





-
毛竹(Phyllostachys edulis (Carrière) J. Houz.)是一种具有重要经济和生态意义的非木材林产品。在毛竹生长过程中,非生物胁迫条件严重制约了其产量和品质[1]。近年来,毛竹基因组测序工作的完成,为从分子水平探讨毛竹抗逆调控方式提供了研究依据。目前在毛竹中,已分离和鉴定出很多参与毛竹非生物胁迫反应相关的功能基因。Wu等研究发现,毛竹中的LBD类转录因子PheLBD29的表达量被PEG、NaCl、ABA和MeJA显著诱导。干旱胁迫条件下,过表达PheLBD29的转基因植株通过降低相对电解质、丙二醛和过氧化物的积累来提高可溶性糖的含量,以及改变LEA、DREB2A、 RD22、 ERD15、Di19和RAB18等胁迫相关基因的表达量来提高转基因植物的耐旱性[2]。同样毛竹中的C2H2类转录因子PheDi19-8的表达量也被PEG、NaCl和ABA显著诱导,成苗期干旱胁迫条件下,转基因株系的存活率(60.34%) 显著高于野生型对照植株(36.7%) ;且在转基因植株中,生物量含量及可溶性糖含量的积累,LEA、 RD29A、DREB2A 和RD22基因的表达量得到了提高[3]。与过表达PheLBD29基因的拟南芥 ( Arabidopsis thaliana (L.) Heynh.) 不同的是,过表达PheDi19-8基因的植株对ABA并不敏感。Gao等研究发现在水稻 (Oryza sativa L.) 中过表达Phehdz1基因也可以显著提高植株对干旱胁迫的耐受性[4]。利用农杆菌转化法在拟南芥中过表达毛竹 WKRY 类转录因子 WKRY83和WKRY86,分别增强了转基因植物对盐和干旱胁迫的耐受性[5-6]。Liu等研究发现在拟南芥和水稻中过表达毛竹 TCP 类转录因子PheTCP10,促进了转基因植物叶片细胞中气孔的关闭,减少了水分的流失,提高了对干旱胁迫的耐受性[7];Xu等进一步地发现,PheTCP10通过调控BTB/TAZ基因(BT2)的表达,进而提高转基因植物对高盐环境的适应性[8]。 在水稻中超表达ASR类转录因子PheASR2,促进了水稻植株气孔关闭,降低了叶片失水速率,增加了抗旱性[9]。此外,毛竹中的PheC3H74、PheCPK1、PhePLATZ1、PeSNAC-1、PheTCP9等基因也被证实在植物的非生物胁迫反应中发挥着重要作用[10-14]。
从前期的研究结果,我们可以看出转录因子在调节毛竹非生物胁迫反应中发挥着重要作用[5-14]。MYB 转录因子家族是植物中最大的转录因子家族[15]。MYB 转录因子在 N 端都有一段保守的MYB 结构域,是 DNA 的结合区(DNA-binding domain)。MYB 转录因子的 C 端有一个转录激活区(transcription activation domain),通过折叠成双亲性的螺旋结构从而激活转录[16-17]。根据MYB 转录因子 N 端 R 结构(R1、R2、R3)的个数 , 可 以 将 MYB 转 录 因 子 大 致 分 为 R1/R2-MYB、R2R3-MYB、R1R2R3-MYB 三个亚类,其中 R2R3-MYB 是植物中数目最多的一类[18],目前已在很多物种中鉴定出与植物逆境相关的R2R3-MYB 转录因子。R2R3-MYB转录因子主要通过两个方面来正向或者负向地调控植物对外界逆境环境的适应性。一是通过与顺式作用元件特异结合实现对逆境的调控。如枸桔 (Poncirus trifoliata (L.) Raf.) 的 PtsrMYB 蛋白能够与ADC基因的启动子区域的 MYBCORE 元件相结合,调控ADC基因表达改变转基因植物体内多胺的含量,从而调控转基因植株的抗旱性[19]。杜梨 (Pyrus betulifolia Bunge) Raf.) 中PbrMYB21蛋白能够POD、SOD和P5CS基因的启动子结合,激活此类非生物胁迫反应基因的表达,从而调控转基因杜梨的抗旱性[20]。Cui等利用EMSA实验证明,AtMYB20能够与PP2Cs基因的启动子序列中的TAACTG元件结合,进而调节植物的耐盐性[21]。
二是R2R3-MYB转录因子通过参与到多种信号通路途径调控植物的耐旱性。大豆 (Glycine max (L.) Merr.) GmMYB84 通过调节转基因大豆中的活性氧物质(ROS)来调控主根的长度,进而提高转基因大豆的耐旱性[22]。拟南芥中过表达AtMYB15 增强了转基因植物对 ABA 的敏感性,且与ABA 合成、信号、响应相关基因的表达模式也得到了增强,表明 AtMYB15 可能通过 ABA 依赖的途径调控了植物对干旱胁迫的响应[23]。毛竹作为重要的非木材林,在毛竹的基因组中,已鉴定出114个R2R3-MYB转录因子[13]。到目前为止,毛竹R2R3-MYB相关转录因子在非生物胁迫中作用的研究不多,只报道了MYB2参与了非生物胁迫,对毛竹低温胁迫和盐胁迫的响应起着重要的作用[24]。然而,目前对毛竹中其它R2R3-MYB相关转录因子在非生物胁迫中的作用却知之甚少。
-
本实验所用毛竹种子来自于中国浙江省天目山国家级自然保护区,种植于温室培养,光周期(光/暗) 16 h/8 h,温度25 ºC,湿度 60%。
拟南芥种子由本实验室保存,种植于光周期(光/暗)为 16 h/8 h,温度为(光/暗) 22 ºC,湿度为 60%的温室中培养。
-
对三个月大小的毛竹种苗,分别浇灌20% PEG-6000和 200 mmoL·L−1 氯化钠溶液,同时选取生长一致的种苗分别放置在不同温度的(42 ℃和4 ℃)培养箱中,模拟高温和低温处理。分别在0、1、12、24、48 和 72 h时,在相同部位,剪取长势相似的叶片10片,每次 3 个生物学重复,−80 ℃保存。
对转基因拟南芥进行扩繁至 T3 代,将消毒后的 T3 代拟南芥OE-6、OE-8、OE-9和野生型株系种子在点铺在MS平板上进行萌发,且MS平板中添加不同浓度的甘露醇。同时,将消毒后的 T3 代拟南芥OE-6、OE-8和野生型株系种子在点铺在MS平板上进行萌发,萌发 10 d后选取大小一致的转基因株系和野生型幼苗移入营养土中生长。待幼苗生长3周时,停止供水,进行缺水干旱处理,分析转基因植株的抗旱表型。
-
毛竹PheMYB2R-4 (PH01000388G0380)的氨基酸序列从(
http://www.bamboogdb.org/ )网站上获取,从phytozome (https://phytozome-next.jgi.doe.gov/ )网站上,下载拟南芥、玉米 (Zea mays L.)、水稻和小麦 (Triticum aestivum L.) 的氨基酸序列,并在 DNAMAN 中进行氨基酸序列比对。 -
利用RNAiso Plus 试剂盒 (Takara货号:9108) 根据说明书提取 RNA,利用 PrimeScript™ RT Master Mix 试剂盒 (Takara货号:RR036A) 根据说明书反转录 RNA。荧光定量实验所用引物由 Primer Express 3.0 进行设计(表1) ,NCBI 网站验证特异性后由通用生物系统 (安徽) 有限公司合成,每对引物扩增产物长度为为 80 ~ 200 bp。使用 TransStart® Green qPCR Super Mix (货号:AQ101-01) 按照说明书实验,使用 2−△△CT计算表达量。TIP41 基因作为毛竹内参基因,Atactin2基因作为拟南芥内参基因。荧光定量引物见表1。
表 1 荧光定量引物
Table 1. Primer for qRT-PCR
基因
Gene引物名称
Primer Name引物序列 (5’-3’)
Primer sequence(5’-3’)TIP41 TIP41F AAAATCATTGTAGGCCATTGTCG TIP41R ACTAAATTAAGCCAGCGGGAGTG PheMYB2R-4 PheMYB2R-4F TTGGAAGCAGGTGGTCGATC PheMYB2R-4R AACCTGCAGCACTCGTAGTC Atactin2 Atactin2F AAGCTGGGGTTTTATGAATGG Atactin2R TTGTCACACACAAGTGCATCAT AtRD29A AtRD29AF TGAAAGGAGGAGGAGGAATGGTTGG AtRD29AR ACAAAACACACATAAACATCCAAAGT AtRD22 AtRD22F CAAACCCTTTCGTGTATAACTATGCA AtRD22R TTTCCCGCGAACCAAGTC AtDREB2A
AtDREB2AF GCGATTTTCATCTGGATCACATT AtDREB2AR GCAGGTTCCTCGCATCCTT AtLEA
AtLEAF GGCGGATTTGAGGGACGAAA AtLEAR GTTATGGTGGCCTGGAAGCT -
设计PheMYB2R-4的转录活性引物序列,F:5'-GgaattcATGGGAAGGGCTCCATGT-3')和R:5'- CGggatccTTACATCATCGTTACCTTCCC-3',通过 PCR 扩增获得两端分别带有 EcoR I和BamH I酶切位点的PheMYB2R-4 基因片段。以 pGBKT7 为基础载体,构建含有目的片段的重组载体,分析PheMYB2R-4的转录激活活性。同时设置阴性对照:pGBKT7空载体;阳性对照:pGBKT7-53 + pGADT7-T。使用热击法将构建好的载体质粒转化到农杆菌GV3101 菌株中。参照唯地生物公司的说明书 (货号: YC1002S),进行酵母的转化。分别取菌液 100 μL 涂布于单缺 (SD/Trp-)和三缺 (SD/Trp-/His-/Ade-/X-α-gal) 固体培养基进行培养 (30 ºC),观察分析并拍照记录。
-
设计 PheMYB2R-4的亚细胞定位引物序列,F:5'-GCtctagaATGGGAAGGGCTCCATGT-3'和R:5'- CGggatccCATCATCGTTACCTTCCC- 3',通过 PCR 扩增获得两端分别带有Xba I和BamH I 酶切位点的PheMYB2R-4 基因片段。以 pCAMBIA-1305为基础载体,构建含有目的片段的重组载体,分析PheMYB2R-4的亚细胞定位。同时设pCAMBIA-1305空载为对照组。分别吸取50 μL pCAMBIA-1305重组质粒、pCAMBIA-1305空载的农杆菌液于摇菌管中同时加入5 mL YEP、5 μL 50 mg·mL−1 Kan和5 μL 25 mg·mL−1 Rif 于恒温摇床中 28 ℃、220 r·min−1培养36 ~ 48 h。 然后吸取2 mL 到无菌的10 mL 无菌管中,常温,5 000 rpm,离心5 min;弃上清,用纯水将菌斑重悬,再次离心; 加入2 mL 纯水,用分光光度计测OD600值,计算,将到菌液浓度调到0.8 ~ 1.0;将菌液注射到生长健康3周的本式烟草叶片中,做好标记。黑暗培养36 ~ 48 h左右, 用激光共聚焦显微镜观察细胞中的GFP信号。
-
设计PheMYB2R-4的过表达载体引物序列,F:5'-GGcgtaccATGGGAAGGGCTCCATGT-3'和R:5'-GCtctagaTTACATCATCGTTACCTTCCC-3',通过 PCR 扩增获得两端分别带有Kpn I和Xba I酶切位点的PheMYB2R-4 基因片段。以 pCAMBIA-1301为基础载体,构建过表达载体。将构建好的重组菌液培养至 OD600 值达到 0.8 ~ 1.0 左右,同时配置转化 Buffer 溶液。处理液使用前还需加入表面活性剂 (Solarbio:627N021)。用转化 Buffer 溶液悬浮离心的菌体,制备侵染溶液,然后将整个花序浸泡在侵染液中 5 min,3 ~ 4 周后果荚成熟即可收取转基因 T0代种子。
-
将 T0 代种子进行消毒清洗发芽后,均匀涂布在含有潮霉素的 MS 培养基平板上。进一步对筛选的阳性植株进行 PCR 分子检测,提取阳性拟南芥植株基因组,并作为 PCR 扩增模板,用目的基因PheMYB2R-4序列引物扩增片段,鉴定转基因拟南芥是否转化成功,阴性对照为野生型拟南芥,阳性对照为含有PheMYB2R-4的农杆菌质粒。
-
叶片相对含水量 (RWC) 和相对电导率(REL)的测定方法按照Wu等的文献中的报道[3,5,9]。丙二醛 (MDA) 具体测定方法按照试剂盒 (Solarbio:BC0020) 说明书进行操作。
-
以毛竹 cDNA 为模板进行 PCR 扩增,结果获得一条约918 bp 的特异性条带,与预期目的基因大小为基本一致(图1A)。测序结果表明,目的片段大小918 bp,与毛竹基因组数据库PH01000388G0380编码区一致,该基因编码一个 305 aa 的蛋白,分子量为 33.95 kD,理论等电点 pI 为 8.4,根据Hou等对毛竹中R2R3-MYB转录因子的鉴定和分析,将该基因命名为 PheMYB2R-4[13]。

图 1 PheMYB2R-4基因克隆及进化树分析
Figure 1. PheMYB2R-4 gene cloning and phylogenetic analysis
-
干旱和高盐处理后,荧光定量结果显示PheMYB2R-4 的转录水平在检测的时间点的都表现出上调状态 (图2)。值得注意的是,PheMYB2R-4的表达在干旱处理72 h时是对照组CK的36倍 (图2A)。在高盐处理72 h时是对照组CK的11倍 (图2B)。然而在高温处理下,处理24 h之后,PheMYB2R-4的表达才开始上升,72 h时是对照组CK的3倍 (图2C)。低温处理下,与对照相比,PheMYB2R-4的表达在24 h之后才有差异(图2D)。
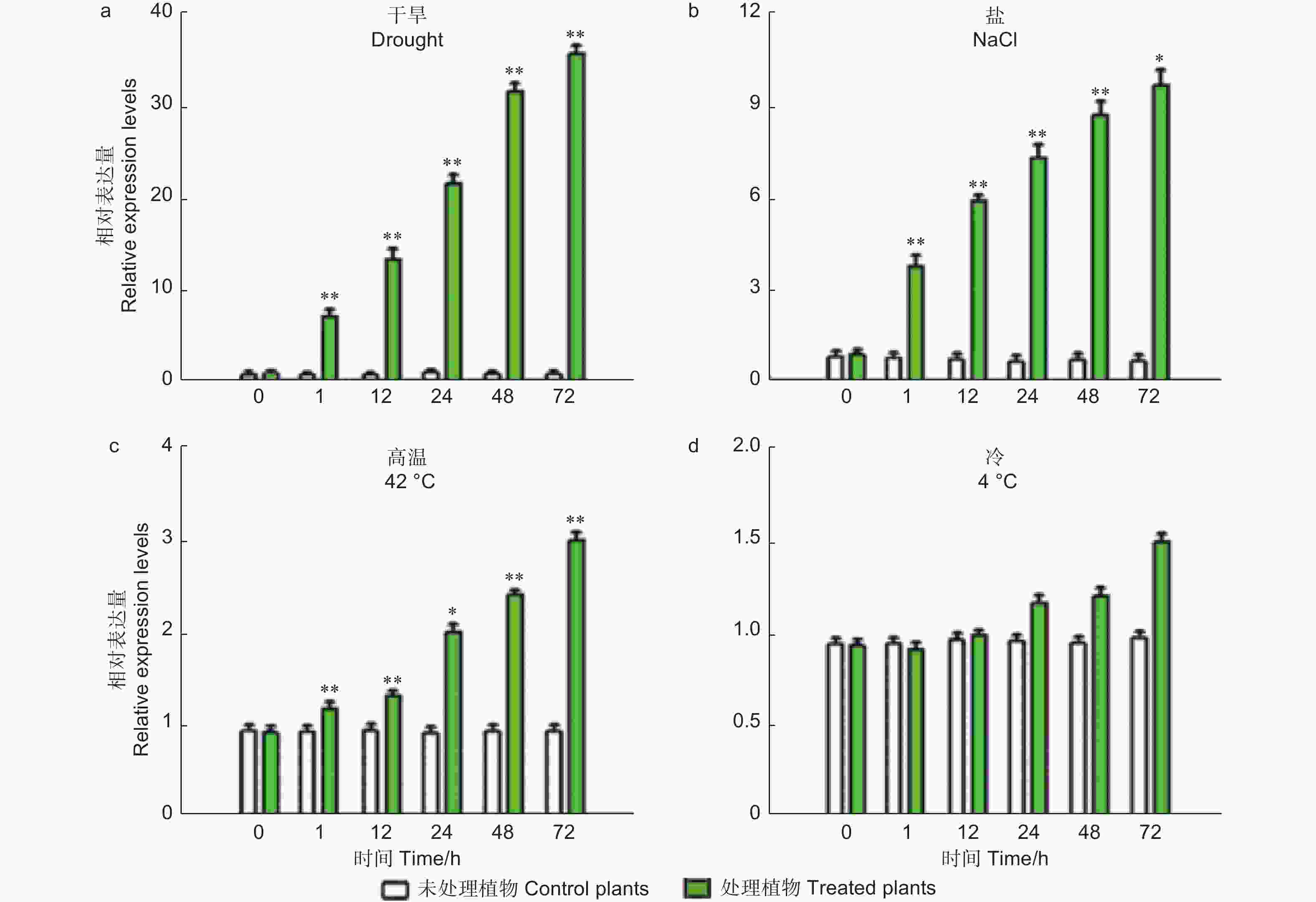
图 2 PheMYB2R-4在干旱、盐、高温及低温胁迫下的基因表达
Figure 2. Relative expression of PheMYB2R-4 in response to drought, salt, high temperature and low temperature treatments
-
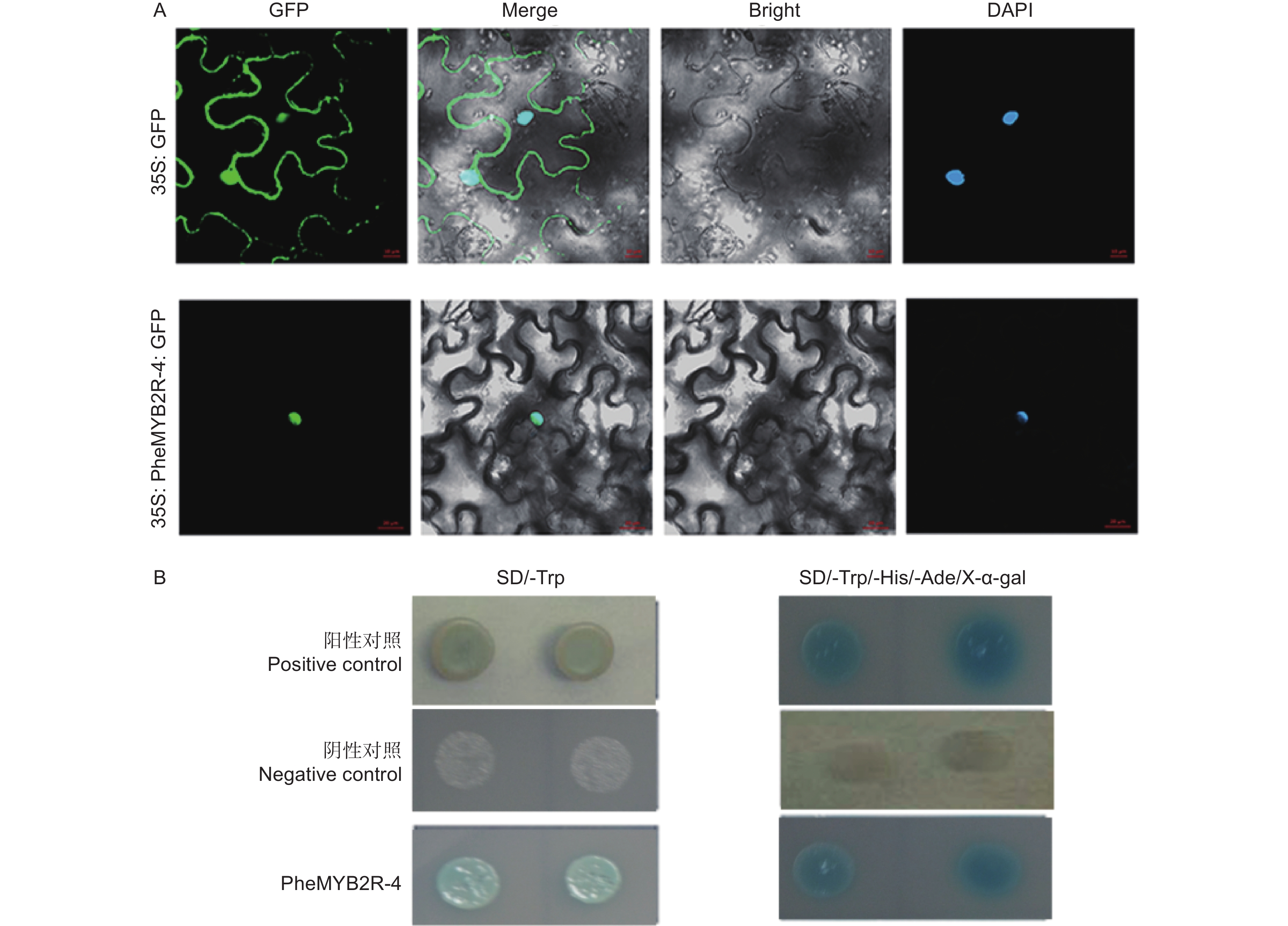
将构建成功的重组质粒 (p1305-35S-PheMYB2R-4-GFP) 分别导入农杆菌,同时将空载作为对照 (p1305-35S-GFP),侵染烟草瞬时表达,观察定位。结果如3A 所示:空白对照的 GFP 信号充满整个细胞中,而实验组(p1305-35S-PheMYB2R-4-GFP) 只在核内检测到信号,并且也与 DAPI 核定位信号相重合,说明PheMYB2R-4 是核定位蛋白。
通过 PEG/ LiAc 法将PheMYB2R-4 基因与 pGBKT7 的重组质粒,阴性对照组的空载 pGBKT7 和阳性对照组 pGBKT7-53 + pGADT7-T 转化至Y2H Gold 酵母中,在SD/-Trp 单缺板,缺陷培养基 SD/-Trp 和 SD/-Trp/-His/-Ade/X-α-gal 三缺平板上进行避光培养。结果如图3B所示:实验组和对照组的酵母在单缺板 SD/-Trp都可以正常生长,说明PheMYB2R-4 基因对酵母无毒性,可以在酵母系统中进行实验。然后再在含有 40 μg·mL−1 的 SD/-Trp/-His/-Ade 固体培养基上进行避光培养 3 ~ 5 d,发现实验组和阳性对照组酵母生长状况良好,并且变蓝色,说明 PheMYB2R-4在酵母菌株中具有自激活活性(图3B)。
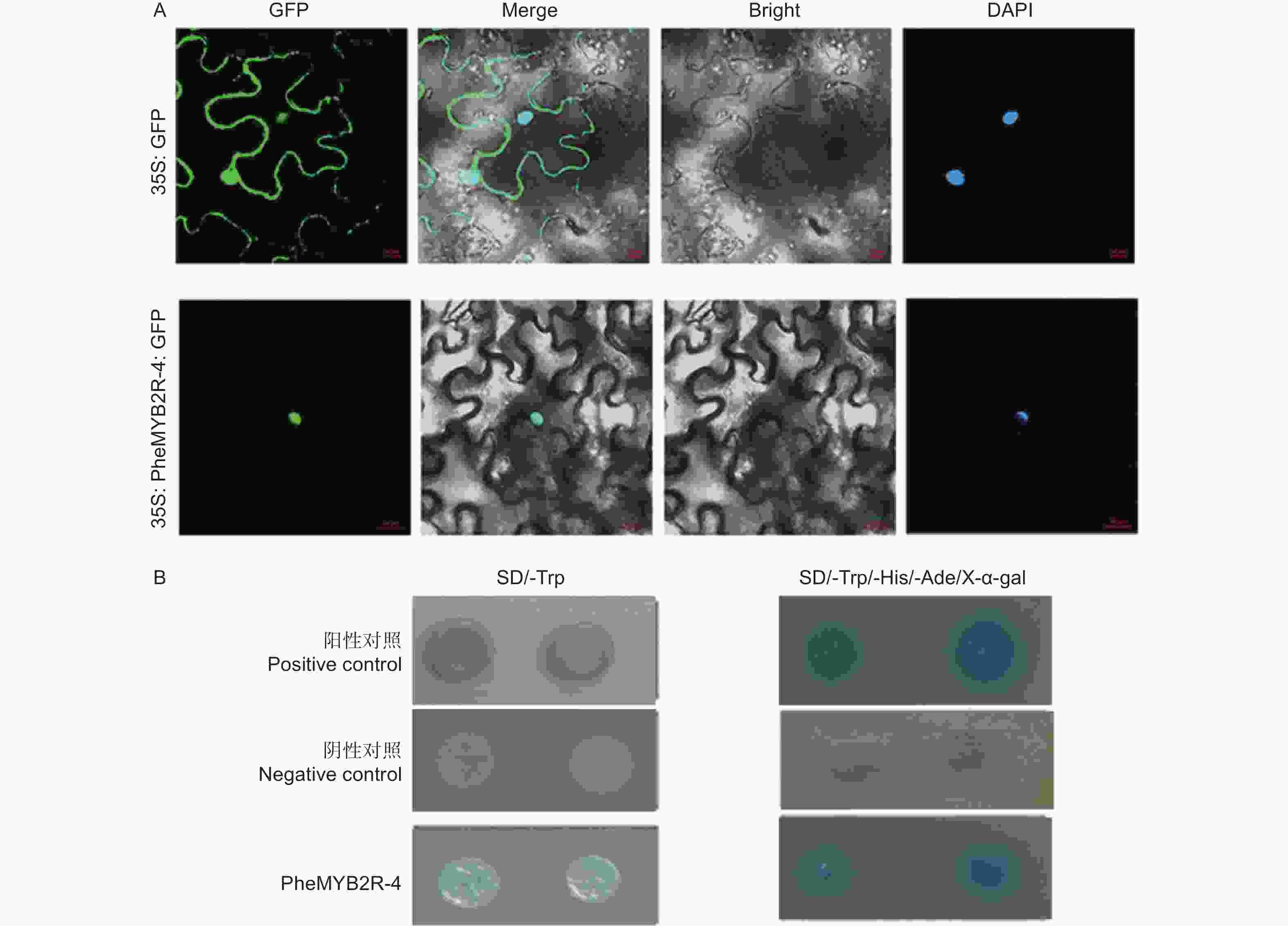
图 3 PheMYB2R-4的分子特征分析
Figure 3. Molecular characteristic analysis of PheMYB2R-4
-
对获得的转基因拟南芥,提取基因组,进行 PCR 克隆验证,并筛选出表达量较高的株系进行功能分析。对 T3 代转基因拟南芥和野生型植株种子在不同浓度的甘露醇 (0、125、200 mmoL·L−1) 溶液中进行萌发,分别于3 d和7 d 后统计萌发率。结果表明,当平板中不含有甘露醇溶液时,转基因植株和野生型拟南芥的萌发率无明显差异,而在含有125 mmoL·L−1和200 mmoL·L−1的甘露醇平板中,3 d时,转基因拟南芥的萌发率分别为32% 和8% 左右,而野生型拟南芥的萌发率分别为 18% 和4% 左右(图4B)。7 d时,转基因拟南芥的萌发率分别为为 68% 和40% 左右,而野生型拟南芥的萌发率分别为 56% 和22% 左右(图4C)。

图 4 野生型和转基因拟南芥在甘露醇下的萌发分析
Figure 4. Germination Analysis of wild type and transgenic Arabidopsis under Mannitol treatment
选取生长时期和状态一致的株系移栽到营养土中生长 3周左右,对转基因和野生型拟南芥进行干旱缺水处理。结果如图5A所示,在干旱处理前,转基因植株和野生型拟南芥生长无明显差别,在缺水处理10 d之后,发现野生型拟南芥较转基因植株受到更加明显的干旱胁迫,表现为叶片卷曲严重,生长受阻,叶片含水量 (RWC) 明显较低 (图5A、B),且野生型植株的绿叶存活率显著低于转基因株系 (图5C),表明转基因植株耐旱性较强。
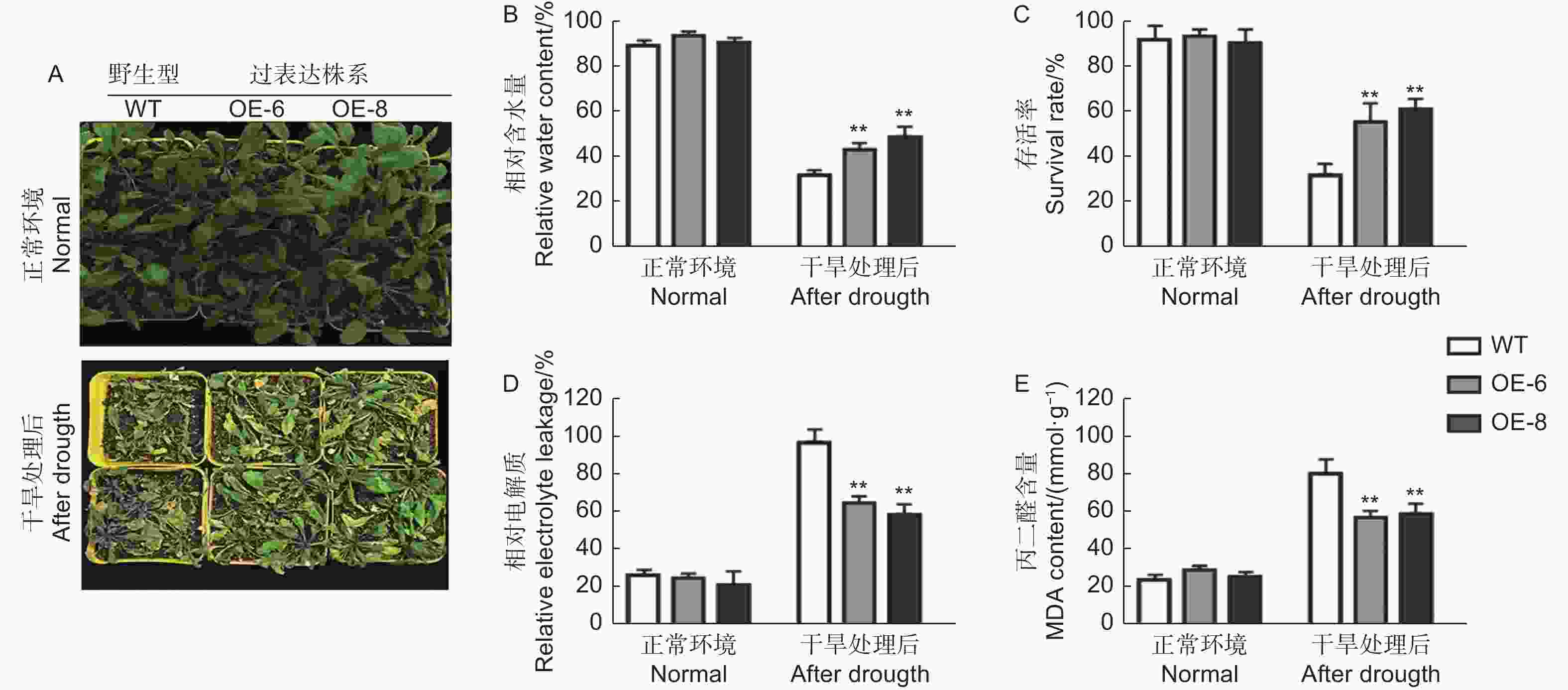
图 5 野生型和转基因拟南芥在干旱下的表型分析
Figure 5. Phenotypic Analysis of wild type and transgenic Arabidopsis under drought treatment
同时,对干旱处理前和处理后植株进行了抗逆相关生理生化指标的测定,重点对相对电导率 (REL) 以及丙二醛 (MDA) 含量进行了分析,如图5D和5E所示,转基因和野生型植株两个所测定的生理生化指标在处理前没有显著差异。在干旱处理后,转基因株系和野生型植株的相对电导率 (REL) 以及丙二醛 (MDA)含量都得到增加,野生型植株上升幅度显著高于转基因植株,进一步说明了转基因植株的耐旱性优于野生型。
-
通过对胁迫相关基因表达的检测发现,正常处理下的RD22 (responsive to dehydration 22)、RD29A (desiccation-responsive protein 29A)、DREB2A (dehydration-responsive element binding protein 2) 和LEA (dehydrin LEA) 在所有株系中的表达水平是相似的。相反,在缺水处理3 d下,转基因株系中的这些基因的表达水平被显著诱导,表明了过表达 PheMYB2R-4 能够调节胁迫响应相关基因的表达,进而提高植物的耐旱性 (图6)。
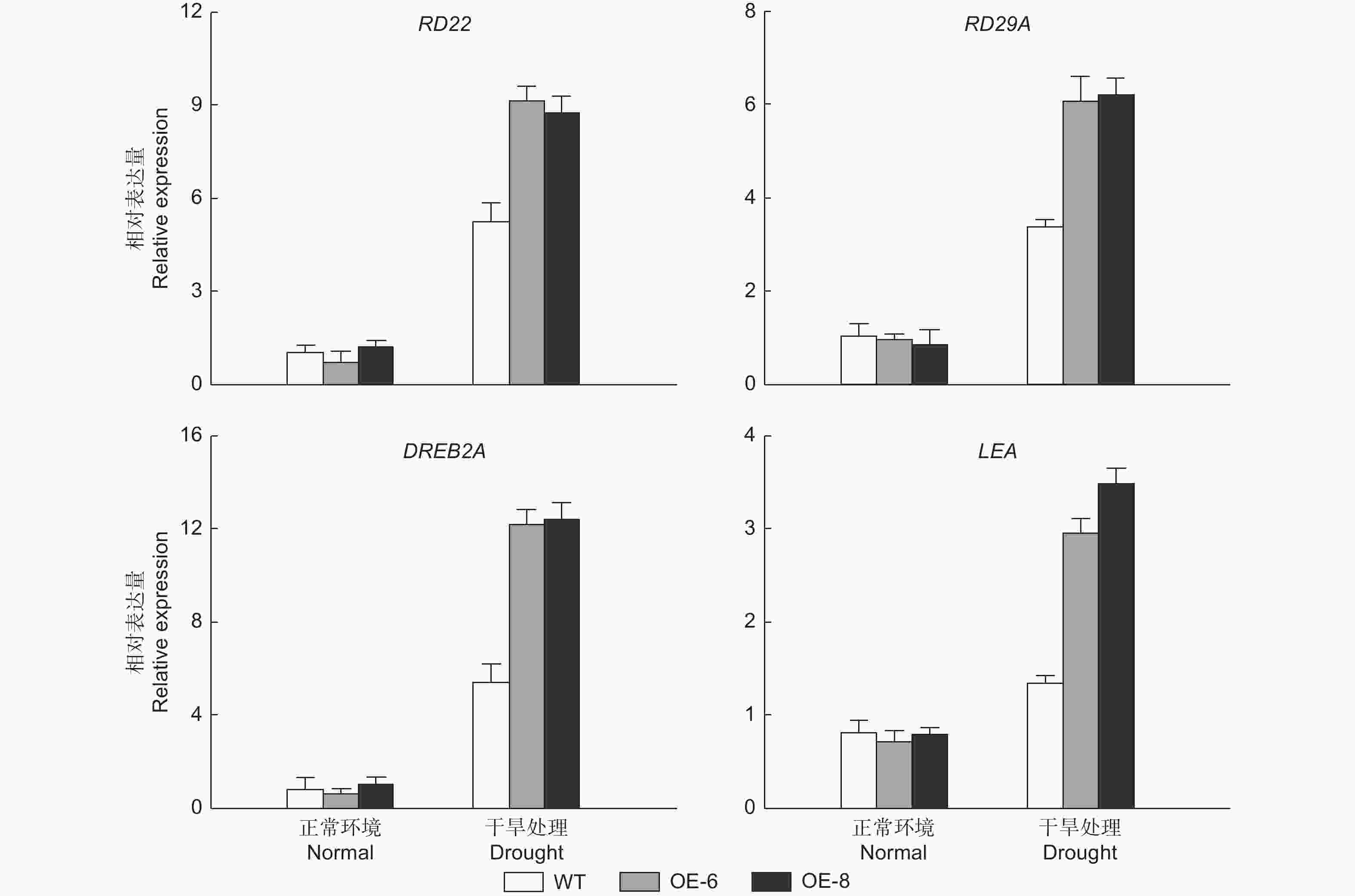
图 6 干旱相关基因的在野生型和转基因拟南芥中干旱处理前后的表达分析
Figure 6. Expression analysis of drought-related genes in wild type and transgenic Arabidopsis before and after drought treatment.
-
MYB 转录因子家族,其中R2R3亚类是目前研究较为广泛的一类MYB转录因子家族。R2R3亚类的MYB转录因子在植物的非生物胁迫反应中发挥着重要作用,目前已在农作物、林木、蔬菜、花卉等物种中已鉴定出相关的功能基因[3, 5, 8-9]。尽管2018年Hou等在毛竹的基因组数据中,鉴定出114个R2R3亚类的MYB转录因子,但是毛竹中关于R2R3亚类的MYB转录因子参与植物干旱胁迫反应的研究却鲜有报道。
本研究从毛竹中克隆1个 MYB基因,该基因全长为 918 bp (图1),编码 305个氨基酸,被命名为PheMYB2R-4,属于 MYB-R2R3基因家族[13]。进化树结果显示,PheMYB2R-4与水稻中的OsMYB55亲缘关系较近(图1)。Casaretto等研究发现,将OsMYB55基因超表达到玉米中,与对照植株相比,在干旱和高温条件下,转基因植株的叶绿素含量明显较高,说明过量表达OsMYB55基因提高了转基因植株对干旱和高温的耐性[25]。此外在本研究中,qRT-PCR 实验表明,PheMYB2R-4 的表达被 20% PEG6000 和200 mmoL·L−1 NaCl显著诱导(图2)。由此猜测 PheMYB2R-4可能参与植物参与植物的非生物胁迫反应。为了验证这一猜测,本研究分别开展了三个方面的工作。
首先,将 PheMYB2R-4 基因异源表达到拟南芥中进行功能研究。结果显示,过表达 PheMYB2R-4 转基因植物展现出了对干旱的耐受性 (图3和图4)。另外,PheMYB2R-4 在拟南芥中过表达对拟南芥生长发育而未产生影响,此种情况在PheDi9-8、PheCDPK1、PheC3H74、PheWRKY86等的转拟南芥实验中亦有报道[3, 6, 10-11]。
其次,干旱会损害植物的生理过程,进而会引起植株一系列的生理生化指标的改变[21]。叶片失水率 (RWC)是评价植物抗逆性的重要指标[26]。研究结果表明,在干旱条件下,转基因PheMYB2R-4 株系相比野生型植株具有更高的 RWC (图5B)。此外,作为评估细胞膜损伤的重要指标相对电导率 (REL) 和反映膜脂过氧化程度的丙二醛 (MDA) 含量[27-28] 。在我们研究中,过表达 PheMYB2R-4 株系的 REL 和 MDA 含量明显低于野生型,这表明转基因株系受到更轻微的膜损伤 (图5D和图5E)。这表明 PheMYB2R-4通过减轻膜损伤和减少水分流失来提高植物的抗旱性。
最后,前期大量研究表明,过量表达抗逆相关的转录因子通常是通过改变下游相关胁迫基因的表达,来提高植物对外界环境的适应性[29-31]。在本研究中,RD22、RD29A、DREB2A和LEA,4个已被证明主要参与干旱胁迫的基因[32-35],干旱胁迫后在OE-6和OE-8中的表达量显著高于WT (图6)。由此推测,干旱胁迫下,PheMYB2R-4 极有可能是通过调控这些干旱胁迫信号通路中关键基因的表达, 最终影响毛竹对干旱胁迫的耐受性,而 PheMYB2R-4 基因直接调控的靶基因,是后续研究的重点。
以上研究结果表明PheMYB2R-4在植物的干旱胁迫中发挥着重要的作用,研究结果将进一步创新和丰富MYB 转录因子的研究理论,并为毛竹抗逆新种质的创建提供新的基因资源。
-
本研究从毛竹的基因组中筛选出 1 个 R2R3-MYB 转录因子PheMYB2R-4,通过对 PheMYB2R-4 基因进行分子特性分析和功能验证,结果显示 PheMYB2R-4 基因定位在细胞核中,且在酵母中具有转录活性。在逆境胁迫过程中对干旱胁迫具有正调控作用,在拟南芥中过表达PheMYB2R-4可以提高转基因拟南芥对干旱胁迫的耐受性。在干旱胁迫下,PheMYB2R-4可以提高 AtRD22、AtRD29A、AtDREB2A和AtLEA在转基因拟南芥中的表达量。
毛竹PheMYB2R-4的克隆与功能分析
Cloning and Functional Analysis of PheMYB2R-4 in Phyllostachys edulis
-
摘要:
目的 研究 MYB 转录因子家族在毛竹干旱胁迫反应中的重要作用,为毛竹的抗逆改良和分子育种提供基因资源。 方法 从毛竹的基因组数据库中获得PheMYB2R-4的基因序列,利用亚细胞定位和转录活性实验分析基因的分子特性、通过实时荧光定量 PCR技术、转基因拟南芥表型分析和逆境相关生理生化指标的测定来确定PheMYB2R-4基因的功能。 结果 毛竹中PheMYB2R-4编码 1 个 305个氨基酸的蛋白PheMYB2R-4,其N端区域有一个保守的R2R3区域,属于R2R3-MYB 亚家族。PheMYB2R-4 基因的表达受到干旱和盐的显著诱导。PheMYB2R-4 是一个核定位蛋白,具转录自激活活性。PheMYB2R-4 的过表达提高了干旱胁迫下拟南芥的叶片相对含水量,降低了相对电导率并减少了丙二醛的积累,说明过表达 PheMYB2R-4 通过提高拟南芥的保水能力和减少氧化损伤来增强转基因拟南芥的耐旱性;同时干旱胁迫下, AtRD22、AtRD29A、AtDREB2A和AtLEA基因的表达量均上调。 结论 MYB 转录因子家族中PheMYB2R-4在干旱胁迫应答反应中具有正调控作用,可以提高植物的耐旱性,增强干旱响应基因的表达量。 -
关键词:
- 毛竹
- / PheMYB2R-4转录因子
- / 非生物胁迫
- / 干旱应答
Abstract:Objective The role of MYB transcription factor family in the drought stress response of Phyllostachys edulis was analyzed to provide candidate genes for stress resistance improvement and molecular breeding of moso bamboo. Method In this study, the gene sequence of PheMYB2R-4 was obtained from the genome database of Phyllostachys edulis, and the molecular characteristics of PheMYB2R-4 was analyzed by subcellular localization and transcriptional activity experiments. The function of PheMYB2R-4 was confirmed by qRT-PCR, phenotype analysis of transgenic Arabidopsis thaliana and determination of stress-related physiological and biochemical indicators. Result PheMYB2R-4 encoded a protein with 305 amino acids, which had a conservative R2R3 domain in its N-terminal region and belonged to the R2R3-MYB subfamily. The expression of PheMYB2R-4 gene was significantly induced by drought and salt. PheMYB2R-4 was a nuclear localization protein and had transcriptional self-activation activity. The overexpression of PheMYB2R-4 increased the relative water content of Arabidopsis thaliana leaves under drought stress and decreased the relative conductivity and the accumulation of malondialdehyde, indicating that the overexpression of PheMYB2R-4 enhanced the drought tolerance of transgenic Arabidopsis thaliana by improving its water retention capacity and reducing oxidative damage. At the same time, under drought stress, AtRD22, AtRD29A, AtDREB2A and AtLEA were all up-regulated. Conclusion PheMYB2R-4 has a positive regulatory role in the response to drought stress, which can improve the drought tolerance of plants and enhance the expression of drought response genes. -
图 2 PheMYB2R-4在干旱、盐、高温及低温胁迫下的基因表达
Figure 2. Relative expression of PheMYB2R-4 in response to drought, salt, high temperature and low temperature treatments
图 4 野生型和转基因拟南芥在甘露醇下的萌发分析
Figure 4. Germination Analysis of wild type and transgenic Arabidopsis under Mannitol treatment
图 5 野生型和转基因拟南芥在干旱下的表型分析
Figure 5. Phenotypic Analysis of wild type and transgenic Arabidopsis under drought treatment
图 6 干旱相关基因的在野生型和转基因拟南芥中干旱处理前后的表达分析
Figure 6. Expression analysis of drought-related genes in wild type and transgenic Arabidopsis before and after drought treatment.
表 1 荧光定量引物
Table 1. Primer for qRT-PCR
基因
Gene引物名称
Primer Name引物序列 (5’-3’)
Primer sequence(5’-3’)TIP41 TIP41F AAAATCATTGTAGGCCATTGTCG TIP41R ACTAAATTAAGCCAGCGGGAGTG PheMYB2R-4 PheMYB2R-4F TTGGAAGCAGGTGGTCGATC PheMYB2R-4R AACCTGCAGCACTCGTAGTC Atactin2 Atactin2F AAGCTGGGGTTTTATGAATGG Atactin2R TTGTCACACACAAGTGCATCAT AtRD29A AtRD29AF TGAAAGGAGGAGGAGGAATGGTTGG AtRD29AR ACAAAACACACATAAACATCCAAAGT AtRD22 AtRD22F CAAACCCTTTCGTGTATAACTATGCA AtRD22R TTTCCCGCGAACCAAGTC AtDREB2A
AtDREB2AF GCGATTTTCATCTGGATCACATT AtDREB2AR GCAGGTTCCTCGCATCCTT AtLEA
AtLEAF GGCGGATTTGAGGGACGAAA AtLEAR GTTATGGTGGCCTGGAAGCT  下载: 导出CSV
下载: 导出CSV
-
[1] 杨振亚, 周本智, 周 燕, 等. PEG模拟干旱对毛竹种子萌发及生长生理特性的影响[J]. 林业科学研究, 2018, 31(6):47-54. doi: 10.13275/j.cnki.lykxyj.2018.06.007 [2] WU M, HE W, WANG L N,et al. PheLBD29, an LBD transcription factor from Moso bamboo, causes leaf curvature and enhances tolerance to drought stress in transgenic Arabidopsis[J]. Journal of Plant Physiology, 2023, 280: 153865. doi: 10.1016/j.jplph.2022.153865 [3] WU M, LIU H L, GAO Y M,et al. The moso bamboo drought-induced 19 protein PheDi19-8 functions oppositely to its interacting partner, PheCDPK22, to modulate drought stress tolerance[J]. Plant Science, 2020, 299: 110605. doi: 10.1016/j.plantsci.2020.110605 [4] GAO Y M, LIU H L, ZHANG K M,et al. A moso bamboo transcription factor, Phehdz1, positively regulates the drought stress response of transgenic rice[J]. Plant cell reports, 2021, 40(1): 187-204. doi: 10.1007/s00299-020-02625-w [5] WU M, LIU H L, HAN G M,et al. A moso bamboo WRKY gene PeWRKY83 confers salinity tolerance in transgenic Arabidopsis plants[J]. Scientific Reports, 2017, 7: 11721. doi: 10.1038/s41598-017-10795-z [6] WU M, ZHANG R, XU Z Y,et al. The moso bamboo WRKY transcription factor, PheWRKY86, regulates drought tolerance in transgenic plants[J]. Plant physiology and biochemistry, 2022, 170: 180-191. doi: 10.1016/j.plaphy.2021.10.024 [7] LIU H L, GAO Y M, WU M,et al. TCP10, a TCP transcription factor in moso bamboo (Phyllostachys edulis), confers drought tolerance to transgenic plants[J]. Environmental and Experimental Botany, 2020, 172: 104002. doi: 10.1016/j.envexpbot.2020.104002 [8] XU Y Z, LIU H L, GAO Y M,et al. The TCP transcription factor PeTCP10 modulates salt tolerance in transgenic Arabidopsis[J]. Plant Cell Reports, 2021, 40(10): 1971-1987. doi: 10.1007/s00299-021-02765-7 [9] WU M, LIU R, GAO Y M,et al. PheASR2, a novel stress-responsive transcription factor from moso bamboo (Phyllostachys edulis), enhances drought tolerance in transgenic rice via increased sensitivity to abscisic acid[J]. Plant physiology and biochemistry, 2020, 154: 184-94. doi: 10.1016/j.plaphy.2020.06.014 [10] CHEN F, LIU H L, WANG K,et al. Identification of CCCH zinc finger proteins family in moso bamboo (Phyllostachys edulis), and PeC3H74 confers drought tolerance to transgenic plants[J]. Front. Plant Sci., 2020, 11: 579255. doi: 10.3389/fpls.2020.579255 [11] WU M, LIU H X, WANG L N,et al. Comparative genomic analysis of the CPK gene family in Moso bamboo (Phyllostachys edulis) and the functions of PheCPK1 in drought stress[J]. Protoplasma, 2023, 260(1): 171-187. doi: 10.1007/s00709-022-01765-y [12] ZHANG K M, LAN Y G, WU M,et al. PhePLATZ1, a PLATZ transcription factor in moso bamboo (Phyllostachys edulis), improves drought resistance of transgenic Arabidopsis thaliana[J]. Plant Physiology and Biochemistry, 2022, 186: 121-134. doi: 10.1016/j.plaphy.2022.07.004 [13] HOU D, ZHAO Y, HU Q T,et al. PeSNAC-1 a NAC transcription factor from moso bamboo (Phyllostachys edulis) confers tolerance to salinity and drought stress in transgenic rice[J]. Tree Physiology, 2020, 40(12): 1792-1806. doi: 10.1093/treephys/tpaa099 [14] XU Y Z, WANG L N, LIU H X,et al. Identifcation of TCP family in moso bamboo (Phyllostachys edulis) and salt tolerance analysis of PheTCP9 in transgenic Arabidopsis[J]. Planta, 2022, 256(1): 5. doi: 10.1007/s00425-022-03917-z [15] CAO, Z H, ZHANG S Z, WANG R K,et al. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene confering abiotic stress tolerance in plants[J]. PloS ONE, 2013, 8(7): e69955. doi: 10.1371/journal.pone.0069955 [16] LIPSICK J S. One billion years of Myb[J]. Oncogene, 1996, 13(2): 223-235. [17] OGATA K, MORIKAWA S, NAKAMURA H,et al. Comparison of the free and DNA-complexed forms of the DMA-binding domain from c-Myb[J]. Nature Structural Biology, 1995, 2(4): 309-320. doi: 10.1038/nsb0495-309 [18] 唐 宁, 陈信波. 植物MYB转录因子与非生物胁迫响应研究[J]. 生物学杂志, 2014, 31(3):74-78. [19] SUN P P, ZHU X F, HUANG X F,et al. Overexpression of a stress-responsive MYB transcription factor of Poncirus trifoliata confers enhanced dehydration tolerance and increases polyamine biosynthesis[J]. Plant Physiology and Biochemistry, 2014, 78: 71-79. doi: 10.1016/j.plaphy.2014.02.022 [20] LI K Q, XING C H, YAO Z Z,et al. PbrMYB21, a novel MYB Protein of Pyrus betulaefolia, functions in drought tolerance and modulates polyamine levels by regulating arginine decarboxylase gene[J]. Plant Biotechnol J, 2017, 15(9): 1186-1203. doi: 10.1111/pbi.12708 [21] CUI M H, YOO K S, HYOUNG S,et al. An Arabidopsis R2R3-MYB transcription factor, AtMYB20, negatively regulates type 2C serine/threonine protein phosphatases to enhance salt tolerance[J]. FEBS Letters, 2013, 587(12): 1773-1778. doi: 10.1016/j.febslet.2013.04.028 [22] WANG N, ZHANG W, QIN M,et al. Drought tolerance conferred in soybean (Glycine max. L) by GmMYB84, a novel R2R3-MYB transcription factor[J]. Plant Cell Physiol, 2017, 58(10): 1764-1776. doi: 10.1093/pcp/pcx111 [23] DING Z H, LI S M, AN X L,et al. Transgenic expression of MYB15 confers enhanced sensitivity to abscisic acid and improved drought tolerance in Arabidopsis thaliana[J]. Genet Genomics, 2009, 36(1): 17-29. doi: 10.1016/S1673-8527(09)60003-5 [24] 肖冬长, 张智俊, 徐英武, 等. 毛竹 MYB 转录因子 PeMYB2 的克隆与功能分析[J]. 遗传, 2013, 35(10):1217-1225. [25] EL-KEREAMY A, BI Y M, RANATHUNGE K,et al. The rice R2R3-MYB transcription factor OsMYB55 is involved in the tolerance to high temperature and modulates amino acid metabolism[J]. PLoS ONE, 2012, 7(12): e52030. doi: 10.1371/journal.pone.0052030 [26] AHAMMED G J, LI X, WAN H, et al. SlWRKY81 reduces drought tolerance by attenuating proline biosynthesis in tomato[J]. Scientia Horticulturae, 2020, 270. [27] QUAN W, HU Y, MU Z,et al. Overexpression of AtPYL5 under the control of guard cell specific promoter improves drought stress tolerance in Arabidopsis[J]. Plant Physiology and Biochemistry, 2018, 129: 150-157. doi: 10.1016/j.plaphy.2018.05.033 [28] CUI J, JIANG N, ZHOU X,et al. Tomato MYB49 enhances resistance to Phytophthora infestans and tolerance to water deficit and salt stress[J]. Planta, 2018, 248(6): 1487-1503. doi: 10.1007/s00425-018-2987-6 [29] KRISHNA R, KARKUTE S G, ANSARI W A,et al. Transgenic tomatoes for abiotic stress tolerance: status and way ahead[J]. 3 Biotech, 2019, 9(4): 143. doi: 10.1007/s13205-019-1665-0 [30] PENG Y, ZHANG J, CAO G,et al. Overexpression of a PLDα1 gene from Setaria italica enhances the sensitivity of Arabidopsis to abscisic acid and improves its drought tolerance[J]. Plant Cell Reports, 2010, 29(7): 793-802. doi: 10.1007/s00299-010-0865-1 [31] XU G Y, ROCHA P S, WANG M L,et al. A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis[J]. Planta, 2011, 234(1): 47-59. doi: 10.1007/s00425-011-1386-z [32] THAMMEGOWDA V, HARSHAVARDHAN V S, SEILER C,et al. AtRD22 and AtUSPL1, members of the plant-specific BURP domain family involved in Arabidopsis thaliana drought tolerance[J]. PloS ONE, 2014, 9(10): e110065. doi: 10.1371/journal.pone.0110065 [33] HUA Z M, YANG X, FROMM M E. Activation of the NaCl- and drought-induced RD29A and RD29B promoters by constitutively active Arabidopsis MAPKK or MAPK proteins[J]. Plant Cell and Environment, 2006, 29(9): 1761-1770. doi: 10.1111/j.1365-3040.2006.01552.x [34] SAKUMA Y, MARUYAMA K, OSAKABE Y,et al. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression[J]. Plant Cell, 2006, 18(5): 1292-1309. doi: 10.1105/tpc.105.035881 [35] CANDAT A, PASZKIEWICZ G, NEVEU M,et al. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress[J]. Plant Cell, 2014, 26(7): 3148-3166. doi: 10.1105/tpc.114.127316 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 2785
- HTML全文浏览量: 1400
- PDF下载量: 82
- 被引次数: 0
