



-
转录因子(transcription factor,TF)又称反式作用因子,可识别并特异性结合靶基因启动子区的相关顺式作用元件,进而激活或抑制基因表达[1-3]。在植物的生长发育、形态建成及适应环境变化过程中转录因子起重要调控作用。碱性亮氨酸拉链(basic region/leucine zipper,bZIP)转录因子家族是真核生物中分布最广泛、成员种类最多的家族之一。bZIP主要由位于N-端的碱性结构域和C-端的亮氨酸拉链区域组成。碱性结构域由20个氨基酸残基组成,包含核定位信号和一段固定的N-X7-R/K基序,能够特异识别并结合启动子上的顺式元件[4-5]。亮氨酸拉链区则由1个或多个重复单位组成,每个重复单位含有7或9个氨基酸,最末位一般是亮氨酸,该区域可以形成两亲性的α螺旋结构,使bZIP蛋白单体形成同源或异源二聚体,从而为bZIP蛋白功能多样性提供基础[6]。在拟南芥(Arabidopsis thaliana (L.) Heynh.)中共鉴定出78个bZIP成员,分为13个亚家族[7]。不同亚家族成员之间的功能存在差异,A亚族成员主要参与ABA信号转导、种子发育和叶片衰老等过程,B和K亚族成员在内质网应激反应中发挥功能,C亚族成员参与种子萌发和环境胁迫过程,D亚族成员在病原体防御方面起重要作用,E和M亚族成员负责花粉发育,F亚族成员是锌缺乏反应的中心调节因子,G、H和J亚家族成员参与光信号转导,I亚家族成员主要调控愈伤组织形成和维管发育,S亚家族成员主要在生长发育和环境适应性中调控碳氮代谢与能量稳态[8-14]。S亚家族包含17个成员,是bZIP家族中最大的一个亚族,但其基因功能报道较少,目前仅对S1组的5个成员(AtbZIP1、AtbZIP2、AtbZIP11/ATB2、AtbZIP44和AtbZIP53)进行了研究。S1类bZIP成员具有独特的无内含子基因结构,编码小而高度同源的蛋白[15]。研究表明,S1-bZIPs能与C类bZIP蛋白形成异源二聚体,在蔗糖非发酵-1-相关蛋白激酶(SUCROSE NON-FERMENTING1 RELATED KINASES1, SnRK1)介导的细胞内低能量以及低糖水平感应信号途径中起重要调控作用[16-17]。在低能量条件下,拟南芥AtbZIP2、AtbZIP11和AtbZIP44直接激活生长素信号负调控因子INDOLE-3-ACETIC ACID INDUCIBLE 3(IAA3)的转录进而抑制生长素输出载体PIN-FORMED(PIN)基因的表达,导致根尖生长素极性运输减弱,从而抑制根的生长[18]。在种子发育早期,AtbZIP44高水平表达[19],并与β-甘露聚糖酶7(Endo-β-mannanases 7,MAN7)启动子区的G-box结合,正调控种子萌发[20]。此外,S1-bZIPs也参与调控植物对非生物胁迫的响应。干旱和盐胁迫下,AtbZIP1表达量显著上调,增强Responsive to dehydration 17(RD17)、RD29A和9-cis-epoxycarotenoid dioxygenase 3(NCED3)等胁迫响应基因的表达,进而提高植株的耐受性[21]。AtbZIP53受盐胁迫诱导,激活下游脯氨酸脱氨酶(PROLINE DEHYDOGENSE1,ProDH)基因的表达,调节植物体内氨基酸代谢重编程以适应盐胁迫[22]。同时C亚族成员AtbZIP10可与AtbZIP53形成异源二聚体,进一步增强其对ProDH启动子的结合活性[23]。
结合拟南芥中S1-bZIPs基因的研究发现,S1-bZIPs广泛参与调控植物体内能量稳态适应非生物胁迫。然而,目前对木本植物S1-bZIPs成员的系统性鉴定和响应胁迫的表达模式研究还比较缺乏。杨树是重要的造林树种和工业用材树种[24]。随着毛果杨(Populus trichocarpa (Torr. & Gray))[25]、新疆杨(Populus alba var. pyramidalis Bunge)[26]、84K杨(Populus alba × P. glandulosa Hyun)[27]和毛白杨(Populus tomentosa Carrière)[28]等杨树全基因组测序的完成以及多种杨树的遗传转化体系日益完善,杨树已成为林木基因工程的模式物种。本研究以毛白杨为对象,利用生物信息学技术从全基因组水平鉴定S1类bZIP家族成员,分析PtoS1-bZIPs的基因结构、系统进化等特征,明确PtoS1-bZIPs基因的组织表达以及对非生物胁迫的响应模式,为后续解析PtoS1-bZIPs基因的生物学功能及其调控植物抗逆性的分子机制奠定基础。
-
从拟南芥基因组网站(
https://www.arabidopsis.org/ )中下载AtS1-bZIP亚家族基因编码的氨基酸序列,通过BLASTP和hmmsearch程序对毛白杨基因组数据库进行搜索,鉴定筛选出10个毛白杨PtoS1-bZIP候选基因。利用NCBI毛白杨基因组数据库(https://www.ncbi.nlm.nih.gov/datasets/genome/GCA_018804465.1/ )查找候选基因的染色体位置信息。通过Expasy ProtParam tool网站(https://www.expasy.org/ )获得PtoS1-bZIP亚家族成员的基本理化信息。利用在线网站(https://wolfpsort.hgc.jp/ )预测PtoS1-bZIPs蛋白的亚细胞定位。 -
使用Clustal软件将毛白杨(Populus tomentosa)、毛果杨(P. trichocarpa)、拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的S1-bZIPs氨基酸序列进行多重序列比对。在MEGA7软件中,通过邻接法将比对的序列构建进化树,在Evoview(
https://www.evolgenius.info/ )网站中进行美化。 -
通过GSDS网站(
http://gsds.cbi.pku.edu.cn/ )分析杨树PtoS1-bZIP亚家族成员的基因结构。利用MEME网站(http://gsds.cbi.pku.edu.cn/ )预测PtoS1-bZIPs的蛋白保守基序。 -
基于毛白杨基因注释文件,使用Tbtools软件绘制PtoS1-bZIPs的染色体定位图。利用MCScanX对毛白杨种内及毛白杨与拟南芥、水稻种间的S1-bZIPs基因进行共线性分析,并通过TBtools对分析结果进行可视化展示。
-
从毛白杨基因组中提取PtoS1-bZIPs基因上游2000 bp序列作为启动子,通过PlantCARE网站(
http://bioinformatics.psb.ugent.be/webtools/plantcare/htm )预测顺式作用元件,并利用Excel筛选和统计各基因的响应类元件。 -
毛白杨无性系材料来源于中国林业科学研究院林木遗传育种国家重点实验室。在含有WPM培养基的组培瓶里生长1个月后,移栽至温室中(昼夜温度: 25 °C/18 °C, 相对湿度: 50%~60%,光照时间: 16 h)培养3个月,选择长势一致的植株,收集每个植株的叶、皮、形成层、木质部和根组织,每次取材进行三次生物学重复。通过CTAB法提取各个组织的RNA,利用PrimeScript™ RT reagent Kit(Perfect Real Time)(Takara, Dalian, China)试剂盒合成cDNA,根据PtoS1-bZIPs基因序列设计特异引物(表1),进行实时荧光定量qRT-PCR分析。每个组织使用三次生物学重复以及四次技术重复。以PtoUBQ10为内参基因,利用2−∆CT法计算相对表达量。
表 1 PtoS1-bZIPs基因qRT-PCR引物
Table 1. Primers for quantitative real-time PCR of PtoS1-bZIP genes
基因名称
Gene names上游引物(5′→3′)
Forward primers (5′→3′)下游引物(5′→3′)
Reverse primers (5′→3′)PtobZIP55 AGGCCGATAACTCAATCTTG CATTAAAGATGCTTGAATCTCC PtobZIP21 GCGTTACTTGAGTGTTGAGG ATTGAAGGTGATCGAGTCTC PtobZIP27 GAGATCCTGACAAGGATGA TAGTCGCAGAAACAGAGGC PtobZIP44 CGAAGCTGAGAACTCTATCC CAAAAGCACCAGTAGTGGG PtobZIP6 GTCACAACACAGCACTTCTT GCACTAGTTTGGAGATCCTC PtobZIP33 CCATCAATGTCACAACACAGC ATCAGGGAGATCCTCGTGATG PtobZIP63 TCAGGCTCTGAAATTGACCCA AACTTTGTTGTCTGATTCGAGAG PtobZIP49 ATCAGGTTCTGACGTTGATGCG TGTTCTGTGATTCGAGAGC PtobZIP15 GGTCGTTGAACTCTGTGTTGC TTGAAACATATCAGCAGATGCC PtobZIP75 TGTGGAATTGACTGAGAGGC AATAGGCTGTACTGAACACG PtoUBQ10 GTTGATTTTTGCTGGGAAGC GATCTTGGCCTTCACGTTGT -
材料为生长1个月的毛白杨组培苗,选择长势一致的植株平均分为四组,转移至1/4 Hoagland营养液中在人工气候室(昼夜温度: 25 °C/18 °C, 相对湿度: 50%~60%,光照时间: 16 h)进行水培,处理组分别进行100 μmol·L−1 ABA和100 mmol·L−1 NaCl处理,以及用10% PEG6000处理来模拟干旱胁迫,以未处理的毛白杨为对照组,每组设置三个生物学重复。对处理24 h后植株顶端向下第3、4片功能叶进行取样,通过CTAB法提取RNA,反转录为cDNA进行qRT-PCR分析,引物序列见表1。每个处理使用三次生物学重复以及四次技术重复。以PtoUBQ10为内参基因,利用2−∆∆CT法进行相对表达量分析。
-
通过生物信息学方法,鉴定出毛白杨bZIP基因家族包含86个成员,分为12个亚家族。按照基因在染色体上的位置顺序,将其依次命名为PtobZIP1~PtobZIP86。利用拟南芥AtS1-bZIPs基因在毛白杨基因组数据库中进行比对,筛选到10个PtoS1-bZIPs基因。PtoS1-bZIP亚家族成员基因的基本信息分析表明,各基因所编码的氨基酸序列长度在139~165 aa之间,分子量分布在15.78~18.71 kDa之间,等电点介于6.05~9.66之间。通过对PtoS1-bZIP亚家族成员进行亚细胞定位预测发现,所有蛋白均定位在细胞核中(表2)。
表 2 PtoS1-bZIP 亚家族基因特征分析
Table 2. Characteristics analysis of PtoS1-bZIP subfamily genes in P. × tomentosa
基因名称
Gene names基因号
Gene ID染色体
Chr内含子
Introns外显子
Exons氨基酸数目
AA分子量
MW (kDa)等电点
PI亚细胞定位
Subcellular locationPtobZIP55 KAG6762870.1 9 0 1 156 17.30 8.09 Nucleus PtobZIP21 KAG6777875.1 4 0 1 156 17.44 8.10 Nucleus PtobZIP27 KAG6773901.1 5 0 1 163 18.25 6.59 Nucleus PtobZIP44 KAG6766766.1 7 0 1 165 18.71 6.20 Nucleus PtobZIP6 KAG6785745.1 2 0 1 157 17.90 6.18 Nucleus PtobZIP33 KAG6774655.1 5 0 1 159 18.32 6.05 Nucleus PtobZIP63 KAG6757882.1 10 0 1 144 16.51 9.66 Nucleus PtobZIP49 KAG6764077.1 8 0 1 144 16.46 9.36 Nucleus PtobZIP15 KAG6787046.1 2 0 1 145 16.50 8.88 Nucleus PtobZIP75 KAG6749792.1 14 0 1 139 15.78 7.84 Nucleus -
为分析PtoS1-bZIPs基因的进化关系,将毛白杨PtoS1-bZIPs基因与毛果杨(Populus trichocarpa)、拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)中的S1-bZIPs基因进行序列比对,构建系统进化树。结果显示,上述S1-bZIPs基因可分为3个进化分支,其中第一分支包含的PtoS1-bZIPs基因数量最多,包括PtobZIP55、PtobZIP21、PtobZIP27、PtobZIP44、PtobZIP6和PtobZIP33,与拟南芥AtbZIP2、AtbZIP11和AtbZIP44亲缘关系较近;第二分支包括PtobZIP15和PtobZIP75,与AtbZIP53同源性较高;第三分支包括PtobZIP63和PtobZIP49,与AtbZIP1亲缘关系密切(图1)。

图 1 毛白杨 (Pto)、毛果杨 (Ptr)、拟南芥 (At)和水稻 (Os) S1-bZIP 亚家族成员的系统发育分析
Figure 1. Phylogenetic analysis of S1-bZIP subfamilies in P. tomentosa (Pto), P. trichocarpa (Ptr), A. thaliana (At) and O. sativa (Os)
-
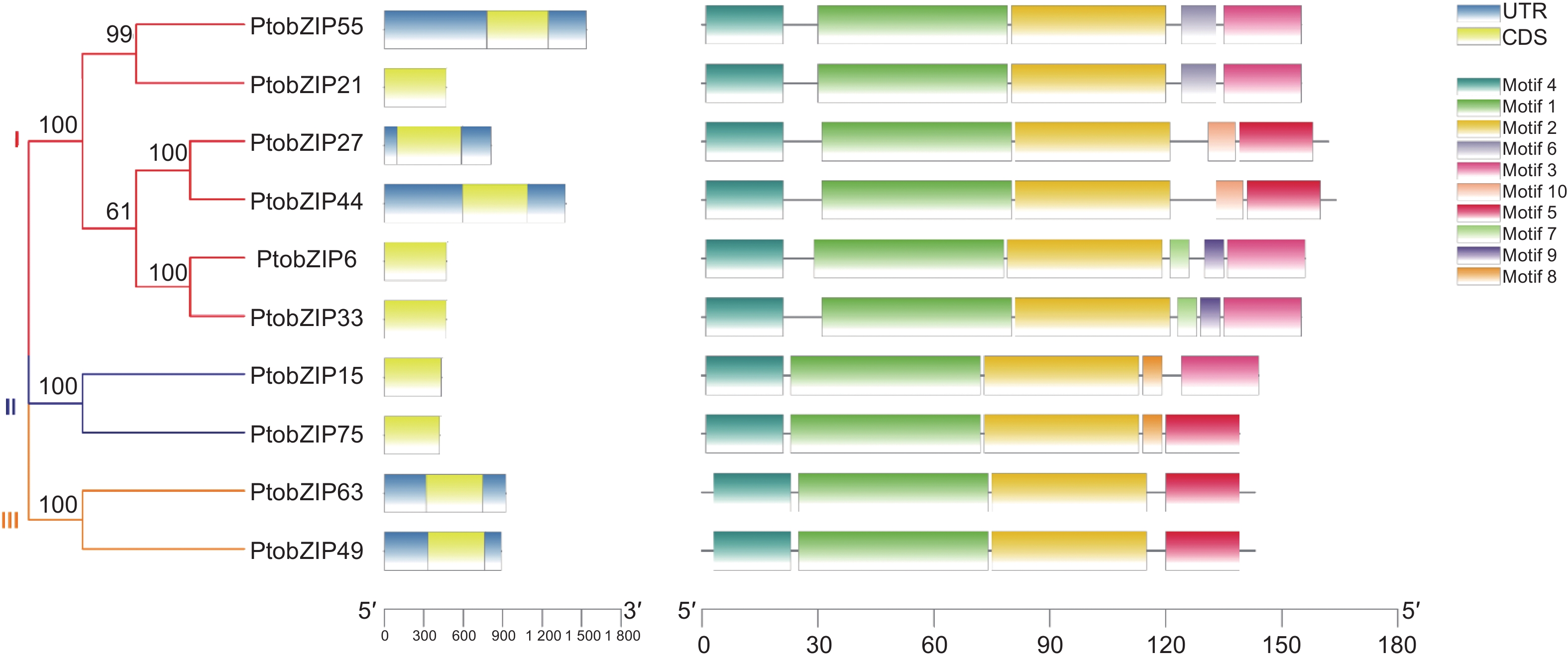
为进一步分析PtoS1-bZIPs基因的序列特征,对其进行基因结构及保守基序分析,结果如图2所示,所有PtoS1-bZIPs成员均无内含子结构。利用在线软件MEME预测PtoS1-bZIPs的蛋白保守基序,共鉴定得到10个保守基序。分析发现,所有PtoS1-bZIP亚家族成员的N-端中均含有motif 1、2、4这3种保守基序,且保守基序分布一致,长度也较为接近。而各成员在C-端的保守基序存在差异,在第一个进化分支中,PtobZIP55和PtobZIP21蛋白的C-端含有motif 3和motif 6,PtobZIP27和PtobZIP44含有motif 5和motif 10,PtobZIP6和PtobZIP33含有motif 3、motif 7和motif 9。在第二个分支中,PtobZIP15蛋白的C-端含有motif 3和motif 8,PtobZIP75则含有motif 5和motif 8。在第三个分支中,PtobZIP63和PtobZIP49蛋白的C-端只含有一个蛋白基序motif 5。同一进化分支内PtoS1-bZIPs含有相似的保守基序,表明其可能具有相似的功能。不同分支中各蛋白C-端保守基序的差异暗示其基因功能可能出现分化。
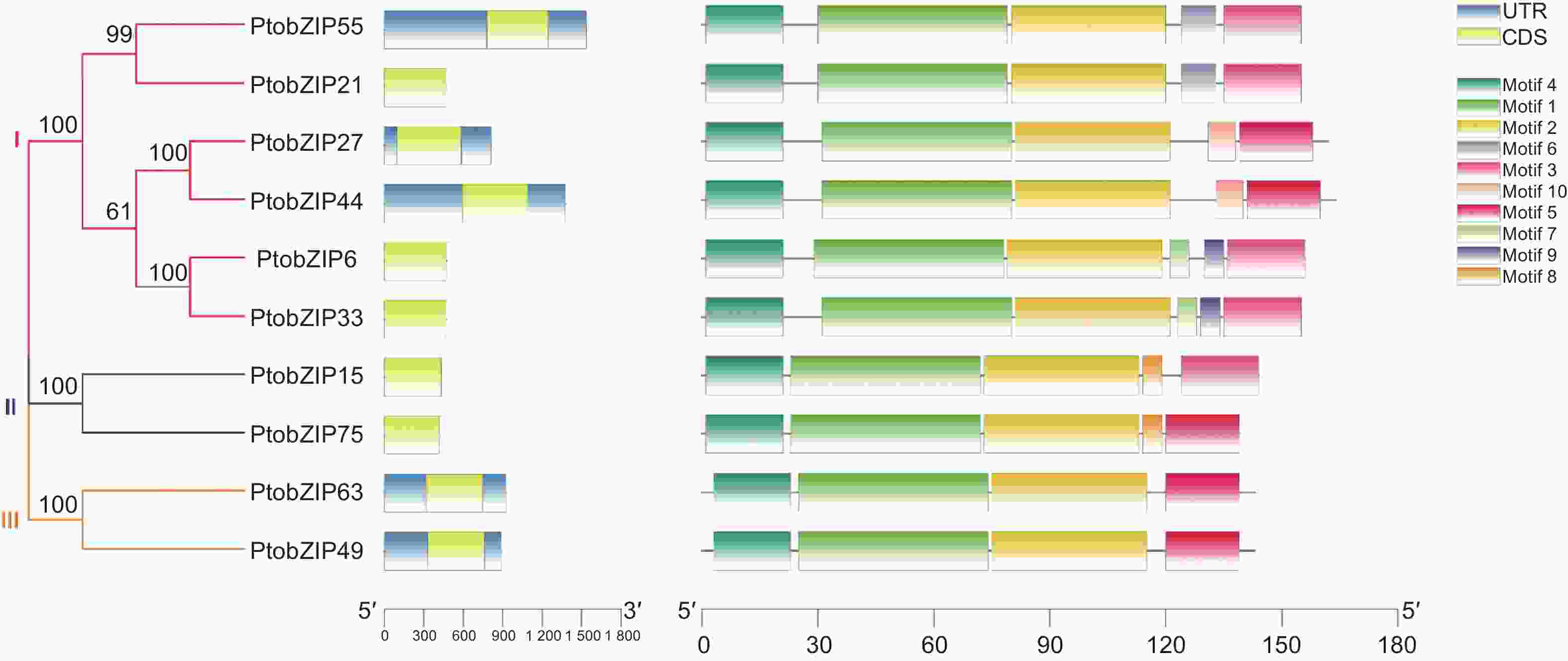
图 2 PtoS1-bZIPs 基因结构与保守基序分析
Figure 2. The gene structures and protein conserved motifs of PtoS1-bZIP genes
-
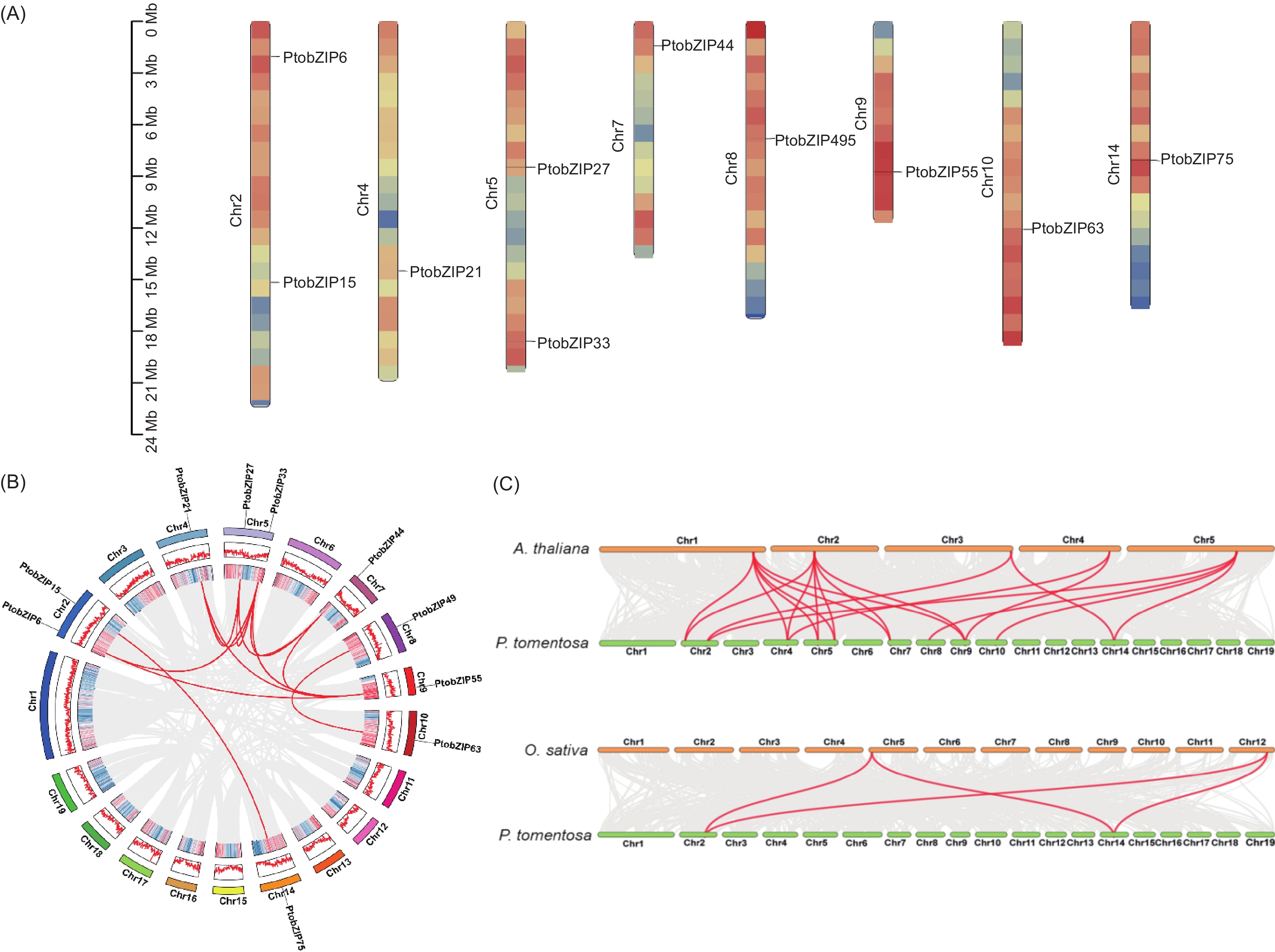
染色体定位结果表明,PtoS1-bZIP亚家族中的10个成员差异分布于毛白杨19条染色体中的8条,分别位于2、4、5、7、8、9、10、14号染色体上,其中2和5号染色体各有2个基因分布,其余6条染色体上各包含1个基因(图3A)。为了分析PtoS1-bZIPs在毛白杨基因组中的复制事件,对其进行种内共线性分析。结果显示,在10个PtoS1-bZIPs基因中共存在12对共线性关系(图3B)。第二个进化分支内的PtobZIP15和PtobZIP75以及第三个进化分支内的PtobZIP63和PtobZIP49分别互为1对共线性基因,而第一个分支内的6个PtobZIPs基因之间出现一个基因同时和多个基因存在共线性关系的情况,说明这些基因在进化过程中发生了扩张事件。为进一步探究S1-bZIPs在不同植物间的进化关系,对毛白杨,拟南芥和水稻的S1-bZIPs基因进行种间共线性分析。结果显示,毛白杨与水稻之间仅有4组基因存在共线性关系,而毛白杨与拟南芥之间具有20组基因存在共线性关系(图3C)。其中大多数PtoS1-bZIPs基因与拟南芥之间至少存在2个共线性基因对,表明毛白杨PtoS1-bZIPs基因与拟南芥之间的亲缘关系更近,推测这些基因可能从共同祖先的基因组中进化而来。

图 3 PtoS1-bZIPs基因在毛白杨染色体上的定位(A)以及在物种内(B)和物种间(C)的共线性分析
Figure 3. Chromosome localization (A), gene duplication (B) and synteny analysis (C) of PtoS1-bZIP genes
-
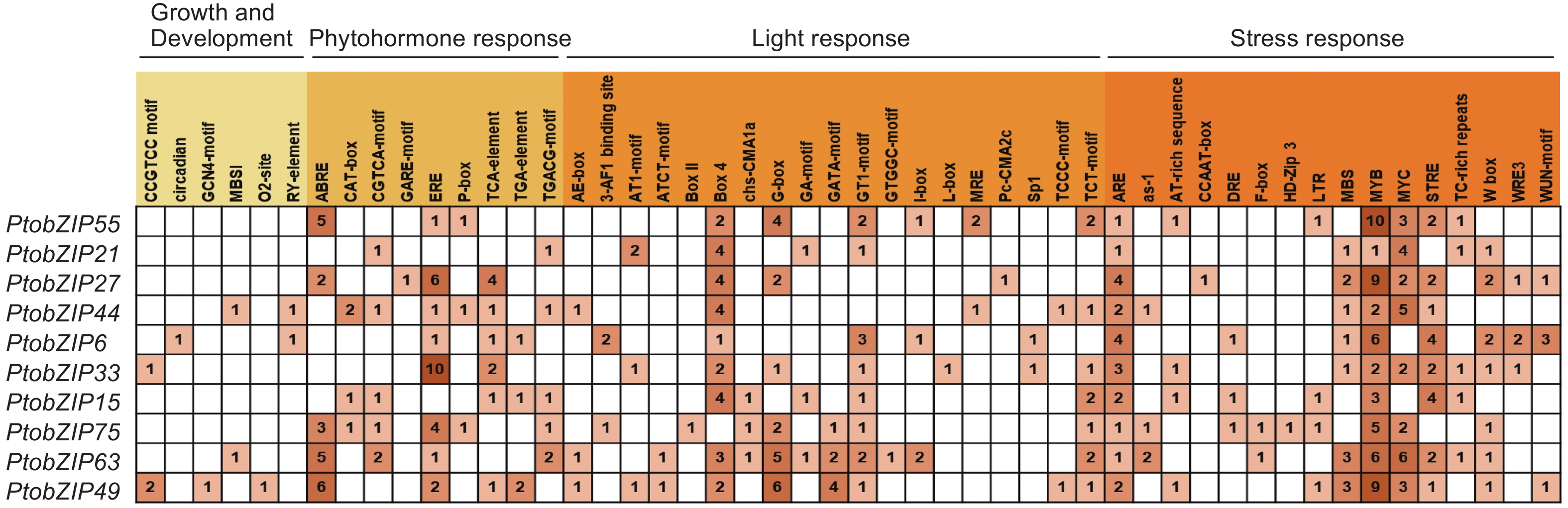
为了预测PtoS1-bZIPs基因的生物学功能,对PtoS1-bZIPs基因上游2 000 bp启动子区域的顺式作用元件进行分析。结果表明,发挥功能的顺式作用元件主要分为4类:植物生长发育类响应元件,激素响应类元件,光响应元件以及生物与非生物胁迫类响应元件(图4)。其中,生长发育类响应元件分布最少,只在5个基因启动子区域存在,其他3类顺式作用元件不均匀地分布在所有PtoS1-bZIPs基因的上游区域。脱落酸响应元件abscisic acid-response element(ABRE)和乙烯响应元件ethylene response element(ERE)是含量较多的激素类响应元件,Box 4和G-box元件在光响应元件中所占比例较高,MYB和MYC作为响应生物与非生物胁迫的重要功能元件,是PtoS1-bZIPs成员中包含最多的顺式作用元件。此外,还有干旱响应元件MYB binding site involved in drought-inducibility(MBS)、厌氧诱导响应元件antioxidant response element(ARE)等广泛存在于PtoS1-bZIPs启动子区域。以上结果表明,PtoS1-bZIPs基因可能在光响应、激素信号转导及环境因子胁迫中发挥重要作用。
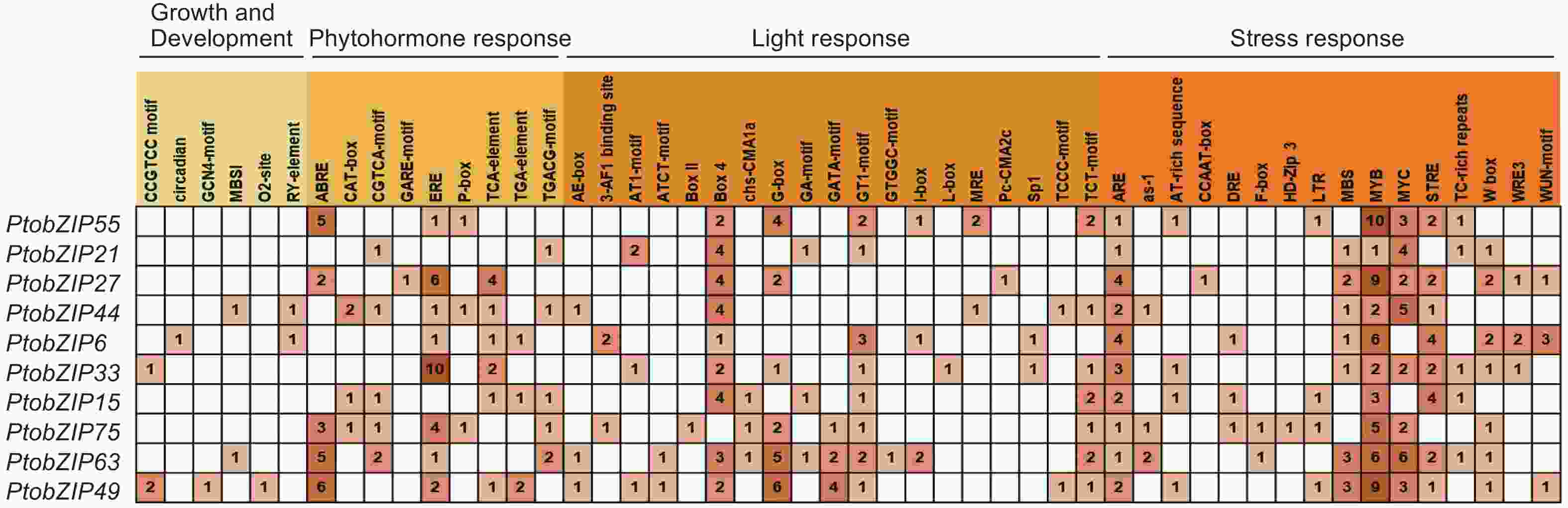
图 4 毛白杨 PtoS1-bZIPs 基因顺式作用元件
Figure 4. Cis-elements of S1-bZIP genes in the genome of P. tomentosa
-
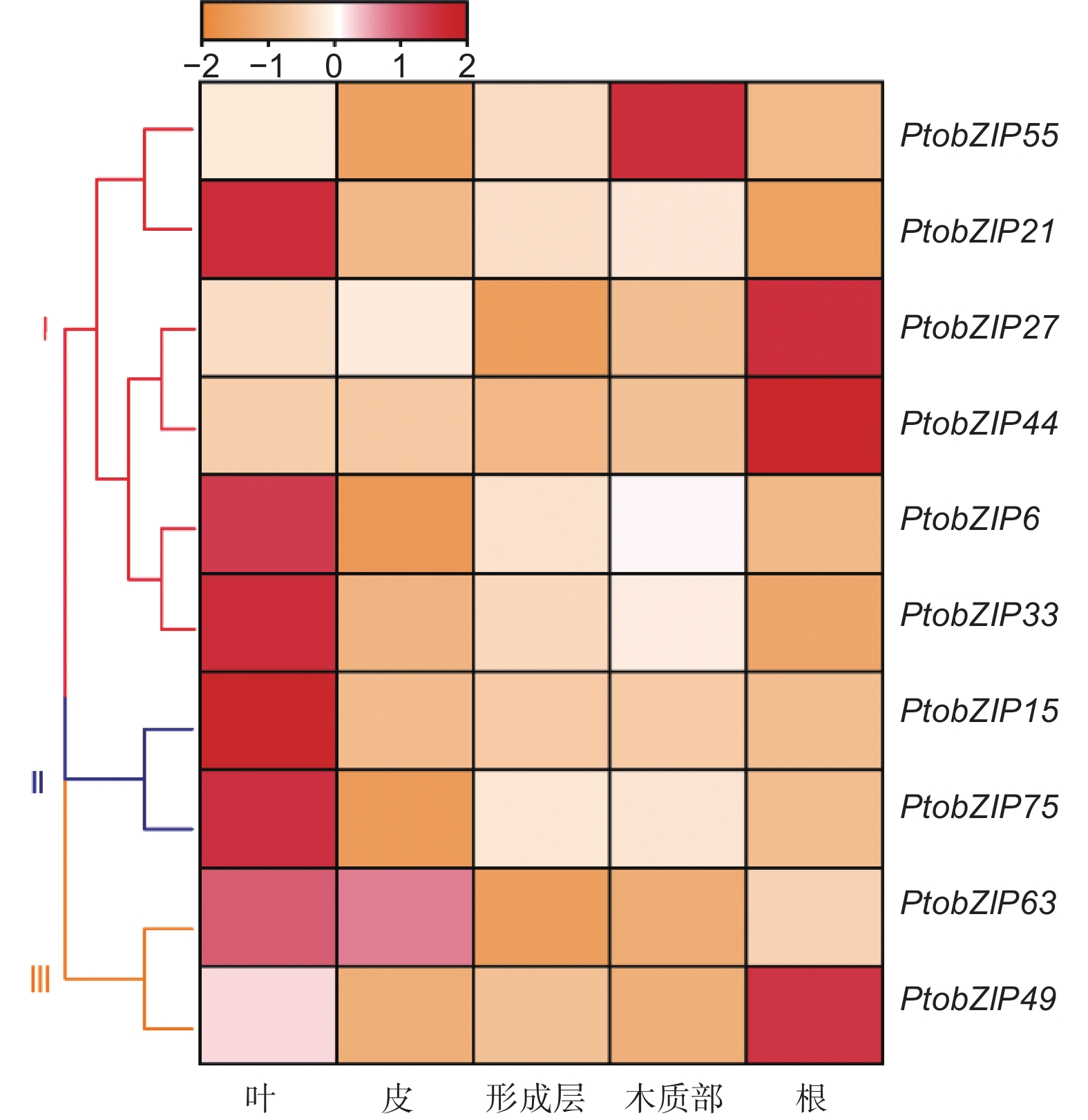
基因在植物不同组织中的表达特异性与其基因功能相关。为探究毛白杨PtoS1-bZIPs的功能,利用qRT-PCR对毛白杨各个组织中的S1-bZIPs表达量进行研究(图5)。结果表明,在第一个进化分支中,PtobZIP55在木质部中有较高的表达丰度,PtobZIP21、PtobZIP6和PtobZIP33在叶中表达量较高,而PtobZIP27和PtobZIP44在根中的表达水平较高。在第二个进化分支中,PtobZIP15和PtobZIP75主要在叶中表现出较高的表达丰度。虽然PtobZIP63和PtobZIP49同属第三个进化分支,但其表达模式有显著差异,PtobZIP63在叶和皮中有较高的表达,而PtobZIP49则在根中的表达量最高。PtoS1-bZIPs基因在不同组织中的差异表达表明该亚家族成员可能存在功能上的差异。

图 5 PtoS1-bZIPs 基因在不同组织中的表达模式
Figure 5. The expression patterns of PtoS1-bZIP genes in different tissues
-
为进一步探究PtoS1-bZIPs基因在激素及非生物胁迫处理下的表达特征,分别对毛白杨进行脱落酸(ABA)、盐(NaCl)和干旱(PEG6000)处理,利用qRT-PCR分析PtoS1-bZIPs基因在不同处理下的表达变化(图6)。结果显示,经脱落酸处理后,第一进化分支的PtobZIP55、PtobZIP21、PtobZIP27和PtobZIP44以及第三进化分支的PtobZIP63和PtobZIP49表达量上调,其中PtobZIP63上调最明显,为对照组的4.1倍。盐胁迫下,第一进化分支中的PtobZIP21、PtobZIP27、PtobZIP44、PtobZIP6和PtobZIP33以及第二进化分支中的PtobZIP15下调表达,而第三进化分支的PtobZIP63和PtobZIP49上调表达,其中PtobZIP27的表达量下调最显著,为对照组的0.2倍,PtobZIP63的表达量上调最明显,为对照组的4.9倍。干旱胁迫下,第一进化分支的PtobZIP55、PtobZIP21和PtobZIP27,第二进化分支的PtobZIP15以及第三进化分支的PtobZIP63和PtobZIP49表达量上调,其中,PtobZIP55的上调表达倍数最高,为对照组的6倍。以上结果表明,PtoS1-bZIPs家族成员在脱落酸、盐和干旱处理下呈现不同的表达特征,推测其在响应非生物胁迫中发挥不同作用。
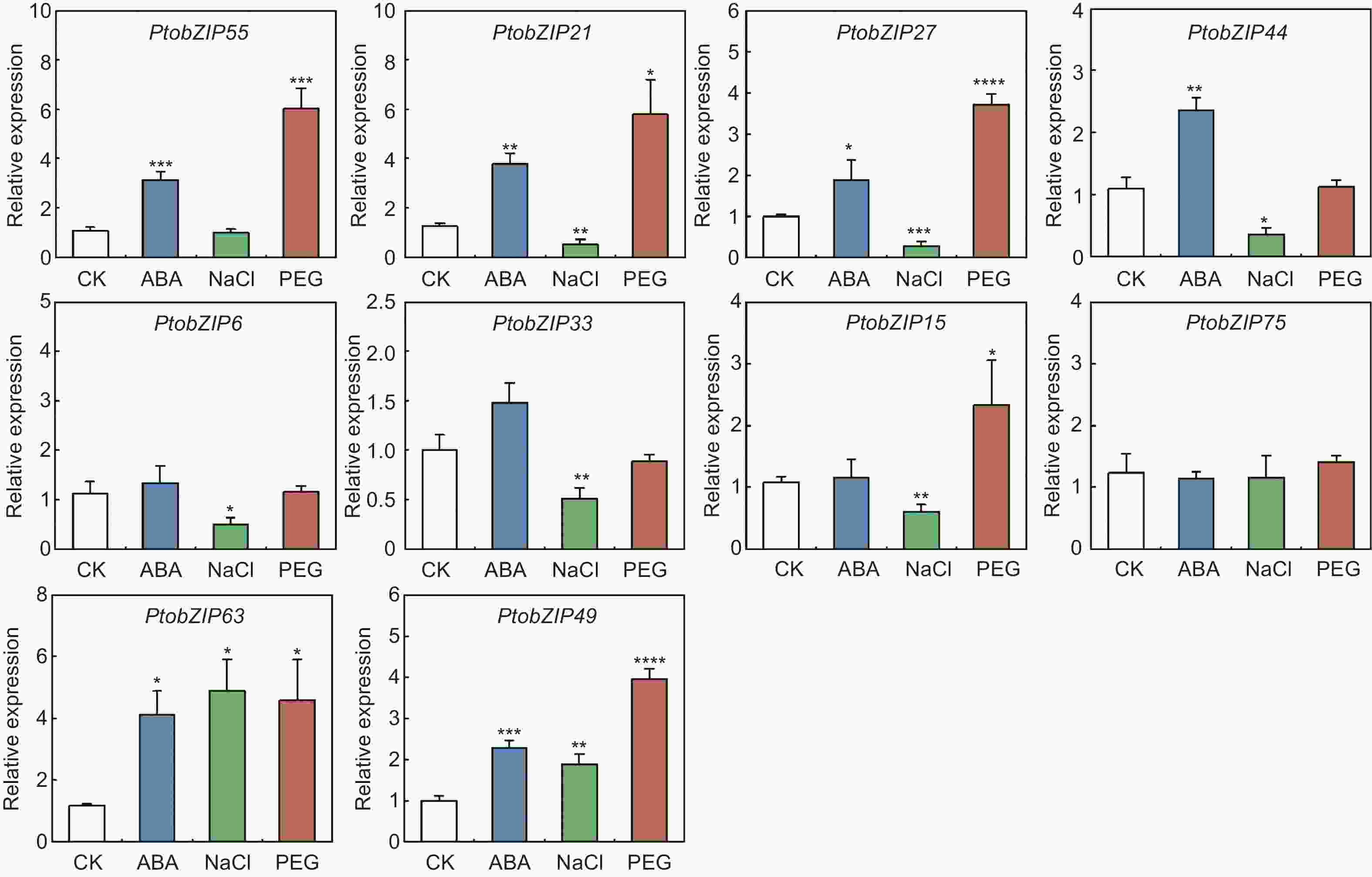
图 6 脱落酸(ABA)、盐(NaCl)及干旱(PEG6000)胁迫下 PtoS1-bZIPs 基因在叶片中的表达水平
Figure 6. The expression levels of PtoS1-bZIP genes in leaves under ABA, NaCl and PEG6000 treatment
-
近年来,对于S1-bZIP亚家族成员的研究主要集中在模式植物拟南芥中,而对木本植物S1-bZIPs成员的系统性鉴定及响应胁迫的表达模式研究较少。本研究以毛白杨为对象,从全基因组水平对PtoS1-bZIP亚家族进行了系统的生物信息学分析,共鉴定得到10个S1-bZIPs基因。进化分析显示,PtoS1-bZIPs蛋白可分为3个进化支,每个进化支中均包含单子叶植物水稻、双子叶植物拟南芥和毛果杨的S1-bZIPs氨基酸序列,说明该基因家族在单、双子叶植物分化之前就已形成。保守基序分析显示,PtoS1-bZIP亚家族各成员在序列的N-端具有相同的保守基序,然而,不同进化分支内的bZIP蛋白在C-端所含有的保守基序存在较大差异,这可能会导致不同分支中S1-bZIPs基因功能出现差异。基因复制分为串联复制和片段复制两种形式,有助于基因家族扩张促进新基因产生和功能分化[29-30]。在PtoS1-bZIP亚家族成员中共检测到12个片段复制基因对,未发现串联重复,说明片段复制事件对于PtoS1-bZIPs成员的进化和扩增具有重要作用。另外,种间共线性分析发现毛白杨与拟南芥之间的共线性关系对最多,表明毛白杨与拟南芥的S1-bZIPs基因可能是由相同祖先进化而来。PtobZIP55和PtobZIP21在毛白杨基因组内以及与拟南芥之间存在的共线性关系对最多,暗示这两个基因在进化过程中发挥关键作用。
基因在不同组织中的表达特异性与其功能密切相关,10个PtoS1-bZIPs基因在各个组织中的表达水平存在明显差异,暗示其基因功能可能不同。启动子区的顺式作用元件在基因表达过程中起重要调控作用,PtoS1-bZIP家族成员含有多个与光、激素及非生物胁迫响应相关元件,其中,MYB和MYC顺式作用元件所占比例最高,而MYB和MYC元件能够应答ABA介导的干旱、盐等非生物胁迫逆境因子[31-32],表明PtoS1-bZIPs可能在逆境响应中发挥重要作用。本研究对毛白杨进行ABA、盐和干旱处理,分析PtoS1-bZIPs的表达特征。第一和第二进化分支中的大部分基因在ABA和干旱处理下上调表达,在盐处理下显著下调,第三个进化分支中的基因在胁迫和ABA处理下均显著上调。结合基因结构分析发现,所有PtoS1-bZIPs成员都没有内含子结构,而缺失内含子的基因能有效减少转录后加工过程,可帮助植物在非生物胁迫下快速转录及翻译[33],推测PtoS1-bZIPs可能在调控植物抗逆性方面发挥重要功能。在盐和干旱胁迫处理下,第一和第二分支有少部分S1-bZIPs成员表达无显著变化,推测这些基因可能为毛白杨在进化过程中产生的冗余S1-bZIPs基因。S1-bZIPs是调控能量代谢的关键因子,在持续黑暗条件下,AtS1-bZIPs通过激活参与支链氨基酸、脯氨酸以及酪氨酸降解途径中相关酶基因的表达来维持细胞内能量稳态[16, 34-35]。拟南芥中研究表明,AtS1-bZIPs可与C亚族成员形成异源二聚体,调控碳水化合物和氨基酸代谢重编程以适应盐胁迫[22]。在番茄中,SlbZIP1通过上调与ABA生物合成以及信号转导相关基因的表达来提高耐盐性和抗旱性[36]。本研究发现,PtoS1-bZIP亚家族第三分支的PtobZIP63和PtobZIP49与SlbZIP1同源性较高,并且在盐和干旱处理下表达量均显著上调,推测PtobZIP63和PtobZIP49可能在盐和干旱胁迫下发挥重要功能。PtoS1-bZIP亚家族第一分支和第二分支中的大部分基因受干旱诱导,受盐胁迫抑制,说明毛白杨PtoS1-bZIPs不同分支中的成员之间可能具有功能分化。PtoS1-bZIP亚家族各分支中关键成员在调控杨树抗逆性中的分子功能还需进一步研究。
-
本研究对毛白杨PtoS1-bZIP亚家族进行了系统的生物信息学分析,共鉴定获得10个S1-bZIPs基因,分布在8条染色体上,均无内含子结构。系统进化分析发现PtoS1-bZIPs可分为3个分支,不同成员的基因表达具有组织特异性。第一和第二分支中多数PtoS1-bZIPs成员的表达受干旱胁迫诱导,而受盐胁迫抑制,第三分支成员在干旱和盐胁迫处理下均上调表达,说明PtoS1-bZIP亚家族不同分支中的成员具有功能分化。本研究结果可为后续PtoS1-bZIPs的生物学功能及其调控植物抗逆性的分子机制研究提供基础。
毛白杨PtoS1-bZIP亚家族成员的鉴定及表达分析
Identification and Expression Analysis of PtoS1-bZIP Subfamily in Populus tomentosa
-
摘要:
目的 鉴定毛白杨PtoS1-bZIP亚家族成员,解析PtoS1-bZIPs基因响应干旱和盐胁迫等非生物逆境的表达模式。 方法 通过生物信息学方法对PtoS1-bZIP亚家族进行系统分析,利用实时荧光定量技术检测该亚家族成员在不同组织中的基因表达特征以及对非生物胁迫的表达响应模式。 结果 从毛白杨基因组中鉴定出10个S1-bZIPs基因,分布在8条染色体上,均无内含子结构。系统进化分析表明,PtoS1-bZIP亚家族可分为3个分支,在毛白杨基因组内有12对共线性基因。顺式作用元件分析发现PtoS1-bZIP亚家族成员的启动子中含有多个光信号、激素与非生物胁迫响应元件。荧光定量结果显示,PtoS1-bZIP亚家族成员的表达具有组织特异性。第一和第二进化分支中的多数成员在ABA和干旱处理下表达量上调,在盐胁迫下表达量下调;第三进化分支中的成员在ABA、干旱和盐处理下均表达上调。 结论 从毛白杨中鉴定出10个PtoS1-bZIPs基因,聚类为3个进化分支,第一和第二分支中多数PtoS1-bZIPs成员的表达受干旱胁迫诱导,而受盐胁迫抑制;第三分支成员的表达受干旱和盐胁迫诱导。表明毛白杨PtoS1-bZIPs不同分支成员可能具有功能分化,在响应非生物胁迫过程中发挥不同作用,研究结果为后续解析PtoS1-bZIPs基因调控杨树抗逆性的分子机制提供了基础。 Abstract:Objective To identify S1-bZIP subfamily members in Populus tomentosa and analyze the expression profiles of PtoS1-bZIP genes in different tissues in response to abiotic stress. Method Members of PtoS1-bZIP subfamily were systematically analyzed using bioinformatic approaches. Real-time PCR was performed to explore the expression patterns of PtoS1-bZIP genes in different tissues, as well as under different hormonal or abiotic stress treatment. Result A total of ten intron-lacking PtoS1-bZIP genes were identified in the genome of Populus tomentosa, which were distributed to eight distinct chromosomes. According to phylogenetic analysis, PtoS1-bZIP subfamily members were divided into three sub-clades, designated as Clades I to III. 12 pairs of fragment replicates were identified in the genome of Populus tomentosa. Cis-acting element prediction showed that the promoter regions of PtoS1-bZIP genes were rich of various regulatory elements involving light, hormone or abiotic stress response. qRT-PCR analysis revealed that the members of PtoS1-bZIP subfamily were specifically expressed across different tissues. The majority of PtoS1-bZIP genes in the first and second clades were upregulated under ABA and drought treatments, while expression levels were downregulated under salt treatment. In contrast, all of the PtoS1-bZIP members in the third clades were upregulated under ABA, drought, and salt treatments. Conclusion In the genome of Populus tomentosa, ten PtoS1-bZIP genes are identified and divided into three clades. The expression of major PtoS1-bZIP genes in the first and second clades were induced by drought stress and inhibited by salt stress. The expression of members in the third clades were induced by both drought and salt stress. It indicates that different clades of PtoS1-bZIP genes may have functional differentiation, which play different roles in response to abiotic stresses. Our results lay a foundation for further investigation of the biological functions of PtoS1-bZIP genes and their molecular mechanisms involved in regulating stress resistance of poplars. -
Key words:
- Populus tomentosa
- / S1-bZIP
- / gene family
- / gene expression
-
图 1 毛白杨 (Pto)、毛果杨 (Ptr)、拟南芥 (At)和水稻 (Os) S1-bZIP 亚家族成员的系统发育分析
Figure 1. Phylogenetic analysis of S1-bZIP subfamilies in P. tomentosa (Pto), P. trichocarpa (Ptr), A. thaliana (At) and O. sativa (Os)
图 2 PtoS1-bZIPs 基因结构与保守基序分析
Figure 2. The gene structures and protein conserved motifs of PtoS1-bZIP genes
图 3 PtoS1-bZIPs基因在毛白杨染色体上的定位(A)以及在物种内(B)和物种间(C)的共线性分析
Figure 3. Chromosome localization (A), gene duplication (B) and synteny analysis (C) of PtoS1-bZIP genes
图 4 毛白杨 PtoS1-bZIPs 基因顺式作用元件
Figure 4. Cis-elements of S1-bZIP genes in the genome of P. tomentosa
图 5 PtoS1-bZIPs 基因在不同组织中的表达模式
Figure 5. The expression patterns of PtoS1-bZIP genes in different tissues
图 6 脱落酸(ABA)、盐(NaCl)及干旱(PEG6000)胁迫下 PtoS1-bZIPs 基因在叶片中的表达水平
Figure 6. The expression levels of PtoS1-bZIP genes in leaves under ABA, NaCl and PEG6000 treatment
表 1 PtoS1-bZIPs基因qRT-PCR引物
Table 1. Primers for quantitative real-time PCR of PtoS1-bZIP genes
基因名称
Gene names上游引物(5′→3′)
Forward primers (5′→3′)下游引物(5′→3′)
Reverse primers (5′→3′)PtobZIP55 AGGCCGATAACTCAATCTTG CATTAAAGATGCTTGAATCTCC PtobZIP21 GCGTTACTTGAGTGTTGAGG ATTGAAGGTGATCGAGTCTC PtobZIP27 GAGATCCTGACAAGGATGA TAGTCGCAGAAACAGAGGC PtobZIP44 CGAAGCTGAGAACTCTATCC CAAAAGCACCAGTAGTGGG PtobZIP6 GTCACAACACAGCACTTCTT GCACTAGTTTGGAGATCCTC PtobZIP33 CCATCAATGTCACAACACAGC ATCAGGGAGATCCTCGTGATG PtobZIP63 TCAGGCTCTGAAATTGACCCA AACTTTGTTGTCTGATTCGAGAG PtobZIP49 ATCAGGTTCTGACGTTGATGCG TGTTCTGTGATTCGAGAGC PtobZIP15 GGTCGTTGAACTCTGTGTTGC TTGAAACATATCAGCAGATGCC PtobZIP75 TGTGGAATTGACTGAGAGGC AATAGGCTGTACTGAACACG PtoUBQ10 GTTGATTTTTGCTGGGAAGC GATCTTGGCCTTCACGTTGT  下载: 导出CSV
下载: 导出CSV
表 2 PtoS1-bZIP 亚家族基因特征分析
Table 2. Characteristics analysis of PtoS1-bZIP subfamily genes in P. × tomentosa
基因名称
Gene names基因号
Gene ID染色体
Chr内含子
Introns外显子
Exons氨基酸数目
AA分子量
MW (kDa)等电点
PI亚细胞定位
Subcellular locationPtobZIP55 KAG6762870.1 9 0 1 156 17.30 8.09 Nucleus PtobZIP21 KAG6777875.1 4 0 1 156 17.44 8.10 Nucleus PtobZIP27 KAG6773901.1 5 0 1 163 18.25 6.59 Nucleus PtobZIP44 KAG6766766.1 7 0 1 165 18.71 6.20 Nucleus PtobZIP6 KAG6785745.1 2 0 1 157 17.90 6.18 Nucleus PtobZIP33 KAG6774655.1 5 0 1 159 18.32 6.05 Nucleus PtobZIP63 KAG6757882.1 10 0 1 144 16.51 9.66 Nucleus PtobZIP49 KAG6764077.1 8 0 1 144 16.46 9.36 Nucleus PtobZIP15 KAG6787046.1 2 0 1 145 16.50 8.88 Nucleus PtobZIP75 KAG6749792.1 14 0 1 139 15.78 7.84 Nucleus
下载: 导出CSV
-
[1] 樊松乐, 王纪坤, 覃 碧, 等. 植物转录因子研究方法及应用[J]. 分子植物育种, 2017, 17(15):5003-5009. [2] NG D W K, ABEYSINGHE J K, KAMALI M. Regulating the regulators: the control of transcription factors in plant defense signaling[J]. International Journal of Molecular Sciences, 2018, 19(12): 3737. doi: 10.3390/ijms19123737 [3] SINGH K B, FOLEY R C, OÑATE-SÁNCHEZ L. Transcription factors in plant defense and stress responses[J]. Current Opinion in Plant Biology, 2002, 5(5): 430-436. doi: 10.1016/S1369-5266(02)00289-3 [4] LANDSCHULZ W H, JOHNSON P F, MCKNIGHT S L. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins[J]. Science, 1988, 240(4860): 1759-1764. doi: 10.1126/science.3289117 [5] JAKOBY M, WEISSHAAR B, DRÖGE-LASER W, et al. bZIP transcription factors in Arabidopsis[J]. Trends in Plant Science, 2002, 7(3): 106-111. [6] SCHÜTZE K, HARTER K, CHABAN C. Post-translational regulation of plant bZIP factors[J]. Trends in Plant Science, 2008, 13(5): 247-255. doi: 10.1016/j.tplants.2008.03.002 [7] DRÖGE-LASER W, SNOEK B L, SNEL B, et al. The Arabidopsis bZIP transcription factor family-an update[J]. Current Opinion in Plant Biology, 2018, 45: 36-49. doi: 10.1016/j.pbi.2018.05.001 [8] BANERJEE A, ROYCHOUDHURY A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress[J]. Protoplasma, 2017, 254(1): 3-16. doi: 10.1007/s00709-015-0920-4 [9] HOWELL S H. Endoplasmic reticulum stress responses in plants[J]. Annual Review of Plant Biology, 2013, 64(1): 477-499. doi: 10.1146/annurev-arplant-050312-120053 [10] FU Z Q, DONG X. Systemic acquired resistance: turning local infection into global defense[J]. Annual Review of Plant Biology, 2013, 64(1): 839-863. doi: 10.1146/annurev-arplant-042811-105606 [11] SHEN H S, CAO K M, WANG X P. A conserved proline residue in the leucine zipper region of AtbZIP34 and AtbZIP61 in Arabidopsis thaliana interferes with the formation of homodimer[J]. Biochemical and Biophysical Research Communications, 2007, 362(2): 425-430. doi: 10.1016/j.bbrc.2007.08.026 [12] INABA S, KURATA R, KOBAYASHI M, et al. Identification of putative target genes of bZIP19, a transcription factor essential for Arabidopsis adaptation to Zn deficiency in roots[J]. The Plant Journal, 2015, 84(2): 323-334. doi: 10.1111/tpj.12996 [13] XU C Y, CAO H F, ZHANG Q Q, et al. Control of auxin-induced callus formation by bZIP59–LBD complex in Arabidopsis regeneration[J]. Nature Plants, 2018, 4(2): 108-115. doi: 10.1038/s41477-017-0095-4 [14] DRÖGE-LASER W, WEISTE C. The C/S1 bZIP network: a regulatory hub orchestrating plant energy homeostasis[J]. Trends in Plant Science, 2018, 23(5): 422-433. doi: 10.1016/j.tplants.2018.02.003 [15] YAMASHITA Y, TAKAMATSU S, GLASBRENNER M, et al. Sucrose sensing through nascent peptide-meditated ribosome stalling at the stop codon of Arabidopsis bZIP11 uORF2[J]. FEBS Letters, 2017, 591(9): 1266-1277. doi: 10.1002/1873-3468.12634 [16] PEDROTTI L, WEISTE C, NÄGELE T, et al. Snf1-RELATED KINASE1-controlled C/S(1)-bZIP signaling activates alternative mitochondrial metabolic pathways to ensure plant survival in extended darkness[J]. The Plant Cell, 2018, 30(2): 495-509. doi: 10.1105/tpc.17.00414 [17] WANG H, ZHANG Y T, NORRIS A, et al. S1-bZIP Transcription factors play important roles in the regulation of fruit quality and stress response[J]. Frontiers in Plant Science, 2021, 12: 802802. [18] WEISTE C, PEDROTTI L, SELVANAYAGAM J, et al. The Arabidopsis bZIP11 transcription factor links low-energy signalling to auxin-mediated control of primary root growth[J]. PLOS Genetics, 2017, 13(2): e1006607. doi: 10.1371/journal.pgen.1006607 [19] WELTMEIER F, RAHMANI F, EHLERT A, et al. Expression patterns within the Arabidopsis C/S1 bZIP transcription factor network: availability of heterodimerization partners controls gene expression during stress response and development[J]. Plant Molecular Biology, 2008, 69(1-2): 107-119. [20] IGLESIAS-FERNáNDEZ R, BARRERO-SICILIA C, CARRILLO-BARRAL N, et al. Arabidopsis thaliana bZIP44: a transcription factor affecting seed germination and expression of the mannanase-encoding gene AtMAN7[J]. The Plant Journal, 2013, 74(5): 767-780. doi: 10.1111/tpj.12162 [21] SUN X L, LI Y, CAI H, et al. The Arabidopsis AtbZIP1 transcription factor is a positive regulator of plant tolerance to salt, osmotic and drought stresses[J]. Journal of Plant Research, 2011, 125(3): 429-438. [22] WELTMEIER F, EHLERT A, MAYER C S, et al. Combinatorial control of Arabidopsis proline dehydrogenase transcription by specific heterodimerisation of bZIP transcription factors[J]. The EMBO Journal, 2006, 25(13): 3133-3143. doi: 10.1038/sj.emboj.7601206 [23] HARTMANN L, PEDROTTI L, WEISTE C, et al. Crosstalk between two bZIP signaling pathways orchestrates salt-induced metabolic reprogramming in Arabidopsis roots[J]. The Plant Cell, 2015, 27(8): 2244-2260. doi: 10.1105/tpc.15.00163 [24] 王瑞文, 郭 赟, 周忠诚. 杨树育种研究进展[J]. 湖北林业科技, 2016, 45(6):33-35 + 80. [25] TUSKAN G A, DIFAZIO S, JANSSON S, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray)[J]. Science, 2006, 313(5793): 1596-1604. doi: 10.1126/science.1128691 [26] MA J C, WAN D S, DUAN B B, et al. Genome sequence and genetic transformation of a widely distributed and cultivated poplar[J]. Plant Biotechnology Journal, 2019, 17(2): 451-460. doi: 10.1111/pbi.12989 [27] QIU D Y, BAI S L, MA J C, et al. The genome of Populus alba × Populus tremula var. glandulosa clone 84K[J]. DNA Research, 2019, 26(5): 423-431. doi: 10.1093/dnares/dsz020 [28] AN X M, GAO K, CHEN Z, et al. High quality haplotype-resolved genome assemblies of Populus tomentosa Carr., a stabilized interspecific hybrid species widespread in Asia[J]. Molecular Ecology Resources, 2021, 22(2): 786-802. [29] GU Z L, STEINMETZ L M, GU X, et al. Role of duplicate genes in genetic robustness against null mutations[J]. Nature, 2003, 421(6918): 63-66. doi: 10.1038/nature01198 [30] LYNCH M, CONERY J S. The evolutionary fate and consequences of duplicate genes[J]. Science, 2000, 290(5494): 1151-1155. doi: 10.1126/science.290.5494.1151 [31] ABE H, YAMAGUCHI-SHINOZAKI K, URAO T, et al. Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression[J]. The Plant cell, 1997, 9(10): 1859-1868. [32] ABE H, URAO T, ITO T, et al. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling[J]. The Plant Cell, 2003, 15(1): 63-78. doi: 10.1105/tpc.006130 [33] JEFFARES D C, PENKETT C J, BÄHLER J. Rapidly regulated genes are intron poor[J]. Trends in Genetics, 2008, 24(8): 375-378. doi: 10.1016/j.tig.2008.05.006 [34] HANSON J, HANSSEN M, WIESE A, et al. The sucrose regulated transcription factor bZIP11 affects amino acid metabolism by regulating the expression of ASPARAGINE SYNTHETASE1 and PROLINE DEHYDROGENASE2[J]. The Plant Journal, 2007, 53(6): 935-949. [35] MA J, HANSSEN M, LUNDGREN K, et al. The sucrose-regulated Arabidopsis transcription factor bZIP11 reprograms metabolism and regulates trehalose metabolism[J]. New Phytologist, 2011, 191(3): 733-745. doi: 10.1111/j.1469-8137.2011.03735.x [36] ZHU M K, MENG X Q, CAI J, et al. Basic leucine zipper transcription factor SlbZIP1 mediates salt and drought stress tolerance in tomato[J]. BMC Plant Biology, 2018, 18(1): 83. doi: 10.1186/s12870-018-1299-0 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 483
- HTML全文浏览量: 165
- PDF下载量: 4
- 被引次数: 0
