

-
作为我国二级保护物种之一的毛红椿(Toona ciliata Roem.var.pubescens(Franch.)Hand.-Mazz.[1],因其速生、珍贵、濒危,近年来越来越受到学者关注。目前,在毛红椿天然种群生态特征[2]、群落结构特征[3]、群体遗传结构[4-5]、结实特性及生殖力[6-7]、种子萌发的影响因素[8-9]和毛红椿资源保护对策和育苗技术[10-12]等方面开展了研究,本课题组探讨了天然林毛红椿种子库、林下幼苗数量、基株个体大小和枯落物厚薄等生境因素对毛红椿天然更新的影响[2, 6, 9]。植物种群更新限制主要来自三种机制:一是种子可获得性低,导致有效传播率低,引起种源限制;二是种子不能扩散到合适地点而受到传播限制;三是扩散到微生境后,受生物和非生物因素影响而不能生长成幼树,导致建成限制[13-15]。微生境生态因子包含多种限制因素,如土壤、水分、光照、枯落物、植物邻居等,这些因子决定着幼树的建成,不同种植物的主导因子存在差异[16]。土壤作为林木生长的重要生态因子,其物理、化学及生物学性质直接影响森林的更新过程[17],森林群落植物的组成及分布,还与其森林系统根系分泌物的化感效应存在密切联系[18]。本文在毛红椿天然优势群落中,研究根区土壤、土壤真菌及土壤性质等对种子萌发和幼苗建成的影响,研究结果有助于寻找制约该物种生存和发展的主要因子,更好理解毛红椿天然更新的障碍因素,为珍稀阔叶树种毛红椿致濒提供有效的保护策略和保育技术。
-
2015年3月,在江西九连山国家级自然保护区大丘田、电站和虾公塘等3个毛红椿天然林群落,以较小径级基株受邻体干扰的有效距离10.0 m为限[2],在距离毛红椿成年大树2.5、5.0、7.5 m树根附近采集除去枯落物0~10 cm表层土壤作为根区土,以远离大树25 m以外的土壤作为非根区土带回实验室,每个群落采取3株毛红椿9个根区土样和3个非根区土样,共36个土样,每个土样采集2.0 kg,分成3份,分别用于播种、土壤理化分析和微生物检测。以新鲜河沙、沙壤土、菜地土为对照,其中,沙壤土为细沙与黄壤土按7∶3均匀混合。供试种子采自九连山毛红椿天然林群落母树,千粒质量7.345 g。3个群落及母树基本情况见表 1。
表 1 九连山毛红椿天然林群落及母树基本情况
Table 1. Geographic location and the mother trees of nature populations T.ciliata var.pubescens stands in Jiulianshan
群落
Stand地形
Geomorphic types经度(E)
Longitude纬度(N)
Latitude海拔
Altitude/m毛红椿相对密度
Relative density of Toona ciliate var. pubescens/%母树平均高度
Mean height of the mother tree/m母树平均胸径
DHB of the mother tree/cm大丘田Daqiutian 山谷Valley 114°26′ 24°34′ 501 37.93 23.5 40.0 电站Dianzhan 山坡Hillside 114°30′ 24°36′ 575 49.22 21.0 29.8 虾公塘Xiagongtang 山谷Valley 114°27′ 24°31′ 760 22.22 22.5 31.9 -
2015年4月,用0.3%的高锰酸钾溶液进行种子消毒30 min,再用蒸馏水清洗残液。将根区土、非根区土及河沙、沙壤土、菜地土分别装入经消毒灭菌的组培瓶(瓶高85 mm,上口径95 mm,底面直径80 mm),每瓶置20粒种子,放入光照培养箱发芽,光照时间14 h·d-1,温度25℃,每天称质量补充水分,5次重复。观察记录种子发芽数、发芽天数及幼苗保存情况,以某处理中幼苗全部死亡结束整个实验,计算毛红椿种子发芽率和幼苗存活率。
种子发芽率=种子发芽数/种子数×100%
幼苗存活率=实验结束时存活幼苗数/种子发芽数×100%
-
采用烘干法(105℃)测定土壤含水率,玻璃电极法测定土壤pH值,水合热重铬酸钾氧化-比色法测定土壤有机质含量,KCl浸提-靛酚蓝比色法测定土壤铵态氮含量,紫外分光光度法测定土壤硝态氮含量,3, 5-二硝基水杨酸比色法测定土壤纤维素酶和蔗糖酶活性[19]。
-
选取播种实验幼苗死亡率较高的毛红椿根区土样,分离培养真菌。称取5 g根区土,梯度稀释法制成土壤悬液,接种至ABPDA培养基(PDA培养基内加100 mg·L-1链霉素和100 mg·L-1青霉素),28℃恒温培养获得单菌落,采用划线分离法纯化菌株,得到根区土壤真菌,再将其接种至PDA培养基斜面上保存待用。
-
挖取1年生毛红椿幼苗3株,洗净根系泥土,在盛有1 000 mL蒸馏水的容器中放置24 h,收集水培液,过滤除菌获得根系分泌物(R)[20]。
-
将消毒处理后的毛红椿种子播于121℃间歇灭菌3次的沙壤土中,光照培养箱内培养15 d至初生叶形成,以备接种之用。
-
从分离出的6株真菌RS1、RS2、RS3、RS4、RS5、RS6中随机选取3株真菌(RS2、RS3、RS5),接种至PDA培养基平皿进行活化培养后,加入适量灭菌水,洗脱孢子,用4层灭菌擦镜纸过滤除去菌丝及杂质。采用血球计数板计数,用无菌水(W)和根系分泌物(R)分别将供试真菌孢子配成浓度为1 × 107个·L-1的无菌水真菌孢子悬液(RSW)和根系分泌物真菌孢子悬浮液(RSR),以无菌水和根系分泌物为对照,均用10 mL对毛红椿幼苗进行灌根接种,8种处理(RS2W、RS3W、RS5W、RS2R、RS3R、RS5R及W、R),3次重复,每重复20株幼苗。置28℃黑暗处理24 h后,置于光照14 h·d-1、温度25℃、相对湿度80%~90%的培养箱内培养,期间补充相应的无菌水或根系分泌物,连续观察15 d,记录菌丝和幼苗生长情况,计算幼苗存活率。
-
采用SPSS 17.0统计软件和Excel 2007进行数据分析,不同处理间参数的差异采用Student-Newman-Keuls法进行多重比较,比较两参数采用独立样本t检验,变量相关性采用双变量pearson相关分析。
-
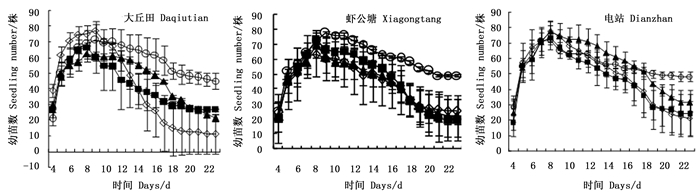
在不同群落土壤中,毛红椿种子的萌发趋势基本一致,即第4天开始萌发,第8~10天达到高峰,第10天全部种子发芽结束。幼苗数呈先上升后下降的趋势,林地根区土幼苗数下降速度比非根区土的快,18 d以后非根区土幼苗数均比根区土的高(图 1)。

图 1 毛红椿种子萌发及幼苗数量随时间的变化
Figure 1. Changes on the number of seed germination and seedling of T. ciliata var. pubescens with the sowing time
-
表 2表明:种子发芽率为62.33%~77.67%,幼苗保存率为15.70%~ 67.14%,各群落2.5、5.0、7.5 m处根区土毛红椿幼苗保存率均低于非根区土。方差分析表明:大丘田、电站和虾公塘3个群落间及不同距离根区间种子发芽率的差异均不显著;大丘田、电站和虾公塘不同距离土壤间幼苗保存率差异显著(F=10.222,P=0.004;F=4.172,P=0.047;F=4.794,P=0.034)。多重比较结果(表 2)表明:大丘田和虾公塘群落2.5、5.0、7.5 m处根区土的毛红椿幼苗保存率均显著低于非根区土的;电站2.5 m处根区土的幼苗保存率显著低于非根区土的,2.5、5.0、7.5 m处根区土间的幼苗保存率差异不显著;2.5、5.0、7.5 m处根区土的毛红椿幼苗保存率总均值间差异不显著,但与非根区土间差异极显著(F=15.135,P=0.000<0.01)。以上结果表明:毛红椿根区土、非根区土种子发芽率差异不显著,而根区土与非根区土间幼苗保存率差异显著,说明根区土对毛红椿幼苗保存有较大影响;2.5、5.0、7.5 m处根区土毛红椿种子发芽率及幼苗保存率总均值间差异不显著,说明在距母树一定周径范围内根区土对毛红椿种子发芽和幼苗保存影响一致。
表 2 距毛红椿母树不同距离的种子发芽率与幼苗保存率
Table 2. Seedling survival rate and seed germination rate of T.ciliata var. pubescens in different distances from the mother tree
离母树距离
Distances from the mother tree发芽率seed germination rate/% 保存率seedling survival rate/% 大丘田
Daqiutian虾公塘
Xiagongtang电站
Dianzhan总均值
Total mean大丘田
Daqiutian虾公塘
Xiagongtang电站
Dianzhan总均值
Total mean2.5 m根区2.5 m root zone 77.00±6.25a 63.33±14.98a 77.67±7.09a 72.67±11.29a 15.70±18.24c 38.84±4.85 b 27.82±14.35b 27.45±15.52B 5.0 m根区5.0 m root zone 66.33±5.13a 73.67±6.35 a 73.67±7.64a 71.22±6.47a 40.89±3.91b 25.77±16.19b 34.54±22.59ab 33.73±15.50B 7.5 m根区7.5 m root zone 62.33±4.93a 70.33±9.87a 77.67±6.51a 70.11±9.23a 35.00±5.32bc 32.52±20.08b 40.92±10.99ab 36.14±12.33B 非根区Non-root zone 71.00±9.85a 77.33±1.51a 72.00±2.65a 73.44±5.94a 64.03±9.42a 63.80±0.61a 67.14±3.88a 64.99±5.35A 注:同列不同小写字母间表示差异显著(P<0.05),同列不同大写字母间表示差异极显著(P<0.01),下同。
Note:Different lowercase letter along the column indicate significant difference at P<0.05,Different capital letter along the column indicate extremely significant difference at P<0.01,the same as below.表 3表明:林地土(根区土、非根区土)种子发芽率的总均值(71.86%)与非林地土(河沙、沙壤土、菜地土)对照的种子发芽率总均值(72.67%)差异不显著,但林地土的保存率总均值(40.58%)极显著低于非林地土的(70.02%)(F=5.352,df=49,P=0.000<0.01);林地根区土与非根区土的种子发芽率差异不显著,但幼苗保存率差异极显著(F=30.074,P=0.000<0.01),林地根区土幼苗存活率较低,仅32.44%。以上结果进一步表明,林地土毛红椿幼苗保存率低的原因主要来源于根区土的影响。
表 3 不同类型土壤毛红椿种子发芽率与幼苗保存率
Table 3. Seedling survival rate and seed germination rate of T.ciliata var. pubescens in different soils
土壤类型Soil type 发芽率Seed germination rate/% 保存率Seedling survival rate/% 均值Mean 总均值Total mean 均值Mean 总均值Total mean 林地土Forest soil 根区土Root zone soil 71.33±8.96a 71.86±8.78a 32.44±14.45B 40.58±19.13B 非根区土Non-root zone soil 73.44±5.94a 64.99±5.35A 非林地土Non-forest soil 河沙River sand 62.00±7.58b 72.67±9.29a 74.74±11.04A 70.02±5.43A 沙壤土Sandy soil 77.00±7.58a 64.09±22.31A 菜地土Vegetable soil 79.00±10.25a 71.24±5.44A -
鉴于非根区土壤幼苗保存率与3种非林地土均差异不显著,故探讨林地土壤(根区土和非根区土)性状与毛红椿幼苗保存率间的关系。
九连山毛红椿林地土部分理化性质和酶活性见表 4。独立样本t检验结果表明:毛红椿根区土的pH值显著高于非根区土的(F=0.049,df=34,P=0.016<0.05),但毛红椿根区土和非根区土的含水率、蔗糖酶、纤维素酶、有机质、铵态氮、硝态氮差异不显著。
表 4 九连山林地土壤的理化性质和酶活性
Table 4. Enzyme activity and physical-chemical properties of Jiulianshan forest soil
土壤性状
Soil traits根区Root zone 非根区Non-root zone 大丘田Daqiutian 虾公塘Xiagongtang 电站Dianzhan 大丘田Daqiutian 虾公塘Xiagongtang 电站Dianzhan 含水率Water content/% 32.57±1.42 34.97±3.82 26.34±3.84 37.67±8.72 39.24±3.65 28.60±3.00 pH值pH value 5.39±0.29 5.40±0.22 6.09±0.14 5.11±0.29 4.90±1.83 5.68±0.36 有机质SOM/(g·kg-1) 15.16±5.44 10.56±0.48 7.40±2.34 17.00±4.37 11.36±0.51 9.36±0.54 蔗糖酶Invertase/(mg·g-1) 4.34±3.24 3.17±0.99 5.61±3.81 6.40±2.37 5.33±3.66 3.72±2.98 纤维素酶Cellulose/(mg·g-1) 1.38±0.86 1.92±0.57 1.41±0.85 7.86±0.79 2.38±0.19 0.95±0.26 铵态氮(NH4+)N/(mg·kg-1) 13.60±5.86 32.55±2.82 21.01±10.33 21.13±11.03 33.13±1.96 16.71±7.43 硝态氮(NO3-)N/(mg·kg-1) 1.61±0.61 1.63±0.28 1.34±0.39 1.86±0.65 1.71±0.40 1.13±0.42 相关分析结果(表 5)表明:不同类型土壤指标与种子发芽率和幼苗保存率相关不显著。
表 5 九连山林地土壤理化性质、酶活性与种子发芽率和幼苗保存率相关分析
Table 5. Correlation of seed germination rate, seedling survival rate and soil physical-chemical properties, enzyme activity of Jiulianshan forest soil
项目Items 相关分析
Correlation含水率
Water contentpH值
pH value有机质
SOM蔗糖酶
Invertase纤维素酶
Cellulose铵态氮
(NH4+)N硝态氮
(NO3-)N发芽率
Seed germination rate皮尔森相关系数Pearson Correlation 0.127 0.033 -0.063 0.210 -0.022 0.054 -0.038 显著性Sig. 0.462 0.848 0.716 0.218 0.900 0.758 0.830 样本数N 36 36 36 36 36 35 35 保存率
Seedling survival rate皮尔森相关系数Pearson Correlation 0.038 -0.107 -0.104 0.114 0.031 0.037 -0.070 显著性Sig. 0.824 0.536 0.546 0.509 0.858 0.832 0.687 样本数N 36 36 36 36 36 35 35 -
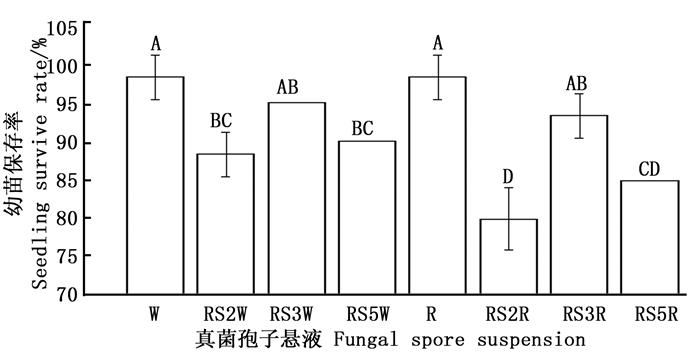
根区土死亡的幼苗表现为苗茎或其叶子着生处感染褐化干枯而死,疑似真菌感染所致。因此,从幼苗死亡率较高的根区土壤中分离出真菌(RS2和RS3来自大丘田群落距毛红椿大树2.5 m处根区土壤,RS5来自虾公塘群落距毛红椿大树5 m处根区土壤),分别用其水(RS2W、RS3W、RS5W)或毛红椿根系分泌物孢子悬液(RS2R、RS3R、RS5R)对毛红椿健康幼苗灌根接种,无菌水(W)和根系分泌物(R)为对照,幼苗保存结果见图 2。
图 2 根区土壤真菌孢子悬浮液对毛红椿幼苗保存率的影响
Figure 2. Effect of the soil-borne fungal spore suspension on the survival of T. ciliata var. pubescens seedlings
方差分析结果表明:处理间差异极显著(F=20.802,df=7,P=0.000<0.01)。多重比较表明:2个对照间差异不显著,无菌水悬浮液RS2W、RS5W和根系分泌物悬浮液RS2R、RS5R处理的幼苗保存率均显著低于其相应的对照;RS3W和RS3R孢子悬浮液处理的幼苗保存率与无菌水(W)和根系分泌物(R)的差异不显著;RS2R处理的幼苗保存率显著低于RS2W处理的,RS5R处理的幼苗保存率低于RS5W处理的,但差异不显著(图 2)。以上结果表明:RS2和RS5真菌可能为毛红椿潜在致病菌,且毛红椿根系分泌物能显著增强RS2真菌的致病力。
-
在林木更新过程中,更新幼苗的发生和存活往往与同种成年树的位置有关[21-22]。本研究结果表明,距离毛红椿母树2.5~7.5 m内根区土的幼苗存活率差异不显著,但显著低于距毛红椿成年大树较远的非根区土的幼苗存活率。10 m是小径级毛红椿受邻体干扰较强的范围[2],说明毛红椿种内的邻体干扰亦来自母树影响范围内的土壤,但根区土壤的理化性状与远离同种大树的非根区土壤差异并不显著。McCarthy-Neumann S等[22]认为,成年母树附近的低光照强度、存在高浓度的化感物质及高的取食压力和聚集的病原菌等可能是导致其幼苗死亡率偏高的原因。本模拟实验是在室内相同光照条件下进行,不存在取食压力问题,幼苗根系分泌物及根区真菌灌根培养结果表明,根系分泌物单独作用并未影响幼苗存活率,但与某些菌同时作用却显著降低幼苗存活率。因此,母树地下根部是病原菌的仓库,而树下有限的光照条件更利于病原菌的繁殖,推测毛红椿大树根区土壤病原菌可能是影响其幼苗建成的一个重要因素。
森林土壤中的真菌及细菌等有较大的空间变异性[23],土壤中病原菌对土壤的改变,将不利于植物的生存和生长,特别是对原有植物种的扩展产生限制作用,在幼苗更新过程中,这种限制作用表现为距同种成年树越近,幼苗死亡率越高。研究已证实,土壤中病害真菌导致热带和亚热带森林中成年树附近幼苗死亡,这可能是由土传病原菌引起的植物-土壤的负反馈作用对植物丰度的调节[24-26]。近年来,越来越多的学者认为,根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素[27]。由于根系分泌物的输入,根际沉积物有不同于非根际沉积物的化学、物理和微生物特性[28]。植物根系周围的特殊区域对土壤中的微生物而言,是一个天然的选择性培养基, 如果大量繁殖的微生物为病原菌,则可能入侵植物根系,导致病害发生[27, 29]。刘艳霞等[30]研究表明,烟草根系分泌物及其酚酸类成分促进烟草青枯病原菌(茄科劳尔氏菌)的生长,而对拮抗菌(短芽孢杆菌)的生长具有抑制作用。某些根系分泌物成分对植物病原菌的毒性物质分泌具有促进作用,如Wu等[31]研究发现,兰州百合根系分泌物成分邻苯二甲酸显著刺激百合枯萎病原菌(尖孢镰刀菌)发病相关水解酶的活性。关于根系分泌物和土壤病原菌对毛红椿实生更新的影响还有待进一步研究。
-
在本研究的不同类型土壤中,毛红椿种子萌发规律基本一致,幼苗数均呈先上升后下降的趋势,且毛红椿林地土壤理化性状与酶活性等均与种子发芽率及幼苗保存率不显著相关,但根系分泌物能够影响某些病原菌的致病性,使毛红椿根区土壤中存在潜在致病真菌导致幼苗死亡,不利于其幼苗保存和建成。建议下一步应侧重于致病微生物鉴定、对根系分泌物趋化性和其成分分析,以期提高毛红椿幼苗存活率,促进幼苗建成。
林地土壤及其真菌对毛红椿种子发芽及幼苗保存的影响
Effects of Forest Soil and Soil-borne Fungi on Seed Germination and Seedling Survival of Toona ciliata var. pubescens
-
摘要:
目的 通过不同群落毛红椿天然林林地土壤及其真菌对种子萌发和幼苗存活影响的研究,探讨影响毛红椿天然更新的障碍因素。 方法 在江西九连山国家级自然保护区内的3个毛红椿天然优势群落中,分别取距离毛红椿母树3个不同距离(2.5、5.0、7.5 m)处的根区土壤和远离毛红椿母树(25 m以外)的非根区土壤,以非林地土为对照开展室内模拟播种和以根区土壤真菌的水或根系分泌物悬液对毛红椿幼苗进行灌根接种,观察种子萌发、幼苗存活和幼苗感病情况。 结果 各群落土壤中种子发芽均呈先上升后下降规律,且基本在第8~10天达到高峰;林地土与非林地土种子发芽率差异不显著,但林地根区土幼苗的存活率极显著低于非林地土和林地非根区土;距母树3个不同距离根区土的幼苗存活率差异不显著。灌根接种不同处理间幼苗存活率差异极显著,具体表现为接种RS2、RS5根区真菌的幼苗感病率显著高于根系分泌物及无菌水空白对照,且RS2真菌的根系分泌物悬浮液幼苗感病率显著高于其无菌水悬浮液。 结论 毛红椿种子萌发不受土壤环境的影响,但幼苗建成受根区范围内土壤的影响,根系分泌物和致病菌的互作显著降低幼苗保存率。由此推断,毛红椿根区土壤内存在幼苗的潜在致病菌及根系分泌物可增强其致病性。 Abstract:Objective The barrier factors influencing the natural regeneration of Toona ciliata var. pubescens were explored through investigating the effects of forest soil and fungi in different natural communities on seed germination and seedling survival. Method Soil samples were collected from the root zone at three distances (2.5, 5.0, and 7.5 m) from the mother trees as well as the natural forest stands 25 m away from the mother trees in Jiulianshan National Nature Reserve. The seed germination trial on soils was carried out in the laboratory with non-forest soil as the control. In addition, the seedling infection trial in soil was also performed in the laboratory with root irrigation by fungi isolated from root zone soil and suspended with water or root exudates. The seed germination, seedling survival and seedling susceptibility were observed. Result The germination of seed in all kinds of soil revealed the tendency of increasing at initial and then decreasing, and reached the peak during the eighth to tenth day after sowing. There were no significant differences between forest soil and non-forest soil in germination rate, while the seedling survival rate of forest soil was significantly lower than that of non-forest soil. There were significant differences between root zone soil and non-root zone soil in seedling survival rate, and the seedling mortality of root zone soil was higher. It was also found that there were no significant differences in seedlings survival rate of root zone soil among different distances from the mother trees. It addition, there were extremely significant differences in seedling infection treatments, and the seedling disease incidence rates of RS2, RS5 soil-borne fungi were significantly higher than those of the two blank controls, water (W) and root exudates (R). Moreover, the disease infection rate of RS2 root exudates suspension was significantly higher than that of RS2 water suspension. Conclusion Forest soil of different natural communities has no effect on seed germination of T. ciliata var. pubescens, However, the seedling establishment is disturbed by root zone soil of the same species; the interaction between root exudates and pathogens can significantly reduce the seedling survival rate. It could be inferred that there are potential pathogens in root zone soil of T. ciliata var. pubescens and root exudates can enhance their pathogenicity. -
Key words:
- Toona ciliata var. pubescens
- / seed germination
- / seedling survival
- / soil-borne fungi
-
图 1 毛红椿种子萌发及幼苗数量随时间的变化
Figure 1. Changes on the number of seed germination and seedling of T. ciliata var. pubescens with the sowing time
图 2 根区土壤真菌孢子悬浮液对毛红椿幼苗保存率的影响
Figure 2. Effect of the soil-borne fungal spore suspension on the survival of T. ciliata var. pubescens seedlings
表 1 九连山毛红椿天然林群落及母树基本情况
Table 1. Geographic location and the mother trees of nature populations T.ciliata var.pubescens stands in Jiulianshan
群落
Stand地形
Geomorphic types经度(E)
Longitude纬度(N)
Latitude海拔
Altitude/m毛红椿相对密度
Relative density of Toona ciliate var. pubescens/%母树平均高度
Mean height of the mother tree/m母树平均胸径
DHB of the mother tree/cm大丘田Daqiutian 山谷Valley 114°26′ 24°34′ 501 37.93 23.5 40.0 电站Dianzhan 山坡Hillside 114°30′ 24°36′ 575 49.22 21.0 29.8 虾公塘Xiagongtang 山谷Valley 114°27′ 24°31′ 760 22.22 22.5 31.9  下载: 导出CSV
下载: 导出CSV
表 2 距毛红椿母树不同距离的种子发芽率与幼苗保存率
Table 2. Seedling survival rate and seed germination rate of T.ciliata var. pubescens in different distances from the mother tree
离母树距离
Distances from the mother tree发芽率seed germination rate/% 保存率seedling survival rate/% 大丘田
Daqiutian虾公塘
Xiagongtang电站
Dianzhan总均值
Total mean大丘田
Daqiutian虾公塘
Xiagongtang电站
Dianzhan总均值
Total mean2.5 m根区2.5 m root zone 77.00±6.25a 63.33±14.98a 77.67±7.09a 72.67±11.29a 15.70±18.24c 38.84±4.85 b 27.82±14.35b 27.45±15.52B 5.0 m根区5.0 m root zone 66.33±5.13a 73.67±6.35 a 73.67±7.64a 71.22±6.47a 40.89±3.91b 25.77±16.19b 34.54±22.59ab 33.73±15.50B 7.5 m根区7.5 m root zone 62.33±4.93a 70.33±9.87a 77.67±6.51a 70.11±9.23a 35.00±5.32bc 32.52±20.08b 40.92±10.99ab 36.14±12.33B 非根区Non-root zone 71.00±9.85a 77.33±1.51a 72.00±2.65a 73.44±5.94a 64.03±9.42a 63.80±0.61a 67.14±3.88a 64.99±5.35A 注:同列不同小写字母间表示差异显著(P<0.05),同列不同大写字母间表示差异极显著(P<0.01),下同。
Note:Different lowercase letter along the column indicate significant difference at P<0.05,Different capital letter along the column indicate extremely significant difference at P<0.01,the same as below.
下载: 导出CSV
表 3 不同类型土壤毛红椿种子发芽率与幼苗保存率
Table 3. Seedling survival rate and seed germination rate of T.ciliata var. pubescens in different soils
土壤类型Soil type 发芽率Seed germination rate/% 保存率Seedling survival rate/% 均值Mean 总均值Total mean 均值Mean 总均值Total mean 林地土Forest soil 根区土Root zone soil 71.33±8.96a 71.86±8.78a 32.44±14.45B 40.58±19.13B 非根区土Non-root zone soil 73.44±5.94a 64.99±5.35A 非林地土Non-forest soil 河沙River sand 62.00±7.58b 72.67±9.29a 74.74±11.04A 70.02±5.43A 沙壤土Sandy soil 77.00±7.58a 64.09±22.31A 菜地土Vegetable soil 79.00±10.25a 71.24±5.44A
下载: 导出CSV
表 4 九连山林地土壤的理化性质和酶活性
Table 4. Enzyme activity and physical-chemical properties of Jiulianshan forest soil
土壤性状
Soil traits根区Root zone 非根区Non-root zone 大丘田Daqiutian 虾公塘Xiagongtang 电站Dianzhan 大丘田Daqiutian 虾公塘Xiagongtang 电站Dianzhan 含水率Water content/% 32.57±1.42 34.97±3.82 26.34±3.84 37.67±8.72 39.24±3.65 28.60±3.00 pH值pH value 5.39±0.29 5.40±0.22 6.09±0.14 5.11±0.29 4.90±1.83 5.68±0.36 有机质SOM/(g·kg-1) 15.16±5.44 10.56±0.48 7.40±2.34 17.00±4.37 11.36±0.51 9.36±0.54 蔗糖酶Invertase/(mg·g-1) 4.34±3.24 3.17±0.99 5.61±3.81 6.40±2.37 5.33±3.66 3.72±2.98 纤维素酶Cellulose/(mg·g-1) 1.38±0.86 1.92±0.57 1.41±0.85 7.86±0.79 2.38±0.19 0.95±0.26 铵态氮(NH4+)N/(mg·kg-1) 13.60±5.86 32.55±2.82 21.01±10.33 21.13±11.03 33.13±1.96 16.71±7.43 硝态氮(NO3-)N/(mg·kg-1) 1.61±0.61 1.63±0.28 1.34±0.39 1.86±0.65 1.71±0.40 1.13±0.42
下载: 导出CSV
表 5 九连山林地土壤理化性质、酶活性与种子发芽率和幼苗保存率相关分析
Table 5. Correlation of seed germination rate, seedling survival rate and soil physical-chemical properties, enzyme activity of Jiulianshan forest soil
项目Items 相关分析
Correlation含水率
Water contentpH值
pH value有机质
SOM蔗糖酶
Invertase纤维素酶
Cellulose铵态氮
(NH4+)N硝态氮
(NO3-)N发芽率
Seed germination rate皮尔森相关系数Pearson Correlation 0.127 0.033 -0.063 0.210 -0.022 0.054 -0.038 显著性Sig. 0.462 0.848 0.716 0.218 0.900 0.758 0.830 样本数N 36 36 36 36 36 35 35 保存率
Seedling survival rate皮尔森相关系数Pearson Correlation 0.038 -0.107 -0.104 0.114 0.031 0.037 -0.070 显著性Sig. 0.824 0.536 0.546 0.509 0.858 0.832 0.687 样本数N 36 36 36 36 36 35 35
下载: 导出CSV
-
[1] 傅立国. 中国植物红皮书(第1册)[M]. 北京: 科学出版社, 1991: 142-669. [2] 杨清培, 付方林, 张露, 等. 九连山自然保护区毛红椿天然林邻体干扰效应[J]. 江西农业大学学报, 2013, 35(4): 748-754. doi: 10.3969/j.issn.1000-2286.2013.04.015 [3] 刘军, 陈益泰, 罗阳富, 等. 毛红椿天然林群落结构特征研究[J]. 林业科学研究, 2010, 16(1): 93-97 [4] 刘军, 陈益泰, 孙宗修, 等. 基于空间自相关分析研究毛红椿天然居群的空间遗传结构[J]. 林业科学, 2008, 44(6): 45-52. doi: 10.3321/j.issn:1001-7488.2008.06.009 [5] 温强, 叶金山, 周诚. 江西毛红椿天然群体的遗传多样性分析[J]. 南方林业科学, 2016, 44(3): 1-6, 55. [6] 黄红兰, 张露, 廖承开. 毛红椿天然林种子雨、种子库与天然更新[J]. 应用生态学报, 2012, 23(4): 972-978. [7] 黄红兰, 张露, 郭晓燕, 等. 九连山毛红椿种群的结实特性及其生殖力[J]. 林业科学, 2013, 49(7): 170-174. [8] 张丽, 张露. 毛红椿种子萌发影响因素初探[J]. 林业科技开发, 2011(6): 51-56. doi: 10.3969/j.issn.1000-8101.2011.06.014 [9] 郭晓燕, 张露, 梁跃龙, 等. 枯枝落叶物厚度影响毛红椿种子出苗的模拟研究[J]. 江西农业大学学报, 2014, 36(2): 332-337. [10] 张汝忠, 彭佳龙, 王坚娅, 等. 毛红椿播种育苗技术及苗期生长规律研究[J]. 浙江林业科技, 2007, 27(4): 51-53. doi: 10.3969/j.issn.1001-3776.2007.04.012 [11] 孙洪刚, 刘军, 董汝湘, 等. 水分胁迫对毛红椿幼苗生长和生物量分配的影响[J]. 林业科学研究, 2014, 27(3): 381-387. [12] 唐强, 李志辉, 吴际友, 等. 毛红椿无性系扦插繁殖试验研究[J]. 中南林业科技大学学报, 2015, 35(1): 67-70. [13] Münzbergová Z, Herben T. Seed, dispersal, microsite, habitat and recruitment limitation: identification of terms and concepts in studies of limitations[J]. Oecologia, 2005, 145(1): 1-8. doi: 10.1007/s00442-005-0052-1 [14] Cordeiro N J, Ndangalasi H J, McEntee J P, et al. Disperser limitation and recruitment of an endemic African tree in a fragmented landscape[J]. Ecology, 2009, 90(4): 1030-1041. doi: 10.1890/07-1208.1 [15] Marques M C M, Burslem D F R P. Multiple stage recruitment limitation and density dependence effects in two tropical forests[J]. Plant Ecology, 2015, 216(9): 1243-1255. doi: 10.1007/s11258-015-0505-5 [16] 李宁, 白冰, 鲁长虎. 植物种群更新限制—从种子生产到幼树建成[J]. 生态学报, 2011, 31(21): 6624-6632 [17] 曾思齐, 甘静静, 肖化顺, 等. 木荷次生林林木更新与土壤特征的相关性[J]. 生态学报, 2014, 34(15): 4242-4250. [18] 陈锋, 孟永杰, 帅海威, 等. 植物化感物质对种子萌发的影响及其生态学意义[J]. 中国生态农业学报, 2017, 25(1): 36-46. [19] 郑必昭. 土壤分析技术指南[M]. 北京: 中国农业出版社, 2012. [20] Khorassani R, Hettwer U, Ratzinger A, et al. Citramalic acid and salicylic acid in sugar beet root exudates solubilize soil phosphorus[J]. BMC Plant Biology, 2011, 11(1): 121-128. doi: 10.1186/1471-2229-11-121 [21] Comita L S, Queenborough S A, Murphy S J, et al. Testing predictions of the Janzen-Connell hypothesis: a meta-analysis of experimental evidence for distance- and density-dependent seed and seedling survival[J]. Journal of Ecology, 2014, 102: 845-856. doi: 10.1111/1365-2745.12232 [22] Mccarthy-Neumann S, Ibáñez I. Plant—soil feedback links negative distance dependence and light gradient partitioning during seedling establishment[J]. Ecology, 2013, 94(94): 780-786. [23] Morris S J. Spatial distribution of fungal and bacterial biomass in southern Ohio hardwood forest soils: fine scale variability and microscale patterns[J]. Soil Biology & Biochemistry, 1999, 31(10): 1375-1386. [24] Packer A, Clay K. Soil pathogens and spatial patterns of seedling mortality in a temperate tree[J]. Nature, 2000, 404(6775): 278-281. doi: 10.1038/35005072 [25] Bagchi R, Gallery R E, Gripenberg S, et al. Pathogens and insect herbivores drive rainforest plant diversity and composition[J]. Nature, 2014, 506(7486): 85-88. doi: 10.1038/nature12911 [26] Liu Y, Yu S, Xie Z, et al. Analysis of a negative plant-soil feedback in a subtropical monsoon forest[J]. Journal of Ecology, 2012, 100(4): 1019-1028. doi: 10.1111/j.1365-2745.2012.01953.x [27] 吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望[J]. 植物生态学报, 2014, 38 (3): 298-310. [28] Liu B, Liu X, Huo S, et al. Properties of root exudates and rhizosphere sediment of Bruguiera gymnorrhiza (L. )[J]. Journal of Soils & Sediments, 2016, 17(1): 1-11. [29] Qi J J, Yao H Y, Ma X J, et al. Soil microbial community composition and diversity in the rhizosphere of a Chinese medicinal plant. Communications in Soil[J]. Science and Plant Analysis, 2009, 40(9-10): 1462-1482. doi: 10.1080/00103620902818104 [30] 刘艳霞, 李想, 蔡刘体, 等. 烟草根系分泌物酚酸类物质的鉴定及其对根际微生物的影响[J]. 植物营养与肥料学报, 2016, 22(2): 418-428. [31] Wu Z, Yang L, Wang R, et al. In vitro study of the growth, development and pathogenicity responses of Fusarium oxysporum to phthalic acid, an autotoxin from Lanzhou lily[J]. World Journal of Microbiology and Biotechnology, 2015, 31(8): 1227-1234. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3481
- HTML全文浏览量: 807
- PDF下载量: 545
- 被引次数: 0
