

-
生境地是指某一植物具体生长地段的生物和非生物因子的综合。生境地在植物生活史中具有重要作用, 是个体、种群完成其生命过程的空间[1-2], 是植物一切生命活动和生命过程的发生基底, 其质量和适宜性高低直接影响植物的分布、数量、存活和繁殖等[3-4]。随着人类生产方式及活动范围的改变, 大量植物赖以生存的环境发生改变, 适宜生境逐渐减少甚至消失, 导致大量物种面临灭绝的风险。因此, 开展植物生境地的研究, 了解植物生境需求, 是植物生物学及生态学研究的基础[5], 也是探索濒危植物的濒危机制及制定保护策略的关键。
藤枣(Eleutharrhena macrocarpa (Diels) Forman)为防己科(Menispermaceae)藤枣属(Eleutharrhena Forman)多年生木质藤本植物, 主要分布于印度阿萨姆、云南(西部、西南部、南部、东南部)和广西西南部的低山沟谷湿热地区, 并以中国云南南部为分布中心[6], 是中国I级重点保护野生植物、世界自然保护联盟的极危种及云南省极小种群保护物种。近年来, 由于社会经济发展及人类对自然的破坏, 藤枣种群数量急剧降低, 因此, 加大对藤枣种群及其生境的保护刻不容缓。目前, 国内外有关藤枣的研究极为缺乏, 主要集中于花粉形态[7]、叶片解剖学[8-10]、种群分布及种群状况[6]。本文以藤枣模式标本采集地普洱地区藤枣分布区为研究对象, 通过对藤枣分布地植被、环境及藤枣种群的调查, 探索藤枣种群与生境地特征, 为深入分析藤枣种群濒危机制奠定基础。
-
研究区域位于云南省普洱市的太阳河自然保护区。地理位置为22.53°~22.63°N, 101.04°~101.30° E, 最高峰海拔1 707 m, 地势总体上是由北与西北向南与东南倾斜, 地貌主体为山地。该区地处热带北缘向南亚热带的过渡地区, 气候主要受印度洋及太平洋季风控制, 形成夏秋季多雨、冬春季干旱、年温差小、日温差大、干湿季明显的气候特征[11]。该地区年平均气温17.7 ℃, 年降水量超过1 500 mm, 雨水主要集中在5—10月, 占全年降水量的87%, 年平均蒸发量1 590 mm, 日照时数约为2 200 h, 太阳总辐射量为130 333卡·cm-2。土壤以赤红壤和山地红壤为主[11]。
云南太阳河自然保护区主要植被类型包括季风常绿阔叶林、季节雨林等。季风常绿阔叶林是该区面积最大、保存最完整的植被类型, 占保护区总面积的60.8%。以短刺锥(Castanopsis echidnocarpa Miq.)、红锥(Castanopsis hystrix Miq.)、西南木荷(Schima wallichii (DC.) Choisy)为代表性物种的季风常绿阔叶林中附生有大量蕨类和兰科(Orchidaceae)植物, 此外, 板根现象和层间植物也是该地区季风常绿阔叶林的一个重要特征。以绒毛番龙眼、千果榄仁为标志的季节雨林, 在滇南、滇西南有较广泛的分布, 构成滇南的地带性植被[12]。云南太阳河自然保护区是该群系的最北分布端, 分布区海拔一般低于1 300 m。季节雨林是保护区生物区系富集的重要生态系统。
-
在云南太阳河自然保护区内, 选择藤枣的主要分布地设置调查样地。样地面积为20 m×20 m。根据不同地点(4个)藤枣分布数量及地形地势, 共设置调查样地5块。利用网格法将每个样地分割成16个5 m×5 m的调查样方, 在样方内对所有胸径(DBH)≥1 cm的植物进行每株调查, 主要记录物种名称、高度、胸径并进行定位。在样地四角及中心样方内各设置1个2 m×2 m的灌木和草本调查小样方, 对小样方内所有植物记录其物种名称、高度、株数(草本为多度)、盖度等。与此同时, 对藤枣在样地出现情况进行全面调查, 记录其长度、基径、胸径(如果有)及坐标。在植被调查的同时, 记录每个样地的郁闭度、海拔、坡度、裸岩覆盖率、距河流距离等环境因子, 并在样地四角及中心位置各取0~10 cm深土壤样品约500 g, 带回实验室进行土壤理化性质分析。
-
(1) 样地的林分因子及物种组成: 根据野外调查数据, 计算各样地平均高、平均胸径、胸高断面积和林分密度等林分因子; 统计各样地物种数量及物种组成情况, 并利用公式重要值=(相对多度+相对盖度+相对频度)/3计算物种重要值。
(2) 群落大小结构: 径级结构按上限排外法共划分6级: Ⅰ(DBH < 5 cm)、Ⅱ(5≤DBH < 10 cm)、Ⅲ(10≤DBH < 20 cm)、Ⅳ(20≤DBH < 40 cm)、Ⅴ(40≤DBH < 100 cm)和Ⅵ(DBH≥100 cm), 分别统计每个样地各径级树木个体多度及物种数, 计算不同径级树木的物种丰富度(平均值±标准误)及个体多度(平均值±标准误)。根据研究的群落特征, 本文高度级结构按上限排外法共划分5级: Ⅰ(H < 5 m)、Ⅱ(5≤H < 10 m)、Ⅲ(10≤H < 20 m)、Ⅳ(20≤H < 30 m)和Ⅴ(H≥30 m)。分别统计各样地不同高度级内树木个体数及物种数, 计算不同高度级树木的物种丰富度(平均值±标准误)及个体多度(平均值±标准误)。
(3) 种群数量及更新: 统计各样地出现的藤枣个体数量, 计算其平均长、平均基径、平均胸径, 绘制其个体数量频率分布图, 并将长度 < 33 cm的个体作为其更新幼苗, 统计其数量及出现的频率等信息。
-
文中所有数据处理与统计均利用SPSS19.0和EXCEL完成。图形利用SigmPlot12.5绘制。
-
在藤枣生境地, 样地林分平均高为11.98 m(9.78 m~14.27 m)、平均胸径为15.13 cm(9.88 cm~18.14 cm)、平均胸高断面积41.42 m2·hm-2(18.99 m2·hm-2~67.90 m2·hm-2)、平均密度1 205株·hm-2(875株·hm-2~1 625株·hm-2)(表 1)。各样地平均高、平均胸径、林分密度数值显示出藤枣生境地林分高度较高, 树木较大, 密度较大, 整体显示出群落的老龄林特征。
表 1 不同样地林分因子
Table 1. Stand factors in different plots
样地号
Plots平均高
Mean height/ m平均胸径
Mean DBH/cm胸高断面积
Basal areas/(m2 · hm-2)密度
Density/(stem · hm-2)1 10.93 15.14 33.61 1 125 2 11.54 15.34 27.68 925 3 13.37 18.14 67.90 1 475 4 9.78 9.88 18.99 1 625 5 14.27 17.13 58.91 875 平均Mean 11.98 15.13 41.42 1 205 -
藤枣生境地中, 各样地物种丰富度在31~42之间, 平均为35.2(表 2)。其中, DBH≥1 cm的物种丰富度平均为21.8(范围为15~28), 而灌草样方中物种丰富度则为22.2(范围为12~29)。
表 2 不同样地物种丰富度
Table 2. Species richness in different plots
样地号
PlotsDBH≥1 cm 灌草
Shrub and herb总物种数
Total species1 24 12 35 2 15 22 31 3 26 24 35 4 28 29 42 5 16 24 33 平均Mean 218 22.2 35.2 通过对所调查样地进行数据统计与计算, 不同样地重要值前5位的物种列于表 3中。结合各样地物种组成情况, 尽管各样地重要物种不尽相同, 但样地1、2、3、5中主要物种均为热带雨林物种, 而样地4中的物种则为季风常绿阔叶林物种, 可见, 在太阳河自然保护区内藤枣主要分布在热带雨林和季风常绿阔叶林内。
表 3 不同样地重要值前5位的物种
Table 3. The top five species of the important values in different plots
样地号
Plot物种
Species相对盖度
Relative coverage相对多度
Relative abundance相对频度
Relative frequency重要值
Important value1 棒柄花(Cleidion brevipetiolatum Pax et Hoffm.) 0.067 1 0.2093 0.2105 0.1623 1 线毛番龙眼(Pometia tomentosa (Bl.) Teysm. et Binn.) 0.108 6 0.0698 0.0789 0.0858 1 香桂(Cinnamomum subavenium Miq.) 0.1715 0.0233 0.0263 0.0737 1 越南安息香(Styrax tonkinensis (Pierre) Craib ex Hartw.) 0.1628 0.0233 0.0263 0.0708 1 大叶桂樱(Laurocerasus zippeliana (Miq.) Yü et Lu) 0.0700 0.0698 0.0526 0.0641 2 歪叶榕(Ficus cyrtophylla Wall, ex Miq.) 0.2417 0.2973 0.2727 0.2706 2 景洪暗罗(Polyalthia cheliensis Hu) 0.0986 0.1622 0.1515 0.1374 2 假桂钓樟(Linderca tonkinensis Lec.) 0.2091 0.0270 0.0303 0.0888 2 秋枫(Bischofia javanica Bl.) 0.1085 0.0541 0.0606 0.0744 2 木奶果(Baccaurea ramiflora Lour.) 0.0114 0.1081 0.0909 0.0701 3 秋枫(Bischofia javanica Bl.) 0.3522 0.1017 0.0435 0.1658 3 线毛番龙眼(Pometia tomentosa (Bl.) Teysm. et Binn.) 0.0724 0.1695 0.1522 0.1314 3 顶果树(Acrocarpus fraxinifolius Wight ex Arn.) 0.2834 0.0169 0.0217 0.1074 3 棒柄花(Cleidion brevipetiolatum Pax et Hoffm.) 0.0204 0.1017 0.1087 0.0769 3 歪叶榕(Ficus cyrtophylla Wall, ex Miq.) 0.0554 0.0508 0.0652 0.0572 4 耳叶柯(Lithocarpus grandifolius (D. Don) Biswas) 0.2460 0.1077 0.1017 0.1518 4 牛矢果(Osmanthus matsumuranus Hayata) 0.0408 0.1077 0.1017 0.0834 4 鱼尾葵(Caryota ochlandra Hance) 0.1362 0.0615 0.0508 0.0828 4 密花树(Rapanea nerifolia (Sieb. et Zucc.) Mez) 0.0356 0.0769 0.0678 0.0601 4 大肉实树(Sarcosperma arboreum Hook. f.) 0.0254 0.0615 0.0678 0.0516 5 千果榄仁(Terminalia myriocarpa Vaniot Huerck et Muell. -Arg.) 0.7216 0.0571 0.0667 0.2818 5 香芙木(Schoepfia fragrans Wall.) 0.0488 0.2571 0.2000 0.1686 5 菜阳河杮(Dispyros caiyangheiensis Tao) 0.0211 0.2000 0.1667 0.1293 5 山牡荆(Vitex quinata (Lour.) Wall.) 0.0633 0.0571 0.0667 0.0624 5 香桂(Cinnamomum subavenium Miq.) 0.037 2 0.0571 0.0667 0.0537 -
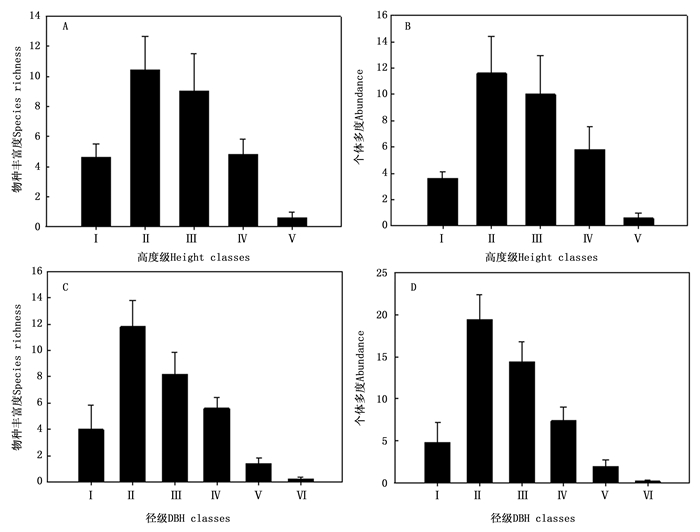
在藤枣分布区内, 不同高度级内物种丰富度和个体多度均呈单峰曲线, 最高值均出现在第Ⅱ高度级(5≤H < 10 m)(图 1A和图 1B), 但第Ⅴ(H≥30 m)高度级仍有一定量的物种和个体出现。与高度级相似, 不同径级内物种丰富度和个体多度也呈单峰曲线, 最高值均出现在第Ⅱ径级(5≤DBH < 10 cm)(图 1C和图 1D), 但第Ⅴ(40≤DBH < 100 cm)径级特别是第Ⅵ(DBH≥100 cm)径级仍有一定量的物种和个体出现, 表明了群落的原始性。

图 1 不同大小级物种丰富度与多度
Figure 1. Species richness and abundance in different size classes
在季风常绿阔叶林(样地4)中, 不同径级和高度级物种丰富度与个体多度分布特征与藤枣分布区内总体分布特征(图 1)相似, 但高度级缺少Ⅴ级, 径级缺少Ⅵ级。最大树高仅为21 m(耳叶柯), 最大胸径为41.8 cm(耳叶柯)。在3层(H < 5 m为灌木层, 5 m≤H < 15 m林下层, H≥15 m为林冠层)的乔木分层中, 林冠层中物种共有8种, 主要包括耳叶柯、红锥(Castanopsis hystrix Miq.)、截果柯(Lithocarpus truncatus (King) Rehd. et Wils.)、黑黄檀(Dalbergia fusca Pierre)等; 林下层中物种共有24种, 牛矢果与耳叶柯个体最多, 胸高断面积同样牛矢果与以耳叶柯最高; 而灌木层中仅有物种4种, 以厚皮香(Ternstroemia gymnanthera (Wight et Arn.) Beddome)和牛矢果为主。
在热带雨林(样地1、2、3、5)中, 不同径级和高度级物种丰富度与个体多度分布特征与藤枣分布区内总体分布特征(图 1)相似。最大树高为32 m(越南安息香), 最大胸径为146 cm(千果榄仁)。H≥30 m共有物种3种, 分别为顶果树、越南安息香和假柿木姜子(Litsea monopetala (Roxb.) Pers.); 树高在20~30 m之间共有物种19种, 绒毛番龙眼和秋枫个体最多, 而秋枫和和千果榄仁胸高断面积最大; 树高在10~20 m之间共有物种25种, 歪叶榕和绒毛番龙眼个体最多, 胸高断面积最大; 树高在5~10 m之间共有物种29种, 木奶果和棒柄花个体最多, 胸高断面积最大; 在H < 5 m共有物种18种, 绒毛番龙眼、香芙木和景洪暗罗个体最多, 绒毛番龙眼和景洪暗罗胸高断面积最大。
-
藤枣生境地中, 海拔范围在964~1 250 m, 平均为1 107 m, 坡度≥22°, 平均值达到陡坡范围(表 4), 坡向则多数为阴坡(北坡), 均处坡下位置。林分郁闭度为76%, 距河流较近5 m~70 m, 平均为31 m, 并伴有一定的裸露岩石(表 4)。藤枣生境土壤接近中性, 平均pH值为6.2, 土壤养分条件较好, 有机质、N、P、K及其速效成分含量较高(表 4)。可见, 藤枣生境总体显示出隐蔽、高湿度、高肥性、动物在其分布区内活动的不易性等特点。
表 4 藤枣生境地环境特征
Table 4. Environment characteristic in the habitat of Eleutharrhena macrocarpa
环境因子Environmental factors 1 2 3 4 5 平均值Mean 海拔Altitude /m 1 052 1 032 964 1 237 1 250 1 107 坡度Slope /。 22 31 31 22 27 26.6 坡向Slope aspect 东坡 北坡 北坡 南坡 北坡 - 坡位Slope position 坡下 坡下 坡下 坡下 坡下 坡下 郁闭度Canopy density 80 70 85 80 65 76 距河流距离Distance to rivers /m 5 70 45 25 10 31 裸岩覆盖率Rocky’s cover /% 5 15 10 8 18 11.2 pH 6.61 6.49 6.52 5.97 5.43 6.20 有机质Organic matter/ (g · kg -1) 120.48 108.54 105.38 95.34 50.24 96.00 全氮Nitrogen/% 0.47 0.43 0.42 0.42 0.25 0.40 全磷Phosphorus/% 0.19 0.15 0.16 0.07 0.08 0.13 全钾Potassiunm% 1.95 2.19 2.26 1.07 0.7 1.67 水解性氮Hydrolysable nitrogen/(mg · kg-1) 359.90 337.50 335.50 372.10 275.51 336.10 有效磷Available phosphorus/(mg · kg-1) 89.65 54.89 57.76 19.38 55.92 55.52 速效钾Available potassium (mg · kg -1) 340.10 230.45 228.00 218.20 247.15 252.78 -
本次调查共发现藤枣28株, 3号样地中个体数量最多为10株, 其次为4号样地9株, 而在1号和5号样地中仅出现1株, 平均每个样地出现5.6株。28株藤枣平均长度为2.39 m, 分布在0.1~10 m之间, 其平均基径仅为0.74 cm, 最大值为1.7 cm, 平均胸径为0.41 cm, 分布在0.2~1.3 cm之间(表 5)。
表 5 藤枣种群数量
Table 5. Population numbers of Eleutharrhena macrocarpa
样地号
Plots个体多度
Abundance平均长度
Mean length/ m平均基径
Mean basal diameter /cm平均胸径
Mean DBH /cm1 1 1.2 0.45 - 2 7 2.38 1.04 0.38 3 10 2.74 0.60 0.33 4 9 4.03 1.02 0.73 5 1 1.6 0.58 0.2 平均Mean 5.6 2.39 0.74 0.41 -
5 m×5 m的样方中, 共有19个样方分布有藤枣。其中, 仅分布1株的有15个样方, 分布2株的有2个样方, 分布4株和5株的各有1个样方。
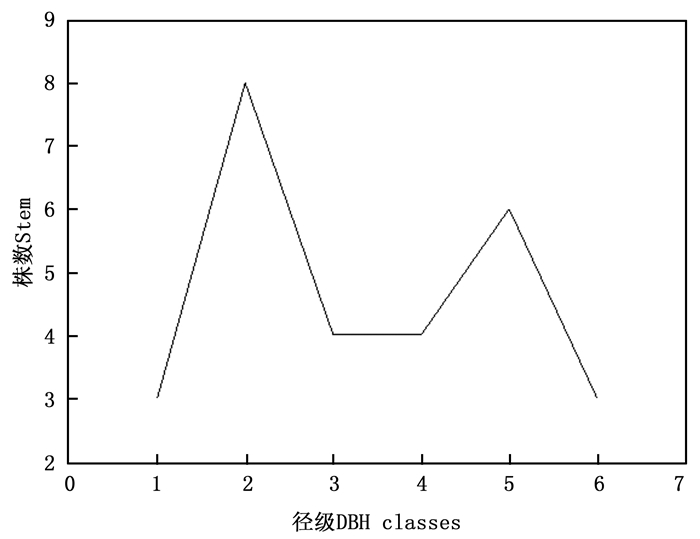
按照上限排外法, 以0.3 cm为基径径阶长度, 分析各径级藤枣种群数量(图 2)。除第二径级(0.3 cm≤基径 < 0.6 cm)藤枣种群数量(8株)及第五径级(1.2 cm≤基径 < 1.5 cm)藤枣种群数量(6株)较高外, 其余各径级均为3株或4株, 种群表现出稳定状态。

图 2 不同径级藤枣株数
Figure 2. The stems of Eleutharrhena macrocarpa in different size classes
-
幼苗的划分标准, 高度H < 33 cm的即为幼苗[13]。本次调查中, 共发现3株藤枣长度小于33 cm, 占藤枣总株数的10.71%。其长度分别为10 cm、15 cm和30 cm, 平均长度为18.33 cm, 其基径分别为0.1 cm、0.1 cm和0.2 cm, 平均为0.13 cm。
-
本文的研究结果显示, 藤枣生境地中林分的平均高、平均胸径均较高, 而林分密度偏低。同时, 群落中分布有高度大于30 m、胸径大于100 cm的物种与个体, 结合太阳河省级自然保护区对该区植被的记载, 本研究中藤枣生境地的群落具有一定的古老特性, 显示出老龄林特征。而不同高度级和径级内物种丰富度和个体多度的分布特征(物种和个体主要出现在中等径级和高度级内)则显示出群落稳定性的一面。侯昭强等[6]对藤枣生境地调查也显示, 藤枣生境地群落结构复杂, 乔木上层平均高35 m, 为原始热带雨林。结合本文与侯昭强等[6]的研究结果, 藤枣分布地群落应为稳定状态的老龄林或原始林。本文同时对所调查5块样地的物种组成进行了分析, 根据重要值前5位物种特性结合样地中的物种组成分析发现, 5块样地植被类型分别属于热带雨林(样地1、2、3、5)和季风常绿阔叶林(样地4)。而此前的报道则显示, 藤枣分布地植被类型为热带雨林(沟谷雨林、季节雨林、山地雨林)[6]。季风常绿阔叶林中藤枣的发现则拓展了藤枣分布地的植被类型, 这也警示作者, 在今后的藤枣调查过程中, 不仅要关注热带雨林, 还要对季风常绿阔叶林开展调查。
-
环境是植物生活的载体, 与植物有着密切的关系[14-15]。环境决定着物种进化的趋势及种群数量[16]。本研究中, 藤枣生境地海拔高度在964~1 250 m, 平均为1 107 m, 处于《云南植物志》和《中国植物志》记载范围内, 也处于侯昭强等[6]研究范围的上限。综合藤枣海拔分布现有研究结果发现, 藤枣的海拔分布处于300~1 500 m范围[6, 17], 该海拔范围植被类型主要包括热带雨林和季风常绿阔叶林, 说明藤枣分布地的植被类型主要为热带雨林和季风常绿阔叶林, 这也在本文的研究结果中得到证实。藤枣分布地的地形主要为陡坡, 阴坡及坡下位置为藤枣主要分布区, 这与侯昭强等[6]的调查结果相同。藤枣分布地林分郁闭度为76%, 同时由于其处于阴坡及坡下位置, 林内光照相对较弱。野外调查中也发现, 藤枣分布地接受光照直射的时间非常短暂(2~3 h), 林内及林冠以接收散射光为主。与此同时, 藤枣分布地距离河(溪)流距离较近为5~70 m, 平均距离仅为31m。距离河(溪)流较近保证了林内较高的湿度。这与“藤枣喜湿热的气候, 常生长在海拔较低的山坡林下阴湿处、沟谷溪边等生境[6]”的研究结果相一致。此外, 酸碱度接近中性、养分条件较好的土壤是藤枣生境中的重要特点之一。由于藤枣分布地为坡下位置, 一方面地表以下水分的从高向低流动带来一定量的养分, 另一方面地表较厚的枯落物分解进一步增加了土壤的养分。因此, 藤枣分布地中土壤养分条件较好。
-
种群数量是种群生态学的核心, 表征了种群个体数量的分配状况、形成过程和发展趋势, 能够揭示种群的形成机制和维持机制[18]。本研究共发现藤枣28株, 这是目前在同一地点通过实地考察发现的最多株数。侯昭强等[6]通过多年对滇西、滇西南、滇南及滇东南的考察共发现藤枣约40株, 其中江城16株为最多, 其余地点均在1~6株之间。截至目前, 藤枣种群的数量众说纷纭, 没有明确的数量大小。藤枣辨识难度高是其种群数量不定的重要原因之一。物种辨识需要专业的知识及丰富的野外经验, 其依据多为叶片、花、果实等器官的特征。然而, 野外调查过程中, 花和果实极难见到, 叶片特征成为其常用的辨识依据。但该种手段对于一些叶片较容易采摘或观察到的物种适用性高, 而对于叶片较少又不容易采摘或观察到的物种则用途有限。藤枣作为多年生木质藤本, 和其它大型木质藤本一样, 成年个体均攀爬到树冠上层。由于其生境地的植被(热带雨林和季风常绿阔叶林)个体均较高大, 树冠一般超过30 m, 加之树冠叶片相互遮挡, 在林下极难观察到或清晰发现藤枣叶片特征, 从而导致种类无法辨识。此外, 野外调查不全面及难度高也是藤枣种群数量不定的另一重要原因。由于藤枣分布区包括滇西、滇西南、滇南、滇东南及广西西南部等地区, 对其可能的分布地点进行全面仔细的普查所需人力、物力、财力要求均较高。同时, 由于其分布地地形均为陡坡, 又邻近沟底, 湿滑及难于攀爬进一步增大了其调查的难度。因此, 要想准确定量藤枣种群数量难度极高。
植物的天然更新是物种自我维持的重要机制, 是种群延续和群落稳定的重要保证[19], 而幼苗数量则决定着物种更新成败。依据现有乔木更新幼苗大小的界定标准, 本研究共发现3株长度小于33 cm的藤枣幼苗, 占总株数的10.71%, 数量较少。然而, 本次调查的数据显示, 长度在0.33 m~1.3 m范围内有6株, 而长度大于1.3 m的19株个体其基径(0.4~1.7 cm)和胸径(0.2~1.3 cm)也均较小, 这表明, 本次调查中所发现的藤枣多数处于其个体发育的前期阶段, 这也在后期成年个体的发现及种子发芽试验中得到部分证实(未出版数据)。由于不同物种其生物学特性的差异, 以及藤本和乔木生长型的差异, 藤枣幼苗的界定采用乔木幼苗界定方法是否科学有待进一步探讨, 而其更新状况也因幼苗界定方法的不同而产生不同的结果。因此, 加强藤枣生物学特性研究是深入探索其种群动态的前期基础, 并能更好的揭示其濒危机制。
-
通过藤枣生境地特征分析表明, 藤枣分布地植被类型除热带雨林外, 季风常绿阔叶林也是其分布的植被类型之一, 这为今后全面调查藤枣资源现状拓展了调查范围。藤枣分布地植物群落树体粗壮、高大, 大径级和高大树木的存在显示出群落的古老性; 同时, 物种和个体多度在不同高度级和径级内的分布特征则显示出群落稳定性的一面。因此, 藤枣分布地群落为稳定状态的老龄林或原始林。藤枣分布地环境以阴坡、坡下、荫蔽为主要特点, 距河(溪)流较近, 空气湿度大, 同时, 土壤养分条件较好。本研究发现的28株藤枣是目前在同一地点通过实地考察发现的最多株数。藤枣资源数量由于其辨识方法单一、调查不全面及难度高而无法准确确定, 因此, 藤枣种类的辨识和可能分布区域的全面调查是当前急需开展的工作。28株藤枣分布于19个样方, 多数为单方单株分布。藤枣幼苗数量较少, 自然更新受限。
濒危植物藤枣的生境与种群结构特征
Habitat and Population Structure Characteristics ofEleutharrhena macrocarpa
-
摘要:
目的 探索藤枣生境及其种群数量、分布、结构及更新等特征。 方法 在藤枣主要分布地设置5块20 m×20 m的调查样地,记录样地中所有胸径(DBH)≥1 cm的植物的物种名称、高度、胸径、及样地的郁闭度、海拔、坡度、裸岩覆盖率、距河流距离、土壤等环境因子,同时在样地中调查藤枣种群特征,分析藤枣藤枣生境地的林分因子、群落特征、生境特征及种群数量与更新。 结果 表明:藤枣生境地林分平均高、平均胸径、平均胸高断面积、林分密度分别为11.98 m、15.13 cm、41.42 m2·hm-2和1 205株·hm-2。群落平均物种丰富度为35.2,群落类型为热带雨林和季风常绿阔叶林;在大小级结构上,不同高度级和径级内物种丰富度和个体多度均呈单峰曲线,最高值均出现在第Ⅱ高度级和径级内,但第Ⅴ(H≥30 m)高度级和第Ⅵ(DBH≥100 cm)径级均有物种和个体分布。藤枣生境地平均海拔高度为1 107 m,坡度为陡坡,坡向则多数为阴坡,处坡下位置;林分郁闭度仅为76%,但距河流较近(31 m),并伴有一定的裸露岩石;土壤接近中性(pH=6.2),养分条件较好。在5块调查样地(面积20 m×20 m)中,共有藤枣28株,分布于19个样方,多数为单方单株分布。28株藤枣平均长度2.39 m,平均基径仅为0.74 cm,平均胸径为0.41 cm。其中,更新幼苗(H < 33 cm)为3株,占藤枣总株数的10.71%。 结论 藤枣分布地植物群落类型主要为热带雨林和季风常绿阔叶林,树木较高,树体较大,单峰曲线是其物种丰富度和个体多度在不同高度级和径级内的分布形式。生境地环境以高温高湿为主,土壤养分条件较好。28株藤枣分布于19个样方,多数为单方单株分布。藤枣幼苗数量较少,自然更新受限。 Abstract:Objective To study the habitat and population structure ofEleutharrhena macrocarpa, an endangered plant species in China. Method The stand parameters, community characteristics, habitat characteristics, population and regeneration of Eleutharrhena macrocarpa were investigated on five plots (20 m×20 m). On each plot, the species, height, and DBH (diameter at breast height) of all individuals with the height (or length) ≥1.3 m were recorded. Result The mean height, mean DBH, basal areas, and density of E. macrocarpa community were 11.98 m, 15.13 cm, 41.42 m2·hm-2, and 1 205 stem·hm-2, respectively. The species richness was 35.2 and the community types were tropical rainforest and monsoon evergreen broad-leaved forest. The species richness and abundance showed a single peak curve with DBH and height increasing. The maximum values of species richness and abundance all appeared in the second DBH and height classes. In the same time, it was also found that the species and stems distributed in the fifth height classes and the sixth DBH classes. The E. macrocarpa mainly distributed at 1 107 m m a.s.l., abrupt slope, shady slope and the bottom part of slope. The canopy density was only 76% and some naked rocks and the neutral and rich nutrient soil existed in the habitat of E. macrocarpa. The position of E. macrocarpa habitat was very near to the river (only 31 m). Twenty-eight stems were found in five plots and distributed in nineteen subplots, only one stem existed in most of subplots. The mean length, mean basal diameter, mean DBH were 2.39 m, 0.74 cm, and 0.41 cm, respectively. Only three seedlings, about 10.71% of total E. macrocarpa stems, were found in all plots. Conclusion The community types are tropical rainforest and monsoon evergreen broad-leaved forest in the habitat of E. macrocarpa, which have the higher and bigger trees. Single peak curve is the mainly distribution type of species richness and abundance among different DBH and height classes. The environment is characterized by high temperature and high humidity. The soil nutrient is also high. Twenty-eight stems of E. macrocarpa distributed in nineteen subplots, one stem in one subplot is the mainly distribution type. The amount of E. macrocarpa seedling is low and regeneration is limited. -
图 2 不同径级藤枣株数
Figure 2. The stems of Eleutharrhena macrocarpa in different size classes
表 1 不同样地林分因子
Table 1. Stand factors in different plots
样地号
Plots平均高
Mean height/ m平均胸径
Mean DBH/cm胸高断面积
Basal areas/(m2 · hm-2)密度
Density/(stem · hm-2)1 10.93 15.14 33.61 1 125 2 11.54 15.34 27.68 925 3 13.37 18.14 67.90 1 475 4 9.78 9.88 18.99 1 625 5 14.27 17.13 58.91 875 平均Mean 11.98 15.13 41.42 1 205  下载: 导出CSV
下载: 导出CSV
表 2 不同样地物种丰富度
Table 2. Species richness in different plots
样地号
PlotsDBH≥1 cm 灌草
Shrub and herb总物种数
Total species1 24 12 35 2 15 22 31 3 26 24 35 4 28 29 42 5 16 24 33 平均Mean 218 22.2 35.2
下载: 导出CSV
表 3 不同样地重要值前5位的物种
Table 3. The top five species of the important values in different plots
样地号
Plot物种
Species相对盖度
Relative coverage相对多度
Relative abundance相对频度
Relative frequency重要值
Important value1 棒柄花(Cleidion brevipetiolatum Pax et Hoffm.) 0.067 1 0.2093 0.2105 0.1623 1 线毛番龙眼(Pometia tomentosa (Bl.) Teysm. et Binn.) 0.108 6 0.0698 0.0789 0.0858 1 香桂(Cinnamomum subavenium Miq.) 0.1715 0.0233 0.0263 0.0737 1 越南安息香(Styrax tonkinensis (Pierre) Craib ex Hartw.) 0.1628 0.0233 0.0263 0.0708 1 大叶桂樱(Laurocerasus zippeliana (Miq.) Yü et Lu) 0.0700 0.0698 0.0526 0.0641 2 歪叶榕(Ficus cyrtophylla Wall, ex Miq.) 0.2417 0.2973 0.2727 0.2706 2 景洪暗罗(Polyalthia cheliensis Hu) 0.0986 0.1622 0.1515 0.1374 2 假桂钓樟(Linderca tonkinensis Lec.) 0.2091 0.0270 0.0303 0.0888 2 秋枫(Bischofia javanica Bl.) 0.1085 0.0541 0.0606 0.0744 2 木奶果(Baccaurea ramiflora Lour.) 0.0114 0.1081 0.0909 0.0701 3 秋枫(Bischofia javanica Bl.) 0.3522 0.1017 0.0435 0.1658 3 线毛番龙眼(Pometia tomentosa (Bl.) Teysm. et Binn.) 0.0724 0.1695 0.1522 0.1314 3 顶果树(Acrocarpus fraxinifolius Wight ex Arn.) 0.2834 0.0169 0.0217 0.1074 3 棒柄花(Cleidion brevipetiolatum Pax et Hoffm.) 0.0204 0.1017 0.1087 0.0769 3 歪叶榕(Ficus cyrtophylla Wall, ex Miq.) 0.0554 0.0508 0.0652 0.0572 4 耳叶柯(Lithocarpus grandifolius (D. Don) Biswas) 0.2460 0.1077 0.1017 0.1518 4 牛矢果(Osmanthus matsumuranus Hayata) 0.0408 0.1077 0.1017 0.0834 4 鱼尾葵(Caryota ochlandra Hance) 0.1362 0.0615 0.0508 0.0828 4 密花树(Rapanea nerifolia (Sieb. et Zucc.) Mez) 0.0356 0.0769 0.0678 0.0601 4 大肉实树(Sarcosperma arboreum Hook. f.) 0.0254 0.0615 0.0678 0.0516 5 千果榄仁(Terminalia myriocarpa Vaniot Huerck et Muell. -Arg.) 0.7216 0.0571 0.0667 0.2818 5 香芙木(Schoepfia fragrans Wall.) 0.0488 0.2571 0.2000 0.1686 5 菜阳河杮(Dispyros caiyangheiensis Tao) 0.0211 0.2000 0.1667 0.1293 5 山牡荆(Vitex quinata (Lour.) Wall.) 0.0633 0.0571 0.0667 0.0624 5 香桂(Cinnamomum subavenium Miq.) 0.037 2 0.0571 0.0667 0.0537
下载: 导出CSV
表 4 藤枣生境地环境特征
Table 4. Environment characteristic in the habitat of Eleutharrhena macrocarpa
环境因子Environmental factors 1 2 3 4 5 平均值Mean 海拔Altitude /m 1 052 1 032 964 1 237 1 250 1 107 坡度Slope /。 22 31 31 22 27 26.6 坡向Slope aspect 东坡 北坡 北坡 南坡 北坡 - 坡位Slope position 坡下 坡下 坡下 坡下 坡下 坡下 郁闭度Canopy density 80 70 85 80 65 76 距河流距离Distance to rivers /m 5 70 45 25 10 31 裸岩覆盖率Rocky’s cover /% 5 15 10 8 18 11.2 pH 6.61 6.49 6.52 5.97 5.43 6.20 有机质Organic matter/ (g · kg -1) 120.48 108.54 105.38 95.34 50.24 96.00 全氮Nitrogen/% 0.47 0.43 0.42 0.42 0.25 0.40 全磷Phosphorus/% 0.19 0.15 0.16 0.07 0.08 0.13 全钾Potassiunm% 1.95 2.19 2.26 1.07 0.7 1.67 水解性氮Hydrolysable nitrogen/(mg · kg-1) 359.90 337.50 335.50 372.10 275.51 336.10 有效磷Available phosphorus/(mg · kg-1) 89.65 54.89 57.76 19.38 55.92 55.52 速效钾Available potassium (mg · kg -1) 340.10 230.45 228.00 218.20 247.15 252.78
下载: 导出CSV
表 5 藤枣种群数量
Table 5. Population numbers of Eleutharrhena macrocarpa
样地号
Plots个体多度
Abundance平均长度
Mean length/ m平均基径
Mean basal diameter /cm平均胸径
Mean DBH /cm1 1 1.2 0.45 - 2 7 2.38 1.04 0.38 3 10 2.74 0.60 0.33 4 9 4.03 1.02 0.73 5 1 1.6 0.58 0.2 平均Mean 5.6 2.39 0.74 0.41
下载: 导出CSV
-
[1] 颜忠诚, 陈永林. 动物的生境选择[J]. 生态学杂志, 1998, 17(2): 43-49. doi: 10.3321/j.issn:1000-4890.1998.02.009 [2] 李世森, 王宇, 邵晨, 等. 虎纹蛙(Hoplobatrachus chinensis)繁殖后期的生境特征及生境选择[J]. 生态学杂志, 2014, 33(11): 3018-3025. [3] Cody M L. Habitat selection and interspecific territoriality among the Sylviid warblers of England and Sweden[J]. Ecological Monographs, 1978, 48(4): 351-396. doi: 10.2307/2937239 [4] 王朋, 孟凡露, 曾治, 等. 大兴安岭林区驯鹿夏季喜栖生境的生态特征[J]. 应用生态学报, 2014, 25(9): 2529-2535. [5] 卢杰, 唐晓琴, 李连强, 等. 色季拉山长鞭红景天种群生境特征[J]. 西部林业科学, 2016, 45(3): 13-19. [6] 侯昭强, 周丹, 侯淑娜, 等. 中国藤枣现状研究[J]. 植物分类与资源学报, 2015, 37(5): 640-646. [7] Ferguson I K. Pollen Morphology of the Tribe Triclisieae of the Menispermaceae in Relation to Its Taxonomy[J]. Kew Bulletin, 1975, 30(5): 49-75. [8] Cutler D F. Anatomical Notes on the Leaf of Eleutharrhena and Pycnarrhena(Menispermaceae)[J]. Kew Bulletin, 1975, 30(1): 41-48. doi: 10.2307/4102874 [9] 王凤春. 气孔群[J]. 植物杂志, 1984, 11(2): 22. [10] 洪亚平, 潘开玉, 陈之端, 等. 防己科植物的叶表皮特征及其系统学意义[J]. 植物学报, 2001, 43(6): 615-623. doi: 10.3321/j.issn:1672-9072.2001.06.011 [11] 刘万德, 苏建荣, 李帅锋, 等. 云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征[J]. 植物生态学报, 2015, 39(1): 52-62. [12] 曹善寿. 菜阳河自然保护区[M]. 昆明: 云南科技出版社, 2003. [13] 曲仲湘. 植物生态学[M]. 北京: 高等教育出版社, 1983. [14] Blondel J. Assessing convergence at the communitywide level[J]. Trends in Ecology and Evolution, 1991, (6): 271-272. [15] Losos J B. The evolution of convergent structure in Caribbean Anolis communities[J]. Systematic Biology, 1992, 41(4): 403-420. doi: 10.1093/sysbio/41.4.403 [16] Samuels C L, Drake J A. Divergent perspectives on community convergence[J]. Trends in Ecology and Evolution, 1997, (12): 427-432. [17] 中国科学院昆明植物所. 云南植物志[M]. 北京: 科学出版社, 1979. [18] 费世民, 何亚平, 陈秀明, 等秦岭水灾迹地油松和华山松更新种群数量特征[J]. 植物生态学报, 2008, 32(1): 95-105. [19] 刘万德, 李帅锋, 张志钧, 等. 滇西北云南红豆杉群落结构与更新特征[J]. 生态学杂志, 2012, 31(12): 3024-3031. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3887
- HTML全文浏览量: 944
- PDF下载量: 672
- 被引次数: 0
