

 下载:
下载:

-
随着人类对原始林的干扰破坏,全球次生林的面积占比不断上升[1-2],特别在中国东北地区,由于历史上高强度的采伐干扰而形成的次生林占天然林面积的65%以上[3-4]。关于次生林方面的研究受到了广大学者的关注,有关研究也在各个方面展开[5-9],其中次生林演替规律及其干扰是研究热度较高的一个分支[3, 10-13]。
次生林演替属于一种特殊的森林演替,指的是原生森林受到强烈干扰形成次生林后的一系列生长发育过程[11, 14],这与森林演替中先锋群落至顶级群落的演替过程基本一致[15]。与森林演替相似,次生林演替按照发展趋向同样可分为进展演替和逆行演替[16]。次生林演替的发生与干扰有密不可分的联系。一般而言,次生林在正向干扰或小强度范围内的逆向干扰下可以保持进展演替,一旦逆向干扰强度超过阈值,次生林演替就会停滞,甚至出现逆行演替的情形[15, 17]。
在中国东北地区,关于次生林演替和干扰的研究从20世纪50年代逐渐展开[18-19],至今已形成了大量的研究成果。本研究分别阐述干扰因素与东北次生林演替的关系,中国东北地区地带性群落受干扰后衍生出主要的次生林的演替过程,以及演替规律和干扰作用在次生林恢复演替过程中的应用,并对未来研究进行展望。
-
(1)采伐林木对森林的影响较为复杂,高强度的皆伐或择伐会移除整个森林群落,使林地从先锋次生林群落甚至次生草地重新开始新的次生演替过程[20],如阔叶红松林衍生的次生林遭到皆伐形成了次生杨桦林[21],云冷杉过伐林在反复择伐后更新困难,形成了稳定的亚高山湿草甸[22];若反复的定向择伐顶级树种红松,会使次生林群落缺少红松种源,演替最终会收敛于蒙古栎林、阔叶混交林和针阔混交林等演替中期群落;而适宜的采伐强度和采伐方式会促进森林群落生长发育和更新演替[9, 20, 23]。梁立东等[24]对黑河地区针阔混交林、白桦林、蒙古栎林和杂木林进行了常规抚育经营,发现抚育经营可显著提高演替层林木占比,降低主林层林木占比,加速了天然次生林向地带性顶级群落的恢复演替进程。Dai等[25]指出长白山地区阔叶红松林衍生的次生林群落,适宜根据林分内是否缺乏顶级树种种源,选择不同强度的单株择伐或带状皆伐等抚育经营措施和补植作业方式,才能更科学地促进次生林的恢复演替。
(2)次生林经过进一步垦地成为农田生态系统,土壤的乔木种子库也会在年复一年的农业经营中丧失,这片土地也丧失了进展演替为森林的能力[26]。若多年后成为撂荒地,则需要从草本灌木群落开始演替,依靠农田周边的乔木或经由鸟类取食种子后的粪便获得种源,进而演替至先锋次生林群落,这个过程的时间长短与撂荒地的斑块大小有关[27]。
(3)放牧和采摘松果可能造成森林的逆行演替,这主要是通过影响树种的更新而产生影响[28]。放牧会造成家畜对顶级阔叶树幼苗的啃食,造成自然更新或补植幼苗的死亡,减缓次生林发展到顶级群落的演替速度[29];采摘则直接减少了植物种子的自然扩散[30]。
(4)次生林内补植幼苗,弥补了因采摘松果、缺少种源造成的目标树种更新稀少的不足,在一定程度上可以促进次生林的演替,但其经济成本要比自然更新更高[19, 31]。补植幼苗常常与次生林的抚育采伐结合进行,红玉等[21]和丁磊等[32]在对辽东山区阔叶红松林衍生次生林的恢复经营研究中指出,建议以现有植被为主体,在抚育后“见缝插针”引进红松、沙松等珍贵针叶树种,并保护和培育林冠下萌生的珍贵阔叶目标幼树,通过多次的抚育经营,加快促进次生林正向演替,恢复为顶级阔叶红松林群落。柏广新等[33]对长白山林区的落叶松—白桦林、杨桦林、杂木林、硬阔林和阔叶红松林 5 种典型幼龄次生林群落进行了对照经营、常规经营和优化经营的不同抚育采伐试验,结果显示依据林木营养空间确定合理抚育强度、采取“栽针保阔”途径恢复次生林的优化经营可以改变幼龄林的演替趋势,加速了向顶级阔叶红松林恢复的演替进程。
(5)人工火干扰对次生林演替的作用一般是积极的,有目的性的用火,保持火势在可控范围内,可清理林下灌草,给顶级树种下种和幼苗的生长提供良好条件;若由于人为疏忽原因,加之当时的气候和地形条件,导致火势未能得到及时的控制,则可能会烧死幼苗幼树和种子,使演替进程停滞,甚至逆行演替至前一阶段;当火势发展至重度火烧时,则几乎会烧死全部植被,火烧区的次生林自然恢复完全需要非火烧区的植被扩散来进行,且火烧迹地的大小及形状等空间特征对次生林的演替恢复同样有重要影响[34-35]。
-
(1)自然干扰中,如长白山火山爆发产生的岩浆和厚厚的火山灰覆盖地表[36]、辽东地区山啸所形成的乱石窖[37],可能造成次生林毁灭性死亡,由于地表土壤缺失,整个演替需要从地衣阶段完成整个原生演替进程才能恢复至先锋次生林群落,然后逐步演替到顶级森林群落,整个过程十分漫长。
(2)自然火灾干扰发生频率较高,特别是东北大兴安岭林区,几乎所有的森林都曾在某一时期火烧过[34]。同人为火干扰类似,火后残余的植被繁殖体对于次生林演替有决定作用,且次生林对火灾强度与大小等同样特别敏感[38],兴安落叶松(Larix gmelinii (Rupr.) Kuzen)林在中度和重度火烧多年后,林分生物量与皆伐措施无显著差异,而轻度火烧多年后的生物量则显著高于中度和重度火烧以及皆伐措施,说明轻度火烧下次生林恢复演替效果较好[39]。由于自然火灾干扰可能存在反复回归的可能,若兴安落叶松林反复受到中等强度的干扰,则可能会改变演替的途径, 导致物种组成及结构的转变,形成对火灾适应性更好的偏途顶级樟子松(Pinus sylvestris var. mongolica Litv.)林,而现实中自然火干扰的回归频率和强度不定,因此火烧后的次生林群落演替将异常缓慢且难以预测[34]。
(3)风雪灾害会造成大部分林木出现树干弯曲、折干和根拔,风雪灾害后的病虫害侵袭还会造成更新困难,可能引起次生林的逆行演替[40-41]。整体而言,风雪灾害对次生林的影响会随风雪灾害严重程度的变化而变化,且一旦对林内环境的改变超过一定阈值,会对林下层植被产生较大影响,进而影响演替的进行[42]。
(4)气候变化作用时间长且作用范围大,对次生林演替有持续性影响,除直接通过影响降水和温度对次生林演替产生影响外,还会通过影响火灾、风雪灾害的频率和强度间接影响次生林演替[43-44]。程肖侠等[43]应用FAREAST模型,模拟了东北地区森林在气候变化条件下的动态演替过程,结果显示东北主要森林群落中阔叶树种明显增加,水平地带性森林植被分布北移,长白山地垂直分布林线上移。次生林早期演替过程对气候变化的响应研究结果显示,在缺乏耐寒性的香脂冷杉(Abies balsamea (L.) Mill)的森林景观中,气候变化可能会增加不耐阴阔叶树种的更新数量,导致次生林群落逆行演替[45]。
以上结果表明,人为干扰对次生林演替的作用有利有弊,而大部分的自然干扰均不利于次生林进展演替,但这也不能否定自然干扰的价值,利用好自然干扰的生态作用,加之适当的人为干扰辅助,同样可以改善次生林演替停滞不前的情形,甚至加快次生林的演替进程。
-
东北地区次生林群落类型多样,但都源生于所在气候带的地带性顶级群落,次生林的演替最终也会收敛于地带性顶级群落,因此将次生林按照其源生的地带性顶级群落类型分类来阐述次生林演替进程更为科学。现以兴安落叶松林、阔叶红松(Pinus koraiensis Sieb.et Zucc.)林和山地云冷杉林这3个东北地区的典型顶级群落为例,来说明东北林区顶级群落受到干扰后形成的主要次生林的生态演替序列与演替过程。由于干扰在群落演替过程中无时无刻不在发生,而干扰类型复杂多样[46],因此在演替序列不同群落之间,不区分干扰类型,仅使用干扰强度来串联,强度干扰造成顶级群落的毁灭,中度干扰造成逆行演替,轻度干扰促进群落进展演替。
-
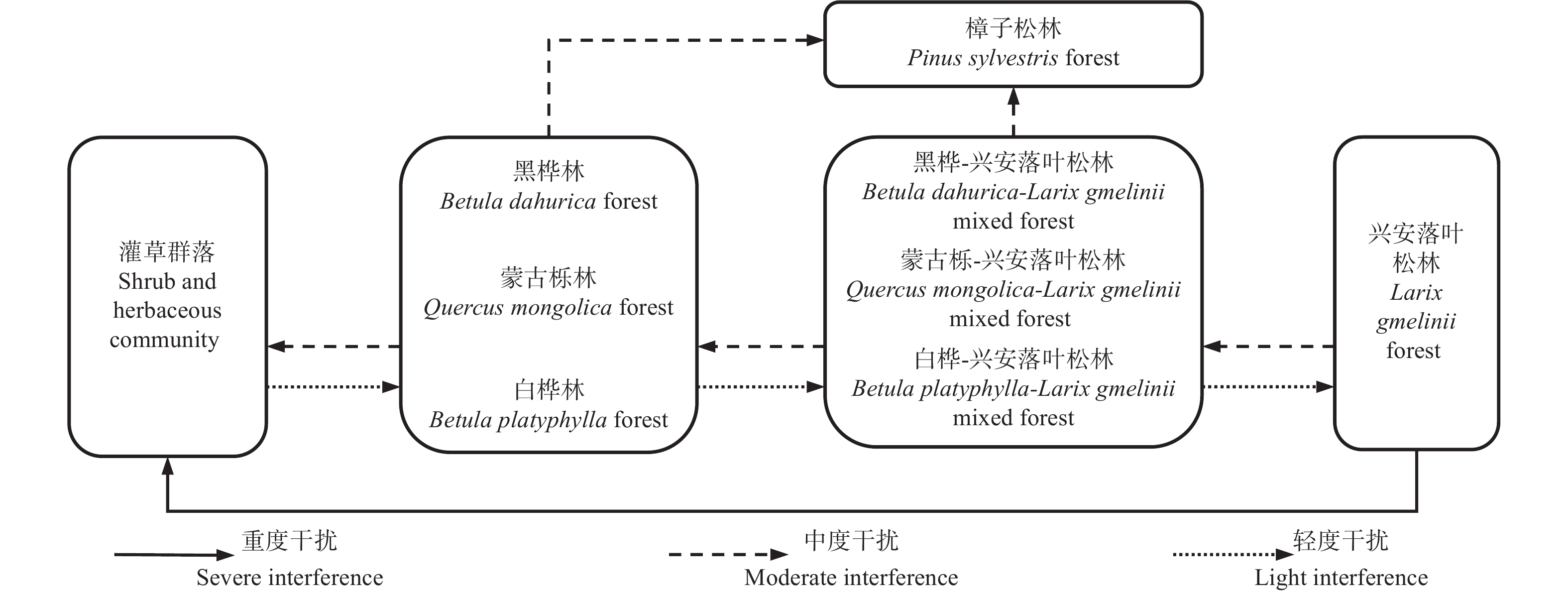
兴安落叶松林主要分布在大兴安岭和小兴安岭北麓,群落几乎全部由兴安落叶松构成[47]。图1展示了兴安落叶松林在干扰条件下形成的主要次生林的生态演替序列[48]。

图 1 兴安落叶松林在干扰下的次生演替序列
Figure 1. Secondary succession of The Larix gmelinii forest under disturbance
兴安落叶松林受到重度干扰(如严重火灾干扰)后,乔木、灌木和草本基本死亡,土壤有机质几乎全部烧掉,存活在土壤中的种子也非常贫乏,整个演替进程需要从灌草群落开始,依赖乔木种子的散播进行森林演替[34]。由于兴安落叶松种子不如白桦(Betula platyphylla Suk.)种子容易传播,因此在仅受轻度干扰条件下,喜光的白桦会率先侵入灌草地,群落演替至白桦次生林,而在火烧迹地周围有大量蒙古栎(Quercus mongolica Fisch. ex Ledeb)和黑桦(Betula dahurica Pall.)种源的情况下,会演替至蒙古栎次生林和黑桦次生林,这3种群落均属于次生林演替的初期阶段;演替初期阶段的3种群落在仅受轻度的干扰下,后期侵入了生命周期长的兴安落叶松,此时,兴安落叶松长势优于先锋树种,会逐渐占据上层,在与先锋树种的不断竞争中,使林分演替至白桦-兴安落叶松林、蒙古栎-兴安落叶松林和黑桦-兴安落叶松林等中期阶段群落;在中期阶段之后,轻度干扰下,兴安落叶松占据森林群落中的主体地位,其他树种很难在群落内更新生长,而兴安落叶松的幼苗数量较多,整个森林群落逐渐开始稳定,进入次生林演替的后期阶段——兴安落叶松林[48-49]。若兴安落叶松林仅受到中度干扰(如小面积的采伐)下,兴安落叶松种源充足,白桦、黑桦和蒙古栎等先锋树种会和兴安落叶松共同更新,进一步形成演替中期的3种群落,若继续施以中度干扰,采伐兴安落叶松,则会逆行演替至演替初期的先锋群落。若中度火灾干扰频繁,则演替初期和中期的群落会趋向于偏途顶级的樟子松林发展[34]。
-
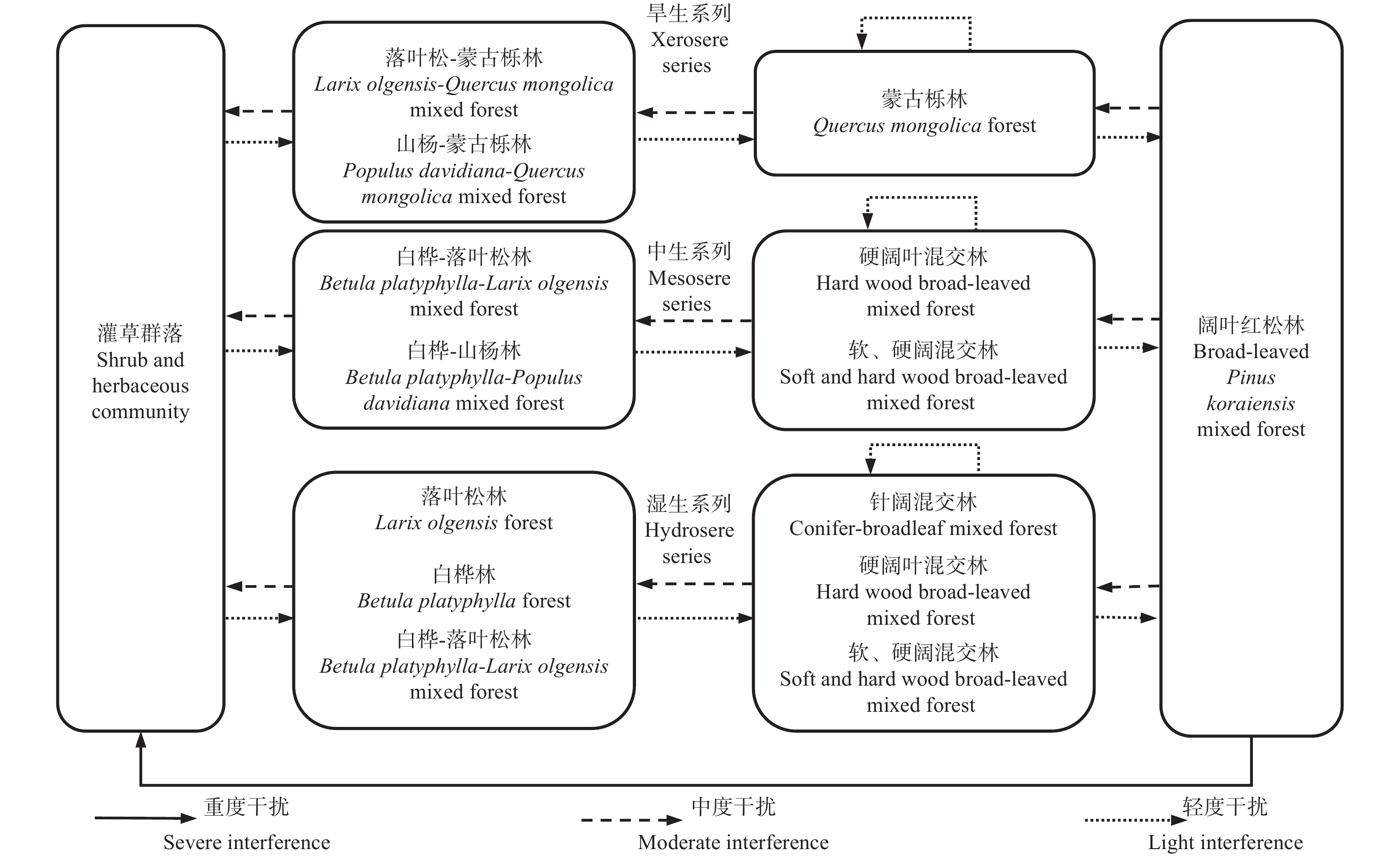
阔叶红松林,即红松阔叶混交林,是东北小兴安岭-长白山地区典型的水平地带性顶极群落,群落中针叶树种以红松为主,同时可能伴生部分鱼鳞云杉(Picea jezoensis var. komarovii (V.Vassil.) Cheng et L.K.Fu)、红皮云杉(Picea koraiensis Nakai)和臭冷杉(Abies nephrolepis (Trautv. ex Maxim.) Maxim.)等针叶树种,阔叶树种主要为紫椴(Tilia amurensis Rupr.)、色木槭(Acer mono Maxim.)、蒙古栎(Quercus mongolica Fisch. ex Ledeb)、枫桦(Betula costata Trautv.)、春榆(Ulmus davidiana var. japonica (Rehd.) Nakai)和3大硬阔树种(水曲柳(Fraxinus mandshurica Rupr.)、黄檗(Phellodendron amurense Rupr.)、胡桃楸(Juglans mandshurica Maxim.))等。由于立地条件的差异,阔叶红松林受到干扰后衍生出的次生林的演替过程可分为3类:旱生、中生和湿生演替序列[50-51]。图2展示了温带阔叶红松林衍生出的主要次生林的3大演替序列。

图 2 阔叶红松林在干扰下的次生演替序列
Figure 2. Secondary succession of broad-leaved Korean pine forest under disturbance
次生林旱生演替一般发生在立地条件比较苛刻的地段,如地势较高的岗脊和阳向陡坡地段,一般水分不足而光照充足,蒙古栎、山杨(Populus davidiana Dode)和落叶松对此环境的耐受性较强,在阔叶红松林受到严重干扰后,可以顺利的从灌草群落中生长发育,会初步形成山杨-蒙古栎林和落叶松-蒙古栎林等初期阶段群落;蒙古栎、山杨和落叶松成林后,改善了林下环境,黑桦等阔叶树种也相继进入群落,而由于山杨和落叶松的幼苗不耐荫蔽,逐步退出森林群落[51],在轻度干扰和无红松种源的条件下,群落收敛于蒙古栎主导的各种派生的蒙古栎林等中期阶段群落;在轻度干扰和有红松种源的条件下,红松进入森林群落,由于红松的寿命较长,在林内其他林木死亡后,会逐步占据主林层,而蒙古栎由于具有较强的萌蘖能力,在老树死亡后,也可迅速形成簇生的小树,因此最终会演替形成蒙古栎和红松为主的后期阶段群落——阔叶红松林[52]。
次生林湿生演替发生的立地条件与旱生演替相反,一般在山坡下腹、谷地和阴坡地段上,林地内排水不畅,生境潮湿,由于白桦和落叶松对此环境的较强适应性,在阔叶红松林受到严重干扰后形成的灌草群落中可以较早地更新发育,进而形成白桦林、落叶松林和白桦-落叶松林等初期阶段群落;在轻度干扰下,白桦、落叶松初步改善了林分内土壤的水分条件,使耐湿的水曲柳和胡桃楸等硬阔叶树种得以进入森林群落,白桦、落叶松由于其幼苗喜光,在郁闭的林分中缺乏更新而逐步淘汰,当缺少红松种源,群落在轻度干扰下收敛于不同树种组成的软硬阔混交林或硬阔叶混交林等中期阶段群落,在落叶松占优势的地段,则会形成针阔混交林;若存在红松种源,在仅受轻度干扰情况下,红松在其他树种死亡后可以占据主林层,但红松死后,其他阔叶树种又得以生长,最终形成以红松、水曲柳和胡桃楸等树种为主的后期阶段群落——阔叶红松林[18]。
次生林中生演替不同于旱生演替和湿生演替,常在山地的中下腹和缓坡地带发生,其演替发生的立地条件也比较适中。阔叶红松林受到严重干扰后形成灌草群落,以白桦和山杨为主的先锋树种在轻度干扰下会率先入侵,形成白桦-山杨林和白桦-落叶松林等初期阶段群落;在轻度干扰下,先锋树种群落改善了林内光照、水分环境后,其他耐阴的紫椴、枫桦和色木槭等阔叶树种开始入侵森林群落,在缺少红松种源的情况下,群落内林木经过多代的死亡与更新,演替收敛于不同树种组成的软、硬阔混交林或硬阔叶混交林等中期阶段群落;而在有红松种源的地段,轻度干扰下的森林群落,红松会逐步占据主林层,而在红松死亡后,其他的阔叶树种则会在林隙内再次回归主林层,最终形成以红松、紫椴、枫桦和色木槭等树种为主的后期阶段群落——阔叶红松林[18]。
虽然将阔叶红松林衍生次生林的演替过程划分为3种类型,但由于山地地形的复杂性和种子的相互传播,这3种演替过程经常会相互重叠,形成多个树种优势度相当的阔叶混交林、针阔混交林,直到演替至红松占优势的阔叶红松林群落。
-
山地云冷杉林主要分布在小兴安岭-长白山林区的较高海拔地带,位于水平地带性群落阔叶红松林林带的上限,是亚高山针叶混交林带的地带性植被[53]。群落树种组成较为单一,以鱼鳞云杉为主,其次是臭冷杉,在排水稍差的地方还有红皮云杉分布,除了针叶树种外,群落间或混生少量的岳桦(Betula ermanii Cham.)、枫桦或花楸(Sorbus dacica Borbás)等阔叶树种。图3展示了山地云冷杉林受到干扰后形成的主要次生林的生态演替序列。
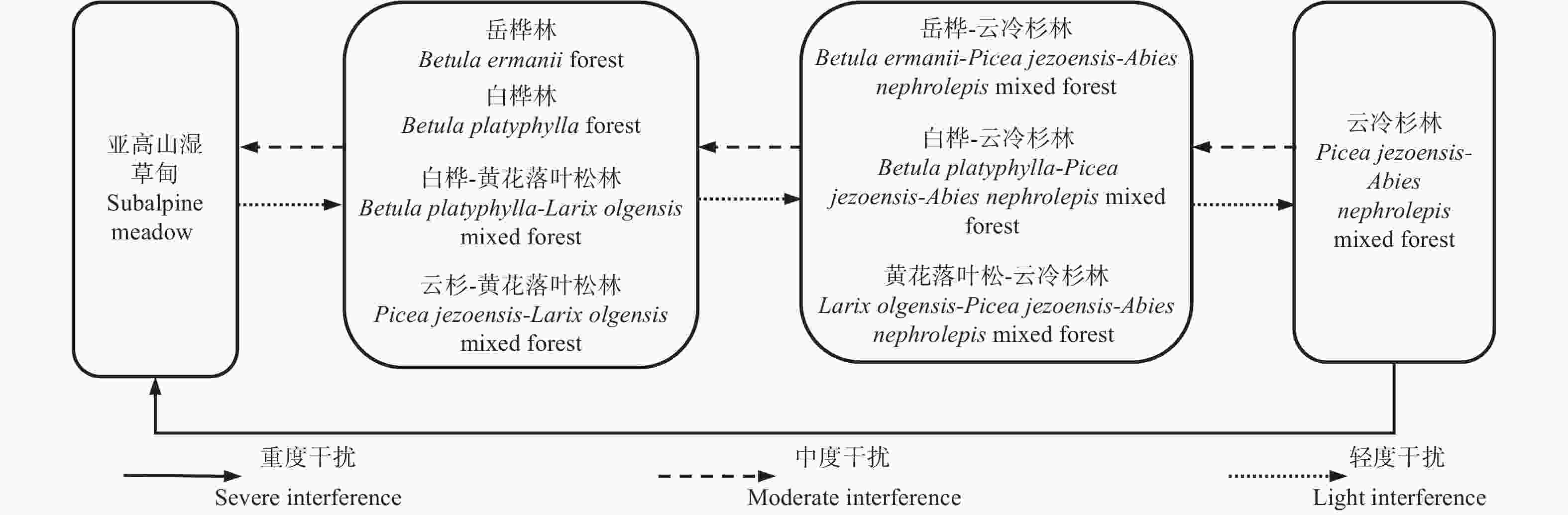
图 3 山地云冷杉林在干扰下的次生演替序列
Figure 3. Secondary succession of montane spruce-fir forest under disturbance
云冷杉林一经干扰破坏,林内光照充足,阳性先锋树种黄花落叶松(Larix olgensis A. Henry)、白桦和岳桦就会侵入群落,当干扰严重致使云冷杉全部死亡或被采伐,则会形成白桦林、岳桦林和白桦-黄花落叶松林等演替初期的先锋群落,其中岳桦的抗风性较其他先锋树种强,因此岳桦林主要分布在高海拔冲风地带,不甚普遍,其他先锋群落则较为常见[53]。先锋群落在轻度干扰下郁闭成林后,耐阴性的云杉、冷杉侵入林下,并在一部分白桦、黄花落叶松、岳桦死亡后,得以进入主林层,演替为岳桦-云冷杉林、白桦-云冷杉林和黄花落叶松-云冷杉林等中期阶段群落;在保持轻度干扰条件下,由于岳桦、白桦和黄花落叶松等先锋树种的寿命不及云杉和冷杉,且云冷杉进入主林层后会将林内小气候推向阴湿状况,先锋树种的幼苗、幼树很难适应而逐渐死亡,群落就进入到演替后期阶段的云冷杉林。当云冷杉林受到的干扰程度较为严重,又缺乏先锋树种种源的情况下,会发展成为以小叶章(Deyeuxia angustifolia (Kom.) Y. L. Chang)为主的亚高山湿草甸,杂草高度1.5~1.7 m,严重影响了云冷杉林的自然更新过程[22],在出现了有利于演替进行的轻度干扰后,由于云杉对于极端条件的耐受度比冷杉强,在潮湿甚至似沼泽的地区,以及更干燥的地区都可以占据优势,因此会形成散生单木的云杉-落叶松林,轻度干扰下,逐代更新后郁闭成林,林下开始侵入冷杉,并进入主林层,落叶松由于幼苗耐阴性差,缺乏更新而被演替淘汰,最终群落演替至顶级云冷杉林[18],但整个过程会非常漫长,必要时需要介入有利的人为干扰,保证云杉和冷杉幼树成活率,并促进其生长[22]。中期阶段的群落若受到中度干扰,云杉和冷杉被采伐,则会逆行演替至初期的先锋群落。
虽然各个次生林的演替进程在物种组成或演替时长上存在差异,也可能经历不完全的演替进程,停滞在其中的某个阶段,但次生林的演替序列几乎都可分为3个主要阶段:①初期:原始林受到严重干扰被破坏,先锋树种侵入形成次生林:②中期:林分形成后改变了原来环境条件,一些更适宜树种再次侵入,并逐渐形成新的林分;相反,林分进一步破坏,则向着结构简单、组成单一、生态条件愈趋恶化的方向发展;③后期:林分向着原地带性植被方向发展,或向着偏途顶极方向逆行发展[54]。
-
对于我国东北大面积的次生林,如何将次生林演替规律和干扰作用融入次生林经营,加快次生林的演替,提高次生林的稳定性,是广大学者关注的重点。自上个世纪50年代以来,历代的林业工作者探索出“栽针保阔”这一基于演替理论的科学论断来指导次生林的恢复演替,经过几十年的实验,验证了施以正向人为干扰措施对于次生林进展演替的有效性[19, 32]。
对于东北地区类型多样的次生林,其适合的干扰促进措施也一定是多样的。在介入经营干扰因素之前,最重要的就是确定次生林的来源,即该次生林群落衍生于何种原始顶级群落,然后判断次生林受干扰的现状,特别是顶级树种的径级结构是否合理、更新与种源是否充足,最后基于该顶级群落的演替序列和次生林的干扰现状指导森林经营。具体措施在于判断干扰现状:(1)若顶级树种大树多更新多,说明群落处于演替后期阶段,实施控制性火干扰可减少林下可燃物储量,避免森林大火的产生引起逆行演替;(2)若顶级树种大树多更新少,则说明可能发生过火灾烧死了幼树幼苗和种子或采摘松果带走了种源,又或是灌草群落茂盛限制了幼苗成活,群落处于向中期阶段演替的逆行演替状态,但由于大树多种源充足,减少松果采摘、对灌草群落进行砍伐,即可依赖自然更新恢复较多的幼苗数量,在新的幼苗长成幼树后,适当的疏伐上层非珍贵树种促进小树成长,即可维持合理的径级结构,促进群落正向演替;(3)若顶级树种大树少更新多,说明演替处于中期阶段,顶级树种种源充足,但可能在生长存活方面受到限制,在更新幼树旁伐除上层先锋树种或非珍贵树种可促进幼树更快进入主林层,促进演替进行;(4)如顶级树种大树少更新少,说明演替处于前期阶段,此时种源若是充足,则可以使用控制性火干扰或砍伐清除林下灌草,促进顶级树种种子萌发,若种源不足,则需要人工补植顶级树种,并适时采伐周围上层先锋树种,促进演替进入下一阶段。
-
干扰与次生林演替的关系涉及到多种生态学原理和林学基础理论,特别是干扰原理、演替原理以及近自然经营理论,“栽针保阔”经营措施就是在这样的科学理论指导下孕育而生。“栽针保阔”经营措施主要包括采伐林木和补植幼树两个过程[19],采伐林木涉及到作业法类型、采伐强度和采伐时间间隔的选择和控制,补植幼树也涉及补植数量、补植位置以及再补植时间间隔的配置选择。目前而言,“栽针保阔”实验取得初步成效,但研究成果并不全面[25, 32-33],如不同树种组成下“栽针保阔”的经营效果差异还有待进一步研究,且补植幼树目前仅遵循“见缝插针”的补植模式,密度制约效应和生物多样性原理均会对补植幼树的生长与存活产生影响,从生态学机理上解释“栽针保阔”后补植幼树的生长存活差异可以更好的指导次生林的恢复演替经营。
东北地区的次生林分布面积广、类型多,单一的“栽针保阔”经营干扰措施并不适用于所有次生林的恢复演替进程,探索因地制宜的次生林干扰模式,是精准促进东北次生林演替的必行之路。另外,演替是一个漫长的过程,我们现在不可能对改造后的次生林进行长达上百年的跟踪调查和后续经营,因此通过研究探讨适宜次生林演替序列内不同演替阶段的经营干扰措施,可以构建次生林恢复演替的全周期经营体系。
干扰不仅包括人为干扰,还包括自然干扰,目前关于东北次生林干扰与演替的关系研究多基于人为干扰,对自然干扰的作用影响研究较少,在气候变化的背景下,极端天气增多,掌握多种自然干扰间的相互作用和复合干扰对演替影响的作用机理[55-56],分析自然干扰后的次生林现状以及随之而来的后续影响,探索实施科学积极的人为干扰,同样是未来需要关注的方向。
东北主要天然次生林干扰与演替规律
Disturbances and Succession Laws of Main Natural Secondary Forests in Northeast China
-
摘要: 对东北地区兴安落叶松林、阔叶红松林和山地云冷杉林三大地带性顶级群落衍生出的主要次生林的生态演替过程规律,以及干扰在次生林演替过程中的作用影响进行了综述。结果显示人为干扰对次生林演替的作用有利有弊,但大部分的自然干扰不利于次生林进展演替,利用好采伐、补植和火烧等人为干扰可以有效地促进次生林的演替进程,次生林演替全周期经营和复杂的自然干扰对东北次生林演替的作用机制是未来进一步的研究重点。Abstract: Due to the excessive logging in the early stages, the natural forests in Northeast China were damaged to varying degrees, thus various types of secondary forests were formed. With the implementation of Natural Forest Protection Project, high-intensity commercial logging has been banned, so that most of the natural secondary forests in Northeast China have been restored to a certain content, but the whole process is slow. Understanding the succession law of secondary forest, using the ecological function of disturbance, and speeding up the succession process of the secondary forest through forest management measures has become the consensus of degraded secondary forest restoration. In this review, we introduce the ecological succession process of the main secondary forests derived from the three zonal top communities of Larix gmelinii forest, broadleaved Pinus koraiensis forest and Mountain spruce-fir forest in Northeast China, as well as the influence of interference on the succession process of secondary forests. It shows that artificial interferences may have both advantages and disadvantages on the succession of secondary forests, but most natural interferences are not conducive to the succession of secondary forests. Making good use of artificlal interferences such as logging, replanting and fire can effectively promote the succession process of secondary forests. Full cycle management of secondary forest successionandthe mechanism of natural disturbance on the succession of secondary forests in Northeast China is the focus of further research in the future.
-
Key words:
- Northeast China
- / natural secondary forest
- / succession
- / disturbance
- / forest management
-
图 1 兴安落叶松林在干扰下的次生演替序列
Figure 1. Secondary succession of The Larix gmelinii forest under disturbance
图 2 阔叶红松林在干扰下的次生演替序列
Figure 2. Secondary succession of broad-leaved Korean pine forest under disturbance
-
[1] Oberleitner F, Egger C, Oberdorfer S, et al. Recovery of aboveground biomass, species richness and composition in tropical secondary forests in SW Costa Rica[J]. Forest Ecology and Management, 2021, 479: 118580. doi: 10.1016/j.foreco.2020.118580 [2] Fao, Global Forest Resources Assessment (FAO, 2015)[R]. Roma: Food and Agriculture Organization of the United Nations, 2015. [3] Jiaojun Z, Mao Z, Hu L, et al. Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern China[J]. Journal of Forest Research, 2007, 12: 403-416. doi: 10.1007/s10310-007-0033-9 [4] Yan Q, Jiaojun Z, Gang Q. Comparison of spatial patterns of soil seed banks between larch plantations and adjacent secondary forests in Northeast China: implication for spatial distribution of larch plantations[J]. Trees, 2013, 27: 1747-1754. doi: 10.1007/s00468-013-0920-y [5] Holz S, Placci G, Quintana R D. Effects of history of use on secondary forest regeneration in the Upper Parana Atlantic Forest (Misiones, Argentina)[J]. Forest Ecology & Management, 2009, 258(7): 1629-1642. [6] Wang Y, Ziv G, Adami M, et al. Upturn in secondary forest clearing buffers primary forest loss in the Brazilian Amazon[J]. Nature Sustainability, 2020, 3(4): 1-6. [7] Chen X, Wang X, Li J, et al. Species diversity of primary and secondary forests in Wanglang Nature Reserve[J]. Global Ecology and Conservation, 2020, 22: e01022. doi: 10.1016/j.gecco.2020.e01022 [8] Muñoz Mazon M, Klanderud K, Finegan B, et al. How forest structure varies with elevation in old growth and secondary forest in Costa Rica[J]. Forest Ecology and Management, 2020, 469: 118191. doi: 10.1016/j.foreco.2020.118191 [9] 全 锋, 周超凡, 段光爽, 等. 基于蓄积生长率的蒙古栎天然次生林抚育间伐研究[J]. 林业科学研究, 2020, 33(2):61-68. [10] Deliang L, Lauren S. P, Dapao Y, et al. Differential responses of tree species to a severe ice storm and their implications to forest composition in the southeast United States[J]. Forest Ecology and Management, 2020, 468: 118177. doi: 10.1016/j.foreco.2020.118177 [11] 朱教君, 刘世荣. 次生林概念与生态干扰度[J]. 生态学杂志, 2007, 26(7):1085-1093. doi: 10.3321/j.issn:1000-4890.2007.07.022 [12] Christensen N, Peet R. Convergence during secondary forest succession[J]. The Journal of Ecology, 1984, 72: 25-36. doi: 10.2307/2260004 [13] 左 政, 郑小贤. 不同干扰等级下常绿阔叶次生林林分结构及树种多样性[J]. 浙江农林大学学报, 2019, 36(1):21-30. doi: 10.11833/j.issn.2095-0756.2019.01.004 [14] Chokkalingam U, De Jong W. Secondary forest: A working definition and typology[J]. International Forestry Review, 2001, 3(1): 19-26. [15] Finegan B. Forest succession[J]. Nature, 1984, 312: 109-114. doi: 10.1038/312109a0 [16] 周灿芳. 植物群落动态研究进展[J]. 生态科学, 2000, 19(2):53-59. doi: 10.3969/j.issn.1008-8873.2000.02.010 [17] 徐文铎, 何兴元, 陈 玮, 等. 长白山植被类型特征与演替规律的研究[J]. 生态学杂志, 2004, 23(5):162-174. doi: 10.3321/j.issn:1000-4890.2004.05.031 [18] 周以良, 赵光仪. 小兴安岭—长白山林区天然次生林的类型、分布及其演替规律[J]. 东北林业大学学报, 1964,(3):33-45. [19] 陈大珂, 周晓峰, 丁宝永, 等. 黑龙江省天然次生林研究(Ⅰ)——栽针保阔的经营途径[J]. 东北林业大学学报, 1984, 12(4):1-12. [20] Prévost M. Effect of cutting intensity on microenvironmental conditions and regeneration dynamics in yellow birch - Conifer stands[J]. Canadian Journal of Forest Research, 2008, 38: 317-330. doi: 10.1139/X07-168 [21] 红 玉, 王 耀, 金 鑫, 等. 抚育经营对杨桦次生林林分结构及土壤性质的影响[J]. 西北林学院学报, 2018, 33(3):67-72. doi: 10.3969/j.issn.1001-7461.2018.03.11 [22] 李玉祥, 焦振英, 张富田, 等. 寒温带云冷杉林次生演替成亚高山草甸人工更新技术的研究[J]. 东北林业大学学报, 1994, 22(4):40-45. [23] Prévost M, Charette L. Selection cutting in a yellow birch–conifer stand, in Quebec, Canada: Comparing the single-tree and two hybrid methods using different sizes of canopy opening[J]. Forest Ecology and Management, 2015, 357: 195-205. doi: 10.1016/j.foreco.2015.08.003 [24] 梁立东, 李明文. 抚育对黑河地区典型天然次生林群落结构的影响[J]. 防护林科技, 2020,(7):1-3+33. [25] Dai L, Shao G, Chen G, et al. Forest cutting and regeneration methodology on Changbai Mountain[J]. Journal of Forestry Research, 2003, 14(1): 56-60. doi: 10.1007/BF02856763 [26] 姚陈健. 伪满时期东北农林生态变迁研究[D]. 辽宁锦州: 渤海大学, 2019. [27] Ram D, Lindström Å, Pettersson L, et al. Forest clear-cuts as habitat for farmland birds and butterflies[J]. Forest Ecology and Management, 2020, 473: 118239. doi: 10.1016/j.foreco.2020.118239 [28] 张海军, 张淑兰, 王长宝. 小兴安岭红松种群天然更新及影响因子的探讨[J]. 林业资源管理, 2015,(2):150-153. [29] 朴春寿, 赵建卓, 李少华, 等. 林间放牧对森林影响的调查与思考[J]. 吉林林业科技, 2012, 41(1):53-54. doi: 10.3969/j.issn.1005-7129.2012.01.019 [30] 金光泽, 杨桂燕, 马建章, 等. 松果采摘对小兴安岭主要林型红松土壤种子库和幼苗库的影响[J]. 自然资源学报, 2010, 25(11):1845-1854. doi: 10.11849/zrzyxb.2010.11.004 [31] Długosiewicz J, Zając S, Wysocka-Fijorek E, et al. Comparative analysis of natural and artificial regeneration in Nowa Dęba Forest District[J]. Folia Forestalia Polonica, 2019, 61: 230-241. doi: 10.2478/ffp-2019-0022 [32] 丁 磊, 胡万良, 丁国泉, 等. 近自然森林经营在辽东山区次生林恢复中的应用效果评价[J]. 东北林业大学学报, 2013, 41(3):30-34. doi: 10.3969/j.issn.1000-5382.2013.03.009 [33] 柏广新, 牟长城. 抚育对长白山幼龄次生林群落结构与动态的影响[J]. 东北林业大学学报, 2012, 40(10):48-55. doi: 10.3969/j.issn.1000-5382.2012.10.012 [34] 李秀珍, 王绪高, 胡远满, 等. 林火因子对大兴安岭森林植被演替的影响[J]. 福建林学院学报, 2004, 24(2):182-187. doi: 10.3969/j.issn.1001-389X.2004.02.022 [35] 马楠楠, 张彦雷, 李 建, 等. 黑龙江呼玛县森林火灾时空分布特征[J]. 东北林业大学学报, 2016, 44(5):20-23. doi: 10.3969/j.issn.1000-5382.2016.05.005 [36] 赵大昌. 长白山火山爆发和植被发展演替关系的初步探讨[J]. 自然资源学报, 1984,(1):72-78. [37] 董厚德, 唐炯炎. 辽东山地“乱石窖”植被演替规律的初步研究[J]. 植物生态学与地植物学丛刊, 1965, 3(1):117-130. [38] 李 威, 周 梅, 赵鹏武, 等. 大兴安岭东麓火烧迹地恢复初期植被特征[J]. 东北林业大学学报, 2020, 48(1):51-55. doi: 10.3969/j.issn.1000-5382.2020.01.009 [39] 何 潇, 李海奎, 曹 磊, 等. 退化森林生态系统中林分碳储量的驱动因素—以内蒙古大兴安岭为例[J]. 林业科学研究, 2020, 33(2):69-76. [40] 李秀芬, 朱教君, 王庆礼, 等. 森林的风/雪灾害研究综述[J]. 生态学报, 2005, 25(1):148-157. doi: 10.3321/j.issn:1000-0933.2005.01.024 [41] Nagel T, Diaci J. Intermediate wind disturbance in an old-growth beech-fir forest in southeastern Slovenia[J]. Canadian Journal of Forest Research, 2011, 36: 629-638. [42] 李秀芬, 朱教君, 王庆礼, 等. 辽东山区天然次生林雪/风灾害成因及分析[J]. 应用生态学报, 2004, 15(6):941-946. doi: 10.3321/j.issn:1001-9332.2004.06.006 [43] 程肖侠, 延晓冬. 气候变化对中国大兴安岭森林演替动态的影响[J]. 生态学杂志, 2007, 26(8):1277-1284. [44] 张晓玉, 田晓瑞. 厄尔尼诺/拉尼娜对大兴安岭森林火险天气的影响[J]. 林业科学研究, 2018, 31(6):55-62. [45] Taylor A, Endicott S, Hennigar C. Disentangling mechanisms of early succession following harvest: implications for climate change adaptation in Canada's boreal-temperate forests[J]. Forest Ecology and Management, 2020, 461: 117926. doi: 10.1016/j.foreco.2020.117926 [46] 陈利顶, 傅伯杰. 干扰的类型、特征及其生态学意义[J]. 生态学报, 2000, 20(4):581-586. doi: 10.3321/j.issn:1000-0933.2000.04.008 [47] 顾云春. 中国的兴安落叶松林[J]. 林业资源管理, 1982,(2):27-30. [48] 顾云春. 大兴安岭林区森林群落的演替[J]. 植物生态学与地植物学丛刊, 1985, 9(1):64-70. [49] 王绪高, 李秀珍, 贺红士, 等. 大兴安岭北坡落叶松林火后植被演替过程研究[J]. 生态学杂志, 2004, 23(5):35-41. doi: 10.3321/j.issn:1000-4890.2004.05.007 [50] 王义弘. 帽儿山地区次生林的天然更新和演替[J]. 东北林业大学学报, 1984, 12(A1):39-46. [51] 陆龙龙. 长白山林区阔叶红松林不同演替阶段群落结构特征研究[D]. 吉林:北华大学, 2019. [52] 陈大珂, 周晓峰, 祝 宁. 天然次生林——结构、功能、动态与经营[M]. 哈尔滨: 东北林业大学出版社, 1994. [53] 周以良, 李景文. 中国东北东部山地主要植被类型的特征及其分布规律[J]. 植物生态学与地植物学丛刊, 1964, 2(2):190-206. [54] 朱教君. 次生林经营基础研究进展[J]. 应用生态学报, 2002, 13(12):1689-1694. doi: 10.3321/j.issn:1001-9332.2002.12.040 [55] Buma B. Disturbance interactions: characterization, prediction, and the potential for cascading effects[J]. Ecosphere, 2015, 6(4): 1-15. [56] Cannon J, Peterson C, O'brien J, et al. A review and classification of interactions between forest disturbance from wind and fire[J]. Forest Ecology and Management, 2017, 406: 381-390. doi: 10.1016/j.foreco.2017.07.035 -

 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 5967
- HTML全文浏览量: 3765
- PDF下载量: 140
- 被引次数: 0
