

-
森林更新是生态系统动态中森林资源再生产的一个极为重要的生物学过程,也是森林生态系统自我繁衍恢复的重要手段之一,对于植物群落在时间和空间上不断延续、发展或发生演替,以及未来森林群落的结构与其生物学多样性具有深远的影响[1-3]。森林自然更新过程极为复杂,更新过程中受到如自然生态因子、更新树种的遗传学、生态学特性等影响,而当前影响因子的研究主要集中在地形条件(如坡位、坡向、海拔)、林分条件(如郁闭度、林分密度)、土壤条件(如土壤厚度、腐殖质厚度)、干扰等方面,如康冰[4]、郑金萍[5]、张希彪[6]、白志强[7]等研究表明林分密度、坡向、海拔、腐殖质及凋落物厚度等对幼树幼苗更新有显著影响,马姜明[8]、杨文云[9]等研究得出母树密度、倒木蓄积量、胸高断面积、灌木层及草本层盖度等因子对天然更新均有不同程度的影响,Xu[10]、Zhu[11]、Gallegos[12]等探讨了干扰、林隙及林下植被等对森林更新再生的作用,等等。从现有研究看,多数研究采用比较分析和简单相关分析的方法,如Yu[13]等比较分析了坡位、坡向等对天然更新的影响,而农友[14]、白登忠[15]、Chai[16]等运用相关分析法探讨了天然更新与环境因子之间的关系,但用综合方法研究多因子对天然更新影响的报道极为少见。
闽楠(Phoebe bournei (Hemsl.) Yang)为樟科常绿乔木,现为国家二级珍稀渐危种,在福建、江西、湖南等海拔1 000 m以下的山地常绿阔叶林中有零星分布。对闽楠天然次生林的地理分布、空间结构及种间竞争[17-19]等方面有较多研究,但对其自然更新影响因子研究尚为缺乏。因此,本文通过综合分析闽楠幼树幼苗自然更新与其环境因子之间的关系,阐明其自然更新的主要影响因子,为促进自然更新经营措施的制订提供参考依据,这对闽楠天然次生林经营与保护具有重要的现实意义。
-
研究区域位于江西省中部吉安市(113°48′~115°56′ E,25°58′~27°57′ N),地貌以丘陵、山地为主,土壤类型主要有红壤、黄壤等,母岩以砂岩、花岗岩为主;属亚热带季风湿润气候,年平均气温17.1~18.6℃,年均降水量1 360~1 577 mm,降水主要集中在春季和初夏,年无霜期281 d。森林资源丰富,主要植被类型有针叶林、常绿阔叶林、针阔混交林和竹林等,森林覆盖率达到67.6%。灌木以木(Loropetalum chinense (R. Br.) Oliv)、杜茎山(Maesa japonica (Thunb.) Moritzi.)为主,草本以铁芒萁(Dicranopteris linearis (Burm.f.) Underw.)、庐山楼梯草(Elatostema stewardii Merr)为主。
-
根据森林资源二类调查资料及当地林业部门的了解,在研究区的安福县、遂川县、井冈山市的闽楠天然次生林分布地进行踏查,选择人为干扰程度轻且在分布地具有代表性的地块设置标准地,标准地面积依据其分布地形等因素而定,为400 m2(20 m×20 m)或600 m2(20 m×30 m),共计标准地23块,并采用相邻网格调查方法,将每块标准地划分为4或6个10 m×10 m小样方,共有100个小样方。以每个小样方为调查单元,调查胸径≥2 cm的所有乔木的林木位置(x、y坐标)、树种、胸径、树高等;更新层的调查在10 m×10 m小样方内记录胸径 < 2 cm的幼苗(H < 1 m)、幼树(1 m < H < 3 m)[20-21]的树种、地径、树高、冠幅等;林下植被在10 m×10 m小样方的上、中、下分别选择具有代表性的样方,灌木样方调查为2 m×2 m,草本样方调查为1 m×1 m,调查记录灌木和草本的种类、数量、高度、盖度等,同时在1 m×1 m小样方内记录凋落物层的盖度和厚度;土壤质地、土层厚度、腐殖质层厚度等均在做土壤剖面时记录;利用GPS测定标准地的地理位置和海拔,同时记录坡位、坡向、坡度和郁闭度等因子。
-
数量化模型Ⅰ的数学原理是建立某个因子和数量因子或非数量因子之间的线性关系,公式如下:
$ Y_{t}=B_{0}+\sum\limits_{i=1}^{m} \sum\limits_{j=1}^{n_{i}} B(i, j) s_{t}(i, j)+\sum\limits_{i=m+1}^{p} B(i) x_{t i} $
其中,Yt为第t点因变量值;B0为常数项;B(i, j)为第i定性因子第j等级的得分;st(i, j)为第t点第i定性因子第j等级的反应值(或0或1);B(i)为第i定量因子的回归系数;xti为第t点第i项定量因子的观测值;ni为第i定性因子的等级数。为衡量各项目在模型中的贡献,计算各项目的得分范围,得分范围越大,则该项目影响就越大。
重要值计算目前应用较多的是Curtis等修改的重要值指数,是相对密度或相对多度、相对频度和相对显著度3项指数的综合。采用下式计算闽楠幼树幼苗的重要值[22]:
重要值=相对密度(%)+相对频度(%)+相对盖度(%)
其中,相对密度=(某种植物的个体数/所有种的个体数)×100%;相对频度=(某种植物的频度/所有种的频度总和)×100%;相对盖度=(某种植物的盖度/所有种盖度总和)×100%。
重要值作为因变量,影响林下植被更新状况的环境因子为自变量,通过多元数量化模型Ⅰ建模,分析各环境因子对闽楠天然次生林闽楠幼树幼苗更新的影响,在此基础上,对影响极显著的定性因子进行控制变量分析,即控制其它影响因子相似的情况下,采用单因素方差分析(one-way ANOVA)方法进一步分析某个因子变化下闽楠幼树幼苗更新状况(平均密度、平均高度)的差异程度。所有建模过程、图表及数据分析过程在SPSS 19.0和Excel 2016中完成。
-
参考相关文献[23-24]及结合闽楠天然次生林的特征,选择影响闽楠幼树幼苗自然更新的环境因子并对其进行分解。环境因子包括林分因子(平均胸径、郁闭度和株数密度)、地形因子(坡度、坡位和坡向)、林下植被因子(林下植被盖度、林下植被高度)、土壤因子(土壤厚度、腐殖质层厚度)、凋落物层厚度以及闽楠的状况(闽楠下种母树株数、闽楠密度和闽楠平均胸径)等14个因子。需要说明的是,由于标准地的海拔在200~400 m范围,跨度小,土壤类型均为红壤,其土壤质地等均相差不大,因此没有考虑海拔及其它土壤因子;此外,考虑到能结实的闽楠数量的多少可能对其幼树幼苗更新影响较大,因此,将结实量较为稳定的闽楠视为下种母树,参考文献[18]和闽楠生长特性[25],闽楠下种母树定义为胸径≥20 cm以上的树木。各环境因子的分解见表 1。
表 1 环境因子分解
Table 1. Decomposition of environmental factors
编号No 项目Item 类目Sub-item 1 2 3 4 1 坡位Position of slope 上坡 中坡 下坡 2 坡向Aspect of slope 阳坡 半阳坡 半阴坡 阴坡 3 坡度Slope gradient 缓坡615° 斜坡1625° 陡坡2535° 急坡3545° 4 林下植被盖度Coverage of understory vegetation/% < 0 3060 6090 >90 5 林下植被高度Hight of understory vegetation/m < 0.5 0.51.0 1.01.5 >1.5 6 郁闭度Canopy density < 0.3 0.30.5 0.50.7 >0.7 7 土层厚度Soil layer thickness/cm 薄(< 40) 中(4080) 厚(>80) 8 腐殖质层厚度Humus layer thickness/cm 薄(< 10) 中(1020) 厚(>20) 9 凋落物层厚度Litter layer thickness/cm 薄(< 1) 中(13) 厚(>3) 10 闽楠密度Density of Phoebe bournei/(N·hm-2) < 500 5 001 000 1 0001 500 >1 500 11 株数密度Strains density/(N·hm-2) < 1 000 1 0002 000 2 0003 000 >3 000 12 闽楠下种母树株数Number of Phoebe bournei parent tree/N 13 闽楠平均胸径Average DBH of Phoebe bournei/cm 14 林分平均胸径Average DBH /cm 以闽楠幼树幼苗的重要值为因变量,选取的环境因子(包括定性和定量项目)为自变量,运用多元化数量模型Ⅰ,对14个环境因子进行建模,根据结果进行偏相关检验,把相关系数小的,差异性不显著的项目剔除掉,再对剩下的项目重新建模,反复进行。考虑到减少建模因子的数量,才能增加模型的准确性,最后筛选出闽楠下种母树株数、郁闭度、坡位、腐殖质层厚度、坡向、林下植被盖度、凋落物层厚度、株数密度8个环境因子,其模型如下:
Y=2.284-3.416x11-2.042x12+0.245x13-1.709x21-0.686x22+0.423x31+0.384x32-0.171x33-1.488x41-1.388x42+6.022x51+1.341x52-0.131x61+0.023x62+0.291x63-0.152x71-0.094x72+0.104x73+0.605x8
在模型检验中,偏相关系数采用t检验,其结果均为极显著或显著;复相关系数用F检验,检验结果为极显著,所以筛选出的8个因子与闽楠天然次生林幼树幼苗自然更新具有显著或极显著的关系。模型运算结果及各类目得分值和项目分值极差见表 2。
表 2 模型运算结果
Table 2. Operational result of the model
项目Item 项目代号Item code 类目代号Sub-item code 得分值Score 分值极差Score range 偏相关系数Partial correlation coefficient t检验t test 坡向Aspect of slope x1 1 -3.416 3.661 0.397 5 2.947 2** 2 -2.042 3 0.245 4 0 坡位Position of slope x2 1 -1.709 1.709 0.602 4 5.196 4** 2 -0.686 3 0 林下植被盖度Coverage of understory vegetation x3 1 0.423 0.423 0.365 1 2.772 1** 2 0.384 3 0.171 4 0 腐殖质层厚度Humus layer thickness x4 1 -1.488 1.488 0.584 8 4.971 3** 2 -1.388 3 0 凋落物层厚度Litter layer thickness x5 1 6.022 6.022 0.359 2 2.718 6** 2 1.341 3 0 郁闭度Canopy density x6 1 -0.131 0.422 0.650 2 5.846 6** 2 0.023 3 0.291 4 0 株数密度Strains density x7 1 -0.152 0.256 0.325 7 2.307 4* 2 -0.094 3 0.104 4 0 闽楠下种母树株树Number of Phoebe bournei parent tree x8 0.605 0.605 0.715 7 6.843 5** 注:*表示在0.05水平上显著相关,**表示在0.01水平上显著相关;t0.01(59)=2.662;t0.05(59)=2.001;F0.01(8, 59)=2.829;复相关系数为0.966;F检验结果为极显著(F=30.96* *)
Note: *indicates a significant correlation at the 0.05 level,** indicates a significant correlation at the 0.01 level;t0.01(59)=2.662;t0.05(59)=2.001;F0.01(8, 59)=2.829;The multiple correlation coefficient was 0.966;F test was extremely significant (F=30.96* *) -
从表 2可知,闽楠下种母树株数对闽楠幼树幼苗更新的贡献率最大。自然更新必须有种源,而闽楠下种母树结实比较稳定,可为更新提供种源,对其幼树幼苗的更新起最主要的作用,大树越多,提供的种子越多,越有利于自然更新。因此必需保证林分中有一定数量的闽楠下种母树,这是其自然更新的基础。通过对标准地调查数据的分析,并参考闽楠种子萌发[26]及其幼树幼苗生长竞争特性[25],要保证其天然更新的顺利进行,闽楠下种母树应保留200株·hm-2 以上。
郁闭度对闽楠幼树幼苗更新影响极显著,其影响程度排名第2位,其中以0.5~0.7的郁闭度最好,其次是0.3~0.5和>0.7, < 0.3最差。进一步通过单因素方差分析表明(表 3),郁闭度0.5~0.7下闽楠幼树幼苗的平均密度显著高于其他郁闭度(P < 0.05),而郁闭度为0.3~0.5的幼树幼苗平均高度显著高于郁闭度 < 0.3和>0.7的平均高度(P < 0.05)。林冠层郁闭度的大小直接决定林下层光照的情况。郁闭度过小,会导致光照强度增大,引起土壤湿度变小,不利于种子的萌发;郁闭度过大,会导致光照强度过弱,不利于幼树幼苗生长。
表 3 不同郁闭度对幼树幼苗更新状况影响
Table 3. Effect of different canopy density on natural regeneration of sapling
郁闭度Canopy density < 0.3 0.30.5 0.50.7 >0.7 幼树幼苗平均密度Sapling average density/(N·hm-2) 789±126b 2 133±208b 6 533±1 973a 1 633±305b 幼树幼苗平均高度Sapling average height/m 0.46±0.23b 1.110±0.35a 0.710±0.01ab 0.550±0.21b 注:同行不同小写字母代表差异显著(P < 0.05),下同。
Note: Data with different lowercase letters in the same line are highly significantly different(P < 0.05). The same below.株数密度以2 000~3 000株·hm-2最好,其次是3 000~4 000株·hm-2和1 000~2 000株·hm-2, < 1 000株·hm-2更新最差。模型运算虽然得出这种结果,但由于株数密度的多少主要是影响光照条件,而在天然林中株数密度对林内光照的影响程度,还与株数按径级的分配比例有很大关系。因此,考虑到株数密度与郁闭度之间存在一定的关系,在经营过程中主要可采用调控郁闭度大小的措施,对郁闭度过大的结合径级结构的调整适当间伐掉一些次要树种,而对郁闭度过小的适当补植树种以提高郁闭度。
-
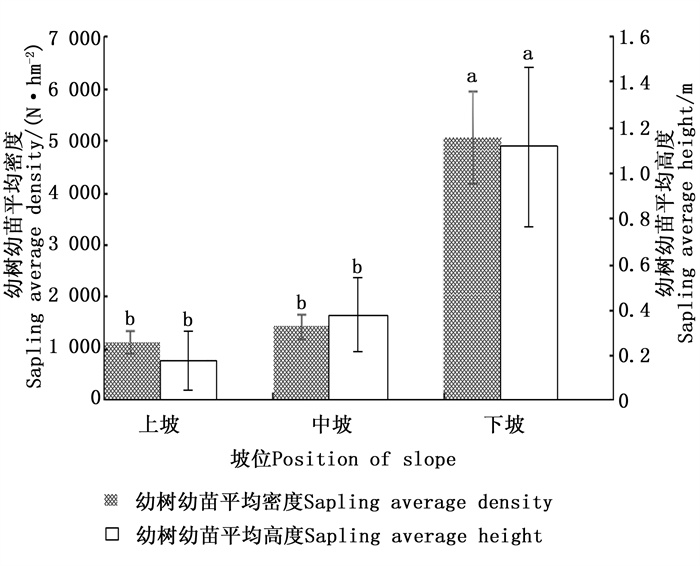
根据表 2结果,坡位对幼树幼苗更新特征的影响排名第3位,从得分值来看,下坡位闽楠幼树幼苗更新最好,其次是中坡,最差的是上坡。图 1单因素方差分析结果表明,下坡幼树幼苗的平均密度和平均高度与中坡、上坡之间均存在显著差异(P < 0.05)。这主要是因为下坡土壤的持水性能高,长期的地表径流作用会导致下坡土壤更为肥沃,下坡土壤更有利于乔木种群的繁殖、种子的萌发以及幼苗种群的扩展,且雨水冲刷导致种子冲到下坡,下坡种子库较上坡丰富,因此,下坡更新更好。而上坡位土壤流失水分多,光照比下坡强,同时养分含量具有抑制效应,不利于种子的萌发和幼树幼苗的生长。
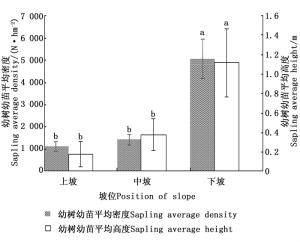
图 1 不同坡位对幼树幼苗更新状况的影响
Figure 1. Effect of different position of slope on natural regeneration
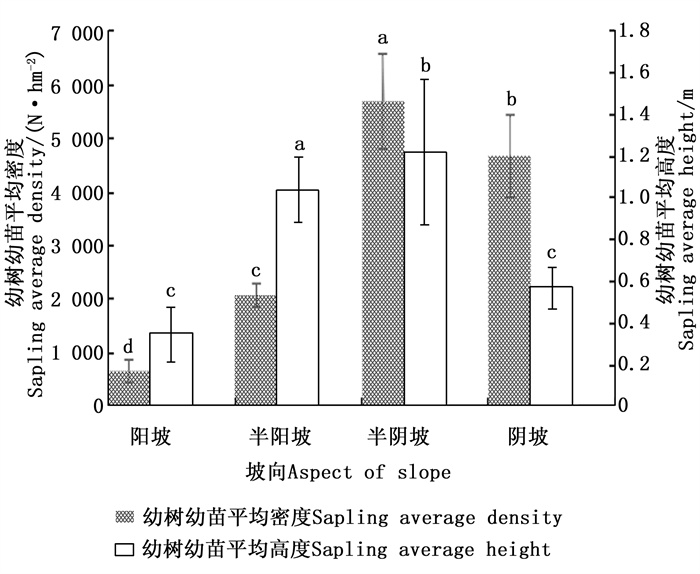
坡向以半阴坡最好,其次是阴坡、半阳坡,阳坡最差。通过单因素方差分析(图 2)可知,阳坡、半阳坡、半阴坡、阴坡闽楠幼树幼苗的平均密度之间的差异均显著(P < 0.05),其中半阴坡闽楠幼树幼苗的平均密度最大,且其平均高度也显著高于其他坡向(P < 0.05)。不同坡向太阳的辐射强度以及日照时数不同,导致水、热和土壤理化性质有所差异。半阴坡和阴坡林分内太阳辐射及日照少导致温度降低和蒸腾作用减小,土壤水分充足,对种子萌发和幼树幼苗生长起促进作用。而阳坡光照强,导致温度升高和水分蒸腾增大,不利于幼树幼苗生长。闽楠是耐阴性树种,对光热的耐受条件有限。因此,对闽楠幼树幼苗来说,半阴坡最适合闽楠更新。
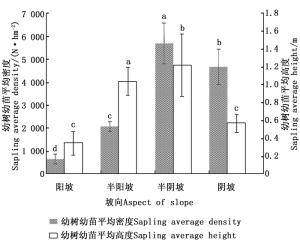
图 2 不同坡向对幼树幼苗更新状况的影响
Figure 2. Effect of different aspect of slope on natural regeneration
-
腐殖质层厚度对闽楠次生林下幼树幼苗更新的影响排在第4位。其中,以>20 cm最好,10~20 cm次之, < 10 cm最差。从单因素方差分析(表 4)可以看出,腐殖质层厚度>20 cm的闽楠幼树幼苗的平均密度和平均高度均显著高于其他腐殖质层厚度的平均密度和平均高度(P < 0.05)。由于土壤中的水分、营养物质等影响着种子的萌发、幼苗的生长和发育,而幼树幼苗的根系较浅,加上腐殖质层的养分、微量元素较为丰富,因此腐殖质层厚度越厚越有利于幼树幼苗的更新。
表 4 不同腐殖质厚度对幼树幼苗更新状况影响
Table 4. Effect of different humus layer thickness on natural regeneration of sapling
腐殖质层厚度Humus layer thickness /cm < 10 1020 >20 幼树幼苗平均密度Sapling average density/(N·hm-2) 1 167±88c 2 620±375b 4 720±712a 幼树幼苗平均高度Sapling average height/m 0.99±0.12b 1.20±0.19b 1.49±0.28a 凋落物层厚度以 < 1 cm最好,其次是1~3 cm,最差是>3 cm。单因素方差分析(表 5)可以看出,凋落物层厚度 < 1 cm闽楠幼树幼苗的平均密度和平均高度均显著高于其他凋落物层厚度的平均密度和平均高度(P < 0.05)。这是由于较厚的凋落物对种子和土壤接触影响较大,不利于种子到达土壤表面,减少了种子萌发的可能性和幼苗定居的机会;同时,凋落物也可能会通过化感作用等抑制幼苗的更新。因此,经营中可通过人为措施对凋落物层厚度进行控制,以促进其更新。
表 5 不同凋落物厚度对幼树幼苗更新影响
Table 5. Effect of different litter layer thickness on natural regeneration of sapling
凋落物层厚度Litter layer thickness /cm < 1 13 >3 幼树幼苗平均密度Sapling average density/(N·hm-2) 4 913±893a 2 320±753b 1 500±169b 幼树幼苗平均高度Sapling average height/m 1.39±0.09a 0.98±0.19b 0.95±0.08b -
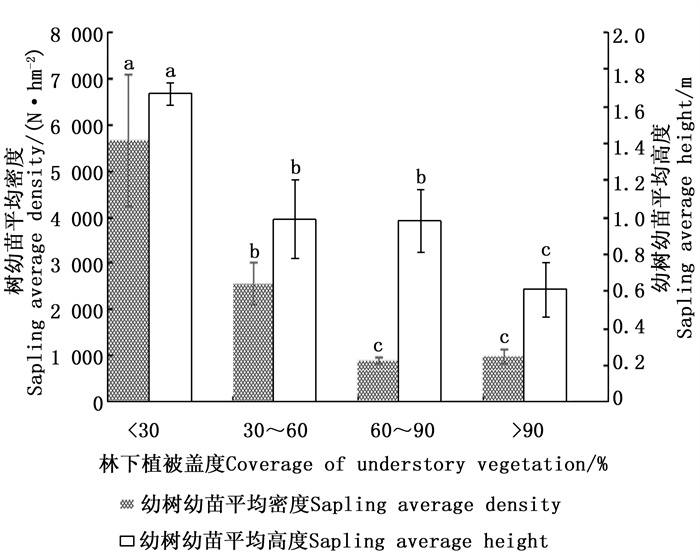
林下植被盖度以 < 30%最好,其次是30%~60%、60%~90%,最差是>90%,从单因素方差分析(图 3)可知,林下植被盖度小于 < 30%的幼树幼苗平均密度和平均高度均显著高于其他林下植被盖度的平均密度和平均高度(P < 0.05)。林下植被盖度的高低直接影响林下幼树幼苗生长空间的大小,其盖度越大林下幼树幼苗与灌草对光照、水分、养分等资源的竞争越强,从而影响幼树幼苗的自然更新。因此在闽楠天然次生林经营中,进行砍杂抚育将有利于其自然更新。
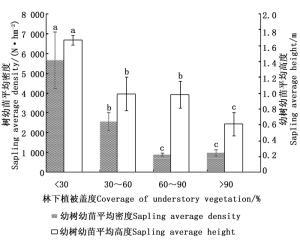
图 3 不同林下植被盖度对幼树幼苗更新状况影响
Figure 3. Effect of different understory vegetation coverage on natural regeneration of sapling
-
在更新过程中,土壤种子库是天然更新的基础[27],母树的数量和母树的繁殖能力对幼树幼苗的更新有着重要的作用,吴大荣[25]等研究表明:闽楠更新很大程度上依赖于种子产量,且大径级株数多的种群中其幼树幼苗种子库最丰富,本研究也得出闽楠下种母树株数(结实量较稳定的闽楠)对闽楠幼树幼苗更新的贡献率最大。因此,保证林分中有一定数量的闽楠下种母树对其自然更新至关重要。
林冠层可通过改变土壤和温度条件[28]以及降低风速和减少蒸腾[29]等为幼树幼苗提供适宜的环境,因此,郁闭度会影响幼树幼苗的更新和生长。本研究得出郁闭度为0.5~0.7时自然更新状况最好,而刘明国[24]等对油松的研究认为0.3~0.5的郁闭度下油松更新更好,这可能是由于闽楠和油松有着不同的生物学特性所造成的。同样,林分密度直接导致林下光、热、水分、土壤理化特性等生态因子的异质性,并直接影响到林下幼树幼苗的生长,林分密度过低会抑制大量中生偏阴生性优势植物幼苗的萌发和生长,过大则林内光照强度减弱有助于幼苗的生长却增大了种内和种间的竞争[30],本研究表明密度在2 000~3 000株·hm-2时,闽楠幼树幼苗更新的状况最好,而郑金萍[5]等对蒙古栎的研究认为林分密度在400~800株·hm-2更新数量最多,造成这种差异的原因可能是株数按径级的分配比例不一样,且闽楠属于阴性树种,而蒙古栎属于阳性树种,它们对光照的要求不同。此外,坡向也影响着光照条件[31],本研究得出半阴坡闽楠幼树幼苗更新状况更好,主要是由闽楠的生物学特性所决定的。
对于坡位及腐殖质层来说,由上坡到下坡,土壤由剥蚀逐渐过渡到堆积,腐殖质层由薄变厚,土壤质地由粗变细,土壤含水量增加,因此,下坡位、厚腐殖质更有助于更新。而对于凋落物层厚度,不少研究均表明其对更新有较大的影响,如张树梓等[32]研究表明天然更新苗状况与凋落物厚度呈负相关,本研究结论与之相一致,这主要是因为凋落物持续积累增厚会对植被的自然更新形成阻碍,表现在物理的机械阻挡、化学的他感作用、生物的动物侵害和微生物致病作用等方面[33]。此外,相关研究也表明,林下植被被认为是决定群落结构和动态的重要因素[34],但林下植被盖度过大,会加大林下更新树种与灌草对光照、水分、养分等资源的争夺[35],从而不利于自然更新,Pham Van Huong[36]等研究结果及本研究结果均验证了这一点。
-
影响闽楠天然次生林自然更新的因子主要有密度(闽楠下种母树株数、郁闭度、株数密度)、坡位、腐殖质层厚度、坡向、林下植被盖度、凋落物层厚度。
密度对闽楠天然次生林自然更新影响最大。其中,闽楠下种母树株数起着最主要的作用,其株数多有利于自然更新,而郁闭度与株数密度过高或过低均不利于其自然更新,郁闭度在0.5~0.7之间其幼树幼苗的平均密度均显著高于郁闭度 < 0.3、0.3~0.5和>0.7的(P < 0.05)。凋落物层越厚、林下植被盖度越大均不利于其自然更新,凋落物层厚度 < 1 cm最好,最差是>3 cm,厚度 < 1 cm的幼树幼苗平均密度和平均高度均显著高于厚度1~3 cm及>3 cm的(P < 0.05);林下植被盖度以 < 30%更新最好,其次是30%~60%、60%~90%,最差是>90%,其 < 30%盖度的幼树幼苗平均密度和平均高度均显著高于其他林下植被盖度(P < 0.05)。腐殖质层越厚越有利于闽楠的自然更新,腐殖质层厚度>20 cm的闽楠幼树幼苗的平均密度和平均高度均显著高于腐殖质层厚度 < 10 cm及10~20 cm的(P < 0.05)。地形对更新的影响主要体现在坡位和坡向,下坡位闽楠幼树幼苗更新最好,最差为上坡,下坡幼树幼苗的平均密度和平均高度均显著高于中坡和上坡(P < 0.05);坡向则以半阴坡最好,阳坡最差,半阴坡幼树幼苗的平均密度和平均高度显著高于其他坡向(P < 0.05)。
为促进闽楠幼树幼苗的自然更新,林分中闽楠下种母树应保留200株·hm-2以上,同时,对郁闭度、林下植被盖度及凋落物层厚度等实施相应的调控措施。
闽楠天然次生林自然更新的影响因子研究
Study on Factors Affecting Natural Regeneration of Natural Secondary Phoebe bournei Forest
-
摘要:
目的 分析闽楠天然次生林自然更新与环境因子之间的关系,为其科学经营及保护提供参考依据。 方法 以江西省吉安市闽楠天然次生林为研究对象,通过标准地调查获取数据,运用多元数量化模型Ⅰ建立闽楠幼树幼苗重要值与环境因子的关系模型,采用单因素方差分析单个环境因子对闽楠幼树幼苗更新的影响。 结果 研究表明:密度(闽楠下种母树株数、郁闭度、株数密度)、坡位、腐殖质层厚度、坡向、林下植被盖度、凋落物层厚度是影响闽楠天然次生林自然更新的主要因素,偏相关系数在0.325 7~0.715 7之间,t检验结果为极显著或显著;模型复相关系数为0.966,F检验结果为极显著(F=30.96**)。 结论 闽楠下种母树株数对其幼树幼苗的更新起着最主要的作用,而郁闭度与株数密度过高或过低、凋落物层越厚、腐殖质层越薄、林下植被盖度越大均不利于其自然更新,同时,半阴坡、下坡位条件下的闽楠幼树幼苗的更新好于其它坡向与坡位。为促进闽楠幼树幼苗的自然更新,林分中闽楠下种母树应保留200株·hm-2以上,以及对郁闭度、林下植被盖度及凋落物层厚度等实施相应的调控措施。 Abstract:Objective To analyze the relationships between natural regeneration of Phoebe bournei and environmental factors in order to provide references for protecting and managing natural secondary P. bournei forest. Method Based on data of sample plot investigation, a relationship model between saplings and seedlings importance values and environmental factors was established by quantitative model Ⅰ, and the effect of single environmental factor on natural regeneration was explored by using one-way ANOVA method in Ji'an Country, Jiangxi Province. Result The results showed that the main elements affecting natural regeneration were density (amount of P. bournei parent tree, canopy density, tree density), position of slope, humus layer thickness, aspect of slope, coverage of understory vegetation, litter layer thickness, with the partial correlation coefficients being 0.325 7-0.715 7, which were significant or very significant by t test. The multiple correlation coefficient of the model reached 0.966, showing very significant by F test (F=30.96**). Conclusion The amount of P. bournei parent tree plays an important role in natural regeneration of saplings and seedlings, while higher or lower of canopy density and tree density, thicker litter layer, thinner humus layer, higher coverage of understory vegetation are not helpful to natural regeneration. Meanwhile, natural regeneration of saplings and seedlings is better in semi-shady slope and lower slope than other aspect of slope and position of slope. Therefore, keeping at least 200 parent trees per hectare in the stand and controlling canopy density, coverage of understory vegetation, thickness of litter layer would be helpful to promote natural regeneration of saplings and seedlings. -
Key words:
- Phoebe bournei
- / natural secondary forest
- / natural regeneration
- / influencing factor
-
图 1 不同坡位对幼树幼苗更新状况的影响
Figure 1. Effect of different position of slope on natural regeneration
图 2 不同坡向对幼树幼苗更新状况的影响
Figure 2. Effect of different aspect of slope on natural regeneration
图 3 不同林下植被盖度对幼树幼苗更新状况影响
Figure 3. Effect of different understory vegetation coverage on natural regeneration of sapling
表 1 环境因子分解
Table 1. Decomposition of environmental factors
编号No 项目Item 类目Sub-item 1 2 3 4 1 坡位Position of slope 上坡 中坡 下坡 2 坡向Aspect of slope 阳坡 半阳坡 半阴坡 阴坡 3 坡度Slope gradient 缓坡615° 斜坡1625° 陡坡2535° 急坡3545° 4 林下植被盖度Coverage of understory vegetation/% < 0 3060 6090 >90 5 林下植被高度Hight of understory vegetation/m < 0.5 0.51.0 1.01.5 >1.5 6 郁闭度Canopy density < 0.3 0.30.5 0.50.7 >0.7 7 土层厚度Soil layer thickness/cm 薄(< 40) 中(4080) 厚(>80) 8 腐殖质层厚度Humus layer thickness/cm 薄(< 10) 中(1020) 厚(>20) 9 凋落物层厚度Litter layer thickness/cm 薄(< 1) 中(13) 厚(>3) 10 闽楠密度Density of Phoebe bournei/(N·hm-2) < 500 5 001 000 1 0001 500 >1 500 11 株数密度Strains density/(N·hm-2) < 1 000 1 0002 000 2 0003 000 >3 000 12 闽楠下种母树株数Number of Phoebe bournei parent tree/N 13 闽楠平均胸径Average DBH of Phoebe bournei/cm 14 林分平均胸径Average DBH /cm  下载: 导出CSV
下载: 导出CSV
表 2 模型运算结果
Table 2. Operational result of the model
项目Item 项目代号Item code 类目代号Sub-item code 得分值Score 分值极差Score range 偏相关系数Partial correlation coefficient t检验t test 坡向Aspect of slope x1 1 -3.416 3.661 0.397 5 2.947 2** 2 -2.042 3 0.245 4 0 坡位Position of slope x2 1 -1.709 1.709 0.602 4 5.196 4** 2 -0.686 3 0 林下植被盖度Coverage of understory vegetation x3 1 0.423 0.423 0.365 1 2.772 1** 2 0.384 3 0.171 4 0 腐殖质层厚度Humus layer thickness x4 1 -1.488 1.488 0.584 8 4.971 3** 2 -1.388 3 0 凋落物层厚度Litter layer thickness x5 1 6.022 6.022 0.359 2 2.718 6** 2 1.341 3 0 郁闭度Canopy density x6 1 -0.131 0.422 0.650 2 5.846 6** 2 0.023 3 0.291 4 0 株数密度Strains density x7 1 -0.152 0.256 0.325 7 2.307 4* 2 -0.094 3 0.104 4 0 闽楠下种母树株树Number of Phoebe bournei parent tree x8 0.605 0.605 0.715 7 6.843 5** 注:*表示在0.05水平上显著相关,**表示在0.01水平上显著相关;t0.01(59)=2.662;t0.05(59)=2.001;F0.01(8, 59)=2.829;复相关系数为0.966;F检验结果为极显著(F=30.96* *)
Note: *indicates a significant correlation at the 0.05 level,** indicates a significant correlation at the 0.01 level;t0.01(59)=2.662;t0.05(59)=2.001;F0.01(8, 59)=2.829;The multiple correlation coefficient was 0.966;F test was extremely significant (F=30.96* *)
下载: 导出CSV
表 3 不同郁闭度对幼树幼苗更新状况影响
Table 3. Effect of different canopy density on natural regeneration of sapling
郁闭度Canopy density < 0.3 0.30.5 0.50.7 >0.7 幼树幼苗平均密度Sapling average density/(N·hm-2) 789±126b 2 133±208b 6 533±1 973a 1 633±305b 幼树幼苗平均高度Sapling average height/m 0.46±0.23b 1.110±0.35a 0.710±0.01ab 0.550±0.21b 注:同行不同小写字母代表差异显著(P < 0.05),下同。
Note: Data with different lowercase letters in the same line are highly significantly different(P < 0.05). The same below.
下载: 导出CSV
表 4 不同腐殖质厚度对幼树幼苗更新状况影响
Table 4. Effect of different humus layer thickness on natural regeneration of sapling
腐殖质层厚度Humus layer thickness /cm < 10 1020 >20 幼树幼苗平均密度Sapling average density/(N·hm-2) 1 167±88c 2 620±375b 4 720±712a 幼树幼苗平均高度Sapling average height/m 0.99±0.12b 1.20±0.19b 1.49±0.28a
下载: 导出CSV
表 5 不同凋落物厚度对幼树幼苗更新影响
Table 5. Effect of different litter layer thickness on natural regeneration of sapling
凋落物层厚度Litter layer thickness /cm < 1 13 >3 幼树幼苗平均密度Sapling average density/(N·hm-2) 4 913±893a 2 320±753b 1 500±169b 幼树幼苗平均高度Sapling average height/m 1.39±0.09a 0.98±0.19b 0.95±0.08b
下载: 导出CSV
-
[1] Nanami S, Kawaguchi H, Tateno R, et al. Sprouting traits and population structure of co-occurring Castanopsis species in an evergreen broad-leaved forest in southern China[J]. Ecological Research, 2004, 19: 341-348. doi: 10.1111/j.1440-1703.2004.00643.x [2] Zhu J J, Matsuzaki T, Li F Q, et al. Effects of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest[J]. Forest Ecology and Management, 2003, 182: 339-354. doi: 10.1016/S0378-1127(03)00094-X [3] 程真, 周光益, 吴仲民, 等. 南陵南坡中段不同群落林下幼树的生物多样性及分布[J]. 林业科学研究, 2015, 28(4): 543-550. doi: 10.3969/j.issn.1001-1498.2015.04.014 [4] 康冰, 王得祥, 李刚, 等. 秦岭山地锐齿栎次生林幼苗更新特征[J]. 生态学报, 2012, 32(9): 2738-2747. [5] 郑金萍, 杨学东, 郭忠玲, 等. 蒙古栎林天然更新状况及影响因素研究[J]. 北华大学学报: 自然科学版, 2015, 16(5): 652-657. [6] 张希彪, 上官周平, 王金成, 等. 子午岭人工油松林群落更新特征及影响因子[J]. 山地学报, 2014, 32(5): 561-567. doi: 10.3969/j.issn.1008-2786.2014.05.006 [7] 白志强, 刘华, 张新平, 等. 新疆额尔齐斯河流域杨树幼苗天然更新影响因子分析[J]. 西北林学院学报, 2011, 26(1): 98-102. [8] 马姜明, 刘世荣, 史作民, 等. 川西亚高山暗针叶林恢复过程中岷江冷杉天然更新状况及其影响因子[J]. 植物生态学报, 2009, 33(4): 646-657. doi: 10.3773/j.issn.1005-264x.2009.04.003 [9] 杨文云. 滇中地区云南松天然林群落结构及天然更新规律[D]. 北京: 中国林业科学研究院, 2010. [10] Xu H, Li Y D, Liu S R, et al. Partial recovery of a tropical rain forest a half-century after clear-cut and selective logging[J]. Journal of Applied Ecology, 2015, 52(4): 1044-1052. doi: 10.1111/1365-2664.12448 [11] Zhu J J, Lu D L, Zhang W D. Effects of gaps on regeneration of woody plants: a meta-analysis[J]. Journal of Forestry Research, 2014, 25(3): 501-510. doi: 10.1007/s11676-014-0489-3 [12] Gallegos S C, Beck S G, Hensen I, et al. Factors limiting montane forest regeneration in bracken-dominated habitats in the tropics[J]. Forest Ecology and Management, 2016, 381: 168-176. doi: 10.1016/j.foreco.2016.09.014 [13] Yu F, Wang D X, Shi X X, et al. Effects of environmental factors on tree seedling regeneration in a pine-oak mixed forest in the Qinling Mountains, China[J]. Journal of Mountain Science, 2013, 10(5): 845-853. doi: 10.1007/s11629-013-2548-1 [14] 农友, 卢立华, 孙冬婧, 等. 岩溶石山降香黄檀人工林的天然更新[J]. 中南林业科技大学学报, 2017, 37(3): 63-68. [15] 白登忠. 祁连山青海云杉林线树木生长、更新的影响因素研究[D]. 北京: 中国林业科学研究院, 2012. [16] Chai Z Z, Wang D X. Environmental influences on the successful regeneration of pine-oak mixed forests in the Qinling Mountains, China[J]. Scandinavian Journal of Forest Research, 2016, 31(4): 368-381. doi: 10.1080/02827581.2015.1062912 [17] 葛永金, 王军峰, 叶荣华, 等. 闽楠地理分布格局及其气候特征研究[J]. 江西农业大学学报, 2012, 34(4): 749-753. doi: 10.3969/j.issn.1000-2286.2012.04.022 [18] 游晓庆, 彭诗涛, 郭孝玉, 等. 闽楠天然次生林林分空间结构研究[J]. 江西农业大学学报, 2016, 38(4): 660-667. [19] 刘宝, 陈存及, 陈世品, 等. 福建明溪闽楠天然林群落种间竞争的研究[J]. 福建林学院学报, 2005, 25(2): 117-120. doi: 10.3969/j.issn.1001-389X.2005.02.005 [20] 于倩, 谢宗强, 熊高明, 等. 神农架巴山冷杉(Abies fargesii)林群落特征及其优势种群结构[J]. 生态学报, 2008, 28(5): 1931-1941. doi: 10.3321/j.issn:1000-0933.2008.05.006 [21] 于大炮, 周莉, 董百丽, 等. 长白山北坡岳桦种群结构及动态分析[J]. 生态学杂志, 2004, 23(5): 30-34. doi: 10.3321/j.issn:1000-4890.2004.05.006 [22] 王育松, 上官铁梁. 关于重要值计算方法的若干问题[J]. 山西大学学报: 自然科学版, 2010, 33(2): 312-316. [23] 欧阳勋志, 廖为明, 彭世揆. 天然阔叶林景观质量评价及其垂直结构优化技术[J]. 应用生态学报, 2007, 18(6): 1388-1392. doi: 10.3321/j.issn:1001-9332.2007.06.036 [24] 刘明国, 殷有, 孔繁轼, 等. 辽西半干旱地区油松人工林天然更新的影响因子研究[J]. 沈阳农业大学学报, 2014, 45(4): 418-423. doi: 10.3969/j.issn.1000-1700.2014.04.007 [25] 吴大荣, 朱政德. 福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究[J]. 林业科学, 2003, 39(1): 23-30. doi: 10.3321/j.issn:1001-7488.2003.01.004 [26] 吴大荣, 王伯荪. 濒危树种闽楠种子和幼苗生态学研究[J]. 生态学报, 2001, 21(11): 1751-1760. doi: 10.3321/j.issn:1000-0933.2001.11.002 [27] Santos D M, Santos J M, Silva K A, et al. Composition, species richness, and density of the germinable seed bank over 4 years in young and mature forests in Brazilian semiarid regions[J]. Journal of Arid Environments, 2016, 129: 93-101. doi: 10.1016/j.jaridenv.2016.02.012 [28] Pastur G J M, Cellini J M, Lencinas M V, et al. Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention[J]. Forest Ecology and Management, 2011, 261(1): 178-186. doi: 10.1016/j.foreco.2010.10.002 [29] Promis A, Caldentey J, Ibarra M. Microclima en el interior de un bosque de Nothofagus pumilio y el efecto de una corta de regeneración[J]. Bosque (Valdivia), 2010, 31(2): 129-139. [30] Aussenac G. Interactions between forest stands and microclimate: ecophysiological aspects and consequences for silviculture[J]. Annals of Forest Science, 2000, 57(3): 287-301. doi: 10.1051/forest:2000119 [31] Bennie J, Huntley B, Wiltshire A, et al. Slope aspect and climate; Spatially explicit and implicit models of topographic microclimate in chalk grassland[J]. Ecological Modelling, 2008, 216(1): 47-59. doi: 10.1016/j.ecolmodel.2008.04.010 [32] 张树梓, 李梅, 张树彬, 等. 塞罕坝华北落叶松人工林天然更新影响因子[J]. 生态学报, 2015, 35(16): 5403-5411. [33] 李霄峰, 胥晓, 王碧霞, 等. 小五台山森林落叶层对天然青杨种群更新方式的影响[J]. 植物生态学报, 2012, 36(2): 109-116. [34] Taylor A H, Jinyanb H, Zhou S. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China: a 12-year study[J]. Forest Ecology and Management, 2004, 200, 347-360. doi: 10.1016/j.foreco.2004.07.007 [35] Zhang H T, Tao J P, Wang L, et al. Influences of herbaceous vines on community characteristics in pioneer succession stages[J]. Acta Ecologica Sinica, 2011, 31(4): 186-191. doi: 10.1016/j.chnaes.2011.03.005 [36] Pham Van Huong, 陈昌雄, 张巧巧, 等. 灌草对胖大海种群更新幼树幼苗密度的影响[J]. 西南林业大学学报, 2016, 36(4): 1-8. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5117
- HTML全文浏览量: 1093
- PDF下载量: 1169
- 被引次数: 0
