

-
我国是木兰科植物资源最丰富的国家,约有140种,木兰科植物是亚热带常绿阔叶林的优势树种之一,多为高大乔木[1-2]。木兰科植物多数属于濒危树种,数量稀少且具有广泛用途,长期以来是引种驯化的热门树种[3-4]。广东含笑(Michelia guangdongensis Y. H. Yan et al.)为木兰科含笑属常绿灌木或小乔木,原产于广东英德,是近年来新发现的木兰科中的广东特有树种,生于海拔1 250~1 400 m的中亚热带山地常绿落叶阔叶混交林及山顶灌丛中,其树形美观,叶片锈色光亮,花大洁白、芬芳,适宜庭园或公园种植,也可盆栽观赏,是优良的园林绿化树种。目前,广东含笑仍处于野生未被广泛开发阶段,在园林中未进行广泛应用。对广东含笑的研究多集中于扩繁、育种、引种驯化等方面[5-8],而对其光合生理特性的研究尚未见报道。对广东含笑的光合参数进行测定与分析及对光合-光响应的最适拟合模型进行研究,对广东含笑在园林绿化中推广应用具有重要指导意义。光响应曲线的拟合模型很多,主要包括直角双曲线模型、非直角双曲线模型、指数模型、改进指数模型和叶子飘光合模型(直角双曲线修正模型)等,不同的模型对同一种植物的光响应曲线的拟合结果存在差异[9]。本研究以嫁接4年生广东含笑为研究对象,利用叶子飘光合模型、改进指数模型、非直角双曲线模型、直角双曲线模型、指数模型和二次多项式回归模型对其光响应曲线进行拟合,探讨6种模型的适用性,筛选最适模型,根据该模型得到广东含笑的光合生理参数,为广东含笑的引种栽培及园林应用提供参考。
-
试验材料广东含笑为广东省林业科学研究院资源收集圃内4年生嫁接植株,选择3株长势优良的植株,每株选取3片健康、完整的植株中上部向阳叶片进行光响应曲线测定。
-
于2016年6月上旬,选择晴朗无风(微风)的天气,在早上9:00—11:30,使用Li-6400便携式光合测定仪(Li-Cor Inc.,Lincoln, NE,USA)进行测定,以小钢瓶内液态CO2为气源,设置成自然CO2浓度条件(400 μmol·mol-1),为保持其他环境因子的稳定,设定叶室温度为(28±0.5)℃,将空气流速设为500 μmol·s-1,样本室相对湿度控制在60%左右,红蓝光源设置叶室内的光合有效辐射强度(I)梯度为:2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2·s-1,测量3株含笑,每株选取3片叶进行测量,每片叶重复测量3次。
-
将光合-光响应曲线的数据从光合仪中导出,每个光合有效辐射对应的测量数据使用SPSS21.0中非线性回归模块完成,得出拟合值。为更好地检验6个模型拟合的精确度,采用拟合值均方误差(MSE)和拟合值平均绝对误差(MAE)2个参数作拟合精确度比较。
$ MSE = \frac{1}{n}\sum\limits_{t = 1}^n {{{\left( {{y_t} - \hat y} \right)}^2}} $
$ MAE = \frac{1}{n}\sum\limits_{t = 1}^n {\left| {{y_t} - {{\hat y}_t}} \right|} $
式中:yt和分别代表实测值和拟合值,误差越小说明拟合值越接近实测值[10]。
通过以下6种模型的拟合方程,计算光合-光响应对应的光饱和点(Isat)、光补偿点(Ic)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)、初始量子效率(φ)、光响应曲角(θ)等生理参数,并和实际测量值进行比较。6种模型如下:
非直角双曲线模型[11]:
$ {P_n} = \frac{{\varphi I + {P_{n\max }} - \sqrt {{{\left( {\varphi I + {P_{n\max }}} \right)}^2} - 4\varphi I\theta {P_{n\max }}} }}{{2\theta }} - {R_d} $
直角双曲线模型[12]:
$ {P_n} = \frac{{\varphi I{P_{n\max }}}}{{\varphi I + {P_{n\max }}}} - {R_d} $
$ {P_n} = \alpha \frac{{1 - \beta I}}{{1 + \gamma I}}\left( {I - {I_C}} \right) $
指数模型[15]
$ {P_n} = {P_{n\max }}\left[ {1 - {{\rm{e}}^{\varphi \left( {I - {I_C}} \right)}}} \right] $
$ {P_n} = \alpha {{\rm{e}}^{( - \beta I)}} - \gamma {{\rm{e}}^{( - \varepsilon I)}} $
二次多项式回归模型[17]:
$ {P_n} = a{I^2} + bI + c $
以上各模型中:Pn为净光合速率(μmol·m-2 ·s-1);Pnmax为最大净光合速率(μmol·m-2 ·s-1);I为光合有效辐射;Ic为光补偿点;Rd为暗呼吸速率;φ为初始量子效率;θ为光响应曲角;叶子飘光合模型中α为I = 0的初始斜率,β为光抑制系数,γ为饱和系数;改进指数模型和二项式中,α、β、ε和a、b、c为待定参数。
-
由表 1可看出:广东含笑光合-光响应曲线除改进指数模型和叶子飘光合模型的拟合值与实测值接近外,其他4个模型的光合有效辐射对应拟合值都不同程度大于实测值。由图 1可知:当光合有效辐射≤400 μmol·m-2·s-1时,广东含笑的净光合速率(Pn)呈线性增长趋势;随着光合有效辐射的增大,广东含笑的Pn缓慢增大至光饱和点,对应出现最大净光合速率(Pnmax);当光合有效辐射在1 400~2 000 μmol·m-2·s-1时,非直角双曲线模型、直角双曲线模型、指数模型、改进指数模型拟合出的Pn均高于实测值,且没有出现光饱和区域的光抑制现象,净光合速率随光合有效辐射的增大持续增大,而二次多项式回归模型拟合曲线虽然有光饱和点,但不能较好的解释光合有效辐射超过此计算所得光饱和点(Isat)后Pn值快速下降的原因;相反,叶子飘光合模型在1 400 μmol·m-2·s-1光合有效辐射下的拟合值小于实测值,且随后净光合速率趋于平稳,具有光抑制现象。因此,叶子飘光合模型的拟合曲线能体现广东含笑光合作用在光饱和点下受到抑制的现象,且随着光合有效辐射的增大,与广东含笑实测Pn的变化幅度基本一致。
表 1 广东含笑光合作用光响应曲线实测值及模型拟合值
Table 1. The measured and estimated data of photosynthesis light-response curve of M. guangdongensis
光合有效辐射
Photosynthetically available radiation/(μmol·m-2·s-1)净光合速率Net photosynthesis rate/(μmol·m-2·s-1) 实测值
Measured values改进指数模型
Modified exp-onential model叶子飘光合模型
Modified rectangular hyperbola model指数模型
Exponential model非直角双曲线模型
Nonrectangular hyperbola model直角双曲线模型
Rectangular hyperbola model二次多项回归模型
Quadratic polynomial regression model2 000 13.66 14.10 13.45 14.23 14.87 15.37 11.19 1 800 13.43 13.97 13.44 14.22 14.67 15.19 12.67 1 600 13.35 13.83 13.37 14.19 14.47 14.96 13.59 1 400 13.27 13.63 13.23 14.13 14.26 14.69 13.95 1 200 12.82 13.37 12.99 14.01 14.02 14.33 13.76 1 000 12.59 13.01 12.61 13.78 13.71 13.87 12.99 800 11.94 12.48 11.99 13.31 13.24 13.21 11.67 600 11.11 11.62 11.01 12.39 12.41 12.25 9.79 400 9.56 10.04 9.31 10.56 10.77 10.69 7.35 200 6.19 6.67 6.11 6.94 7.31 7.72 4.35 150 4.73 5.29 4.83 5.57 5.95 6.49 3.52 100 3.12 3.58 3.29 3.95 4.28 4.91 2.65 50 1.03 1.52 1.17 2.03 2.27 2.78 1.74 20 -0.35 0.11 -0.37 0.71 0.83 1.11 1.18 0 -1.45 -0.91 -1.47 -0.24 -0.22 -0.24 0.80 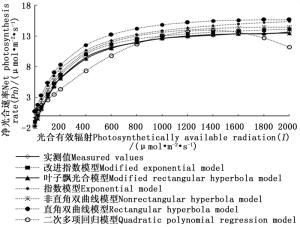
图 1 6种模型拟合的光合-光响应曲线
Figure 1. Light-response curves of photosynthesis for M. guangdongensis
-
利用6种模型分别对广东含笑实测光响应曲线进行拟合,得到光合参数,由表 2可知:除二次多项式回归模型的决定系数R2为0.940外,其余5个模型的拟合度均较高,决定系数R2均>0.990,但R2值越大只说明该模型的拟合度较高,并不能保证拟合结果与实测值相符,因此,引入测量学中平均绝对误差(MAE)和均方误差(MSE)对拟合结果的精确度进行检验。6个模型中,叶子飘光合模型的MAE与MSE最小,其次是改进指数模型,直角双曲线模型和二次多项回归模型较大,说明拟合值与实测值间相差较大;R2越大,MAE和MSE越小,说明拟合效果越好,拟合值和实测值越接近。二次多项式的拟合系数较低,对广东含笑光响应曲线拟合的整体效果不太好,且由回归方程拟合出的暗呼吸速率为正数,植物暗呼吸速率实际为负值,因此,用二项式回归分析法分析广东含笑的光响应曲线不符合植物的光响应特征。在光合参数拟合中,叶子飘光合模型拟合的光补偿点Ic和光饱和点Isat值与实测值最接近;而非直角双曲线模型和直角双曲线模型一般是借助低光合有效辐射下(I≤200 μmol·m-2·s-1)直线回归后计算各光合参数,拟合的光饱和点Isat值远低于实测值,最大光合速率则远大于实测值,初始量子效率偏大,这与前人应用这2个模型拟合的结论一致[18]。因此,这2种模型不适合用于广东含笑的光响应拟合。指数模型一般需要假设Pn为90% Pnmax或99%Pnmax所对应的光合有效辐射,以达到求出光饱和点(Isat)。通过假设Pn=99%Pnmax计算得出广东含笑的Isat为378.83 μmol·m-2·s-1,同样远小于实测值。改进指数模型计算得出的广东含笑的Isat和Pnmax均偏小于实测值,在实际运用中,改进指数模型更多的被学者运用于阴生植物的光响应拟合中。因此,6个模型中,叶子飘光合模型拟合的光合-光响应曲线与广东含笑实测值最接近,最适宜广东含笑的光响应曲线拟合。由叶子飘光合模型计算出的广东含笑的最大净光合速率为13.46 μmol·m-2·s-1,光饱和点为1 598.54 μmol·m-2·s-1,光补偿点为26.74 μmol·m-2·s-1,暗呼吸速率为-1.58 μmol·m-2·s-1,初始量子效率为0.06在0.04~0.07之间。
表 2 6个模型精确度及各项光合参数的比对
Table 2. Six models accuracy and comparison of photosynthetic parameters
参数
Parameters实测值
Measured values叶子飘光合模型
Modified rectangular hyperbola model改进指数模型
Modified exponential model指数模型
Exponential model非直角双曲线模型
Nonrectangular hyperbola model直角双曲线模型
Rectangular hyperbola model二次多项回归模型
Quadratic polynomial regression model决定系数R2 - 0.999 0.995 0.993 0.992 0.992 0.940 平均绝对误差MAE - 0.096 0.499 0.999 1.200 1.500 1.851 均方误差MSE - 0.013 0.252 1.043 1.444 2.305 1.653 光补偿点Ic/ (μmol·m-2·s-1) ≈25 26.74 28.66 26.10 28.25 26.56 50.57 光饱和点Isat/ (μmol·m-2·s-1) ≈1 600 1 598.54 1 056.59 378.83 438.13 433.06 1 400.43 最大净光合速率Pnmax/ (μmol·m-2·s-1) 13.66 13.46 10.83 14.50 17.81 17.48 11.20 暗呼吸速率Rd/ (μmol·m-2·s-1) -1.45 -1.58 -1.42 -1.25 -1.41 -1.75 0.80 初始量子效率φ - 0.06 0.05 0.05 0.05 0.07 - 曲角θ - - - - 0.517 - - -
光适应特性在植物间存在差异,植物的光响应曲线反映了光合速率随光照强度的变化特性。进行光响应的研究有利于分析光合产物积累与环境的关系,而分析环境因素影响是植物生长与发育的关键[19]。光响应曲线是确定光合速率、初始量子效率、光饱和点、光补偿点等植物重要生态生理参数的主要手段[20-22]。本文利用二次多项式、直角双曲线模型、非直角双曲线模型、叶子飘光合模型、指数模型及改进指数模型对广东含笑的光合-光响应曲线数据进行了拟合比较,结果表明:应用二次多项式拟合得到的光响应模型相关系数较低,R2值仅0.940,其他5种模型对广东含笑的光响应拟合效果均较好(R2>0.990),其中,叶子飘光合模型的R2值最高,为0.999。陈卫英等[10]提出,均方误差(MSE)和平均绝对误差(MAE) 越小,拟合模型具有较高的精确性和适宜性,用MSE和MAE检验6种模型的精确度,结果表明:叶子飘光合模型拟合值的MSE、MAE均小于其它模型。因此,叶子飘光合模型为广东含笑光响应曲线的最佳拟合模型,这与叶子飘提出的叶子飘光合模型更适合拟合植物在不同生境下的光响应曲线的结果一致[23]。直角双曲线模型、非直角双曲线模型和指数模型拟合的Pnmax值比实测值大,而Isat偏小,从曲线上看,在达到光饱和点后,三者均没表现出净光合速率受到抑制的现象。本研究中,改进指数模型拟合的光补偿点接近实测值,但光饱和点和净光合速率均小于实测值。前人研究认为,改进指数模型多数用于阴生植物光合-光响应曲线的拟合[10, 24-25],对阳生植物是否同样适用还有待进一步研究。
利用叶子飘光合模型可以计算出广东含笑的Ic、Isat、φ、Rd、Pnmax等参数,为广东含笑植物的生理生态研究提供基础理论依据。广东含笑的Pnmax为13.46 μmol·m-2·s-1、Isat为1 598.54 μmol·m-2 ·s-1、Ic为26.74 μmol·m-2·s-1、初始量子效率为0.06,Rd为-1.58 μmol·m-2·s-1。Isat和Ic是植物的2个重要光合生理指标,反映了植物光照条件的要求,是判断植物耐阴性的2个重要指标[26]。一般认为,阳性植物的光饱和点为1 500~2 000 μmol·m-2 ·s-1,或者更高,光补偿点为50~100 μmol·m-2 ·s-1;阴性植物的光饱和点较阳性植物低,光补偿点 <20 μmol·m-2·s-1。本研究中,广东含笑的Isat稍大于1 500 μmol·m-2 ·s-1,具有较强的光能利用能力;广东含笑的光补偿点为26.74 μmol·m-2·s-1小于50 μmol·m-2 ·s-1,说明广东含笑能够在较弱的光照环境下积累光合产物,具有一定的耐荫能力,这与胡小菱[27]研究广东含笑天然分布于气候比较荫凉湿润的常绿阔叶林,属于一种稍耐侧荫的树种的结论一致。广东含笑在半阴或者阳光充足的环境都能生长,但形态上具有一定的差异,在半阴环境下,叶片的叶色较绿,新叶红锈色光泽不明显;在强光环境中,叶色鲜红,带红锈色的叶片数量显著多于半阴生的环境。此外,广东含笑的初始量子效率均较高,而初始量子效率能够反映植物利用弱光进行光合作用的能力,初始量子效率越高,说明植物能利用弱光进行光合作用的能力较强,在 < 200 μmol·m-2·s-1的弱光环境下,随着光合有效辐射的加强,光合速率能够迅速上升[19]。图 1也表明,在弱光下,广东含笑的光合速率迅速增加。
-
叶子飘光合模型不仅拟合效果最佳,且与广东含笑的光响应曲线的实测值最接近,相关系数为0.999,其拟合的光合参数均与实测值相吻合,能够反映出广东含笑在强光条件下出现光抑制的生理规律,是广东含笑光响应特性拟合的最佳模型;广东含笑的光饱和点为1 598.54 μmol·m-2 ·s-1,是阳性植物,具有较好的光能利用能力,光补偿点为26.74 μmol·m-2·s-1、初始量子效率为0.06,具有一定的耐荫能力,对光照的适应范围较宽,对弱光利用能力较强,适应能力也非常强,广东含笑具有广阔的开发前景。因此,在对广东含笑进行引种栽培或扦插时,应充分考虑其光照适应性,为其提供充足的光照条件。
广东含笑的光响应特性及其最适模型研究
Photosynthetic Light Response Characteristics of Michelia guangdongensis and Practicability of Six Models
-
摘要:
目的 通过对广东含笑光合-光响应曲线及其最适拟合模型的研究,了解广东含笑的光合生理参数,为广东含笑的引种栽培及园林应用提供参考。 方法 选用直角双曲线模型、非直角双曲线模型、叶子飘光合模型、指数模型、改进指数模型和二次多项式6种模型对广东含笑光响应曲线进行拟合,探讨不同光响应模型对广东含笑光合特性的适用性,筛选最佳光响应模型。 结果 表明:(1)广东含笑叶片光合速率与光合有效辐射之间存在非线性关系,叶子飘光合模型拟合的光合-光响应曲线最佳,具有符合植物生长规律的光抑制现象,拟合效果优于其它5种模型;(2)在叶子飘光合模型的光合参数中,最大净光合速率、光饱和点、光补偿点和暗呼吸速率与实测值最接近,决定系数为0.999,6个模型中最大,平均绝对误差和均方误差最小,模型具有较高的拟合精确度;(3)根据叶子飘光合模型拟合出广东含笑叶片的光饱和点为1 598.54 μmol·m-2 ·s-1、光补偿点为26.74 μmol·m-2·s-1、最大净光合速率为13. 46 μmol·m-2·s-1、暗呼吸速率为-1.58 μmol·m-2·s-1,初始量子效率为0.06。 结论 叶子飘光合模型为广东含笑光合-光响应曲线的最佳拟合模型,广东含笑为阳生植物,喜温暖、湿润气候,耐荫,具有广阔的开发前景,这为深入研究广东含笑的生理生态提供了参考。 Abstract:Objective In order to reveal the physiological and ecological characteristics of Michelia guangdongensis, and provide a theoretical basis for its cultivation in landscape greening, the photosynthesis light response curves and fitting of application model were studied. Method Six typical models of light-response curve of leaf net photosynthesis, such as quadratic polynomial regression model, non-rectangular hyperbola model, rectangular hyperbola model, modified rectangular hyperbola model, exponential model and modified exponential model, were used to fit the photosynthesis light response curves of M. guangdongensis and the best fitting model was determined. Result (1) There was a nonlinear relationship between the leaf photosynthesis rate and photosynthetically available radiation. The modified rectangular hyperbola model had the best photosynthetic-optical response curve, and had the light suppression phenomenon in accordance with the law of plant growth. The fitting effect is better than that of the other five models. (2) The parameters of Pnmax, Isat, Ic, and Rd, calculated by modified rectangular hyperbola model were the closest to the measured values. The determined coefficient R2 was 0.999, which was the largest among the six models. In addition, the MAE and MSE were the smallest and the highest fitting accuracy among these models. (3) The light saturation point (Isat) was 1 598.54 μmol·m-2 ·s-1 and the light compensation point (Ic) was 26.74 μmol·m-2 ·s-1, the maximum net photosynthesis rate (Pnmax) was 13.46 μmol·m-2·s-1, the dark respiration rate (Rd) was-1.58 μmol·m-2·s-1 and the Initial quantum efficiency (φ) was 0.06. Conclusion The modified rectangular hyperbola model is the best fitting model for the photosynthesis light response curve of M. guangdongensis. M. guangdongensis is a sun-pollinating plant, which is warm, humid and shade-tolerant, so this species has an expansive developing foreground. -
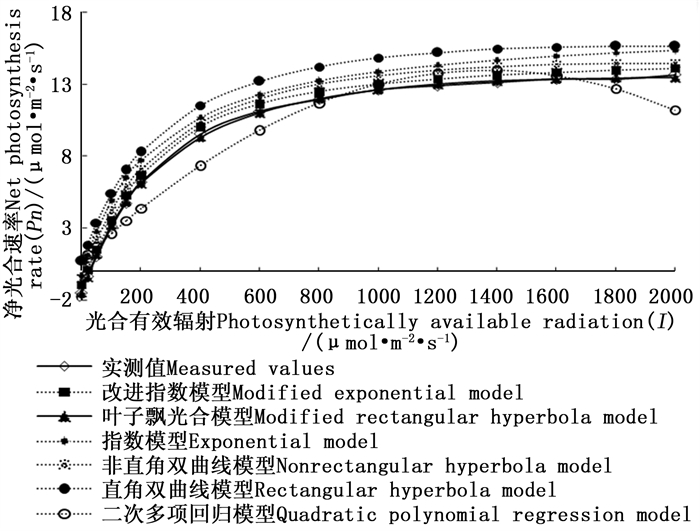
图 1 6种模型拟合的光合-光响应曲线
Figure 1. Light-response curves of photosynthesis for M. guangdongensis
表 1 广东含笑光合作用光响应曲线实测值及模型拟合值
Table 1. The measured and estimated data of photosynthesis light-response curve of M. guangdongensis
光合有效辐射
Photosynthetically available radiation/(μmol·m-2·s-1)净光合速率Net photosynthesis rate/(μmol·m-2·s-1) 实测值
Measured values改进指数模型
Modified exp-onential model叶子飘光合模型
Modified rectangular hyperbola model指数模型
Exponential model非直角双曲线模型
Nonrectangular hyperbola model直角双曲线模型
Rectangular hyperbola model二次多项回归模型
Quadratic polynomial regression model2 000 13.66 14.10 13.45 14.23 14.87 15.37 11.19 1 800 13.43 13.97 13.44 14.22 14.67 15.19 12.67 1 600 13.35 13.83 13.37 14.19 14.47 14.96 13.59 1 400 13.27 13.63 13.23 14.13 14.26 14.69 13.95 1 200 12.82 13.37 12.99 14.01 14.02 14.33 13.76 1 000 12.59 13.01 12.61 13.78 13.71 13.87 12.99 800 11.94 12.48 11.99 13.31 13.24 13.21 11.67 600 11.11 11.62 11.01 12.39 12.41 12.25 9.79 400 9.56 10.04 9.31 10.56 10.77 10.69 7.35 200 6.19 6.67 6.11 6.94 7.31 7.72 4.35 150 4.73 5.29 4.83 5.57 5.95 6.49 3.52 100 3.12 3.58 3.29 3.95 4.28 4.91 2.65 50 1.03 1.52 1.17 2.03 2.27 2.78 1.74 20 -0.35 0.11 -0.37 0.71 0.83 1.11 1.18 0 -1.45 -0.91 -1.47 -0.24 -0.22 -0.24 0.80  下载: 导出CSV
下载: 导出CSV
表 2 6个模型精确度及各项光合参数的比对
Table 2. Six models accuracy and comparison of photosynthetic parameters
参数
Parameters实测值
Measured values叶子飘光合模型
Modified rectangular hyperbola model改进指数模型
Modified exponential model指数模型
Exponential model非直角双曲线模型
Nonrectangular hyperbola model直角双曲线模型
Rectangular hyperbola model二次多项回归模型
Quadratic polynomial regression model决定系数R2 - 0.999 0.995 0.993 0.992 0.992 0.940 平均绝对误差MAE - 0.096 0.499 0.999 1.200 1.500 1.851 均方误差MSE - 0.013 0.252 1.043 1.444 2.305 1.653 光补偿点Ic/ (μmol·m-2·s-1) ≈25 26.74 28.66 26.10 28.25 26.56 50.57 光饱和点Isat/ (μmol·m-2·s-1) ≈1 600 1 598.54 1 056.59 378.83 438.13 433.06 1 400.43 最大净光合速率Pnmax/ (μmol·m-2·s-1) 13.66 13.46 10.83 14.50 17.81 17.48 11.20 暗呼吸速率Rd/ (μmol·m-2·s-1) -1.45 -1.58 -1.42 -1.25 -1.41 -1.75 0.80 初始量子效率φ - 0.06 0.05 0.05 0.05 0.07 - 曲角θ - - - - 0.517 - -
下载: 导出CSV
-
[1] 盘李军, 冼杆标, 陈伟光, 等. 深山含笑等18个景观树种光合生理特性研究[J]. 广东林业科技, 2013, 29(4): 33-37. doi: 10.3969/j.issn.1006-4427.2013.04.007 [2] 姜卫兵, 曹晶, 李刚, 等. 我国木兰科观赏新树种的开发及在园林绿化中的应用[J]. 上海农业学报, 2005, 21(2): 68-73. doi: 10.3969/j.issn.1000-3924.2005.02.016 [3] 王献溥, 蒋高明. 中国木兰科植物受威胁的状况及其保护措施[J]. 植物资源与环境学报, 2001, 10(4): 43-47. doi: 10.3969/j.issn.1674-7895.2001.04.010 [4] 李晓储, 黄利斌, 张永兵, 等. 四种含笑叶解剖性状与抗旱性的研究[J]. 林业科学研究, 2006, 19(2): 177-181. doi: 10.3321/j.issn:1001-1498.2006.02.009 [5] 杨科明, 陈新兰. 广东含笑的引种繁育与园林应用研究[J]. 广东园林, 2011, 33(1): 44-46. doi: 10.3969/j.issn.1671-2641.2011.01.011 [6] 姜春宁, 林伟, 黄永芳. 低温胁迫对广东含笑嫁接苗生理生化指标的影响[J]. 福建林业科技, 2016, 43(2): 123-128. [7] 姜春宁, 黄永芳. 广东含笑嫁接试验[J]. 广东林业科技, 2013, 29(2): 7-10. doi: 10.3969/j.issn.1006-4427.2013.02.002 [8] 张宗享. 优良绿化树种——广东含笑[J]. 林业科技通讯, 1982(5): 6-7, 2. [9] 刘强, 李凤日, 谢龙飞. 人工长白落叶松冠层光合作用-光响应曲线最优模型[J]. 应用生态学报, 2016, 27(8): 2420-2428. [10] 陈卫英, 陈真勇, 罗辅燕, 等. 光响应曲线的指数改进模型与常用模型比较[J]. 植物生态学报, 2012, 36(12): 1277-1285. [11] 叶子飘, 李进省. 光合作用对光响应的直角双曲线修正模型和非直角双曲线模型的对比研究[J]. 井冈山大学学报: 自然科学版, 2010, 31(3): 38-44. [12] Baly E C. The kinetics of photosynthesis[J]. Proceedings of the Royal Society of London Series B (Biological Sciences), 1935, 117(804): 218-239. [13] Ye Z P. A new model for relationship between light irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica, 2007, 45(4): 637-640. doi: 10.1007/s11099-007-0110-5 [14] Ye Z P, Yu Q. A coupled model of stomatal conductance and photosynthesis for winter wheat[J]. Photosynthetica, 2008, 64(4): 637-640. [15] Potvin C, Lechowicz M J, Tardif S. The Statistical Analysis of Ecophysiological Response Curves Obtained from Experiments Involving Repeated Measures[J]. Ecology, 1990, 71(4): 1389-1400. doi: 10.2307/1938276 [16] Chen Z Y, Peng Z S, Yang J, et al. A mathematical model for describing light-response curves in Nicotiana tabacum L[J]. Photosynthetica, 2011, 49(3): 467-471. [17] 梁文斌, 聂东伶, 吴思政, 等. 短梗大参光合作用光响应曲线及模型拟合[J]. 经济林研究, 2014, 32(4): 38-44. [18] 段爱国, 张建国. 光合作用光响应曲线模型选择及低光强属性界定[J]. 林业科学研究, 2009, 22(6): 765-771. doi: 10.3321/j.issn:1001-1498.2009.06.003 [19] 王雁, 马武昌. 扶芳藤、紫藤等7种藤本植物光能利用特性及耐荫性比较研究[J]. 林业科学研究, 2004, 17(3): 305-309. doi: 10.3321/j.issn:1001-1498.2004.03.007 [20] 邓云鹏, 雷静品, 潘磊, 等. 不同种源栓皮栎光响应曲线的模型拟合及参数比较[J]. 生态学杂志, 2016, 35(2): 387-394. [21] 郑益兴, 彭兴民, 吴疆翀, 等. 印楝种源不同生长季节冠层间的光响应特征[J]. 林业科学研究, 2011, 24(2): 176-183. [22] 刘娟, 梁军生, 王健敏, 等. 思茅松干季光合生理日动态及光响应特征分析[J]. 林业科学研究, 2009, 22(5): 677-682. doi: 10.3321/j.issn:1001-1498.2009.05.011 [23] 叶子飘. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J]. 生态学杂志, 2007, 26(8): 1323-1326. [24] 陈兰英, 黎云祥, 钱一凡, 等. 改进指数模型对紫茉莉光合-光响应及CO2响应适用性研究[J]. 广西植物, 2013, 33(6): 839-845. [25] 罗辅燕, 陈卫英, 陈真勇. 指数改进模型在大麦光合-CO2响应曲线中的适用性[J]. 植物生态学报, 2013, 37(7): 650-655. [26] 巨关升, 武菊英. 观赏狼尾草光合特性的研究[J]. 核农学报, 2005, 19(6): 451-455. doi: 10.3969/j.issn.1000-8551.2005.06.010 [27] 胡小菱. 含笑的园林应用与苗木培育[J]. 安徽林业科技, 2005(4): 38, 37. -

 点击查看大图
点击查看大图
图(1) / 表(2)
计量
- 文章访问数: 3991
- HTML全文浏览量: 1461
- PDF下载量: 605
- 被引次数: 0
