 1.657 0.189 3 72.161 0.000 海拔
Altitude2 3.163 0.051 2 1.231 0.301 器官 × 海拔
Organ × Altitude6 2.206 0.058 6 1.054 0.403
下载: 导出CSV
表 3 不同海拔急尖长苞冷杉NSC及源-汇含量
Table 3. Non-structural carbohydrates and source-sink content of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
AltitudeNSC含量/%
NSC concentrationNSC源/%
NSC sourceNSC汇/%
NSC sink源/汇
Source/Sink生长季
The growing season3 500 3.06 ± 0.34bB 4.42 ± 0.20aB 2.61 ± 0.07bB 1.69 ± 0.10aB 3 900 3.50 ± 043aB 3.95 ± 0.11aB 3.35 ± 0.17aB 1.18 ± 0.07bB 4 300 3.12 ± 0.37bB 4.19 ± 0.08aB 2.76 ± 0.14bB 1.52 ± 0.11aB 非生长季
The non-growing season3 500 9.17 ± 0.43bA 9.13 ± 0.48aA 9.18 ± 0.31bA 0.99 ± 0.05aA 3 900 10.08 ± 0.71aA 8.95 ± 0.36aA 10.46 ± 0.30aA 0.86 ± 0.05aA 4 300 10.07 ± 0.64aA 9.72 ± 0.23aA 10.18 ± 0.32aA 0.95 ± 0.04aA 注:不同小写字母表示不同海拔之间的差异显著(最小差异显著法(LSD),P<0.05),不同大写字母表示季节间的差异显著。
Note: Different lowercase letters indicate significant difference between different altitudes (LSD, P < 0.05), and different uppercase letters indicate significant difference between seasons.
下载: 导出CSV
表 4 不同海拔急尖长苞冷杉可溶性糖、淀粉含量及其比值
Table 4. The content and ratio of soluble sugar and starch of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
Altitude可溶性糖/%
Soluble sugar淀粉/%
Starch可溶性糖/淀粉
Soluble sugar/Starch生长季
The growing season3 500 1.31 ± 0.15bB 1.76 ± 0.20abB 0.76 ± 0.03bB 3 900 1.64 ± 0.22aB 1.86 ± 0.21aB 0.85 ± 0.04aB 4 300 1.46 ± 0.19abB 1.66 ± 0.19bB 0.85 ± 0.03aB 非生长季
The non-growing season3 500 5.03 ± 0.48bA 4.14 ± 0.35bA 1.70 ± 0.35aA 3 900 5.33 ± 0.56abA 4.75 ± 0.52aA 1.84 ± 0.43aA 4 300 5.61 ± 0.54aA 4.46 ± 0.52abA 2.17 ± 0.60aA 注:不同小写字母表示不同海拔之间的差异显著(最小差异显著法(LSD,P<0.05),不同大写字母表示季节间的差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference between different altitudes (LSD, P < 0.05), and different uppercase letters indicate significant difference between seasons (P < 0.05).
下载: 导出CSV
表 5 不同海拔急尖长苞冷杉各器官可溶性糖与淀粉比值
Table 5. The ratio of soluble sugar and starch in various organs of Abies georgei var. smithii at different altitudes
季节
Season海拔/m
Altitude叶
Leaf枝
Branch干
Trunk根
Root生长季
The growing season3 500 0.80 ± 0.05aB 0.79 ± 0.01aB 0.77 ± 0.05aA 0.67 ± 0.07b 3 900 0.83 ± 0.07aB 0.79 ± 0.07aB 0.80 ± 0.06aA 0.99 ± 0.03a 4 300 0.94 ± 0.06aB 0.73 ± 0.03aB 0.81 ± 0.04aA 0.91 ± 0.08a 非生长季The non-growing season 3 500 4.22 ± 0.31bA 1.14 ± 0.07aA 0.39 ± 0.05aB 1.07 ± 0.06a 3 900 4.90 ± 0.45abA 1.17 ± 0.19aA 0.30 ± 0.03aB 0.98 ± 0.05a 4 300 5.97 ± 1.37aA 1.41 ± 0.08aA 0.43 ± 0.06aB 0.86 ± 0.07a 注:不同小写字母表示不同海拔之间的差异显著(P<0.05),不同大写字母表示季节间的差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference between different altitudes (P < 0.05), and different uppercase letters indicate significant difference between seasons (P < 0.05).
下载: 导出CSV
表 6 不同海拔急尖长苞冷杉C、N、P含量及化学计量特征
Table 6. The content and stoichiometric characteristics of C, N and P in Abies georgei var. smithii at different altitudes
季节
Season器官
Organ海拔/m
AltitudeC/(g·kg−1) N/(g·kg−1) P/(g·kg−1) C:N C:P N:P 生长季
The growing
season叶
Leaf4 300 508.06 ± 3.07Ac 12.19 ± 0.93Ad 1.22 ± 0.14Ac 41.9 ± 3.45Aa 422.28 ± 53.81Aab 10.05 ± 0.46Ab 3 900 511.65 ± 4.16Ad 12.36 ± 0.84Ad 1.13 ± 0.1Ac 41.57 ± 3.01Aa 453.68 ± 40.29Aa 10.92 ± 0.63Bc 3 500 518.33 ± 4.38Bc 10.89 ± 0.28Bd 1.31 ± 0.12Ac 47.6 ± 1.25Ba 399.01 ± 34.25Aa 8.38 ± 0.69Cd 枝
Branch4 300 488.61 ± 7.78Ab 9.75 ± 1.18Ac 1.78 ± 0.48Ad 50.72 ± 6.17Aa 288.74 ± 64.77Aa 5.64 ± 0.79Aa 3 900 502.46 ± 7.83Bc 8.32 ± 1.21Ac 1.18 ± 0.16Ac 61.46 ± 9.21Ba 433.89 ± 68.47Ba 7.06 ± 0.36Aa 3 500 507.63 ± 7.83Bb 8.17 ± 0.73Ac 1.47 ± 0.25ABc 62.49 ± 4.54Ba 351.48 ± 52.3ABa 5.64 ± 0.85Bb 干
Trunk4 300 481.29 ± 2.62Aa 1.54 ± 0.27Aa 0.23 ± 0.06Ba 319.19 ± 47.81Ab 2 151.11 ± 477.56Bc 6.69 ± 0.69Aa 3 900 477.29 ± 4.64Aa 1.39 ± 0.42Aa 0.18 ± 0.06Ba 371.05 ± 118.38Ab 2 834.81 ± 876.68Bb 7.64 ± 0.72Aa 3 500 477.64 ± 2.06Aa 1.13 ± 0.22Aa 0.16 ± 0.03Ba 436.14 ± 94.91Ac 3 071.08 ± 726.3Bb 7.04 ± 0.56Ac 根
Root4 300 492.21 ± 5.88Bb 4.21 ± 0.66Ab 0.68 ± 0.13Bb 119.36 ± 19.54Ac 748.86 ± 150.39Ab 6.28 ± 0.83Aa 3 900 488.13 ± 5.99Bb 4.2 ± 0.42Ab 0.45 ± 0.05Ab 117.18 ± 12.9Aa 1 084.65 ± 123.2Ba 9.33 ± 1.36Bb 3 500 483.49 ± 7.29Ba 3.64 ± 0.44Ab 0.87 ± 0.23Bb 134.79 ± 19.68Ab 590.7 ± 162.69Aa 4.35 ± 0.78Ca 非生长季
The non-growing
season叶
Leaf4 300 510.94 ± 3.71Ac 13.91 ± 1.1Bd 1.2 ± 0.43Ac 36.92 ± 3.04Aa 501.81 ± 281.83Ba 13.43 ± 6.98Ab 3 900 513.82 ± 4.74ABd 13.18 ± 1.09Bd 1.17 ± 0.11Ac 39.19 ± 3.32Aa 444.23 ± 44.52Ba 11.33 ± 0.4Ac 3 500 514.62 ± 4.39Bd 12.12 ± 1.18Ad 1.21 ± 0.12Ac 42.77 ± 4.02Ba 427.03 ± 39Ba 10 ± 0.52Ac 枝
Branch4 300 493.16 ± 8.36Ab 8.68 ± 1.04Bc 1.76 ± 0.42Bd 57.53 ± 7.25Aa 291.76 ± 58.16Aa 5.04 ± 0.58Aa 3 900 503.07 ± 7.5Bc 8.18 ± 1.17ABc 1.27 ± 0.25Ac 62.71 ± 10.53ABa 411.54 ± 104.27Aa 6.51 ± 0.68Ba 3 500 500.59 ± 9.95Bc 6.38 ± 1.54Ac 1.12 ± 0.37Ac 83.49 ± 26.07Ba 526.34 ± 303.4Aa 6.01 ± 1.33Aa 干
Trunk4 300 473.25 ± 3.78Aa 1.71 ± 0.65Ba 0.25 ± 0.15Ba 302.03 ± 85.46Ac 2 322.86 ± 929.47Ab 7.46 ± 1.48Aa 3 900 471.58 ± 1.3Aa 1.16 ± 0.13Aa 0.14 ± 0.05Aa 411.09 ± 217.85Bc 3 551.93 ± 981.43Bc 8.51 ± 1.68Ab 3 500 475.4 ± 4.46Aa 1.04 ± 0.15Aa 0.14 ± 0.03Aa 466.29 ± 63.67Bc 3 527.29 ± 653.63Bc 7.77 ± 2.25Ab 根
Root4 300 483.01 ± 2.69Bb 5.29 ± 0.77Bb 0.83 ± 0.14Ab 92.85 ± 48.11Ab 595.52 ± 104.91Ba 6.52 ± 1.5Ba 3 900 485.41 ± 8.01Bb 4.26 ± 0.32ABb 0.6 ± 0.32Ab 114.37 ± 9.07Ab 945.23 ± 328.75Cb 8.21 ± 2.81Cb 3 500 481.92 ± 6.2Bb 4.21 ± 0.39Ab 1.41 ± 0.57Bb 115.15 ± 9.94Ab 383.73 ± 129.7Ab 3.35 ± 1.1Ab
下载: 导出CSV
-
[1] 李婷婷, 薛璟祺, 王顺利, 等. 植物非结构性碳水化合物代谢及体内转运研究进展[J]. 植物生理学报, 2018, 54(1):25-35. [2] 杜建会, 邵佳怡, 李升发, 等. 树木非结构性碳水化合物含量多时空尺度变化特征及其影响因素研究进展[J]. 应用生态学报, 2020, 31(4):1378-1388. [3] C. Barbaroux, N. Bréda, E. Dufrêne. Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species(Quercus petraea and Fagus sylvatica)[J]. New Phytologist, 2010, 157(3): 605-615. [4] Chapin F. The ecology and economics of storage in plants[J]. Annual Review of Ecology and Systematics, 1990, 21(1): 423-447. doi: 10.1146/annurev.es.21.110190.002231 [5] 岳剑云, 杜常健, 纪 敬, 等. 银杏枝条部位和年龄对不定根形成的影响及其与非结构碳水化合物含量的关系[J]. 林业科学研究, 2018, 31(5):153-158. [6] 樊金拴. 中国冷杉林[M]. 北京: 中国林业出版社, 2007: 169-172. [7] 汪 松, 解 焱. 中国物种红色名录(第一卷: 红色名录)[M]. 北京: 高等教育出版社, 2004. [8] Marie G, Li M, Luo T, et al. Linking carbon supply to root cell-wall chemistry and mechanics at high altitudes in Abies georgei[J]. Annals of Botany, 2001, 107: 311-320. [9] Stevens G C, Fox J F. The causes of treeline[J]. Annual Review of Ecology & Systematics, 1991, 22(1): 177-191. [10] KöRner C. A re-assessment of high elevation treeline positions and their explanation[J]. Oecologia, 1998, 115(4): 445-459. doi: 10.1007/s004420050540 [11] 李江荣, 高 郯, 郑维列, 等. 急尖长苞冷杉4种林型土壤养分特征[J]. 中南林业科技大学学报, 2021, 41(11):108-119. doi: 10.14067/j.cnki.1673-923x.2021.11.013 [12] 高 郯, 李江荣, 卢 杰, 等. 色季拉山急尖长苞冷杉林不同坡向土壤养分及肥力研究[J]. 生态学报, 2020, 40(4):1331-1341. [13] 吴强盛. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2016. [14] 王学奎. 植物生理生化试验原理和技术[M]. 北京: 高等教育出版社, 2006. [15] 中国林业科学研究院林业研究所. LY/T 1271-1999 森林植物与森林枯枝落叶层全氮、磷、钾、钠、钙、镁的测定[S]//国家林业局. 森林土壤分析方法. 北京: 中国标准出版社, 1999: 295-297. [16] 王 彪, 江 源, 王明昌, 等. 芦芽山不同海拔白杄非结构性碳水化合物含量动态[J]. 植物生态学报, 2015, 39(7):746-752. doi: 10.17521/cjpe.2015.0071 [17] Li M H, Xiao W F, Wang S G, et al. Nitrogen and carbon source–sink relationships in trees at the Himalayan treelines compared with lower elevations[J]. Plant, Cell and Environment, 2008, 31: 1377-1387. doi: 10.1111/j.1365-3040.2008.01848.x [18] 施 征, 白登忠, 张维诚, 等. 青海云杉休眠前后非结构性碳水化合物含量随海拔变化[J]. 林业科学研究, 2017, 30(6):908-915. [19] 周永斌, 吴栋栋, 于大炮, 等. 长白山不同海拔岳桦非结构碳水化合物含量的变化[J]. 植物生态学报, 2009, 33(1):118-124. doi: 10.3773/j.issn.1005-264x.2009.01.013 [20] 郑云普, 王贺新, 娄 鑫, 等. 木本植物非结构性碳水化合物变化及其影响因子研究进展[J]. 应用生态学报, 2014, 25(4):1188-1196. doi: 10.13287/j.1001-9332.2014.0110 [21] Körner C, Paulsen J. A world-wide study of high altitude treeline temperatures[J]. Journal of Biogeography, 2004, 31: 713-732. doi: 10.1111/j.1365-2699.2003.01043.x [22] Li M H, Xiao W F, Wang S G, et al. Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation[J]. Tree Physiology, 2008, 28(8): 1287-1296. doi: 10.1093/treephys/28.8.1287 [23] Lehmann M M, Rinne K T, Blessing C, et al. Malate as a key carbon source of leaf dark-respired CO2 across different environmental conditions in potato plants[J]. Journal of Experimental Botany, 2015, 66: 5769-5781. doi: 10.1093/jxb/erv279 [24] Maunoury F, Berveiller D, Lelarge C, et al. Seasonal, daily and diurnal variations in the stable carbon isotope composition of carbon dioxide respired by tree trunks in a deciduous oak forest[J]. Oecologia, 2007, 151: 268. doi: 10.1007/s00442-006-0592-z [25] Piper F I, Cavieres L A, Marjorie Reyes-Díaz, et al. Carbon sink limitation and frost tolerance control performance of the tree Kageneckia angustifolia D. Don (Rosaceae) at the treeline in central Chile[J]. Plant Ecology, 2006, 185(1): 29-39. doi: 10.1007/s11258-005-9081-4 [26] 郭其强. 西藏急尖长苞冷杉种群生态学特性研究[D]. 杨凌: 西北农林科技大学, 2016. [27] 章异平, 曹鹏鹤, 徐军亮, 等. 秦岭东段栓皮栎叶片非结构性碳水化合物含量的季节动态[J]. 生态学报, 2019, 39(19):7274-7282. [28] 魏龙鑫, 章异平, 李艺杰, 等. 栓皮栎叶片和枝条非结构性碳水化合物调配关系研究[J]. 南京林业大学学报:自然科学版, 2021, 45(2):96-102. [29] 欧阳明, 杨清培, 祁红艳, 等. 亚热带落叶与常绿园林树种非结构性碳水化合物的季节动态比较[J]. 南京林业大学学报:自然科学版, 2014, 38(2):105-110. [30] 康喜坤, 陈小红, 龚 伟, 等. 圆叶玉兰叶片非结构性碳水化合物与氮、磷含量对海拔的响应[J]. 生态学报, 2019, 39(11):4049-4055. [31] 刘万德, 苏建荣, 李帅锋, 等. 云南普洱季风常绿阔叶林主要树种非结构性碳水化合物变异分析[J]. 林业科学, 2017, 53(6):1-9. doi: 10.11707/j.1001-7488.20170601 [32] 上官淮亮, 刘鸿雁, 胡国铮, 等. 干旱林线区不同树种非结构性碳水化合物的季节格局及其主导因子[J]. 北京大学学报:自然科学版, 2019, 55(3):553-560. [33] 刁浩宇, 王安志, 袁凤辉, 等. 特定化合物同位素分析技术在树木非结构性碳水化合物研究中的应用[J]. 应用生态学报., 2020, 31(12):4291-4300. [34] Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J]. Journal of Applied Ecology, 2003, 40(3): 523-534. doi: 10.1046/j.1365-2664.2003.00820.x [35] Koerselman W, Meuleman A F M. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33: 1441-1450. doi: 10.2307/2404783 [36] Güsewell S. N: P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004, 164: 243-266. doi: 10.1111/j.1469-8137.2004.01192.x [37] Tian D, Yan Z B, Niklas K J, et al. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent[J]. National Science Review, 2018, 5: 723-739. [38] 马玉珠, 钟全林, 靳冰洁, 等. 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子[J]. 植物生态学报, 2015, 39:159-166. doi: 10.17521/cjpe.2015.0015 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3024
- HTML全文浏览量: 2301
- PDF下载量: 58
- 被引次数: 0
