

-
凋落物分解在森林生态系统养分循环和能量流动起着重要作用[1]。与地上叶凋落物相比, 地下细根一年四季在不停的生长、死亡和分解, 每年对土壤有机质的贡献率最高达到80%, 是生态系统碳和养分输入的主要途径, 尤其是深层土壤中有机质的重要来源[2]。同时对养分受限的土壤也非常重要[3]。据估计, 若森林生态系统物质循环中忽略细根产量、死亡及分解, 对于森林生态系统而言, 土壤中的物质循环将被低估20%~80%[4]。因此研究细根分解对于了解森林生态系统生物化学循环具有重要意义。
三峡库区位于长江流域的咽喉, 属生态敏感区[5]。马尾松(Pinus Massoniana Lamb.)是三峡库区主要植被组成之一, 也是三峡库区分布面积最大的森林类型[6]。目前国内对三峡库区马尾松细根分解开展了相关研究, 主要包括分解动态变化、养分释放(主要是C、N、P元素)、影响因素等方面[5-7-9], 但是系统的研究不同径级根系的较少。本文对三峡库区马尾松 < 0.5 mm、0.5~1 mm和1~2 mm细根进行研究, 探讨不同径级细根的分解速率和C、N、P、K、Ca、Mg养分释放动态变化, 为了解三峡库区马尾松细根分解自然过程和养分循环提供基础。
-
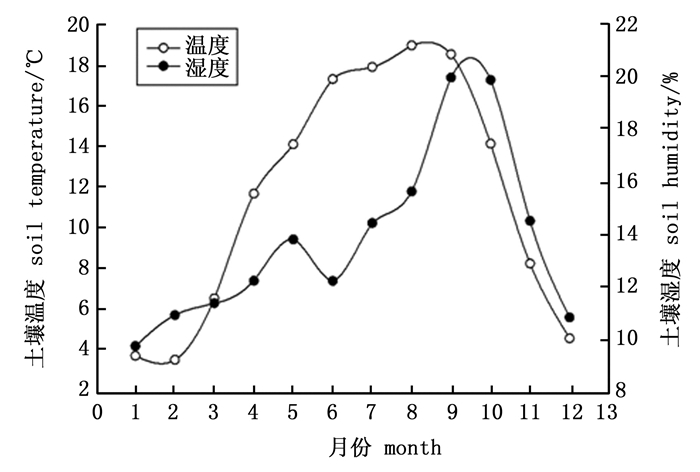
研究地点位于湖北省秭归县三峡库区九岭头林场, 地理位置30° 59′ N, 110° 47′ E, 海拔1 225 m, 属于亚热带季风气候, 年平均温度16.9℃, 年平均降水量范围在1 000~1 250 mL, 多集中在4~9月。区域地带性土壤类型为黄壤或黄棕壤[10]。调查样地内马尾松林为70年代飞播林, 样地内林木分布均匀, 马尾松为优势种, 盖度为80%, 平均胸径和树高分别为20.35 cm和12.4 m。林下灌木平均胸径和高分别为5 cm和5.6 m, 林下灌木主要有火棘(Pyracantha fortuneana (Maxim.) Li), 木姜子(Litsea pungens Hemsl.)等。草本主要有狗脊(Woodwardia japonica (L. F.) Sm)、苔草(Carex tristachya Thunb)、三脉紫菀(Aster ageratoides Turcz)、中日金星蕨(Parathelypteris nipponica (Franch. et Sav.) Ching)等。试验设置3块标准地, 面积为20 m×20 m, 在每个标准地内按上、中、下坡设置3个1 m×1 m的小样方。土壤表层温湿度变化如下图 1。

图 1 土壤表层(0~10 cm)的平均温湿度
Figure 1. Ttemperature and moisture at 0~10 cm soil layer
-
本实验采用埋袋法研究细根分解。于2014年3月, 在林内随机收集表层土中(马尾松细根多集中在土壤0~30 cm处)的根系, 洗净, 去除颜色发暗、无弹性的死根, 自然状态下风干, 分为 < 0.5 mm、0.5~1 mm和1~2 mm。一部分于80℃下烘干至恒重, 用于初始化学成分分析(包括C、N、P、K、Ca、Mg)和测定风干系数。按每袋风干质量1 g装入大小为10 cm×10 cm、孔径为0.1 mm的尼龙网袋内。于当月将网袋埋入各样方10 cm土层处, 表层覆盖凋落物。在2014年5月~2015年3月, 每隔2个月取样一次, 但由于九岭头林场海拔较高, 冬季(次年1月到2月)气温较低, 封山阶段, 故无法进行实验取样。每次随机抽取各径级样品9袋, 除去附着的土壤、杂物和新长入的细根后, 然后迅速用蒸馏水漂洗, 在80℃烘箱中烘干至恒重后, 用于失重测定。每次细根失重测定完后, 将同径级的烘干样品利用液氮研磨, 过100目孔径筛, 用于分解细根C、N、P、K、Ca、Mg的分析。
-
植物根样采用C-N元素分析仪测定马尾松细根C、N含量, HNO3-H2O2消解-ICP法测定细根P、K、Ca、Mg元素含量(相关实验均通过3次重复实验)。
-
$ {\text { 养分保持率 }}(\%)=\frac{C i W i}{C 0 W 0} \times 100 \% $
式中, W0为细根初始干重(g), Wi为细根分解t时间后的干重, C0 为细根初始养分浓度, Ci为细根分解t时间后的养分浓度。
采用单因素方差分析(one way ANOVA)和最小极差法(LSD)检验 < 0.5 mm、0.5~1 mm和1~2 mm细根初始养分浓度的差异显著性。并且采用Person相关分析法分析土壤温度和湿度对细根分解的影响。数据的前期处理、统计分析及绘图分别在Excel2010、SPSS19.0和SigmaPlot11.0中完成。
-
细根初始化学成分分析结果表明, 马尾松不同径级细根C、N、P、K、Ca、Mg含量差别较大(表 1)。细根N、P、K、Ca、Mg含量随直径增大而减小, 且各径级养分两两差异显著(P < 0.05)。1~2 mm细根的C含量明显高于 < 0.5 mm和0.5~1 mm细根的C含量, 且差异显著(P < 0.05), 但 < 0.5 mm和0.5~1 mm细根C浓度差异不显著(P > 0.05)。
表 1 马尾松不同径级细根养分初始浓度
Table 1. Nutrient concentration in different diameters fine roots of Pinus massoniana(mean±SE, n=3)
直径Diameter/mm C/(g·kg-1) N/(g·kg-1) P/(g·kg-1) K/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) < 0.5 mm 442.224±0.119a 9.725±0.011a 0.506±0.002a 3.270±0.021a 6.086±0.076a 1.745±0.010a 0.5~1 mm 444.510±0.194a 7.484±0.022b 0.401±0.012b 2.470±0.018b 5.149±0.039b 1.453±0.012b 1~2 mm 460.430±0.108b 5.470±0.012c 0.306±0.007c 1.657±0.015c 3.498±0.060c 1.078±0.020c 小写字母不同代表差异显著(p < 0.05), 相同代表差异不显著(p < 0.05)。 -
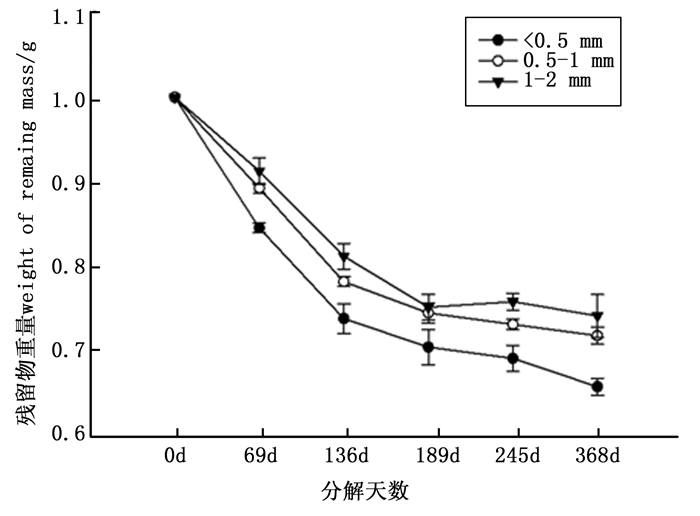
从图 2可以看出, 在马尾松细根分解过程中, 细根残留重量总体上随时间而下降, 且细根分解总体表现为初始阶段分解最快, 后期分解逐渐减慢。细根开始分解的前136 d内, 分解快速, < 0.5 mm、0.5~1 mm和1~2 mm失重率分别达为26.00%、21.67.00%和18.67%, 呈显著差异(p < 0.05), 136 d之后分解速率减缓, 分解368 d后, 马尾松 < 0.5 mm细根约分解34%, 0.5~1 mm约分解28%, 1~2 mm约分解25.72%。

图 2 马尾松不同径级细根残留物重量变化
Figure 2. Dynamics in the remaining mass of Pinus massoniana fine root with different diameters
-
细根(< 0.5 mm、0.5~1 mm、1~2 mm)分解速率与土壤温湿度的Person相关性分析表明(表 2): 土壤温度和各径级细根分解速率呈显著正相关关系(p < 0.05);土壤湿度和各径级细根分解速率呈正相关关系, 其中, < 0.5 mm和0.5~1 mm细根分解速率与土壤湿度未达到显著水平(p < 0.05), 而1~2 mm细根分解速率与土壤湿度达到显著水平(p < 0.05)。
表 2 马尾松不同径级细根分解速率与土壤温湿度的Person相关性
Table 2. Person correlations between soil temperature of moisture anddecomposition rate of Pinus massoniana fine root with different diameters
影响因子
influence factor< 0.5 mm 0.5~1 mm 1~2 mm r p r p r p 土壤温度soil temperature 0.617 0.014 0.817 0.000 0.967 0.000 土壤湿度soil humidity 0.206 0.461 0.356 0.192 0.732 0.002 -
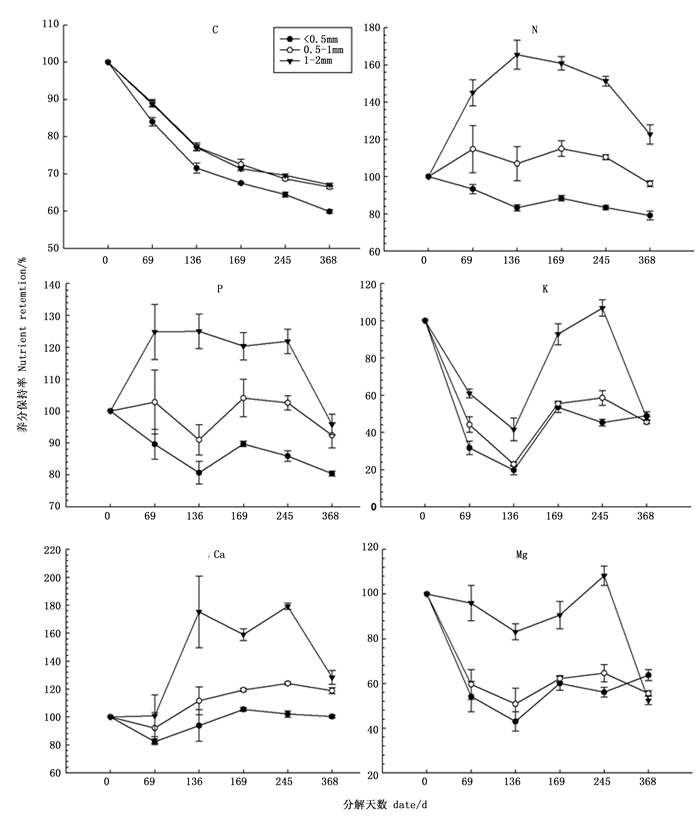
从图 3看出, 随着分解时间增加, 细根C、K、Mg元素均表现为释放状态, 分解到368 d, < 0.5 mm、0.5~1 mm和1~2 mm细根C释放率分别达到40.12%、33.52%和32.92%, K释放率分别为51.09%、54.48%、52.29%, Mg释放率分别为36.32%、44.47%、47.63%;Ca元素整体表现为富集, 保持率分别是100.32%、118.78%、128.3%;N、P元素在不同径级根系分解过程中表现不同, < 0.5 mm细根在整个分解过程中表现为N、P释放, 释放率分别为20.83%、19.61%, 0.5~1 mm细根N和P在分解过程中, 出现释放和富集现象, 在整个分解过程中N表现为释放, 释放率为3.86%, P表现为微弱富集, 保持率为101.41%, 1~2 mm细根N、P元素表现为富集现象, 保持率分别为122.72%、115.77%。

图 3 尾松细根分解过程中养分保持率
Figure 3. Change in nutrient remaining rate of Pinus massoniana roots with different diameters during root decomposition
-
根系分解具有明显的阶段性, 初始阶段分解最快, 后期分解逐渐减慢[11]。一般认为, 细根分解初期较快的原因主要是因为初期细根中碳水化合物的含量相对高, 为真菌和微生物的分解提供了良好条件[12]。本研究中, 马尾松细根3个径级均呈现2个快慢不同的分解阶段, 分解前136 d, 分解较快, 分解368 d后, < 0.5 mm、0.5~1 mm和1~2 mm失重率分别达为26.00%、21.67.00%和18.67%(图 2)。
根分解受到很多环境因子的影响, 如土壤温度、水分等。土壤温度、水分主要是通过影响土壤微生物和酶的活性来影响根分解[13]。细根分解速率随温度升高而增大这一结论已得到多数研究者的证实[11, 13]。本研究同样发现土壤温度与细根分解速率呈显著正相关关系。通过实验发现土壤含水量对细根分解的影响存在一个范围值, 当年降水量为600~2 000 mm时, 呈正相关关系[13], 当降水量过低或过高时, 直接导致土壤含水量的过低或过高, 都不利于土壤微生物生理活性, 从而降低细根分解速率[14]。研究区年平均降水量范围在1 000~1 250 mm, 结果显示土壤湿度与细根分解速率存在正相关关系。
直径大小影响根系分解速率。不同直径细根的化学属性(如N、P含量等)存在显著差异, 表现为随直径增大, N、P含量等减小(表 1)。在相同气候条件下, 细根的化学属性(如C、N、P含量等)决定细根分解速率[15]。如N含量越高, 细根分解越快[16], 表现出直径越大根分解越慢这一普遍规律[17]。本研究结果得出马尾松 < 0.5 mm细根分解速率最快, 1~2 mm细根分解速率最慢, 与王瑞丽等[5]、Fan等[18]的研究结果基本相同, 均符合这一结论。但也有研究表明, 根系的分解速率不是与直径存在始终一致的相关关系, 尤其是细根, 出现异速分解现象, 表现为直径越小分解速率越低[13, 19]。Sun等[20]对4种亚热带树种细根连续4年的分解研究发现 < 0.5 mm细根的分解速率小于0.5~2 mm细根的分解速率。林成芳等[7]对米储细根分解的研究同样发现1~2 mm细根的分解速率高于0~1 mm的细根。这可能与细根中N浓度高, 易与木质素结合成N-木质素络合物从而抑制细根分解有关[21]。也有人解释此种现象是因为外生菌根的作用。在植物细根中, 直径越小, 菌根侵染的程度就越大, 菌根极大地改变了细根本身的物理结构和化学性质, 形成抗分解的保护机制, 降低了分解速率(即“菌根假说”)[22-23]。本研究结果在一定程度上否定以上外生菌根对分解的抑制作用。这种差异的存在可能是因为不同外生菌根真菌对细根分解的速率不同而引起的[24]。
-
细根分解过程中表现的养分释放或富集将影响细根对土壤养分的贡献[25]。本研究结果表明马尾松细根C、K、Mg元素迁移模式表现为释放, 平均释放率达到44%, Ca元素基本表现为富集, 而N、P元素在不同径级中表现不同, 其中直径 < 0.5 mm的细根在分解时表现为N、P释放, 而直径0.5~2 mm的细根在分解时存在N、P富集阶段, 与Wang等[26]对直径 < 0.5 mm和直径0.5~2 mm的细根在分解时元素迁移模式相似。从理论上来说, 在分解过程中, 养分随分解最终应释放到土壤中, 但对于分解1年后, 存在某些元素表现为富集这个现象, 是否此元素释放存在一个时间的滞后性, 这还需进一步的研究证实。
细根分解过程中, 初始养分浓度影响养分释放[7, 27]。根系中若某个养分元素的浓度相对较高, 能满足微生物所需, 则表现为释放; 相反, 浓度低的养分元素一般容易发生富集[28]。直径越小, 马尾松细根N、P元素等浓度越高, 且两两之间差异显著(表 1)。这可能从一方面解释了马尾松不同径级细根N、P等元素表现为释放或积累的现象。同时, 研究表明初始养分浓度大小存在阈值可作为养分释放或积累的分界值。Chen等[29]估算了根系分解过程中N释放或富集的初始N浓度平均阈值为0.4%, 如果大于0.4%, 则释放N, 如果小于0.4%, 则富集N。Guo等[30]对澳大利亚辐射松根的研究结果符合这一观点。本研究中马尾松 < 0.5 mm、0.5~1 mm细根初始N浓度分别为0.97%、0.75%, 远大于0.4%, 分解一年后均表现为N释放, 符合以上结论。
环境因子影响细根的养分释放[7, 27]。降雨, 一方面通过淋溶作用加速了养分的释放, 尤其是如K这类较活跃的元素, 另一方面可改变了土壤水分含水量, 土壤水分通过影响分解者的活动以及微生物的活性影响其养分释放。如细根C元素的释放动态变化, 从雨季到非雨季期间, 释放速率逐渐减小, 与张秀娟等[31]、李吉枚等[32]研究结果一致。同时在研究中发现, 直径越小, 细根养分释放越易受到降水的影响。但是相比其它元素而言, N、P养分释放过程比较复杂, 在分解过程中, 直径0.5~2 mm的细根会出现富集现象(图 3)。土壤养分状况影响细根养分释放。Osterag等[33]对铁心木的研究结果发现在缺磷立地上分解一年后N含量是初始的2.5倍, P含量是初始的1.5倍。从全球尺度角度来说, 热带或亚热带地区土壤P含量普遍偏低[34]。三峡库区属于低纬度、亚热带季风气候区, 马尾松直径 < 0.5 mm细根分解368 d后N、P表现为不同程度的富集(图 3)。但究竟是什么原因导致N、P的富集目前还不清楚, 可能与分解微生物对元素的固定有关。
元素本身的特性和在根系中存在的形态也可能影响细根养分释放[7]。马尾松细根C、K、Mg元素表现为释放, Ca元素表现为富集, 与李吉枚等[32]、翟明普等[35]、Yang等[36]的研究结果类似。元素C、K、Mg在植物组织中较为活跃, 移动性较强, 其中元素K还是非植物细根的结构性成分, 不需要微生物活动的参与, 最易受到外界环境条件如降水导致的淋溶过程等的影响, 而Ca是植物组织内稳定性强的元素, 移动性差, 并且是植物组织的构建成分[37], 因此表现出不同程度的释放或富集模式。
-
本研究分析了不同径级马尾松细根分解速率以及细根分解过程中C、N、P、K、Ca、Mg养分释放动态变化, 得出以下结论:
1) 细根分解速率与土壤温度和直径大小呈显著相关。其中细根分解速率随温度的升高而增大, 随直径的增大而减小;
2) 在细根分解过程中, 不同养分元素的表现状态不同。其中, 直径 < 0.5 mm、0.5~1mm、1~2mm细根C、K、Mg元素表现为释放和Ca元素表现为富集; 直径 < 0.5 mm细根N、P元素表现为释放, 0.5~2 mm细根N、P元素表现为释放-富集;
3) 在细根分解过程中, 元素初始养分浓度、降水、土壤养分状况, 以及元素本身的特性和在根系中存在的形态都影响细根养分释放。
三峡库区马尾松细根分解及其养分释放
Fine Root Decomposition and Nutrient Release of Pinus Massonianain the Three Gorges Reservoir Area
-
摘要:
目的 为认识马尾松细根对土壤养分库的贡献, 方法 本研究以三峡库区秭归县九岭头林场马尾松为研究对象,采用埋袋法进行细根分解实验,探讨 < 0.5 mm、0.5~1 mm和1~2 mm细根的分解动态和养分释放(C、N、P、K、Ca、Mg)。 结果 结果表明:(1)细根分解368 d后, < 0.5 mm、0.5~1 mm和1~2 mm细根干重残留率分别为66.0%、72.0%和74.33%,且细根分解速率随直径增加而减小;(2)细根分解速率与土壤温度呈显著正相关关系(p < 0.05),与土壤湿度呈正相关关系(p < 0.05);(3)细根C、K和Mg元素迁移模式表现为释放,Ca元素表现为富集;(4)细根N、P元素在不同径级细根中迁移模式不同, < 0.5 mm细根N、P元素表现为释放,0.5~2 mm细根N、P元素在分解过程中出现富集阶段。 结论 马尾松细根分解与土壤温度和直径大小显著相关,其中分解与温度呈正相关,与直径呈负相关;在细根分解过程中,马尾松细根不同直径大小的不同养分元素的表现状态不一致,或富集,或释放。 Abstract:Objective The purpose of this study is to determine the contribution of fine root ofPinus massoniana L. on soil nutrient pool. Method Decomposition experiment, using the buried bag method, was conducted to examine < 0.5 mm, 0.5~1 mm and 1~2 mm fine root decomposition dynamics and nutrient release (C, N, P, K, Ca, Mg) in Pinus massoniana at Jiulingtou Forest Farm of Zigui county, Three Gorges Reservoir Area, China. Result The results showed that: (1) the decomposition rate decreased with root diameter increasing of Pinus massoniana, and after 368 days, the remaining percentage of < 0.5 mm, 0.5~1 mm and 1~2 mm fine root were 66.0%, 72.0% and 74.33% respectively. (2) The soil temperature had significant positive effect on decomposition rate, and soil moisture had positive effect on decomposition rate. (3) The transfer model for C, K and Mg of fine root was releasing, while enriching for Ca. (4) The transfer models for N and P of < 0.5 mm, 0.5~1 mm and 1~2 mm fine root were different, the transfer models for N and P of < 0.5 mm fine root were releasing, but N and P of 0.5~2 mm fine root appeared enriching stage during the decomposition process. Conclusion Fine root decomposition rate of Pinus massoniana and soil temperature significantly positivel correlated, andy decomposition rate and diameter significantly negatively correlated; In decomposition process, different nutrient elements of different root diameter performance status were different, or enrichment, or release. -
图 2 马尾松不同径级细根残留物重量变化
Figure 2. Dynamics in the remaining mass of Pinus massoniana fine root with different diameters
图 3 尾松细根分解过程中养分保持率
Figure 3. Change in nutrient remaining rate of Pinus massoniana roots with different diameters during root decomposition
表 1 马尾松不同径级细根养分初始浓度
Table 1. Nutrient concentration in different diameters fine roots of Pinus massoniana(mean±SE, n=3)
直径Diameter/mm C/(g·kg-1) N/(g·kg-1) P/(g·kg-1) K/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) < 0.5 mm 442.224±0.119a 9.725±0.011a 0.506±0.002a 3.270±0.021a 6.086±0.076a 1.745±0.010a 0.5~1 mm 444.510±0.194a 7.484±0.022b 0.401±0.012b 2.470±0.018b 5.149±0.039b 1.453±0.012b 1~2 mm 460.430±0.108b 5.470±0.012c 0.306±0.007c 1.657±0.015c 3.498±0.060c 1.078±0.020c 小写字母不同代表差异显著(p < 0.05), 相同代表差异不显著(p < 0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 马尾松不同径级细根分解速率与土壤温湿度的Person相关性
Table 2. Person correlations between soil temperature of moisture anddecomposition rate of Pinus massoniana fine root with different diameters
影响因子
influence factor< 0.5 mm 0.5~1 mm 1~2 mm r p r p r p 土壤温度soil temperature 0.617 0.014 0.817 0.000 0.967 0.000 土壤湿度soil humidity 0.206 0.461 0.356 0.192 0.732 0.002
下载: 导出CSV
-
[1] Yuan Z, Chen H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses[J]. Critical Reviews in Plant Sciences, 2010, 29(4): 204-221. doi: 10.1080/07352689.2010.483579 [2] 陈曦, 张乃莉, 周晓梅, 等. 细根分解研究进展及存在问题[J]. 吉林师范大学学报: 自然科学版, 2012, (2): 36-40. [3] 林成芳, 杨玉盛, 陈光水, 等. 杉木人工林细根分解和养分释放及化学组成变化[J]. 亚热带资源与环境学报, 2008, 3(1): 15-23. doi: 10.3969/j.issn.1673-7105.2008.01.003 [4] 王存国, 韩士杰, 周玉梅, 等. 长白山阔叶红松林群落的细根现存量及养分内循环[J]. 林业科学, 2012, 48(3): 0148-0153. [5] 王瑞丽, 程瑞梅, 肖文发, 等. 三峡库区马尾松人工林细根生产和周转[J]. 应用生态学报, 2012, 23(9): 2346-2352. [6] 肖文发, 雷静品. 三峡库区森林植被恢复与可持续经营研究[J]. 长江流域资源与环境, 2004, 13(2): 138-144. doi: 10.3969/j.issn.1004-8227.2004.02.008 [7] 林成芳. 中亚热带森林细根分解动态及影响因素[D]. 福建: 福建师范大学, 2008. [8] 罗达. 南亚热带格木、马尾松幼龄纯林及其混交林碳氮特征研究[D]. 北京: 中国林业科学研究院, 2014. [9] 王卫霞. 南亚热带不同树种人工林生态系统碳氮特征研究[D]. 北京: 中国林业科学研究院, 2013. [10] 葛晓改, 肖文发, 曾立雄, 等. 三峡库区不同林龄马尾松土壤养分与酶活性的关系[J]. 应用生态学报, 2012, (02): 445-451. [11] Berg B, McClaugherty C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration[J]. Berlin: Springer, 2003. [12] Lin C F, Yang Y S, Guo J F, et al. Fine root decomposition of evergreen broadleaved and coniferous tree species in mid-subtropical China: dynamics of dry mass, nutrient and organic fractions[J]. Plant Soil, 2010, 338: 311-327. [13] 许玉庆, 项文化, 曾叶霖, 等. 中国森林生态系统细根分解格局及调控因子研究进展[J]. 广西林业科学, 2015, 44(2): 149-155. doi: 10.3969/j.issn.1006-1126.2015.02.012 [14] 王新源, 赵学勇, 李玉霖, 等. 环境因素对干旱半干旱区凋落物分解的影响研究进展[J]. 应用生态学报, 2013, 11: 3300-3310. [15] Silver W L, Myia R K. Global patterns in root decomposition: comparisons of climate and litter quality effects[J]. Oecologia, 2001, 129(3): 407-419. doi: 10.1007/s004420100740 [16] Hishi T. Heterogeneity of individual roots within the fine root architecture: causal links between physiological and ecosystem functions[J]. Journal of Forestry Research, 2007, 12(2): 126-133. doi: 10.1007/s10310-006-0260-5 [17] 勒贝贝, 国庆喜. 蒙古栎、白桦根系分解及养分动态[J]. 生态学报, 2013, (8): 2416-2424. [18] Fan P P, Guo D L. Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soil[J]. Oecologia, 2010, 163(2): 509-515. doi: 10.1007/s00442-009-1541-4 [19] 刘利芳, 徐程杨. 影响森林细根分解的机理研究进展[J]. 山东林业科技, 2012, 3: 0097-0105. [20] Sun T, Mao Z J, Han Y Y. Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in for temperate tree species[J]. Plant and Soil, 2013, 372(1): 445-458. [21] Chen H, Harmon M E, Grifiths R E. Decomposition and nitrogen relaese from decomposing woody roots in coniferous forests of the Pacific Northwest[J]. Can J For Res, 2001, 31: 246-260. doi: 10.1139/x00-167 [22] Langley J A, Hungate B A. Mycorrhizal controls on below-ground litter quality[J]. Ecology, 2003, 84: 2302-2312. doi: 10.1890/02-0282 [23] Langley J A, Chapman S K, Hungate B A. Ectomycorrhizal colonization slows root decomposition: the postmortem fungl legacy[J]. Ecology Letters, 2006, 9: 955-959. doi: 10.1111/j.1461-0248.2006.00948.x [24] Koide R T, Malcolm G M. N concentration controls decomposition rates of different strains of ectomycorrhizal fungi[J]. Fungal Ecology, 2009, 2: 197-202. doi: 10.1016/j.funeco.2009.06.001 [25] 王存国, 陈正侠, 马承恩, 等. 细根异速分解的3各可能影响途径[J]. 北京林业大学学报, 2016, 4(38): 123-128. [26] Wang W, Zhang X Y, Tao N, et al. Effects of litter types, microsite and root diameters on litter decomposition in Pinus silvestris plantations of north China[J]. Plant and Soil, 2014, 374(1-2): 677-688. doi: 10.1007/s11104-013-1902-y [27] Hobbie S E, Vitousek P M. Nutrient limitation of decomposition in Hawaiian forests[J]. Ecology, 2000, 81: 1867-1877. doi: 10.1890/0012-9658(2000)081[1867:NLODIH]2.0.CO;2 [28] 王瑾, 黄建辉. 暖温带地区主要树种叶片凋落物分解过程中主要元素释放的比较[J]. 植物生态学报, 2001, 25(3): 375-380. doi: 10.3321/j.issn:1005-264X.2001.03.019 [29] Chen H, Harmon M E, Sexton J, et al. Fine-root decomposition and N dynamics in coniferous forests of the Pacific Northwest, USA[J]. Canadian Journal of Forest Research, 2002, 32: 320-331. doi: 10.1139/x01-202 [30] Guo L B, Halliday M J, Gifford R M. Fine root decomposition under grass and pine seedlings in controlled environmental conditions[J]. Applied Soil Ecology, 2006, 33(1): 22-29. doi: 10.1016/j.apsoil.2005.09.004 [31] 张秀娟, 吴楚, 梅莉, 等. 水曲柳和落叶松人工林根系分解与养分释放[J]. 应用生态学报, 2006, 17(8): 1370-1376. doi: 10.3321/j.issn:1001-9332.2006.08.003 [32] 李吉枚, 张毓涛, 李建贵, 等. 模拟氮沉降对天山云杉细根分解及其养分释放的影响[J]. 生态环境学报, 2015, 35(1): 0182-0188. [33] Ostertag R, Hobbie S E. Early stages of root and leaf decomposition in Hawaiian forest effect of nutrient availability[J]. Oecologia, 1999, 121: 564-573. doi: 10.1007/s004420050963 [34] Lambers H, Brundrett M C, Raven J A, et al. Plant mineral nutrition in ancient landscapes: high plant species diversityon infertile soils is linked to functional diversity for nutritional strategies[J]. Plant and Soil, 2011, 348: 7-27. doi: 10.1007/s11104-011-0977-6 [35] 翟明普, 蒋三乃, 贾黎明. 杨树刺槐混交林细根养分动态研究[J], 林业科学, 2004, 40(4): 0046-0051. [36] Yang Y S, Chen G S, Guo J F, et al. Decomposition dynamic of fine roots in a mixed forest of Cunninggamia laceolate and Tsoongiodendron odorum in midsubtropics[J]. Annals of Forest Science, 2004, 61: 65-72. doi: 10.1051/forest:2003085 [37] 陈灵芝, 黄建辉, 严昌荣. 中国森林生态系统养分循环[M]. 北京: 中国气象出版社, 1997. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3918
- HTML全文浏览量: 821
- PDF下载量: 730
- 被引次数: 0
