


-
沙棘(Hippophae rhamnoides)属胡颓子科(Elaeagnaceae)沙棘属,落叶灌木或小乔木,广泛分布于欧亚大陆的温带地区,在我国十多个省份均有分布,总面积超过2.133万km2 [1-2]。沙棘具有耐干旱、耐土壤贫瘠、萌芽力和萌蘖力强的特点,在我国“三北”防护林生态建设中发挥着水土保持和防风固沙的重要作用[3]。沙棘果及其枝叶富含维生素、不饱和脂肪酸、黄酮类和磷脂类等多种生物活性物质,对于一些心脑血管疾病具有一定的治疗作用[4-5]。以沙棘为主原料加工生产的一些饮料、保健品及化妆品也得到了市场的广泛认可,经济效益显著。目前,在我国北方干旱半干旱地区,沙棘已经发展成为一种重要的生态—经济型树种[3]。
沙棘绕实蝇(Rhagoletis batava obseuriosa Kol.)隶属双翅目(Diptera)实蝇科(Tephritidae)绕实蝇属,其幼虫蛀食沙棘果,导致受害沙棘果丧失其经济价值,大发生时能够造成沙棘果减产90%以上[6]。范仁俊等人报道称1987年沙棘绕实蝇在山西省右玉县大规模发生[7]。辽宁建平县罗福沟乡等地区的沙棘以及阜新县的沙棘林也曾遭受到该虫危害[8-9]。另外,在陕西北部[10]及黑龙江北部地区[11]也有该虫发生。目前,该实蝇在内蒙古西部地区乌兰布和沙漠中的人工沙棘林[12]和新疆阿勒泰、塔城部分县市的沙棘种植基地大规模爆发,给当地造成了巨大的经济损失,致使沙棘产业的发展前景受到质疑。
同绕实蝇属的其它种类如苹果绕实蝇、樱桃绕实蝇一样,沙棘绕实蝇也属于一化性昆虫,且文献记录国外仅在俄罗斯和蒙古国发生[8],因此目前尚无对沙棘绕实蝇生物学习性详细的报道。作者于2015年和2016年在内蒙古西部地区乌兰布和沙漠中的人工沙棘林选取样地,通过室内饲养结合野外观察,对沙棘绕实蝇的羽化、交尾、产卵、化蛹等生物学习性进行了研究,以期为更精准有效地监测和控制沙棘绕实蝇提供理论指导。
-
实验样地位于内蒙古磴口县中国林业科学研究院沙漠林业实验中心第三实验场,地理位置:40° 25.935′ N,106° 43.442′ E,海拔1 048~1 053 m,面积100 m × 60 m,树龄约30年,株行距2.5 m × 1 m,胸径平均6 cm,郁闭度可达1.0。
-
于2016年5月份在沙棘林地中采集沙棘绕实蝇越冬蛹,取出3 000头蛹,分别放入10个聚丙烯(PP)保鲜盒(长18.5 cm ×宽11.5 cm ×高7 cm,保鲜盒盖凿两个直径0.5 cm的透气孔,盒底凿一个16 cm × 9 cm的矩形孔,均覆以80目尼龙纱网)中,每盒放入试虫蛹300头:先在保鲜盒中铺入3 cm厚的湿润沙土,将沙棘绕实蝇蛹均匀散布其上,再覆以3 cm厚的细沙土。盖好盒盖后将保鲜盒埋于野外沙棘林中树冠投影下方,盒盖与地表平,上覆枯枝落叶。每天9:00定时打开盒盖观察成虫羽化情况。当观察到沙棘绕实蝇首次羽化后,于每日6:00以后每隔1 h观察并记录一次,直至连续10天无沙棘绕实蝇成虫羽化为止。
-
在沙棘林中,通过树冠下铺地膜以监测沙棘绕实蝇老熟幼虫钻出果实化蛹的起始日期;在实蝇化蛹盛期,于野外沙棘林中剪取受到危害的沙棘枝果(沙棘枝切口要保持湿润,防止沙棘枝叶及果实萎蔫),带回室内放入塑料盆(φ= 35 cm,H= 12 cm)中,每隔1小时观察记录一次从沙棘果中钻出的老熟幼虫数量,连续观察3天;用细毛笔轻轻将34头同一时期钻出果实的老熟幼虫转移到培养皿中(φ= 7 cm,H= 3 cm,培养皿中先放入2 cm厚的湿润沙土),每隔15 min观察记录一次老熟幼虫的化蛹情况。
-
于2016年6月—8月以平行线方式在实验样地沙棘林中不同位置悬挂黄色粘虫板4组,黄板中央添加沙棘绕实蝇引诱剂,黄板悬挂高度距地面约1.5 m。每日7:00于林间设置黄板,18:00收取黄板并统计沙棘绕实蝇成虫诱捕量,黄板及诱剂每天更换一次。样地内设置一台FSR-5A型超声波自动气象站(北京天裕德科技有限公司),监测林间气温变化(采样时间间隔1 h)。每天记录天气状况。
-
室内试验用虫来自于2015年5月在沙棘林中采集的越冬蛹,在室温条件下羽化后,用养虫盒(长18.5 cm×宽11.5 cm×高7 cm,盒盖留2个直径0.5 cm的透气孔并覆以纱网)饲养并观察。
-
成虫交尾:挑选新羽化的健康雌、雄成虫各20头,分别放入20个饲养盒中,每盒雌、雄成虫各一头。饲养盒中饲料(蛋白质:糖= 1:4)和水分充足。在每盒中放入长约10 cm的沙棘枝(保留叶和果实,模拟野外沙棘林微环境)。每隔30 min观察一次沙棘绕实蝇交尾情况。
成虫产卵:在上述交尾实验中,每日8:00更换新鲜的沙棘枝果,将取出的前一天果实放在显微镜下解剖,观察产卵情况。于2015年6月4日至7月7日每日测量30粒沙棘果的果径,确定产卵期果径大小。
-
对于沙棘绕实蝇成虫不同时间羽化量和不同时间钻出果实化蛹幼虫数,采用one-way ANVOA(Tukey法)进行多重比较。分析前原始数据加1后采用平方根(sqrt)进行数据转换。所有处理均使用SPSS 21.0软件完成。
-
在内蒙古西部磴口地区,沙棘绕实蝇成虫于每年6月中旬开始羽化,至7月下旬基本羽化结束。羽化2~3日后达到性成熟,便可开始交尾。羽化6~8天后开始产卵。卵期约7天。幼虫孵化后在沙棘果内以蛀食果肉为生,当取食完一粒沙棘果的果肉后幼虫会转移至相邻的果实中继续为害,一头幼虫最多为害10粒沙棘果。约30天后,老熟幼虫落地化蛹。野外用树冠下铺设地膜法监测,发现老熟幼虫于7月中旬开始出果化蛹,至9月上旬化蛹结束,以蛹越冬,生活史见表 1。
表 1 沙棘绕实蝇生活史(磴口,内蒙古)
Table 1. The life cycle of Rhagoletis batava obseuriosa in Dengkou county, Inner Mongolia, China
虫态
Developmental stage6月June 7月July 8月August 9月September 10月—次年5月
October to next MayF M L F M L F M L F M L 蛹pupa △ △ △ △ △ △ 成虫adult + + + + + + 卵egg ● ● ● ● ● ● 幼虫larva — — — — — — 蛹pupa △ △ △ △ △ △ △ △ △ 注:△:蛹;+:成虫;●:卵;—:幼虫; F, M, L分别表示每月的上、中、下旬。
Note: △: pupa;+: adult;●: egg;—: larva; F, M and L represent the first 10 days, the middle 10 days and the last 10 days of each month, respectively.
图 1 沙棘绕实蝇不同虫期
Figure 1. Different developmental stage of Rhagoletis batava obseuriosa
-
沙棘绕实蝇通过额囊的膨大将蛹壳顶破,依靠躯体向前蠕动和额囊胀缩,脱离蛹壳,挤开土壤,慢慢爬出地面。刚出土的成虫躯体细长,翅呈皱缩状,体壁柔软,体色也较浅。出土后迅速爬行寻找相对安全的地方静静展翅,通过额囊活动和腹部收缩运动,逐渐将皱缩的翅伸直展开,最后再用后足整理抚平。约经半小时,翅完全舒展。此时的实蝇体色尚浅,腹部细长,头部额囊也未收回。再经半小时左右,腹部不断收缩至饱满,额囊也收缩成扁圆形,体壁完全硬化,体色和翅斑变为正常。此后,成虫逐渐活跃,时而搓足,时而梳理头部和翅,直至起飞。
-
在磴口县,沙棘绕实蝇蛹最早于6月17日开始羽化,7月2日之后不再有成虫羽化,羽化期持续15天左右。羽化初期雄虫居多,羽化后期多雌虫,总体雌雄性比接近1:1。沙棘绕实蝇一天中的羽化时间较为集中,在6:00—10:00羽化量最多,占当日羽化量的81.3%,0:00—6:00及10:00—15:00有少量羽化,15:00—24:00几乎无成虫羽化(见图 2),不同时间段的羽化量有显著差异(F = 23.643,df = 109,P < 0.01)。当日0:00—7:00雌虫羽化居多,此后雄虫逐渐多于雌虫。
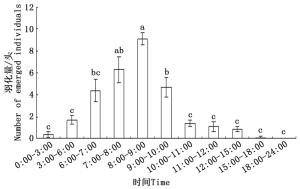
图 2 沙棘绕实蝇一天中不同时段的羽化节律
Figure 2. Note: Data are mean ± SE. The different lowercase letters on the columns mean significant difference at P < 0.01, Tukey's multiple comparison test.
-
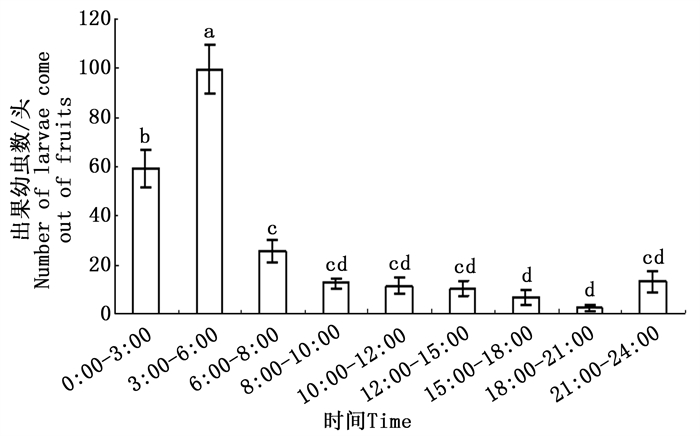
沙棘绕实蝇老熟幼虫在当地最早于7月17日开始钻出果实落地化蛹,至9月上旬基本结束。幼虫钻出果实的时间集中在0:00—6:00,其中以3:00 —6:00钻出果实的幼虫数最多,与其他时间段钻出果实化蛹的老熟幼虫数量有明显的差异(F = 40.717, df = 26, P < 0.01),6:00—18:00钻出果实的幼虫数量会逐渐减少,18:00—24:00出果虫数又开始增多(见图 3)。钻出果实的老熟幼虫很快开始化蛹,平均化蛹时间为276 ± 15 min,最短165 min,最长450 min。

图 3 沙棘绕实蝇老熟幼虫一天中不同时段的化蛹情况
Figure 3. The pupation pattern of Rhagoletis batava obseuriosa at different times of the day
-
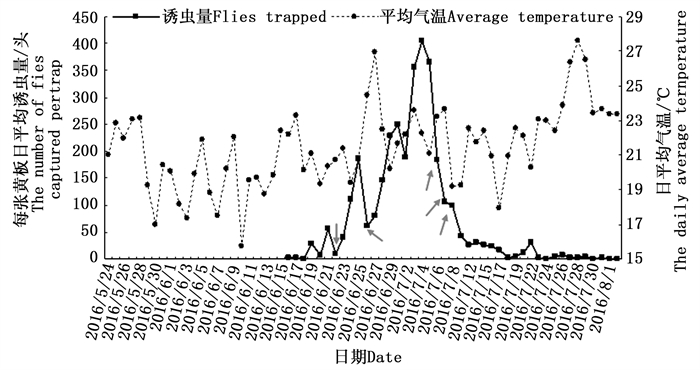
在观测地,整个沙棘绕实蝇成虫发生期持续45天左右(图 4)。沙棘绕实蝇发生期可以人为划分为四个阶段:第一阶段是从6月16日至6月22日,为沙棘绕实蝇发生初期,虫口密度较低;第二阶段从6月23日至7月7日,为沙棘绕实蝇发生中期,在此时期内虫口密度达到最高;从7月8日至7月22日为发生后期,虫口密度迅速下降;7月23日至8月1日为沙棘绕实蝇发生末期。沙棘绕实蝇虫口发生量与成虫期日平均气温并无明显的关系,但受天气的影响较大,如图 4所示,6月22日(阴天)、6月26日(阵雨,大风)和7月6—8日(阴转小雨)的天气变化造成了实蝇种群数量的较大波动。
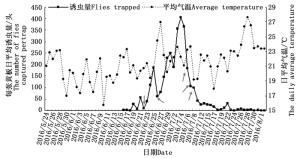
图 4 2016年沙棘绕实蝇野外发生动态
Figure 4. Population dynamic of Rhagoletis batava obseuriosa in the field monitored by trap from June to August in 2016
-
沙棘绕实蝇成虫羽化后非常活跃,一般补充一定营养后,羽化次日雌雄成虫即可交尾。交尾方式为雌下雄上重叠式。当雄虫渴望交尾时,它会寻找一个合适的区域,不停地用后足摩擦拨弄腹部末端的外生殖器,然后后足相互摩搓并涂抹双翅,有雌虫靠近时,会高频率地震动翅膀。性成熟的雌虫在枝果或者叶片上休息时,其前足时而摩擦时而拨弄头部,有时产卵器会伸出体外,后足不停地梳洗产卵器并摩擦涂抹双翅及腹部背面,有雄虫靠近时也会剧烈抖动翅膀。雌雄虫相遇时,如果彼此没有交尾欲望,一接触便会迅速跳开飞离。如果双方有交尾意愿,雄虫会在爬行移动的过程中瞅准时机,迅速飞跃到雌虫背部,前足和中足紧紧抱住雌虫胸背部及腹部前端区域,腹部微弯,后足挑拨雌虫产卵器与自身外生殖器对接,对接成功后雄虫后足抵触叶面或果面支撑身体以减轻对雌虫的压力。交尾过程中雌虫背负着雄虫可以进行游走、取食等活动,一般不会轻易分开。交尾结束时,有时是雄虫自愿结束交尾飞离雌虫的,有时是雌虫后足拨弄雄虫使其脱落,但有时雄虫不愿结束交尾,抱住雌虫不分开。
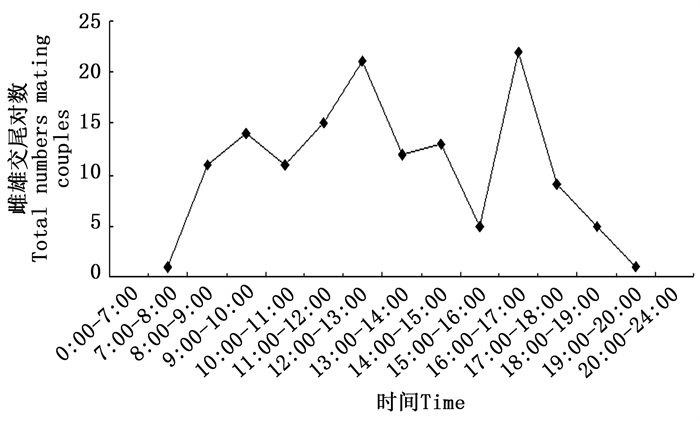
对沙棘绕实蝇交尾开始时间进行统计,发现沙棘绕实蝇多在白天交尾。最早发生在7:00左右,最晚的能够持续到凌晨3:00左右,夜间无交尾起始发生。一天中有两次交尾高峰期,第一次高峰期在12:00—13:00,第二次高峰期在16:00—17:00(见图 5)。成虫交尾持续时间最短约30 min,最长约720 min,平均持续时长为239 ± 11.86 min。雌雄虫一生中平均交尾次数为7.05 ± 3.69(SD)次。交尾后雌虫寿命(14.24 ± 3.38 d, SD)略长于雄虫(13 ± 3.12 d, SD)。
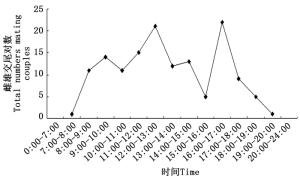
图 5 沙棘绕实蝇不同时间段交尾情况
Figure 5. The temporal pattern of mating of Rhagoletis batava obseuriosa
-
沙棘绕实蝇在羽化后68天开始产卵,产卵期沙棘果大小为长径6.65 ± 0.28 mm (SD),宽径5.47 ± 0.40 mm (SD)。沙棘绕实蝇的产卵过程可分为定位试探、穿刺产卵和清洁休整三个阶段。首先雌成虫搜索、飞行降落到沙棘果上,在沙棘果上一边绕圈游走一边不停振动翅膀,有时用产卵器刺探沙棘果,当找到果实的最佳产卵部位后,躯体做出“俯卧撑”姿态,腹部呈90°向下弯曲并伸出产卵器,通过腹部上下蠕动收缩,将卵产在果皮下。成虫产卵结束后拔出产卵器并用后足梳理清洁产卵器。沙棘绕实蝇产一粒卵耗时10 min左右。
成虫在一枚沙棘果中只产一粒卵,野外受害的沙棘果中有极少数的果中有23条幼虫,可能是幼虫在转果危害时相遇造成的结果。解剖镜下检查沙棘果时发现有极少部分卵镶嵌在果皮上,即一半暴露于空气中,一半包埋于果皮下,可能是沙棘绕实蝇产卵时的姿势不规整或者过早拔出产卵器所造成。
-
野外采用黄板诱捕沙棘绕实蝇成虫可以在一定程度上反映当地沙棘绕实蝇种群发生数量。通过对野外沙棘绕实蝇成虫发生动态的监测,发现沙棘绕实蝇成虫野外发生量与当时的气温关系并不明显,但天气情况对于诱捕数量影响较大,尤其是阴雨天对沙棘绕实蝇活动影响较大。降雨后(该地多为短时间的阵雨),土壤湿度增大,对于沙棘绕实蝇成虫羽化也产生一定影响。因此,有必要对各项气象因子,如环境湿度、风速、光照度等对沙棘绕实蝇成虫活动的影响进行研究,为今后利用引诱剂控制沙棘绕实蝇提供理论指导。
沙棘绕实蝇将卵产在果皮下,幼虫蛀食果肉,蛹在土壤中,所以控制沙棘绕实蝇的关键在成虫期。目前沙棘绕实蝇的防治主要是地表和林冠喷洒化学农药[9],但收效甚微,原因之一就是对沙棘绕实蝇的生活史不甚了解,尤其是对其防治最佳时期——成虫期了解不准确。所以,研究沙棘绕实蝇在当地的发生动态对于沙棘绕实蝇的精准防治意义较大。不过,虽然在大发生时可采用化学农药压低虫口密度,但应尽量少用,以保证果品质量。目前沙棘绕实蝇仅在我国北方地区零星爆发,但扩散和流行的可能性存在。因此,在其大范围发生之前应找到有效的绿色防治措施。
应用引诱剂监测和控制实蝇是当前实蝇防控工作中采用的主要技术[13-18],目前作者已初步研制出沙棘绕实蝇的引诱剂并用于监测和防治沙棘绕实蝇的成虫。另外,沙棘绕实蝇在一粒沙棘果中只产一粒卵,猜测雌成虫在产卵结束后会于沙棘果表面留下标记信息素。如果能够提取到该标记信息素并人工合成,就可在成虫产卵期于沙棘林喷洒以驱避实蝇,减轻危害。此外,采用不育技术防治实蝇危害在国际上已有先例[19-20],鉴于沙棘绕实蝇在我国内蒙、新疆主要危害成片的大果沙棘林,因此未来可考虑采用不育技术控制其危害。
沙棘在我国分布范围广,但是沙棘绕实蝇成虫自然扩散能力并不强,主要还是通过苗木运输造成远距离传播。例如苗木移植过程中根际携带的土块中可能包含沙棘绕实蝇活体蛹,所以在跨区域苗木移植过程中应加强检验检疫,阻止其扩散蔓延。
-
沙棘绕实蝇在内蒙古西部磴口地区一年发生一代,幼虫蛀食果肉,并可转果危害。在地表土层化蛹,蛹期长达9个多月。成虫期较短,自6月中下旬至8月上旬。成虫多在凌晨和上午羽化,性比约为1 : 1,寿命15天左右。交尾多发生在白天,可多次交配。防治工作应在成虫期进行。
沙棘绕实蝇的生物学习性
Biological Characteristics of Rhagoletis batava obseuriosa (Diptera: Tephritidae)
-
摘要:
目的 沙棘绕实蝇严重危害沙棘果实,使沙棘种植地蒙受巨大经济损失。研究其生物学习性,可以为有效监测和控制沙棘绕实蝇提供理论指导。 方法 通过室内人工饲养和野外观察,研究沙棘绕实蝇的羽化、交尾、产卵、化蛹等生物学习性和野外发生动态。 结果 沙棘绕实蝇成虫在野外从6月中旬至8月上旬均可见到,发生量受天气影响较大;成虫主要集中在6:00—10:00羽化,羽化量占当日羽化量的81.3%,羽化高峰出现在8:00—9:00;化蛹时间主要集中在凌晨0:00—6:00,尤其是3:00—6:00钻出果实化蛹的幼虫数量较多,与其它时间段有明显差异;交尾时间主要集中在白天光照比较强的时间段,一天当中有两次交尾高峰,分别出现在12:00—13:00和16:00—17:00,交尾平均持续时长为239±11.86 min;沙棘绕实蝇产卵期沙棘果长径6.65±0.28 mm,宽径5.47±0.40 mm,一果只产一卵。 结论 沙棘绕实蝇在内蒙古磴口地区一年发生一代,幼虫钻蛀果实,蛹隐藏在地下,成虫期较短,防治工作应集中在成虫期进行。 Abstract:Objective To study the biological characteristics of fruit fly, Rhagoletis batava obseuriosa Kol. (RBO) (Diptera: Tephritidae). Method The life cycle and biological characteristics of RBO, including eclosion, mating, oviposition, pupation and population dynamics were studied through laboratory and field observation in Dengkou county, Inner Mongolia. Result Adult RBO occurred from the middle June to the early August, and weather could have impact on adult amount. About 81.3% of RBO emerged from 6:00 to 10:00, and the peak time was 8:00 to 9:00. The larvae came out of fruit and pupated mainly in the early morning from 0:00 to 6:00, especially from 3:00 to 6:00. The mating time mainly occurred in the daytime and there were two peaks from 12:00 to 13:00 and 16:00 to 17:00. Usually, the mating lasts 239 ± 11.86 min. The fruit fly preferred to lay one egg into one fruit which the length was 6.65 ± 0.28 mm and the width was 5.47 ± 0.40 mm. The mating and egg-laying behavior of R. batava obseuriosa were studied in details too. Conclusion There is only one generation of RBO per year in Dengkou county, Inner Mongolia. The larvae develop within the fruit of Hippophae rhamnoides and pupate in the soil. Though adult period was short, it should be an exact stage for controlling the pest. -
Key words:
- Rhagoletis batava obseuriosa
- / Hippophae rhamnoides
- / eclosion
- / mating
- / population dynamic
-
图 2 沙棘绕实蝇一天中不同时段的羽化节律
Figure 2. Note: Data are mean ± SE. The different lowercase letters on the columns mean significant difference at P < 0.01, Tukey's multiple comparison test.
图 3 沙棘绕实蝇老熟幼虫一天中不同时段的化蛹情况
Figure 3. The pupation pattern of Rhagoletis batava obseuriosa at different times of the day
图 4 2016年沙棘绕实蝇野外发生动态
Figure 4. Population dynamic of Rhagoletis batava obseuriosa in the field monitored by trap from June to August in 2016
图 5 沙棘绕实蝇不同时间段交尾情况
Figure 5. The temporal pattern of mating of Rhagoletis batava obseuriosa
表 1 沙棘绕实蝇生活史(磴口,内蒙古)
Table 1. The life cycle of Rhagoletis batava obseuriosa in Dengkou county, Inner Mongolia, China
虫态
Developmental stage6月June 7月July 8月August 9月September 10月—次年5月
October to next MayF M L F M L F M L F M L 蛹pupa △ △ △ △ △ △ 成虫adult + + + + + + 卵egg ● ● ● ● ● ● 幼虫larva — — — — — — 蛹pupa △ △ △ △ △ △ △ △ △ 注:△:蛹;+:成虫;●:卵;—:幼虫; F, M, L分别表示每月的上、中、下旬。
Note: △: pupa;+: adult;●: egg;—: larva; F, M and L represent the first 10 days, the middle 10 days and the last 10 days of each month, respectively. 下载: 导出CSV
下载: 导出CSV
-
[1] 廉永善, 陈学林. 沙棘的生态地理分布及其植物地理学意义[J]. 植物分类学报, 1992, 18(4): 11-12. [2] 张建国, 黄铨, 罗红梅. 沙棘优良杂种选育研究[J]. 林业科学研究, 2005, 18(4): 381-386. doi: 10.3321/j.issn:1001-1498.2005.04.003 [3] 陈云明, 刘国彬, 徐炳成, 等. 我国沙棘水土保持功能研究进展与展望[J]. 中国水土保持科学, 2004, 2(2): 88-92. [4] 丁小林, 秦利平. 沙棘中的营养成分与生物活性物质研究进展[J]. 中国食物与营养, 2008(9): 57-59. [5] 刘勇, 廉永善, 王颖莉, 等. 沙棘的研究开发评述及其重要意义[J]. 中国中药杂志, 2014, 39(9): 1547-1552. [6] 武福亨, 赵玉珍. 前苏联沙棘病虫害的研究与防治[J]. 国际沙棘研究与开发, 2004, 2(4): 44-48. [7] 范仁俊, 董晋明, 曹满, 等. 山西省沙棘病虫草害及相关天敌的调查[J]. 沙棘, 1994, 7(3): 10-13. [8] 葛葆蔚, 李桂和, 张玉伟, 等. 沙棘果实蝇的初步研究[J]. 辽宁林业科技, 1988(3): 45-46. [9] 葛葆蔚. 沙棘果实蝇防治技术的研究[J]. 中国森林病虫, 1991(2): 24-25. [10] 陈孝达, 党心德, 李锋. 陕西省沙棘昆虫区系的特点及分析[J]. 沙棘, 2001, 14(2): 23-26. [11] 刘金江. 黑龙江省沙棘主要病虫草害及综合防治[J]. 沙棘, 2005, 18(4): 11-12. [12] 魏建荣, 苏智, 刘明虎, 等. 沙棘果实的重要检疫害虫-沙棘绕实蝇的发生与危害[J]. 内蒙古林业科技, 2012, 38(4): 55-57. [13] 张钧. 引诱剂对实蝇类害虫的诱捕效果[J]. 植物检疫, 1991(6): 401-404. [14] Prokopy RJ, Miller NW, Piero JC, et al. Effectiveness of GF-120 fruit fly bait spray applied to border area plants for control of melon flies (Diptera: Tephritidae)[J]. Journal of Economic Entomology, 2003, 96(5): 1485-1493. [15] Castrejón-Gómez VR, Aluja M, Arzuffi R, et al. Two low-cost food attractants for capturing Toxotrypana curvicauda (Diptera: Tephritidae) in the field[J]. Journal of Economic Entomology, 2004, 97(2): 310-315. [16] 黄素青, 韩日畴. 桔小实蝇的研究进展[J]. 昆虫知识, 2005, 42(5): 479-484. [17] Khrimian A, Jang EB, Nagata J, et al. Consumption and metabolism of 1, 2-dimethoxy-4-(3-fluoro-2-propenyl) benzene, a fluorine analog of methyl eugenol, in the oriental fruit fly Bactrocera dorsalis (Hendel)[J]. Journal of Chemical Ecology, 2006, 32(7): 1513-1526. [18] 杜迎刚, 陈家骅, 季清娥. 一种新型蛋白诱剂对橘小实蝇引诱作用. 福建林学院学报, 2007, 27(3): 259-262. [19] Mcinnis DO, Shelly TE, Komatsu J. Improving Male Mating Competitiveness and Survival in the Field for Medfly, Ceratitis Capitata (Diptera: Tephritidae) SIT Programs[J]. Genetica, 2002, 116(1): 117-124. [20] Enkerlin WR. Impact of fruit fly control programmes using the sterile insect technique. In: Dyck VA, Hendrichs J, Robinson AS. Sterile insect technique[M]. Springer Netherlands. 2005: 651-676. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5720
- HTML全文浏览量: 1700
- PDF下载量: 632
- 被引次数: 0
