


-
山苍子(Litsea cubeba (Lour) Pers.)为樟科(Lauraceae)木姜子属(Litsea Lam.)落叶灌木或小乔木,是我国重要的香料植物资源之一。其植物精油具有特殊的芳香气味及抗菌、抗虫、抗氧化等多种特征,被广泛用于香料、化妆品、医药行业[1-3]。随着生物学技术的发展及山苍子基因组图谱的完成,有助于解读山苍子中基因在各个组织、器官、个体在不同发育时期表达差异,全面理解基因分子生物学功能[4-5]。开展山苍子基因功能研究,将山苍子中重要的功能基因导入到山苍子植株中进行表型鉴定和挖掘,已成为一种主要方法,其中组培再生体系和遗传转化体系的建立是重要研究内容。
植物再生体系是遗传转化的基础,不同外植体再生能力有较大差异,尤其是木本植物生长周期长、再生困难,通过调整外源激素比例可提高其再生效率[6-9]。目前,有关山苍子的腋芽器官离体再生和愈伤诱导研究已有一定的进展[10-13],但由于遗传转化体系的缺乏,利用基因工程技术进行山苍子遗传改良的研究尚未展开。农杆菌介导法是植物遗传转化采用最多的一种方法,具有成本低、操作简便、可重复性好等优点。农杆菌介导遗传转化法原理:植物受伤部位产生酚类物质(乙酰丁香酮等),在酚类物质诱导下农杆菌聚集和vir诱导因子表达,促进T-DNA复合物产生,然后T-DNA 复合物转移并整合到植物细胞的基因组中[14-15]。遗传转化步骤主要包括农杆菌液准备、侵染和共培养、转化组织的筛选和分化、转化植株的检测[16-19]。在筛选培养阶段至少需要添加两种抗生素进行筛选,一种能抑制农杆菌生长,另一种能杀死未转化细胞并保留转化成功的细胞。头孢霉素能显著抑制农杆菌生长,对植物生长影响较小,而潮霉素能显著杀死未转化的植物细胞组织,大幅降低假阳性的产生,这两种抗生素被广泛应用于植物的遗传转化[20-22]。然而,如抗生素浓度使用过度,会影响愈伤组织的正常生长和分化,甚至造成已转化成功愈伤组织的死亡,且不同植物对抗生素忍耐性不同[23]。因此,选择合适的抗生素浓度能显著提升遗传转化的效率。
本研究以快繁的无菌组培苗为试验材料,进一步优化山苍子组培再生过程,研究头孢霉素和潮霉素对山苍子再生的影响,初步建立山苍子的遗传转化体系,为山苍子种质资源繁育及基因功能研究和遗传改良提供技术支撑。
-
本研究以‘安徽3号’优株为山苍子组培苗材料,采集杭州市富阳区新沙岛繁育基地的当年生带芽茎段为外植体,经过消毒处理后在继代培养基中诱导不定芽20~30 d左右,取不定芽更换至新的继代培养基进行增殖培养,以继代培养3~5 cm高的无菌苗进行愈伤诱导及生根等试验,具体消毒过程及苗继代培养基的配方参照林丽媛等方法[24]。试验所需农杆菌菌株采用LBA4404;载体为pCAMBIA1300-GFP,带绿色荧光蛋白标记(GFP)和潮霉素筛选标记;MS固体培养基(M519);6-BA、IBA、TDZ、NAA、IAA先用1 mol ·L−1 NaOH溶解加无菌水配制成1 mg·mL−1母液并4 ℃保存;卡那霉素、头孢霉素、潮霉素用无菌水配制50 mg·mL−1母液并−20 ℃保存,乙酰丁香酮和利福平直接用二甲基亚砜溶解分别配制成200 mg·mL−1、20 mg·mL−1母液并-20 ℃保存。所有培养基pH调至5.8,除6-BA、IBA、TDZ、NAA激素外,其余激素和抗生素均灭菌冷却至50 ℃左右加入。
-
将长至3~5 cm高的幼嫩组培苗去掉叶片和芽并切成0.5 cm左右的茎段,用镊子将茎段夹至愈伤组织诱导培养基中(MS 4.43 g + 蔗糖30 g + 琼脂粉7.5 g + 6-BA 2.0 mg·L−1 + IBA 0.2 mg·L−1,pH 5.8),弱光培养3周,然后弱光继代培养1周。在预实验的基础上,将继代培养1周的愈伤组织接种至含有3种不同浓度的激素组合6-BA(0.1~2.0 mg·L−1)、IBA(0.01~0.10 mg·L−1)、TDZ(0.01~0.50 mg·L−1)MS培养基中光照培养,观察愈伤组织生长和出芽情况。每个组合30个愈伤组织,进行3次以上独立重复试验。
-
将长至3~5 cm高的幼嫩组培苗去掉基部的大部分愈伤组织,接种至含有3种不同浓度的激素组合NAA、6BA、IBA的1/2 MS培养基(1/2 MS 2.17 g + 蔗糖20 g,pH 5.8),光照培养并观察苗状态。
-
将诱导0.5 cm左右的茎段接种至含有0、200、300、500 mg·L−1头孢霉素的愈伤诱导培养基中诱导愈伤组织,观察愈伤组织诱导及生长状态。
-
将诱导0.5 cm左右的茎段和继代培养1周后的愈伤组织分别接种至含有0、5、10、20、30、50 mg·L−1 潮霉素的愈伤诱导培养基和愈伤组织继代培养基中,观察愈伤组织诱导及生长状态。
-
将目的载体质粒转入农杆菌LBA4404中,2 d后挑单菌落接种于2 mL LB液体培养基中(含50 mg·L−1卡那霉素,20 mg·L−1利福平),28 ℃下200 r·min−1振荡培养过夜,取1 mL转移至100 mL含相同抗生素培养基继续培养10 h左右,至菌液OD值在0.5~0.8之间,3500 g离心5 min,用侵染液悬浮农杆菌OD600至0.6~0.8左右(MS 2.22 g·L−1 + 20 g·L−1蔗糖 + 10 g·L−1葡萄糖 + 10 mmol ·L−1MES + 200 mg·L−1 AS)。
-
选取继代培养1周长势良好且鲜嫩的愈伤组织放入侵染液中侵染30 min,用漏勺将侵染后的愈伤组织放至无菌滤纸吸干多余水分,接种至共培养基中(MS 4.43 g·L−1 + 30 g·L−1蔗糖 + 10 g·L−1葡萄糖 + 1 mg·L−1 6BA + 0.1 mg·L−1 IBA + 0.5 mg·L−1 TDZ + 200 mg·L−1 AS),于22~25 ℃暗培养2~3 d。
-
将共培养好的愈伤放入无菌水中清洗3次,每次5 min,再用含300 mg·L−1头孢霉素的无菌水清洗3次,每次5 min。放置无菌滤纸上完全干燥后,夹至筛选培养基中(MS 4.43 g·L−1 + 30 g·L−1蔗糖 + 1 mg·L−1 6BA + 0.1 mg·L−1 IBA + 300 mg·L−1头孢霉素 + 5 mg·L−1潮霉素)。光照条件下筛选培养至刚好可见新的愈伤,然后不断降低头孢霉素浓度、增加潮霉素筛选浓度对抗性愈伤组织进行筛选培养,每15 d更换培养基。
-
将1~2 cm抗性愈伤组织接种至愈伤分化培养基中(MS 4.43 g·L−1 + 30 g·L−1 蔗糖 + 2 mg·L−1 6BA + 0.01 mg·L−1 IBA + 0.05 mg·L−1 TDZ)诱导不定芽,将分化出的芽接种至含20 mg·L−1潮霉素的继代培养基中,进行扩繁。
-
提取3 cm以上抗性苗叶片DNA(共16株),设计3对引物进行验证,第1对引物为插入基因片段,扩增长度为904 bp(21/MAD31-272 bp PF: ATGAATCTGTGCAGTGAAGAAGGAA和eGFP-79 bp PR: CGTCGCCGTCCAGCTCGACCAG);第2对引物为潮霉素基因片段,扩增长度为506 bp(PF: GCGTCTGCTGCTCCATACA和PR: TGACATTGGGGAGTTTAGCG);第3对引物为T-DNA插入区外的卡那霉素基因片段,扩增片段长度为443 bp(PF: ATGTTGCTGTCTCCCAGGTCG和CGGTATAAAGGGACCACCTATGA)。
-
DNA提取方法参考艾德莱公司试剂盒说明书,Southern杂交检测流程如下:取转化得到的抗性愈伤组织和未转化野生型愈伤组织的DNA,各 10 ug,采用Hind III限制酶酶切回收后,按照罗氏公司Southern Blot杂交试剂盒说明书DIG High Prime DNA Labeling and DetectionStarter Kit I,依次进行DNA电泳分离和转膜、使用地高辛标记的潮霉素基因进行杂交、洗膜和显色、拍照等步骤。
-
诱导培养21 d的山苍子愈伤组织置于6-BA、IBA、TDZ 3种不同激素配比培养基下培养。试验显示:6-BA浓度在1.0~2.0 mg·L−1时的愈伤组织生长状态较好,且能保持持续分裂;IBA浓度在0.1 mg·L−1左右时有利于愈伤组织的增殖继代,而在0.01 mg·L−1 IBA低浓度时有利于愈伤组织的分化;TDZ有助于细胞生长及分裂,在加入0.05~0.50 mg·L−1 TDZ后,愈伤组织表现出持续生长和分化,有效防止了褐化。外源激素组合为1.0 mg·L−1 6-BA + 0.1 mg·L−1 IBA + 0.5 mg·L−1 TDZ有利于愈伤增殖继代培养, 愈伤组织仅在2.0 mg·L−16-BA + 0.01 mg·L−1 IBA + 0.05 mg·L−1 TDZ激素组合下出现分化,分化率在16.67%~36.67%之间(表1)。
表 1 外源激素对山苍子愈伤组织分化的影响
Table 1. Effects of exogenous hormones on callus differentiation in L. cubeba
6-BA/
(mg·L−1)IBA/
(mg·L−1)TDZ/
(mg·L−1)愈伤组织生长状态
Callus growth state6-BA/
(mg·L−1)IBA/
(mg·L−1)TDZ/
(mg·L−1)愈伤组织生长状态
Callus growth state0.1 0.01 0.01 褐化严重
Serious browning1.0 0.01 0.10 褐化严重
Serious browning0.1 0.01 0.05 褐化严重
Serious browning1.0 0.01 0.20 轻褐化
Little browning0.1 0.01 0.10 褐化严重
Serious browning1.0 0.01 0.50 轻褐化
Little browning0.1 0.01 0.20 褐化严重
Serious browning1.0 0.05 0.01 轻褐化
Little browning0.1 0.01 0.50 褐化严重
Serious browning1.0 0.05 0.05 轻褐化
Little browning0.1 0.05 0.01 褐化严重
Serious browning1.0 0.05 0.10 轻褐化
Little browning0.1 0.05 0.05 褐化严重
Serious browning1.0 0.05 0.20 轻褐化
Little browning0.1 0.05 0.10 褐化严重
Serious browning1.0 0.05 0.50 轻褐化
Little browning0.1 0.05 0.20 褐化严重
Serious browning1.0 0.10 0.01 轻褐化
Little browning0.1 0.05 0.50 褐化严重
Serious browning1.0 0.10 0.05 轻褐化
Little browning0.5 0.01 0.01 褐化严重
Serious browning1.0 0.10 0.10 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.05 褐化严重
Serious browning1.0 0.10 0.20 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.10 褐化严重
Serious browning1.0 0.10 0.50 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.20 褐化严重
Serious browning2.0 0.01 0.01 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.50 褐化严重
Serious browning2.0 0.01 0.05 淡黄绿色,致密,分化率16.67%~36.67%
Yellowish green, compact,
differentiation rate 16.67%~36.67%0.5 0.05 0.01 褐化严重
Serious browning2.0 0.01 0.10 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.05 褐化严重
Serious browning2.0 0.01 0.20 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.10 褐化严重
Serious browning2.0 0.01 0.50 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.20 褐化严重
Serious browning2.0 0.05 0.01 褐化严重
Serious browning0.5 0.05 0.50 褐化严重
Serious browning2.0 0.05 0.05 褐化严重
Serious browning0.5 0.10 0.01 褐化严重
Serious browning2.0 0.05 0.10 褐化严重
Serious browning0.5 0.10 0.05 褐化严重
Serious browning2.0 0.05 0.20 褐化严重
Serious browning0.5 0.10 0.10 褐化严重
Serious browning2.0 0.05 0.50 褐化严重
Serious browning0.5 0.10 0.20 褐化严重
Serious browning2.0 0.10 0.01 褐化严重
Serious browning0.5 0.10 0.50 褐化严重
Serious browning2.0 0.10 0.05 褐化严重
Serious browning1.0 0.01 0.01 褐化严重
Serious browning2.0 0.10 0.10 褐化严重
Serious browning1.0 0.01 0.05 褐化严重
Serious browning2.0 0.10 0.20 褐化严重
Serious browning -
将长至高3~5 cm的山苍子无菌苗去除基部愈伤组织,置于不同浓度的NAA、IBA、IAA配比下进行生根诱导。结果表明:NAA不能诱导苗生根,而IBA在适宜浓度下能诱导苗生根,但生根率较低,且基部有愈伤形成,阻碍了养分的运输及根的形成,导致叶片掉落,苗生长状态差。在培养基中仅添加IAA时植株生长状态较好,基部无愈伤组织形成,在0.5 mg·L−1 IAA时生根率达97.33%(表2)。
表 2 外源激素对山苍子组培苗生根的影响
Table 2. Effects of exogenous hormones on rooting of cultured L. cubeba seedlings
NAA/
(mg·L−1)IBA/
(mg·L−1)IAA/
(mg·L−1)生根率
Rooting rate/%植株状态
Plant growth state0.0 0.2 0.0 0.00 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.5 0.5 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.0 0.5 1.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.0 0.5 1.0 28.67±3.06 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.8 0.2 49.33±4.16 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 1.0 0.0 14.67±1.15 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.0 0.5 97.33±2.31 正常生根,基部不长愈伤
Normal rooting, no callus in root bases0.0 0.0 1.0 79.33±5.03 正常生根,基部不长愈伤
Normal rooting, no callus in root bases0.1 0.5 0.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.1 1.0 0.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.5 0.0 0.0 12.67±1.15 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases -
头孢霉素能抑制农杆菌的生长,浓度范围一般控制在200~500 mg·L−1,浓度低于200 mg·L−1不能有效抑制农杆菌的生长,浓度高于500 mg·L−1时影响愈伤的生长及分化。本试验在0、200、300、500 mg·L−1不同头孢霉素浓度下诱导茎段愈伤,在浓度200~300 mg·L−1时愈伤组织颜色鲜绿保持正常,与不添加头孢霉素保持一致。在浓度500 mg·L−1时愈伤组织颜色比正常愈伤更为深绿,出现老化状态。因此,筛选培养前期选择头孢霉素浓度300 mg·L−1作为农杆菌抑制浓度,后期应不断降低头孢霉素浓度,防止对愈伤分化产生影响(表3)。
表 3 不同浓度头孢霉素对愈伤组织诱导及生长的影响
Table 3. Effects of different concentrations of cefotaxime on callus induction and growth
头孢霉素浓度
Cefotaxime
concentrations/
(mg·L−1)愈伤诱导率
Callus induction
rate/%愈伤生长状态
Callus growth
state0 100 颜色鲜绿
Fresh green200 100 颜色鲜绿
Fresh green300 100 颜色鲜绿
Fresh green500 100 颜色深绿
Dark green -
潮霉素对愈伤组织诱导及生长影响较大,潮霉素浓度在5 mg·L−1时,能很大程度地诱导愈伤组织,但愈伤组织整体状态差且诱导培养3周后愈伤组织全部褐化;随着潮霉素浓度的增加,其浓度10 mg·L−1时已不能诱导愈伤组织(表4)。将继代培养1周后的愈伤组织在含潮霉素培养基中培养3周,随着潮霉素浓度的增加愈伤组织死亡越多,10 mg·L−1浓度下愈伤组织出现大面积褐化死亡,浓度30 mg·L−1的愈伤组织表面已无可见愈伤。因此,综合考虑遗传转化过程中多步骤操作及农杆菌对愈伤的影响,以潮霉素浓度5 mg·L−1为第1次筛选浓度,持续时间为7~10 d,潮霉素浓度20~30 mg·L−1 作为愈伤组织临界筛选浓度,持续时间应不短于2个月。
表 4 潮霉素对山苍子愈伤组织诱导及生长的影响
Table 4. Effects of hygromycin on callus induction and growth of L. cubeba
潮霉素浓度/
(mg·L−1)
Hygromycin
concentrations愈伤组织诱导
Callus induction愈伤组织生长
Callus growth0 100%成活,鲜绿色
100% survival, fresh green100%成活
100% survival5 100%成活,暗淡
100% survival, with dull callus80%成活,愈伤暗淡
80% survival, with dull callus10 无愈伤,逐渐褐化
No callus, gradually browning15%成活,大面积褐化
15% survival, many browning20 无愈伤,逐渐褐化
No callus, gradually browning5%成活,大面积褐化
5% survival, many browning30 全部褐化死亡
All browning death全部褐化死亡
All browning death50 全部褐化死亡
All browning death全部褐化死亡
All browning death100 全部褐化死亡
All browning death全部褐化死亡
All browning death -
山苍子农杆菌遗传转化流程见图1。首先在光照条件下诱导愈伤组织约21 d,并继代培养7~10 d,然后采用农杆菌侵染继代培养后的愈伤组织30 min,放置于无菌滤纸上超净台吹干至愈伤组织表面无水分;将侵染后愈伤组织转移至共培养基黑暗共培养2~3 d至刚好肉眼可见农杆菌,立即用头孢霉素清洗愈伤组织并吹干,此处应完全吹干至愈伤组织表面发白为止,防止农杆菌污染;转移愈伤组织至含5 mg·L−1潮霉素筛选培养基初步筛选抗性愈伤组织30 d左右,再将愈伤组织转移至含30 mg·L−1潮霉素进行继代培养约60 d;最后将新长出的抗性愈伤组织在10 mg·L−1潮霉素下进行抗性愈伤组织分化约30~60 d,并将诱导出的芽继代培养约60~90 d。从农杆菌侵染愈伤组织至产生抗性幼苗需要约7~9个月时间。

图 1 农杆菌介导的山苍子遗传转化流程
Figure 1. Agrobacterium mediated genetic transformation process of L. cubeba
阳性苗的验证首先采用PCR检测目的基因的方法,共检测了16株抗性幼苗,利用第1对和第2对引物均扩增出目的条带,送北京擎科生物科技有限公司测序拼接后,与插入基因和载体的序列比对完全正确和无移码突变,表明目的基因已插入山苍子基因组中(图2)。且通过第3对引物扩增未扩增出条带,说明没有农杆菌质粒的干扰,排除了假阳性的存在(图2)。共计转化了300个左右愈伤组织,得到约80个抗性愈伤,共2个愈伤分化,得到38株幼苗,转化率为0.67%。
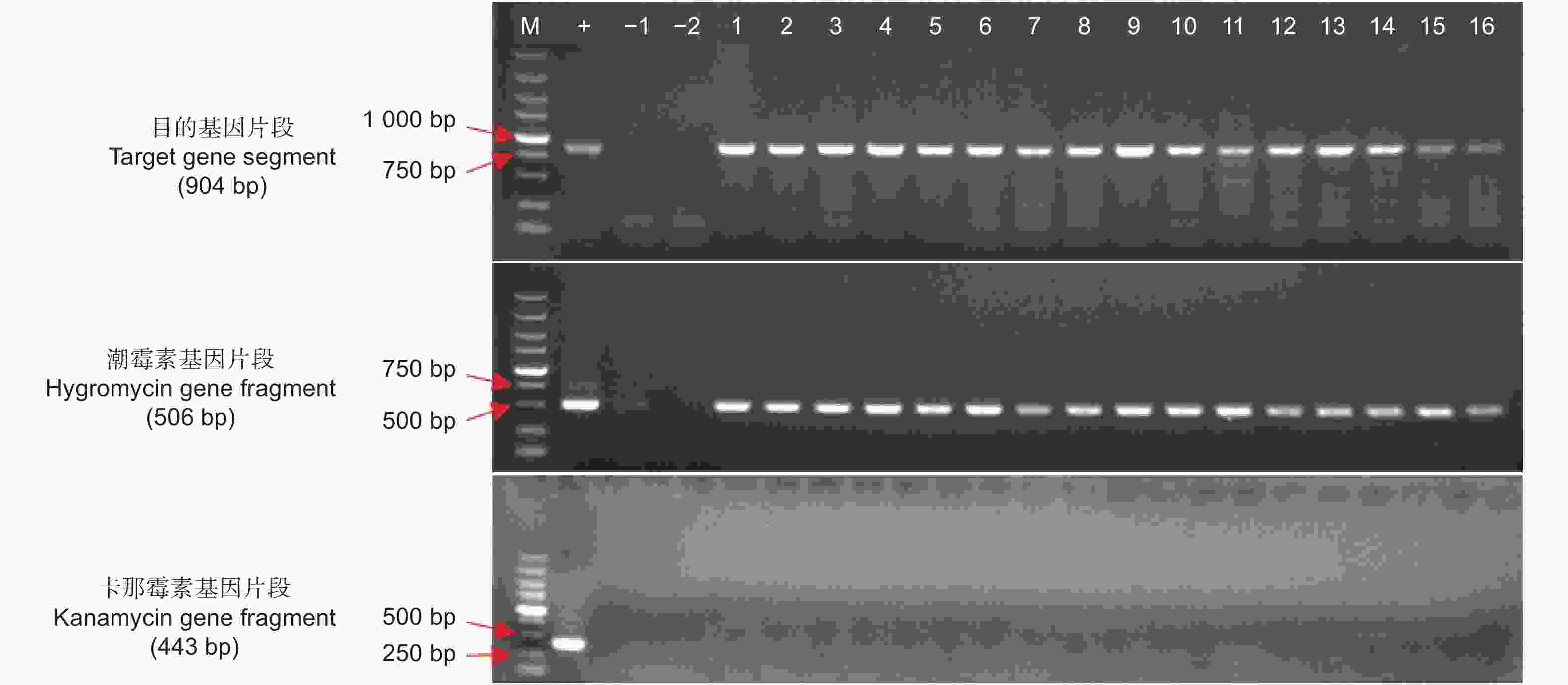
图 2 转基因幼苗的PCR鉴定
Figure 2. PCR identification of transgenic plants
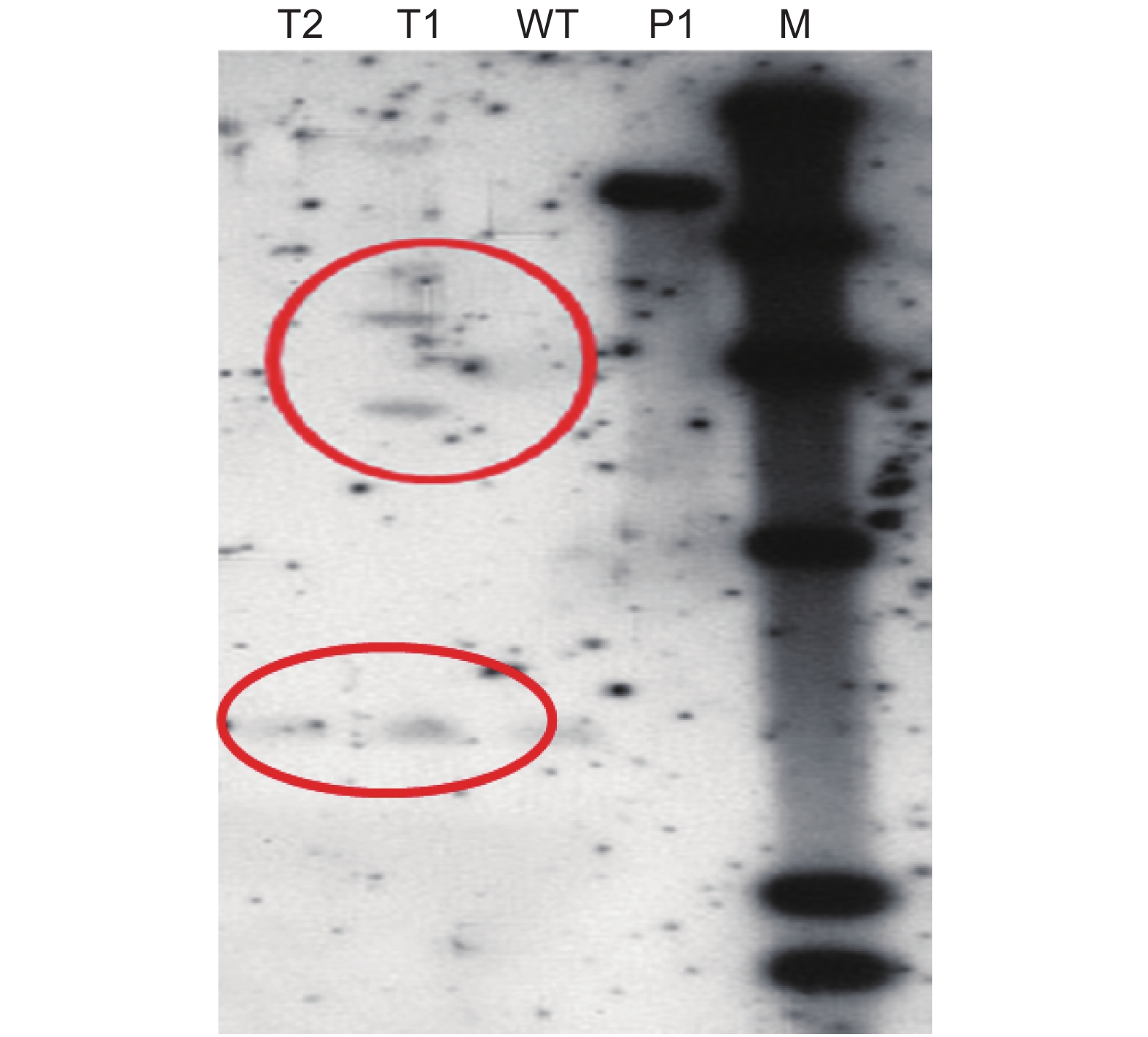
PCR结果主要是利用特异性引物的聚合酶链式反应,可能存在一定的假阳性。而Southern杂交法是当前鉴定外源基因整合及表达的权威方法,具有灵敏性高、特异性强、假阳性低的优点。因此,通过Southern杂交试验进一步验证目的基因是否插入山苍子基因组中。结果表明,转化的抗性愈伤组织中有阳性条带,外源基因已插入山苍子基因组中(图3)。
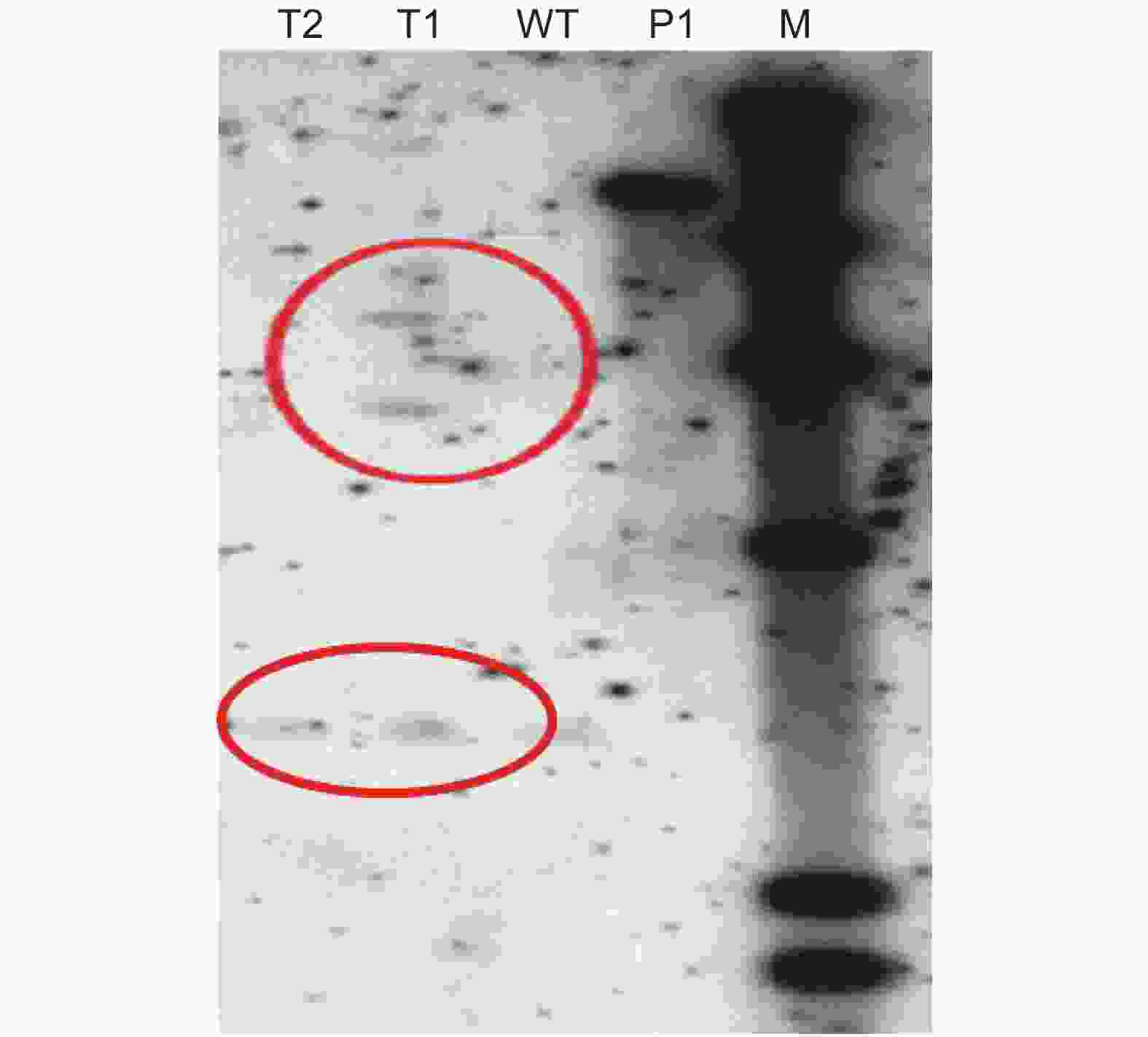
图 3 转基因愈伤组织Southern鉴定
Figure 3. Southern identification of transgenic callus
-
山苍子育种主要以提高果实产量及其品质为目标,利用遗传转化技术能将外源目的基因导入山苍子中进行遗传改良,有助于提高果实产量和品质,创制新的优良种质资源。山苍子全基因组测序的完成,对全面解析和利用山苍子基因分子生物学功能具有重要意义[4]。目前山苍子转基因体系尚未建立,高效稳定的离体再生体系又是影响遗传转化成功的一个重要因素。已有文献报道中愈伤分化率较低,易出现褐化现象且不稳定,不利于开展山苍子基因功能的研究[24]。因此,本研究优化了茎段组织离体再生过程并初步建立山苍子遗传转化流程。
不同植物再生效率具有较大差异,可通过调整激素的种类及配比促进植株再生[25-26]。细胞分裂素能增强细胞的分裂与膨大,生长素促使细胞的伸长,当细胞分裂素/生长素比例较高时能促进芽的分化,比例适中有利于愈伤组织的诱导与苗的生长,比例较低时促进根的生长[27-29]。先前研究表明,山苍子愈伤组织诱导最适激素比例6-BA∶IBA为20∶1[24]。本试验结果中也得到证实,6-BA浓度低于1.0 mg·L−1时愈伤组织生长较差,6-BA∶IBA比例在10∶1至20∶1时能诱导愈伤组织且正常生长繁殖。当降低IBA浓度使细胞分裂素/生长素比例达到200∶1时愈伤组织开始分化形成不定芽,而低于此比例愈伤不分化。因此,山苍子愈伤组织诱导生长培养基6-BA浓度采用2.0 mg·L−1,IBA浓度采用0.1 mg·L−1,在愈伤分化阶段应提高6-BA∶IBA的比值,采用2.0 mg·L−1 6BA和0.01 mg·L−1 IBA浓度。TDZ是一种多功能性人工合成的植物类激素,能诱导芽和愈伤组织的形成与增殖,促进体细胞胚的发生,降低褐化,被广泛应用于木本植物组织培养[30-31]。山苍子愈伤组织分化较困难,长期培养易出现褐化现象。先前山苍子组织培养过程中发现,仅添加6-BA和IBA的激素组合,愈伤组织不能保持持续的增殖继代,而本研究中添加0.05~0.50 mg·L−1浓度范围的TDZ后能有效地保存愈伤组织的继代繁殖能力,褐化现象降低。同时,只有在TDZ存在时,愈伤组织才能分化出不定芽,表明TDZ是一种对山苍子高效再生的激素,在遗传转化过程中发挥重要作用。
不定芽生根是组培再生过程形成完整植株的重要步骤,发达粗壮的根系有利于幼苗的移栽成活和植株的生长。细胞分裂素能抑制植株生根,而生长素有利于植株生根[32-34]。已有山苍子组培生根文献报道中,NAA、IBA、IAA 3种生长激素不同组合均能诱导生根,但生根过程中植株下部叶片脱落,基部有愈伤形成,根数量少且形成一个长的主根,移栽成活率低[11-12,35-36]。本研究3种激素试验发现,IAA有助于山苍子不定芽生根,单独添加IAA诱导生根率达97.33%,且较少有叶片的衰老掉落现象。这可能是诱导生根过程中,在不定芽底部产生伤口后,添加高活性生长素NAA、IBA诱导了愈伤组织形成,阻碍整个植株吸收培养基养分,表现出叶片掉落,甚至出现植株死亡的现象(活性NAA>IBA>IAA)。试验中基本无侧根产生,但移栽穴盆1个月后能产生大量的侧根,形成发达的根系。原因可能是植物具有适应环境的能力,培养基中营养成分比在土壤中更易吸收,不需要产生更多的侧根吸收营养来保证植株的正常生长发育,或由于培养基中某种元素成分相对丰富抑制了侧根的发育,具体原因有待研究。同时存在诱导生根过程中过小的苗大部分死亡,过大组培苗也不能诱导生根的现象,且移栽成活率不高。因此,应充分考虑植株大小、生根时间和移栽过程,提高植株存活率。
-
遗传转化过程中抗生素临界浓度的选择十分重要。抗生素浓度过高不利于愈伤组织的分化及芽的正常生长,过低不能有效筛选出抗性植株。头孢霉素对农杆菌抑制范围一般选择在200~500 mg·L−1[37-39]。本试验中,发现头孢霉素浓度在500 mg·L−1时愈伤组织颜色较深绿,成熟过早不利于愈伤的分化。200~300 mg·L−1浓度范围愈伤组织与不添加头孢霉素无显著性差异,因此选择头孢霉素浓度300 mg·L−1作为农杆菌临界浓度,但后期步骤中应不断降低浓度,减少头孢霉素对愈伤分化增殖的影响。
潮霉素能显著抑制非转化植株的生长,假阳性低,被广泛应用于遗传转化体系中,且随转化过程不断调整抗生素浓度,以达到最佳转化目的。大多数文献报道中,一般采用先高后低的原则,即先使用高浓度潮霉素最大程度地杀死非转化细胞,降低后期筛选的工作量,保证极少的嵌合体存在。而后因潮霉素不断累积毒害作用,应适当降低潮霉素浓度,保证愈伤分化及芽的生长[40-43]。本试验前期预实验结果发现,在筛选培养初期阶段,由于采用高浓度的潮霉素和农杆菌对愈伤的双重毒害,愈伤褐化严重,并局部出现溃烂现象,转化的愈伤已不能存活。因此,在高浓度潮霉素筛选前,先采用5 mg·L−1低浓度潮霉素预筛选7~10 d左右,保证已转化愈伤的正常生长,再使用30 mg·L−1高浓度潮霉素进行筛选,愈伤20~30 d后有抗性愈伤的产生。
在遗传转化过程中,外源基因导入植物细胞的频率较低,通常只有小部分细胞获得了外源DNA,而目的整合到基因组并实现表达的转化细胞则更少。必须采用多种方法反复印证,才能精确筛选和鉴定出转基因植株,包括基因组、转录、翻译水平上的检测。PCR检测具有方便快速、需求更少样本进行转基因植株验证的优点。本试验前期采用PCR方法对插入片段进行检测,较准确地筛选出阳性植株且测序验证,并大概率排除了农杆菌残留的污染。但PCR检测仍可能存在假阳性,某些未转化植株操作在DNA提取过程中的污染也能扩增出相应的片段。Southern杂交法是进行核酸序列分析、重组子鉴定及检测外源基因整合表达的有力手段,具有灵敏性高、特异性强的特点,可清除操作过程中的污染以及转化愈伤组织中质粒残留所引起的假阳性信号[44-45]。本研究中,通过Southern杂交方法检测到阳性条带,证实了目的基因已插入山苍子基因组中。山苍子是一种次生代谢产物丰富的木本油料植物,在遗传转化过程中极易产生褐化导致遗传转化困难,转化效率较低。因此,后续将进一步提高农杆菌转化效率,如侵染农杆菌菌液浓度、时间、共培养时间及AS浓度的条件优化,尤其在遗传转化过程中减少愈伤组织褐化方面进行重点研究。
-
植物组培再生体系和遗传转化体系是培育优良种质资源的重要基础。本研究优化了山苍子组培再生体系,愈伤分化率为16.67%~36.67%,生根率为97.33%。对农杆菌介导的山苍子遗传转化体系进行初步研究,确定了遗传转化过程中最适头孢霉素和潮霉素浓度,且已成功将目的基因导入山苍子基因组中,初步建立山苍子遗传转化体系,为进一步开展基因功能研究及遗传改良提供了技术支持。
农杆菌介导的山苍子遗传转化体系的构建
Establishment of Agrobacterium Mediated Genetic Transformation System of Litsea cubeba
-
摘要:
目的 筛选适合山苍子愈伤组织分化的激素比例,研究其对抗生素的耐受性,构建山苍子遗传转化体系。 方法 以山苍子无菌苗茎段诱导的愈伤组织为试验材料,研究不同激素浓度配比对山苍子愈伤组织诱导不定芽及不定芽生根的影响,探讨愈伤组织对潮霉素和头孢霉素的临界筛选浓度,并通过农杆菌介导法将外源基因导入山苍子愈伤组织中。 结果 筛选出诱导愈伤组织不定芽分化培养基为:MS + 2.0 mg·L−1 6-BA + 0.01 mg·L−1 IBA + 0.05 mg·L−1 TDZ,分化率为16.67%~36.67%;不定芽生根培养基为1/2MS + 0.5 mg·L−1 IAA,生根率为97.33%。遗传转化初期筛选抗性愈伤组织的潮霉素浓度为5 mg·L−1(约7~10 d),通过逐步增加潮霉素浓度至30 mg·L−1进行筛选培养,头孢霉素最适浓度为300 mg·L−1。采用农杆菌介导法将外源基因转入愈伤组织中,并设计PCR引物进行鉴定,共检测16株抗性苗均含有目的条带,表明目的基因已插入山苍子基因组中,转化率为0.67%。通过Southern检测表明,目的片段已插入山苍子愈伤组织中。 结论 初步建立山苍子再生及遗传转化体系,且已获得多个基因的抗性愈伤组织,为进一步开展基因功能研究及遗传改良提供技术支持。 Abstract:Objective To screen the hormone proportion suitable for callus differentiation of Litsea cubeba and clarify its tolerance to antibiotics, and preliminarily establish the genetic transformation system of L. cubeba. Method The effects of different hormone concentrations on adventitious bud induction and adventitious bud rooting of L. cubeba callus were studied. The critical screening concentrations of hygromycin and cefotaxime were discussed, and the foreign gene was introduced into L.cubeba callus by Agrobacterium mediated method. Result The optimum medium for inducing adventitious bud differentiation of callus was MS + 2.0 mg·L−1 6-BA + 0.01 mg·L−1 IBA + 0.05 mg·L−1 TDZ, and the differentiation rate was 16.67%~36.67%; The optimum medium for adventitious bud rooting was 1/2MS + 0.5 mg·L−1 IAA, and the rooting rate was 97.33%. The initial concentration of hygromycin for resistant callus screening was 5 mg·L−1 (about 7~10 days), and then the critical screening culture was carried out by gradually increasing the hygromycin screening concentration to 30 mg·L−1. the optimum concentration of cephalosporin was 300 mg·L−1. Finally, foreign gene was transferred into the callus by Agrobacterium mediated method, and PCR primers were designed for identification. A total of 16 resistant seedlings contained the target band, indicating that the target gene had been inserted into the L. cubeba genome, with a transformation rate of 0.67%. In addition, our research group obtained the resistant calli of multiple genes through this method. Southern detection showed that the target fragment had been inserted into the calli of L. cubeba. Conclusion The regeneration and genetic transformation system of L. cubeba has been preliminarily established, and the resistant calli of multiple genes have been obtained, which provides technical support for further gene function research and genetic improvement. The next step is to optimize the genetic transformation system and improve its transformation efficiency. -
Key words:
- Litsea cubeba
- / regeneration
- / antibiotic
- / genetic transformation
- / Agrobacterium mediated method
-
图 1 农杆菌介导的山苍子遗传转化流程
Figure 1. Agrobacterium mediated genetic transformation process of L. cubeba
表 1 外源激素对山苍子愈伤组织分化的影响
Table 1. Effects of exogenous hormones on callus differentiation in L. cubeba
6-BA/
(mg·L−1)IBA/
(mg·L−1)TDZ/
(mg·L−1)愈伤组织生长状态
Callus growth state6-BA/
(mg·L−1)IBA/
(mg·L−1)TDZ/
(mg·L−1)愈伤组织生长状态
Callus growth state0.1 0.01 0.01 褐化严重
Serious browning1.0 0.01 0.10 褐化严重
Serious browning0.1 0.01 0.05 褐化严重
Serious browning1.0 0.01 0.20 轻褐化
Little browning0.1 0.01 0.10 褐化严重
Serious browning1.0 0.01 0.50 轻褐化
Little browning0.1 0.01 0.20 褐化严重
Serious browning1.0 0.05 0.01 轻褐化
Little browning0.1 0.01 0.50 褐化严重
Serious browning1.0 0.05 0.05 轻褐化
Little browning0.1 0.05 0.01 褐化严重
Serious browning1.0 0.05 0.10 轻褐化
Little browning0.1 0.05 0.05 褐化严重
Serious browning1.0 0.05 0.20 轻褐化
Little browning0.1 0.05 0.10 褐化严重
Serious browning1.0 0.05 0.50 轻褐化
Little browning0.1 0.05 0.20 褐化严重
Serious browning1.0 0.10 0.01 轻褐化
Little browning0.1 0.05 0.50 褐化严重
Serious browning1.0 0.10 0.05 轻褐化
Little browning0.5 0.01 0.01 褐化严重
Serious browning1.0 0.10 0.10 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.05 褐化严重
Serious browning1.0 0.10 0.20 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.10 褐化严重
Serious browning1.0 0.10 0.50 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.20 褐化严重
Serious browning2.0 0.01 0.01 淡黄绿色,致密
Yellowish green, compact0.5 0.01 0.50 褐化严重
Serious browning2.0 0.01 0.05 淡黄绿色,致密,分化率16.67%~36.67%
Yellowish green, compact,
differentiation rate 16.67%~36.67%0.5 0.05 0.01 褐化严重
Serious browning2.0 0.01 0.10 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.05 褐化严重
Serious browning2.0 0.01 0.20 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.10 褐化严重
Serious browning2.0 0.01 0.50 淡黄绿色,致密
Yellowish green, compact0.5 0.05 0.20 褐化严重
Serious browning2.0 0.05 0.01 褐化严重
Serious browning0.5 0.05 0.50 褐化严重
Serious browning2.0 0.05 0.05 褐化严重
Serious browning0.5 0.10 0.01 褐化严重
Serious browning2.0 0.05 0.10 褐化严重
Serious browning0.5 0.10 0.05 褐化严重
Serious browning2.0 0.05 0.20 褐化严重
Serious browning0.5 0.10 0.10 褐化严重
Serious browning2.0 0.05 0.50 褐化严重
Serious browning0.5 0.10 0.20 褐化严重
Serious browning2.0 0.10 0.01 褐化严重
Serious browning0.5 0.10 0.50 褐化严重
Serious browning2.0 0.10 0.05 褐化严重
Serious browning1.0 0.01 0.01 褐化严重
Serious browning2.0 0.10 0.10 褐化严重
Serious browning1.0 0.01 0.05 褐化严重
Serious browning2.0 0.10 0.20 褐化严重
Serious browning 下载: 导出CSV
下载: 导出CSV
表 2 外源激素对山苍子组培苗生根的影响
Table 2. Effects of exogenous hormones on rooting of cultured L. cubeba seedlings
NAA/
(mg·L−1)IBA/
(mg·L−1)IAA/
(mg·L−1)生根率
Rooting rate/%植株状态
Plant growth state0.0 0.2 0.0 0.00 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.5 0.5 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.0 0.5 1.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.0 0.5 1.0 28.67±3.06 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.8 0.2 49.33±4.16 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 1.0 0.0 14.67±1.15 叶片掉落,基部长少量愈伤
Leaf all, little callus in root bases0.0 0.0 0.5 97.33±2.31 正常生根,基部不长愈伤
Normal rooting, no callus in root bases0.0 0.0 1.0 79.33±5.03 正常生根,基部不长愈伤
Normal rooting, no callus in root bases0.1 0.5 0.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.1 1.0 0.0 0.00 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases0.5 0.0 0.0 12.67±1.15 叶片掉落,基部长大量愈伤
Leaf all, many callus in root bases
下载: 导出CSV
表 3 不同浓度头孢霉素对愈伤组织诱导及生长的影响
Table 3. Effects of different concentrations of cefotaxime on callus induction and growth
头孢霉素浓度
Cefotaxime
concentrations/
(mg·L−1)愈伤诱导率
Callus induction
rate/%愈伤生长状态
Callus growth
state0 100 颜色鲜绿
Fresh green200 100 颜色鲜绿
Fresh green300 100 颜色鲜绿
Fresh green500 100 颜色深绿
Dark green
下载: 导出CSV
表 4 潮霉素对山苍子愈伤组织诱导及生长的影响
Table 4. Effects of hygromycin on callus induction and growth of L. cubeba
潮霉素浓度/
(mg·L−1)
Hygromycin
concentrations愈伤组织诱导
Callus induction愈伤组织生长
Callus growth0 100%成活,鲜绿色
100% survival, fresh green100%成活
100% survival5 100%成活,暗淡
100% survival, with dull callus80%成活,愈伤暗淡
80% survival, with dull callus10 无愈伤,逐渐褐化
No callus, gradually browning15%成活,大面积褐化
15% survival, many browning20 无愈伤,逐渐褐化
No callus, gradually browning5%成活,大面积褐化
5% survival, many browning30 全部褐化死亡
All browning death全部褐化死亡
All browning death50 全部褐化死亡
All browning death全部褐化死亡
All browning death100 全部褐化死亡
All browning death全部褐化死亡
All browning death
下载: 导出CSV
-
[1] Si L L, Chen Y C, Han X J, et al. Chemical composition of essential oils of Litsea cubeba harvested from its distribution areas in China[J]. Molecules, 2012, 17(6): 7057-7066. doi: 10.3390/molecules17067057 [2] Gao M, Lin L Y, Chen Y C, et al. Digital gene expression profiling to explore differentially expressed genes associated with terpenoid biosynthesis during fruit development in Litsea cubeba[J]. Molecules, 2016, 21(9): 1251-1267. doi: 10.3390/molecules21091251 [3] 张爱华, 唐春艳, 胡 楠, 等. 我国山苍子产业发展现状及对策[J]. 生物质化学工程, 2020, 54(6):25-32. doi: 10.3969/j.issn.1673-5854.2020.06.005 [4] Chen Y C, Li Z, Zhao Y X, et al. The Litsea genome and the evolution of the laurel family[J]. Nature Communications, 2020, 11(1): 1-14. doi: 10.1038/s41467-019-13993-7 [5] Zhao Y, Chen Y, Gao M, et al. Overexpression of geranyl diphosphate synthase small subunit 1 (LcGPPS. SSU1) enhances the monoterpene content and biomass[J]. Industrial Crops and Products, 2020, 143: 111926. doi: 10.1016/j.indcrop.2019.111926 [6] 徐 麟, 吴 双. 植物再生的理论发展和应用革新[J]. 植物生理学报, 2020, 56(8):1651-1652. doi: 10.13592/j.cnki.ppj.2020.1001 [7] 王栋鑫, 彭 棣, 张 爽. 农杆菌介导木本植物遗传转化的研究进展[J]. 北方园艺, 2018(2):181-185. doi: 10.11937/bfyy.20173329 [8] Sugimoto K, Temman H, Kadokura S, et al. To regenerate or not to regenerate: factors that drive plant regeneration[J]. Current Opinion in Plant Biology, 2019, 47: 138-150. doi: 10.1016/j.pbi.2018.12.002 [9] Bidabadi S S, Jain S M. Cellular, molecular, and physiological aspects of in vitro plant regeneration[J]. Plants, 2020, 9(6): 702. doi: 10.3390/plants9060702 [10] 赵佐敏. 影响山苍子离体快繁效果的主要因素研究[J]. 安徽农业科学, 2005, 33(9):1650-1652. doi: 10.3969/j.issn.0517-6611.2005.09.054 [11] 周传明, 吕曼芳, 陈 奎, 等. 山苍子继代培养中芽增殖效果研究[J]. 广西科学, 2012, 19(4):374-376. doi: 10.3969/j.issn.1005-9164.2012.04.021 [12] 赵 军, 傅体华. 山苍子组织培养快繁试验[J]. 农村经济与科技, 2021, 32(20):31-33. doi: 10.3969/j.issn.1007-7103.2021.20.012 [13] 张付豪, 李佳洛, 王 阳, 等. 山苍子春梢和秋梢带芽茎段最适消毒方法的筛选[J]. 经济林研究, 2019, 37(4):204-209. doi: 10.14067/j.cnki.1003-8981.2019.04.028 [14] Tzfira T, Citovsky V. Agrobacterium-mediated genetic transformation of plants: biology and biotechnology[J]. Current Opinion in Biotechnology, 2006, 17(2): 147-154. doi: 10.1016/j.copbio.2006.01.009 [15] Gelvin S B. Plant proteins involved in Agrobacterium-mediated genetic transformation[J]. Annual Review of Phytopathology, 2010, 48: 45-68. doi: 10.1146/annurev-phyto-080508-081852 [16] Matveeva T V, Otten L. Widespread occurrence of natural genetic transformation of plants by Agrobacterium[J]. Plant Molecular Biology, 2019, 101(4): 415-437. [17] Niazian M, Sadat-Noori S A, Tohidfar M, et al. Agrobacterium-mediated genetic transformation of ajowan (Trachyspermum ammi (L.) Sprague): an important industrial medicinal plant[J]. Industrial Crops and Products, 2019, 132: 29-40. doi: 10.1016/j.indcrop.2019.02.005 [18] Bakhsh A. Development of efficient, reproducible and stable Agrobacterium-mediated genetic transformation of five potato cultivars[J]. Food Technology and Biotechnology, 2020, 58(1): 57-63. doi: 10.17113/ftb.58.01.20.6187 [19] Sood P, Singh R K, Prasad M. An efficient Agrobacterium-mediated genetic transformation method for foxtail millet (Setaria italica L.)[J]. Plant Cell Reports, 2020, 39(4): 511-525. doi: 10.1007/s00299-019-02507-w [20] Tiwari N N, Singh R K, Singh S P. Impact of cefotaxime on Agrobacterium mediated BT gene transformation in sugarcane[J]. Journal of Pharmacognosy and Phytochemistry, 2018, 7(2): 3952-3955. [21] Kumar R, Mamrutha H M, Kaur A, et al. Synergistic effect of cefotaxime and timentin to suppress the Agrobacterium overgrowth in wheat (Triticum aestivum L.) transformation[J]. Asian Journal of Microbiol Biotechnology Environment Science, 2017, 19(4): 961-967. [22] Nai Y S, Lee M R, Kim S, et al. Relationship between expression level of hygromycin B-resistant gene and Agrobacterium tumefaciens-mediated transformation efficiency in Beauveria bassiana JEF-007[J]. Journal of Applied Microbiology, 2017, 123(3): 724-731. doi: 10.1111/jam.13529 [23] Ortiz J P A, Reggiardo M I, Ravizzini R A, et al. Hygromycin resistance as an efficient selectable marker for wheat stable transformation[J]. Plant Cell Reports, 1996, 15(12): 877-881. doi: 10.1007/BF00231579 [24] 林丽媛, 韩小娇, 陈益存, 等. 山鸡椒的愈伤组织诱导及植株再生[J]. 植物生理学报, 2013, 49(10):1047-1052. doi: 10.13592/j.cnki.ppj.2013.10.006 [25] Kim M J, An D J, Moon K B, et al. Highly efficient plant regeneration and Agrobacterium-mediated transformation of Helianthus tuberosus L[J]. Industrial Crops and Products, 2016, 83: 670-679. doi: 10.1016/j.indcrop.2015.12.054 [26] Ikeuchi M, Ogawa Y, Iwase A, et al. Plant regeneration: cellular origins and molecular mechanisms[J]. Development, 2016, 143(9): 1442-1451. doi: 10.1242/dev.134668 [27] Lee K, Seo P J. Dynamic epigenetic changes during plant regeneration[J]. Trends in Plant Science, 2018, 23(3): 235-247. doi: 10.1016/j.tplants.2017.11.009 [28] Qiu Y, Guan S C, Wen C, et al. Auxin and cytokinin coordinate the dormancy and outgrowth of axillary bud in strawberry runner[J]. BMC Plant Biology, 2019, 19(1): 1-16. doi: 10.1186/s12870-018-1600-2 [29] Su Y H, Liu Y B, Zhang X S. Auxin–cytokinin interaction regulates meristem development[J]. Molecular Plant, 2011, 4(4): 616-625. doi: 10.1093/mp/ssr007 [30] Cheng Z J, Zhu S S, Gao X Q, et al. Cytokinin and auxin regulates WUS induction and inflorescence regeneration in vitro in Arabidopsis[J]. Plant Cell Reports, 2010, 29(8): 927-933. doi: 10.1007/s00299-010-0879-8 [31] Ahmad N, Faisal M. Thidiazuron: from urea derivative to plant growth regulator[M]. Singapore: Springer Singapore, 2018: 61-94. [32] 许智宏, 张宪省, 苏英华, 等. 植物细胞全能性和再生[J]. 中国科学:生命科学, 2019, 49(10):1282-1300. [33] Mao C, He J, Liu L, et al. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development[J]. Plant Biotechnology Journal, 2020, 18(2): 429-442. doi: 10.1111/pbi.13209 [34] Liu J, Moore S, Chen C, et al. Crosstalk complexities between auxin, cytokinin, and ethylene in Arabidopsis root development: from experiments to systems modeling, and back again[J]. Molecular Plant, 2017, 10(12): 1480-1496. doi: 10.1016/j.molp.2017.11.002 [35] Neogy A, Singh Z, Mushahary K K K, et al. Dynamic cytokinin signaling and function of auxin in cytokinin responsive domains during rice crown root development[J]. Plant Cell Reports, 2021, 40(8): 1367-1375. doi: 10.1007/s00299-020-02618-9 [36] 文明玲, 徐 茜, 黎 华, 等. 山苍子组培快繁关键技术研究[J]. 陕西农业科学, 2012, 58(3):51-53. doi: 10.3969/j.issn.0488-5368.2012.03.018 [37] 吴幼媚, 蔡 玲, 黄金使, 等. 山苍子组培继代芽瓶外生根技术研究[J]. 广西林业科学, 2011, 40(3):169-172. doi: 10.3969/j.issn.1006-1126.2011.03.003 [38] Gambhir G, Kumar P, Srivastava D K. Effect of antibiotic sensitivity on different cultured tissues and its significance in genetic transformation of cabbage Brassica oleracea[J]. Bioscience Biotechnology Research Communications, 2017, 10(4): 652-661. doi: 10.21786/bbrc/10.4/7 [39] Parmar N, Thakur A K, Kanwar K. Standardization of Agrobacterium tumefaciens-mediated genetic transformation protocol in Punica granatum L. cv. Kandhari Kabuli[J]. Crop Research, 2017, 52(6): 268-276. doi: 10.5958/2454-1761.2017.00028.6 [40] Tsuboyama S, Kodama Y. AgarTrap protocols on your benchtop: Simple methods for Agrobacterium-mediated genetic transformation of the liverwort Marchantia polymorpha[J]. Plant Biotechnology, 2018, 35(2): 93-99. doi: 10.5511/plantbiotechnology.18.0312b [41] 王 坚, 李永玲, 刘 炜. 潮霉素B在遗传转化中应用的研究进展[J]. 宁夏农林科技, 2017, 58(12):36-43. doi: 10.3969/j.issn.1002-204X.2017.12.013 [42] 陈 兰, 朱 晨, 李小桢, 等. 茶树遗传转化体系研究进展[J]. 安徽农业科学, 2019, 47(12):14-18. doi: 10.3969/j.issn.0517-6611.2019.12.004 [43] Van Eck J. The status of Setaria viridis transformation: Agrobacterium-mediated to floral dip[J]. Frontiers in Plant Science, 2018, 9: 652. doi: 10.3389/fpls.2018.00652 [44] Liu H, Zhao H, Wu L, et al. A genetic transformation method for cadmium hyperaccumulator Sedum plumbizincicola and non-hyperaccumulating ecotype of Sedum alfredii[J]. Frontiers in Plant Science, 2017, 8: 1047. doi: 10.3389/fpls.2017.01047 [45] 代 军. Southern杂交技术在农作物遗传转化研究中的应用[J]. 安徽农业科学, 2015, 43(1):20-23. doi: 10.3969/j.issn.0517-6611.2015.01.006 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4705
- HTML全文浏览量: 1927
- PDF下载量: 86
- 被引次数: 0
