

-
钙在大多数土壤溶液中浓度为50~100 μg·g-1, 占土壤中阳离子总量的60%~80%。在自然条件下, 土壤缺钙现象一般较少出现, 但在盐基饱和度较低及酸沉降严重的土壤上容易出现[1]。目前, 我国已成为继欧美之后的第3大酸沉降区, 且有不断恶化的趋势, 我国的酸性土壤主要分布在华中、华南和西南等15个省区, 总面积达218 km2, 约占全国土地总面积的22.7%, 其土壤类型包括黄壤、黄棕壤、红壤、砖红壤和赤红壤等[2]。酸性土壤中阳离子交换量较低, Ca2+作为盐基成分的主导离子容易大量淋失, 导致中国南方的赤红壤、砖红壤等酸性土壤缺钙现象严重[3]。当土壤的pH值降至4左右时, 土壤中的Ca2+、Mg2+等阳离子几乎消耗殆尽[4]。钙是植物生长发育的必需元素之一, 具有维持细胞壁、细胞膜及膜结合蛋白的稳定性, 对细胞内各种生长发育具有调控作用, 是耦联胞外信号与胞内生理生化反应的第二信使[5]。当土壤缺钙时, 植物体内Ca2+库平衡被破坏, 细胞的分裂与形成受到影响, 细胞壁不能形成, 对种子和根系的发育、根尖和茎尖分生组织的生长有较大影响, 同时还对植物的伸长生长和木质坚固也有影响, 植株容易产生缺钙现象[6]。
马尾松(Pinus massoniana Lamb.)作为中国南方的主要造林树种, 占中国用材林面积的17.6%, 具有较高的经济效益、社会效益和生态效益, 为避免马尾松林生产力下降, 实现马尾松人工林可持续经营已成为当前的一项迫切任务。然而, 长期种植马尾松会造成林地水土流失加重, 土壤养分如Ca、Mg和K等流失加快, 引起林地养分缺乏, 导致马尾松生产力降低[7-8]; 而周期性采伐使大量养分元素随木材及其他器官的收获而被移出, 造成林地养分净消耗, 影响了钙在生态系统中的循环, 致使钙流失, 从而导致地力衰退[9-12]。此外, 酸雨作用也会导致林地土壤酸化, 使土壤中盐基离子淋失, 特别是Ca2+的流失[13-14]。研究结果表明, 在马尾松林中土壤钙含量降低可能是导致马尾松林分衰退及生产力降低的重要原因之一。
前人对黔中地区马尾松生长的影响因子进行研究, 结果表明, 马尾松幼树的株高生长与土壤中CaO、Fe2O3含量及pH值呈负相关, 土壤中钙含量对马尾松生长发育影响较大[15]。目前, 专门针对钙与马尾松相关的研究主要集中在养分含量分析[16-18]及种源[19]等方面, 而有关不同Ca2+水平对马尾松生长及生理特征的影响尚不清楚。本文以半年生马尾松幼苗为研究对象, 用不同Ca2+水平的Hoagland营养液对马尾松幼苗进行砂培, 研究其生长情况及针叶生理指标的变化特征, 掌握马尾松适宜生长的钙浓度范围, 阐明不同供Ca2+水平下马尾松生理指标的变化规律及响应机理, 为不同Ca2+水平下马尾松人工林培育及合理施用钙肥提供参考。
-
本试验于2015年3月至8月在贵州大学林学院温室内采用营养液砂培试验。石英砂的处理参照宫杰芳等[20]的方法, 然后装入聚乙烯塑料盆(30 cm×30 cm×40 cm, 底部带绿豆大小的排水孔8个), 每盆10 kg。选取同一规格的半年生马尾松苗(苗高10~11 cm)移植于盆内, 每盆3株。试验共设6个处理, 每处理10盆重复。在移植马尾松苗1周后开始浇营养液, 每3 d浇1次(250 mL·盆-1), 并于第3次浇营养液前一天用大量蒸馏水充分淋洗石英砂以防盐分积累, 之后均按此法浇营养液直至试验结束。本试验根据毛达如[21]的方法采用Hoagland营养液, 其成分如下: 5.0 mmol·L-1 KNO3, 2.0 mmol·L-1 MgSO4·7H2O, 0.5 mmol·L-1 KH2PO4, 46 μmol·L-1 H3BO3, 10 μmol·L-1 MnCl2·4H2O, 0.8 μmol·L-1 ZnSO4·7H2O, 0.56 μmol·L-1 CuSO4·5H2O, 0.4 μmol·L-1 H2MoO4·4H2O和25 μmol·L-1 Na2Fe-EDTA。试验以标准Hoagland营养液(4.0 mmol·L-1 Ca2+)为对照, 6个供Ca2+水平分别为0.0、0.4、1.0、2.0、3.0、4.0 mmol·L-1(CK), 其中, Ca2+以Ca(NO3)2形式添加, 而不足的NO3- 以NaNO3形式添加, 并用稀盐酸或氢氧化钠调节溶液pH值至5.5。分别于幼苗移植后第20、40、60、80天, 随机选取6株马尾松幼苗东、南、西、北向健康针叶(功能叶)进行生理指标测定。于3月(幼苗移植初)和8月分别测量各处理的株高和地径。
-
参照汤绍虎等[22]的方法测定功能叶的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性以及丙二醛(MDA)和脯氨酸含量, 参照张亚宏等[23]的方法测定抗坏血酸过氧化物酶(APX)活性, 参照汤章城[24]和彭建等[25]的方法测定活性氧(ROS)中氧阴离子自由基(O2-)产生速率和过氧化氢(H2O2)浓度, 参照Queval等[26]的方法测定抗坏血酸(AsA)及谷胱甘肽(GSH)的含量, 参照李合生等[27]的方法测定类胡萝卜素的含量。
-
试验数据为平均值±标准差(3次重复), 使用Excel进行数据统计与作图, SPSS(V20.0)进行数据分析。采用隶属函数法对马尾松生理指标进行综合评定; 采用80 d时各处理的生理和生长指标进行相关性分析。
-
从表 1可看出: 随着供Ca2+水平的增加, 马尾松幼苗株高增量和地径增量均呈先增加后降低的趋势, 其中, 供Ca2+水平为2.0 mmol·L-1的株高增量和地径增量均最大, 分别比CK高51.84%和47.78%, 而供Ca2+水平为4.0 mmol·L-1(CK)和0.0 mmol·L-1的株高增量和地径增量最小。供Ca2+水平为2.0 mmol·L-1的株高增量和地径增量除与1.0 mmol·L-1处理的差异不显著外, 均显著高于其他处理。
表 1 不同供Ca2+水平下马尾松的生长情况
Table 1. Growth of P. massoniana seedling under different Ca2+ concentrations
Ca2+浓度
Ca2+ concentration/(mmol·L-1)株高增量
Height increment /cm地径增量
Stem increment/mm0.0 8.72±1.12bc 0.91±0.02c 0.4 9.96±0.45bc 1.09±0.16c 1.0 11.02±1.12ab 1.47±0.19ab 2.0 12.77±0.51a 1.67±0.08a 3.0 10.08±0.53bc 1.20±0.04bc 4.0(CK) 8.41±0.54c 1.13±0.07bc 注: 同列不同小写字母表示差异显著(P < 0.05), 下同。
Note: Data with different lowercase letters in the same column are highly significantly different(P < 0. 05). The same below. -
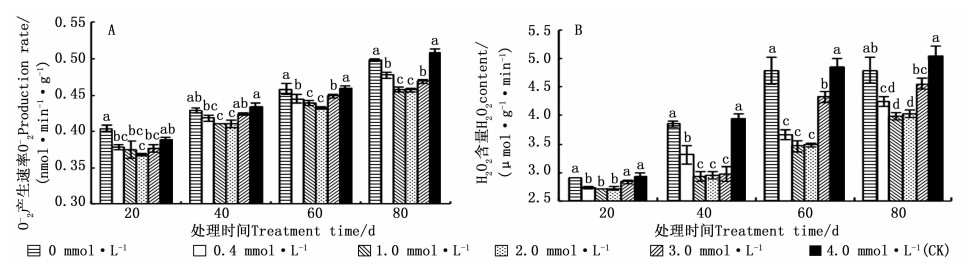
图 1 A表明: 不同供Ca2+水平下, 马尾松在相同的生长期内O2-产生速率均呈先降低后升高的趋势。处理20、40、60、80 d时, 针叶中O2-产生速率均在2.0 mmol·L-1供Ca2+水平时最低, 比CK分别降低了5.12%、5.33%、5.92%和10.02%。处理20 d时, O2-产生速率在0 mmol·L-1供Ca2+水平时最高, 且比CK高4.26%;而处理40、60、80 d时, O2-产生速率均在4.0 mmol·L-1供Ca2+水平时最高。相同供Ca2+水平的马尾松针叶内的O2-产生速率随生长期的延长呈不断上升趋势。

图 1 不同供Ca2+水平下马尾松针叶O2-产生速率和H2O2含量的变化
Figure 1. Change of O2- generation rate and H2O2 content of P. massoniana seedling under different Ca2+ concentrations
-
图 1 B表明: 不同供Ca2+水平下, 马尾松幼苗针叶内的H2O2浓度随生长期的延长呈不断增加的趋势。处理20、40、60、80 d时, 针叶内的H2O2浓度均在1.0 mmol·L-1供Ca2+水平时最低, 分别比对照低7.63%、25.75%、28.72%、20.88%;均在4.0 mmol·L-1供Ca2+水平时最高, 而0.0 mmol·L-1供Ca2+水平的次之, 仅比4.0 mmol·L-1(CK)处理依次低0.67%、2.48%、1.55%和4.87%。
-
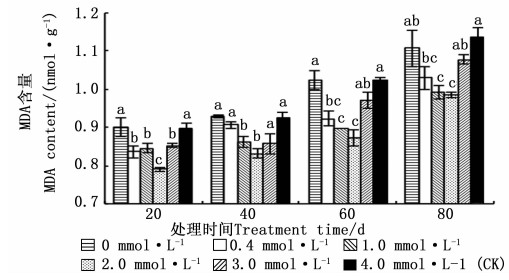
从图 2看出: 在相同的生长期内, 马尾松幼苗针叶内的MDA含量随供Ca2+水平的增加均呈先降低后增加的趋势。处理20、40、60、80 d时, MDA含量均在2.0 mmol·L-1供Ca2+水平最低, 分别比4.0 mmol·L-1(CK)下降了11.98%、10.25%、14.69%、13.25%;除处理80 d外, MDA含量的最高值均在0.0 mmol·L-1供Ca2+水平, 分别比4.0 mmol·L-1(CK)增加了0.31%、0.14%、0.20%, 而处理80 d时, MDA含量最高值在4.0 mmol·L-1(CK)供Ca2+水平。相同供Ca2+水平的马尾松幼苗针叶中的MDA含量随着生长期的延长呈上升趋势。

图 2 不同供Ca2+水平下马尾松针叶MDA含量的变化
Figure 2. Change of MDA content of P. massoniana seedling under different Ca2+ concentrations
-
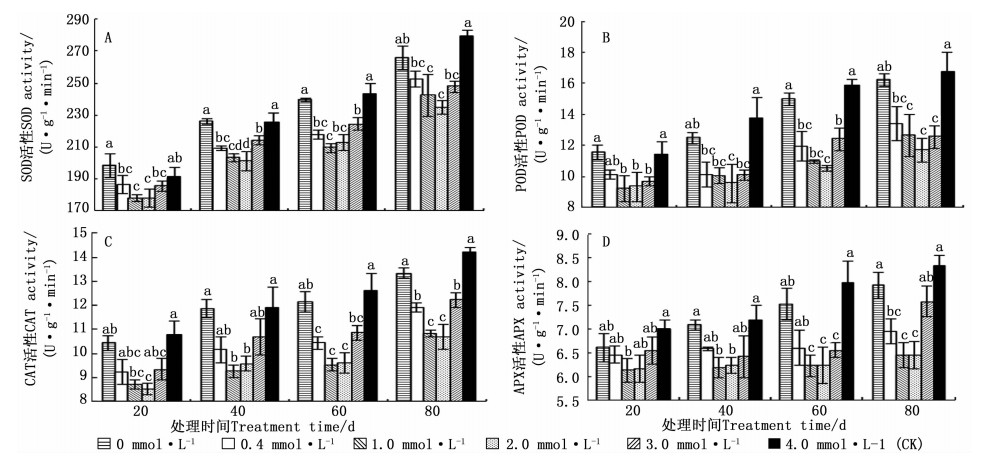
不同供Ca2+水平的马尾松幼苗, 在相同生长期内, 马尾松针叶中的SOD、POD、CAT和APX活性随供Ca2+水平的增加先降低后增加; 而相同供Ca2+水平下, 马尾松针叶中的SOD、POD、CAT和APX活性随生长期的延长呈不断上升的趋势(图 3A~D)。0.0、4.0 mmol·L-1 Ca2+处理的马尾松针叶内的抗氧化酶活性在不同取样期内均处于相对较高的水平, 而2.0、1.0 mmol·L-1Ca2+处理的则处于相对较低水平。马尾松幼苗针叶的SOD、POD、CAT、APX活性最低值, 在处理20 d时, 分别出现在2.0、1.0、2.0、1.0 mmol·L-1 Ca2+处理中, 分别比对照低7.23%、19.20%、20.83%、12.46%;处理40 d时, 分别出现在2.0、2.0、1.0、1.0 mmol·L-1Ca2+处理中, 分别比对照低10.86%、30.30%、22.16%、13.84%;处理60 d时, 分别出现在1.0、2.0、1.0、1.0 mmol·L-1Ca2+处理中, 分别比对照低13.88%、33.49%、24.37%、21.97%;处理80 d时, 分别出现在2.0、2.0、2.0、1.0 mmol·L-1Ca2+处理中, 分别比对照低15.93%、30.07%、24.69%、22.78%。

图 3 不同供Ca2+水平下马尾松SOD、POD、CAT和APX活性的变化
Figure 3. Change of SOD, POD, CAT and APX activities of P. massoniana seedling under different Ca2+ concentrations
-
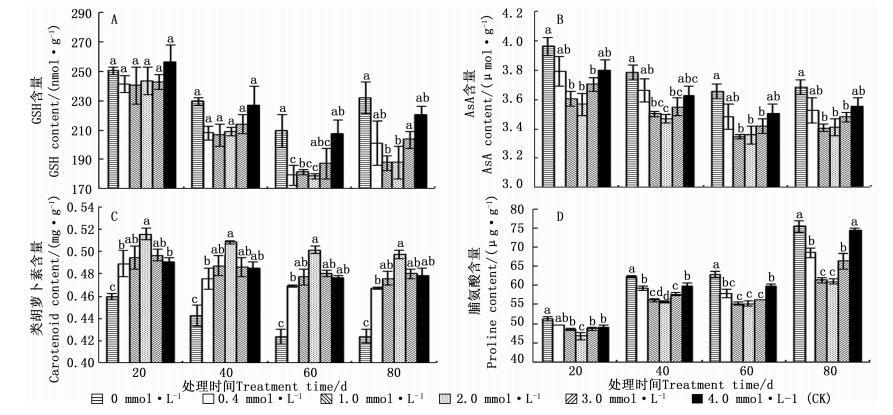
图 4A~C表明: 相同生长期内, 随着供Ca2+水平的增加, 马尾松针叶内的AsA和GSH含量先降低后升高, 类胡萝卜素含量变化则先升高后降低; 而在不同生长期内, 相同Ca2+水平的马尾松幼苗针叶内的AsA、GSH和类胡萝卜素含量变化存在一定差异。在不同生长期内, 马尾松针叶内的AsA含量在0.0 mmol·L-1 Ca2+处理的均最高, 在2.0或1.0 mmol·L-1 Ca2+处理的均最低; 类胡萝卜素含量在2.0 mmol·L-1 Ca2+处理的均最高, 在0.0 mmol·L-1 Ca2+处理的均最低; GSH含量最高值除20 d时出现在4.0 mmol·L-1(CK)Ca2+处理外, 在40、60、80 d时均出现在0 mmol·L-1 Ca2+处理中, 而最低值均出现在1.0或2.0 mmol·L-1 Ca2+处理中。

图 4 不同供Ca2+水平下马尾松AsA、GSH、类胡萝卜素和脯氨酸含量的变化
Figure 4. Change of AsA, GSH, carotenoids and proline contents of P.massoniana seedling under different Ca2+ concentrations
-
在相同生长期内, 马尾松幼苗针叶中的脯氨酸含量均随供Ca2+水平的增加先降低后增加, 最小值均出现在2.0 mmol·L-1 Ca2+处理中, 在20、40、60、80 d时, 分别比4.0 mmol·L-1(CK)降低了5.01%、7.16%、7.92%、18.09%;马尾松幼苗针叶中的脯氨酸含量最大值均出现在0.0 mmol·L-1 Ca2+处理中, 在20、40、60、80 d时, 分别比4.0 mmol·L-1(CK)升高了4.21%、3.90%、4.73%、1.23%。相同供Ca2+水平下, 马尾松幼苗针叶内的脯氨酸含量随处理时间的延长总体呈上升趋势, 而对照80 d时的脯氨酸含量比60 d时的增加24.27%(图 4D)。
-
通过对马尾松幼苗的株高净增量和地径净增量及针叶内各生理指标间进行相关性分析(表 2)可知: 马尾松株高增量与针叶内的SOD活性呈显著负相关(P<0.05), 而与O2-产生速率、H2O2浓度、MDA含量、POD活性、CAT活性、APX活性、AsA含量、GSH含量、脯氨酸含量均呈极显著负相关(P<0.01);马尾松地径净增量与H2O2浓度、MDA含量、AsA含量和脯氨酸含量均呈显著负相关(P<0.05), 而与O2-产生速率、SOD活性、POD活性、CAT活性、APX活性和GSH含量均呈极显著负相关(P<0.01);类胡萝卜素含量与株高增量和地径增量关系均不显著。
表 2 马尾松幼苗生理指标与生长的相关性分析
Table 2. Correlation analysis between physiological indexes and growth indexes of P.massoniana seedling
指标
Index株高增量
Height increment地径增量
Stem incrementO2-产生速率O2- generation rate/ (nmol·min-1·g-1) -0.746** -0.669** H2O2浓度H2O2 content/(μmol·g-1·min-1) -0.609** -0.558* MDA含量MDA content/(nmol·g-1) -0.751** -0.533* SOD活性SOD activity/(U·g-1·min-1) -0.580* -0.651** POD活性POD activity/(U·g-1·min-1) -0.626** -0.689** CAT活性CAT activity/(U·g-1·min-1) -0.793** -0.648** APX活性APX activity/(U·g-1·min-1) -0.729** -0.590** AsA含量AsA content/(μmol·g-1) -0.622** -0.558* GSH含量GSH content/(nmol·g-1) -0.614** -0.598** 类胡萝卜素含量Carotenoid content/(mg·g-1) 0.411 10.451 脯氨酸含量Proline content/(μg·g-1) -0.615** -0.561* 注: *、** 分别表示在0.05水平(双侧)和0.01水平(双侧)上显著相关。
Note: *、** means that correlation is significant at the 0.05 level(2-tailed), 0.01 level (2-tailed), respectively.利用模糊数学的隶属函数法对生理指标进行分析, 马尾松幼苗生理指标受不同供Ca2+水平影响由强到弱的供Ca2+浓度顺序为: 4.0(CK)>0.0>3.0>0.4>1.0>2.0 mmol·L-1(表 3)。
表 3 马尾松生理指标的平均隶属函数均值
Table 3. Synthetical membership function value of P. massoniana seedling in physiological indices
Ca2+浓度
Ca2+ concentration/(mmol·L-1)隶属函数值
Subordinate function value排序
Order0.0 0.816 2 0.4 0.363 4 1.0 0.130 5 2.0 0.110 6 3.0 0.399 3 4.0(CK) 0.867 1 -
本试验中, 2.0 mmol·L-1供Ca2+水平的马尾松株高和地径增量均最高, 且与1.0 mmol·L-1供Ca2+水平差异不显著, 但与其他处理差异显著, 表明马尾松较适宜生长的供Ca2+水平为1.0~2.0 mmol·L-1。马尾松对供Ca2+水平为1.0~2.0 mmol·L-1的环境具有良好的适应性, 对低Ca2+(<1.0 mmol·L-1)环境也有一定的耐性, 但当供Ca2+水平>2.0 mmol·L-1后, 随供Ca2+水平的升高其适应性逐渐降低。马尾松在不同供Ca2+水平下, 其生理适应能力强弱的钙浓度顺序为2.0>1.0>0.4>3.0>0.0>4.0 mmol·L-1(CK), 这可能是马尾松经过长期进化和自然选择的结果。在马尾松分布区内酸沉降严重并不断恶化的情势下, 土壤中的Ca2+作为盐基成分的主导离子容易大量淋失, 土壤缺钙现象严重[3]。对于酸沉降、土壤酸化等环境问题引起的土壤Ca2+大量淋失而产生的低质低效马尾松林, 或土壤中Ca2+水平较低(<1.0 mmol·L-1)的马尾松林, 应根据林地中土壤有效钙含量情况配合施用钙肥或石灰石粉[28], 多施有机肥, 并注意防止林地土壤复酸; 当土壤Ca2+浓度较高时, 可采用增施适量的硫磺粉和硫酸亚铁等措施, 形成有利于马尾松生长和恢复的良好生态环境, 从而提高林分产量。
-
逆境条件下, 植物细胞内产生大量的ROS, 引起膜脂氧化伤害, 而抗氧化系统是植物抵御逆境伤害的重要机制[29]。本试验中, 当供Ca2+水平为1.0~2.0 mmol·L-1时, 马尾松针叶中的O2-产生速率、H2O2浓度、MDA含量、抗氧化酶活性(SOD、POD、CAT和APX)、抗氧化物质含量(GSH、AsA)和脯氨酸含量与其它处理相比均较低, 而类胡萝卜素含量相对较高, 表明供Ca2+水平为1.0~2.0 mmol·L-1时马尾松针叶内ROS的产生和清除处于动态平衡中, 确保原生质胶体及细胞组织中的各种代谢得以稳定, 防止细胞内底物与反应物接触而引起生理代谢紊乱, 因而植株生长表现较好。秦晓佳等[30]对马尾松低磷胁迫的研究表明, 生长在适宜磷浓度下, 马尾松针叶中的MDA含量、抗氧化保护酶及游离脯氨酸的含量最低, 本试验结果与该结论相似, 说明马尾松在适宜的环境中生长, 体内各代谢处于动态平衡中, 膜的稳定性较强, 植株生长旺盛。
当供Ca2+水平 < 1.0 mmol·L-1或在3.0~4.0 mmol·L-1处理下, 马尾松幼苗针叶内的抗氧化酶(SOD、POD、CAT和APX)活性、AsA和GSH含量及脯氨酸含量均比供Ca2+水平为1.0~2.0 mmol·L-1的有所升高, 表明针叶内的抗氧化系统对ROS的清除能力有所增强, 降低了膜脂过氧化水平, 在一定程度上对生物膜起到保护作用, 从而提高了马尾松对环境的适应性; 同时, 脯氨酸含量的增加对原生质胶体及细胞组织中的代谢过程具有稳定作用, 防止细胞脱水[31], 从而在一定程度上对细胞膜的相变和流动性起保护作用, 渗透调节能力得以增强, 这可能是马尾松幼苗在供Ca2+水平低于1.0 mmol·L-1或在3.0~4.0 mmol·L-1处理后一段时间内仍能继续生长的重要原因之一; 然而, O2-产生速率和H2O2浓度升高, 类胡萝卜素含量降低, 表明马尾松体内ROS产生和清除的动态平衡被打破, ROS大量积累, 而过量的ROS可直接对膜系统进行攻击, 导致膜脂过氧化, 细胞膜透性增加。此外, 过量的ROS也能引起叶绿体和线粒体的结构和功能损伤, 同时对蛋白质、核酸、脂类等生物大分子也产生氧化损伤, 导致细胞膜遭到不同程度的破坏, 从而引起MDA含量升高。因此, 本试验中, 马尾松幼苗在供Ca2+水平为0.0~1.0 mmol·L-1或在3.0~4.0 mmol·L-1处理下生长受抑, 并最终表现为马尾松幼苗株高及地径生长量降低。今后在马尾松营林过程中, 应对土壤有效钙含量进行测定, 并结合植株生长情况及生理特征进行综合分析和评价后, 科学合理施肥, 从而提高林分生产力。
-
(1) 马尾松幼苗在Ca2+水平为1.0 mmol·L-1和2.0 mmol·L-1的株高增量和地径增量较高, 其中, Ca2+水平2.0 mmol·L-1的株高增量和地径增量最高。马尾松幼苗对Ca2+水平适应能力强弱顺序为2.0>1.0>0.4>3.0>0>4.0 mmol·L-1(CK)。
(2) 在不同生长期内, 随着供Ca2+水平的增加, 马尾松针叶中ROS(O2-产生速率、H2O2浓度)、MDA含量、抗氧化酶(SOD、POD、CAT和APX)活性、抗氧化物质(GSH、AsA)含量、脯氨酸含量均先降低后增加, 最小值均出现在供Ca2+水平1.0 mmol·L-1或2.0 mmol·L-1处理中; 类胡萝卜素含量先增加后降低, 其最高值均出现在供Ca2+水平2.0 mmol·L-1处理中。
(3) 马尾松幼苗的株高增量和地径增量与类胡萝卜素含量关系不明显, 而与其他生理指标均呈显著或极显著负相关。
综上, 本文研究钙浓度对马尾松生长及生理指标的影响, 明确了适宜马尾松生长的钙浓度及生理指标的变化规律, 为马尾松人工林培育及合理施用钙肥提供一定的理论支持。
钙浓度对马尾松幼苗生长和生理特征的影响
Effects of Calcium Concentration on Growth and Physiological Characteristics of Pinus massoniana Seedling
-
摘要:
目的 在不同供Ca2+水平下对马尾松幼苗生长及生理特性进行研究,以确定适宜马尾松生长的钙浓度,为马尾松人工林培育及合理施用钙肥提供参考。 方法 以半年生马尾松幼苗为试材,采用温室内砂培,研究不同供Ca2+水平(0.0、0.4、1.0、2.0、3.0、4.0 mmol·L-1(CK))对马尾松生长及生理指标的影响。 结果 不同供Ca2+水平处理5个月后,在Ca2+水平为1.0、2.0 mmol·L-1时,马尾松幼苗的株高增量和地径增量较高,其中,2.0 mmol·L-1Ca2+水平的株高增量和地径增量最高。马尾松幼苗对Ca2+水平适应能力强弱顺序为2.0 > 1.0 > 0.4 > 3.0 > 0.0 > 4.0 mmol·L-1(CK)。在不同生长期内,随着供Ca2+水平的增加,马尾松针叶中ROS(O2-产生速率、H2O2浓度)、MDA含量、抗氧化酶(SOD、POD、CAT和APX)活性、抗氧化物质(GSH、AsA)含量、脯氨酸含量均先降低后增加,最小值均在1.0 mmol·L-1或2.0 mmol·L-1处理中;类胡萝卜素含量先增加后降低,其最高值均在2.0 mmol·L-1处理中。 结论 马尾松幼苗的株高增量和地径增量与类胡萝卜素含量关系不明显,而与其他生理指标均呈显著负相关。马尾松幼苗适宜在1.0~2.0 mmol·L-1Ca2+水平的环境中生长。在马尾松林地中,应对土壤有效钙含量进行测定,并结合植株的生长和生理特征进行综合评价后,科学合理施肥以提高生产力。 Abstract:Objective ect the optimal Ca2+ concentration for Pinus massoniana seedling growth and to provide evidence for plantation cultivation and rational application of calcium fertilization by studying the effects of Ca2+ concentration on growth and physiological characteristics of P. massoniana seedlings. Method A greenhouse sand culture experiment was carried out to study the growth and physiological characteristics of half-year-old seedlings of P. massoniana in different Ca2+ concentrations (0.0, 0.4, 1.0, 2.0, 3.0 mmol·L-1 and 4.0 mmol·L-1 (CK)). Result The results showed that both the height and stem increments of P. massoniana under 1.0-2.0 mmol·L-1 Ca2+ for five months were higher, and seedlings grew best in 2.0 mmol·L-1 Ca2+. The adaptable strong-to-weak sequence of P. massoniana seedlings was 2.0, 1.0, 0.4, 3.0, 0.0 mmol·L-1 and 4.0 mmol·L-1(CK) of Ca2+. At different growth periods, these indices of pine needles, including O2- generation rate, H2O2 content, protective enzyme activities (SOD, POD, CAT and APX), the contents of GSH, AsA and proline, decreased at first and then increased subsequently along with the increasing Ca2+ concentration, and all minimums occurred at 1.0 mmol·L-1 or 2.0 mmol·L-1 of Ca2+. Conclusion P. massoniana seedlings is suitable to grow in the soil with Ca2+ concentration of 1.0-2.0 mmol·L-1 according to the experiment results. Based on comprehensive analysis to soil available calcium of P. massoniana forestland, the growth and physiological characteristics of seedlings, it is necessary to applying rationally calcium fertilization so as to boost the productivity of P. massoniana. -
Key words:
- Pinus massoniana
- / calcium concentration
- / growth
- / physiological properties
-
图 1 不同供Ca2+水平下马尾松针叶O2-产生速率和H2O2含量的变化
Figure 1. Change of O2- generation rate and H2O2 content of P. massoniana seedling under different Ca2+ concentrations
图 2 不同供Ca2+水平下马尾松针叶MDA含量的变化
Figure 2. Change of MDA content of P. massoniana seedling under different Ca2+ concentrations
图 3 不同供Ca2+水平下马尾松SOD、POD、CAT和APX活性的变化
Figure 3. Change of SOD, POD, CAT and APX activities of P. massoniana seedling under different Ca2+ concentrations
图 4 不同供Ca2+水平下马尾松AsA、GSH、类胡萝卜素和脯氨酸含量的变化
Figure 4. Change of AsA, GSH, carotenoids and proline contents of P.massoniana seedling under different Ca2+ concentrations
表 1 不同供Ca2+水平下马尾松的生长情况
Table 1. Growth of P. massoniana seedling under different Ca2+ concentrations
Ca2+浓度
Ca2+ concentration/(mmol·L-1)株高增量
Height increment /cm地径增量
Stem increment/mm0.0 8.72±1.12bc 0.91±0.02c 0.4 9.96±0.45bc 1.09±0.16c 1.0 11.02±1.12ab 1.47±0.19ab 2.0 12.77±0.51a 1.67±0.08a 3.0 10.08±0.53bc 1.20±0.04bc 4.0(CK) 8.41±0.54c 1.13±0.07bc 注: 同列不同小写字母表示差异显著(P < 0.05), 下同。
Note: Data with different lowercase letters in the same column are highly significantly different(P < 0. 05). The same below. 下载: 导出CSV
下载: 导出CSV
表 2 马尾松幼苗生理指标与生长的相关性分析
Table 2. Correlation analysis between physiological indexes and growth indexes of P.massoniana seedling
指标
Index株高增量
Height increment地径增量
Stem incrementO2-产生速率O2- generation rate/ (nmol·min-1·g-1) -0.746** -0.669** H2O2浓度H2O2 content/(μmol·g-1·min-1) -0.609** -0.558* MDA含量MDA content/(nmol·g-1) -0.751** -0.533* SOD活性SOD activity/(U·g-1·min-1) -0.580* -0.651** POD活性POD activity/(U·g-1·min-1) -0.626** -0.689** CAT活性CAT activity/(U·g-1·min-1) -0.793** -0.648** APX活性APX activity/(U·g-1·min-1) -0.729** -0.590** AsA含量AsA content/(μmol·g-1) -0.622** -0.558* GSH含量GSH content/(nmol·g-1) -0.614** -0.598** 类胡萝卜素含量Carotenoid content/(mg·g-1) 0.411 10.451 脯氨酸含量Proline content/(μg·g-1) -0.615** -0.561* 注: *、** 分别表示在0.05水平(双侧)和0.01水平(双侧)上显著相关。
Note: *、** means that correlation is significant at the 0.05 level(2-tailed), 0.01 level (2-tailed), respectively.
下载: 导出CSV
表 3 马尾松生理指标的平均隶属函数均值
Table 3. Synthetical membership function value of P. massoniana seedling in physiological indices
Ca2+浓度
Ca2+ concentration/(mmol·L-1)隶属函数值
Subordinate function value排序
Order0.0 0.816 2 0.4 0.363 4 1.0 0.130 5 2.0 0.110 6 3.0 0.399 3 4.0(CK) 0.867 1
下载: 导出CSV
-
[1] McLaughlin S B, Wimmer R. Calcium physiology and terrestrial ecosystem processes[J]. The New Phytologist, 1999, 142(3): 373-417. doi: 10.1046/j.1469-8137.1999.00420.x [2] 赵其国. 中国东部红壤地区土壤退化的时空变化、机理及调控[M]. 北京: 科学出版社, 2002. [3] 周卫, 林葆. 土壤中钙的化学行为与生物有效性研究进展[J]. 土壤肥料, 1996(5): 19-22, 44. [4] 吴飞华, 刘廷武, 裴真明. 酸雨引起森林生态系统流失研究进展[J]. 生态学报, 2010, 30(4): 1081-1088. [5] White P J, Broadley M R. Calcium in plants[J]. Annals of Botany, 2003, 92(4): 487-511. doi: 10.1093/aob/mcg164 [6] 陈兴业, 叶林茂, 张硌. 土壤水分植物生理与肥料学[M]. 北京: 海洋出版社, 2010. [7] 唐万鹏, 潘磊, 崔鸿侠, 等. 三峡库区马尾松天然林林分结构特征分析[J]. 林业科学研究, 2015, 28(5): 681-685. doi: 10.3969/j.issn.1001-1498.2015.05.012 [8] 赵汝东, 樊剑波, 何园球, 等. 退化马尾松林下土壤障碍因子分析及酶活性研究[J]. 土壤学报, 2011, 48(6): 1287-1292. [9] Stupak I, Nordfjell T, Gundersen P. Comparing biomass and nutrient removals of stems and fresh and predried whole trees in thinnings in two Norway spruce experiments[J]. Canadian Journal of Forest Research, 2008, 38: 2660-2673. doi: 10.1139/X08-108 [10] Lattimore B, Smith C T, Titus B D, et al. Environmental factors in woodfuel production: Opportunities, risks, and criteria and indicators for sustainable practices[J]. Biomass and Bioenergy, 2009, 33(10): 1321-1342. doi: 10.1016/j.biombioe.2009.06.005 [11] Schaberg P G, DeHayes D H, Hawley G J. Anthropogenic calcium depletion: a unique threat to forest ecosystem health?[J]. Ecosystem Health, 2001, 7(4): 214-228. doi: 10.1046/j.1526-0992.2001.01046.x [12] Tomlinson G H. Acidicdeposition, nutrient leaching and forest growth[J]. Biogeochemistry, 2003, 65(1): 51-81. doi: 10.1023/A:1026069927380 [13] 凌大炯, 章家恩, 欧阳颖. 酸雨对土壤生态系统影响的研究进展[J]. 土壤, 2007, 39(4): 514-521. doi: 10.3321/j.issn:0253-9829.2007.04.004 [14] 刘菊秀, 周国逸. 土壤累积酸化对鼎湖山马尾松林物质元素迁移规律的影响[J]. 浙江大学学报: 农业与生命科学版, 2005, 31(4): 381-391. doi: 10.3321/j.issn:1008-9209.2005.04.006 [15] 周政贤. 中国马尾松[M]. 北京: 中国林业出版社, 2001. [16] 季孔庶, 邹永梅, 陈天华, 等. 不同种源马尾松灰分矿质营养变异的研究[J]. 南京林业大学学报, 1996, 20(1): 22-25. [17] 郭峰, 周运超. 不同密度马尾松林针叶养分含量及其转移特征[J]. 南京林业大学学报: 自然科学版, 2010, 34(4): 93-96. doi: 10.3969/j.issn.1000-2006.2010.04.021 [18] 周运超, 王德炉, 谌红辉. 不同营林措施对马尾松针叶养分的影响[J]. 浙江林业科技, 2001, 21(1): 5-8. doi: 10.3969/j.issn.1001-3776.2001.01.002 [19] 旷远文, 温达志, 周国逸, 等. 鼎湖山异龄马尾松针叶长度序列元素分布[J]. 植物生态学报, 2006, 30(1): 33-39. doi: 10.3321/j.issn:1005-264X.2006.01.005 [20] 宫杰芳, 周运超, 李小永, 等. 马尾松生理指标对高锰胁迫的响应[J]. 生态学杂志, 2012, 31(3): 520-525. [21] 毛达如. 植物营养研究法[M]. 北京: 中国农业大学出版社, 2005. [22] 汤绍虎, 罗充. 植物生理学实验教程[M]. 重庆: 西南师范大学出版社, 2012. [23] 张亚军, 孙万仓, 魏文慧, 等. 自交对甘蓝型油菜叶片SOD, CAT, APX活性的影响[J]. 华北农学报, 2008, 23(1): 105-108. [24] 汤章城. 现代植物生理学实验指南[M]. 北京: 科学出版社, 1999. [25] 彭建, 王丹英, 徐春梅, 等. 钼酸铵法测定水稻过氧化氢酶活性[J]. 中国农学通报, 2009, 25(16): 61-64. [26] Queval G, Issakidis-Bourguet E, Hoeberichts F A, et al. Conditional oxidative stress responses in the Arabidopsis photorespiratory mutant cat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression, and define photoperiod as a crucial factor in the regulation of H2O2-induced cell death[J]. Plant Journal, 2007, 52: 640-657. doi: 10.1111/j.1365-313X.2007.03263.x [27] 李合生, 孙群, 赵世杰, 等. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000. [28] 潘伟, 王彦辉, 于澎涛, 等. 石灰石粉混施剂量对酸雨区马尾松细根特征的影响[J]. 林业科学院研究, 2009, 22(6): 840-845. [29] Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J]. Plant Physiology and Biochemistry, 2010, 48(12): 909-930. doi: 10.1016/j.plaphy.2010.08.016 [30] 秦晓佳, 丁贵杰. 磷胁迫对不同种源马尾松氮钾吸收与利用的影响[J]. 中南林业科技大学学报, 2012, 34(4): 32-36. doi: 10.3969/j.issn.1673-923X.2012.04.007 [31] Bavei V, Shiran B, Arzani A. Evaluation of salinity tolerance in sorghum (Sorghum bicolor L. ) using ion accumulation, proline and peroxidase criteria[J]. Plant Growth Regulation, 2011, 64: 275-285. doi: 10.1007/s10725-011-9568-z -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3409
- HTML全文浏览量: 971
- PDF下载量: 614
- 被引次数: 0
