






-
毛竹(Phyllostachys edulis (Carrière) J. Houz.)具有生长快、成材早、产量高、用途广、收益大等优点,是我国南方的重要森林资源。水分作为植物生长的重要条件,与毛竹生产力有着密切联系,因此干旱对毛竹林生产力具有重要潜在影响。毛竹忍耐或者逃避干旱生理生态策略值得深入研究探讨。叶片既是植物光合产物形成的主要场所,又是整株植物的水力瓶颈、应对灾难性水力失调的安全阀门[1]。叶片气孔通过调节开张度降低蒸腾来响应水力导度或叶片膨压的下降,进而避免植物水势变化导致的水力失调[2-3]。叶水势是反映植物体内水分亏缺最灵敏的生理指标[4]。其中凌晨叶水势可更好地反映植物的水分亏缺状况,与午后叶水势比较,有受大气状况变化的影响更小、较稳定(低标准差)、直接与土壤水分变化相关的优点[4-5]。由于凌晨叶水势变化稳定,可认为同一植物品种在相同生育阶段凌晨叶水势临界值也具有稳定性,其值在不同生育阶段的高低可表征植物对干旱胁迫的敏感性差异[5]。国内学者较多研究报道了作物在不同生育期的凌晨叶水势临界值及对应时期的土水势、土壤含水量指标等[5-6]。对不同水分状况下许多树木叶水势的研究表明,叶片水势与土壤含水量随干旱胁迫的发展而降低,但不同树种降低的趋势是不同的[6]。关于毛竹干旱胁迫的生理机制已有较多的研究[7],但在不同生长阶段干旱胁迫敏感性如何变化,干旱胁迫对毛竹不同生长季节和不同年龄个体叶水势影响的定量化研究仍较缺乏。仅袁佳丽等[8]研究探讨了毛竹从笋长成幼竹过程中不同年龄毛竹个体叶片水势变化差异,发现依靠水势,母竹将水分源源不断地供给新竹,以保证新竹健康快速生长,揭示水势是驱动毛竹快速生长的关键因素。
目前国内学者对干旱研究主要集中采用室内外盆栽苗半控水的试验方法,从植物水分生理生态、土壤水分动态、植物蒸腾耗水、林地水分平衡等方面进行了广泛的研究[4-7, 9]。野外人工截雨模拟干旱对树木进行研究的方法在国内使用较少[10],而在美国、非洲和东南亚的热带雨林地区应用较广泛[11-12]。本文通过毛竹野外人工截雨干旱试验,探讨毛竹凌晨水势对干旱胁迫的响应特征季节差异及其与无截雨自然生长状态毛竹水势变化的差异,阐明不同生长季节、不同年龄个体毛竹之间存在的水分供给关系,间接揭示毛竹体内水分可利用性的季节差异,旨在为全球气候背景下毛竹人工林可持续高效经营提供理论依据。
-
试验在国家林业局钱江源森林生态系统定位研究站庙山坞辅站进行,位于杭州西郊富阳区境内(119°56′~120°02′ E,30°03′~30°06′ N)。该区域地形属浙西低山丘陵区天目山系余脉,属于中亚热带季风气候区,年平均气温16.1℃(最高气温40.2℃,最低气温-14.4℃),多年平均降水量1 441.9 mm。土壤属于酸性红壤。
-
试验区毛竹林为20世纪60年代种植,处于粗放经营的自然更新林。样地坡度20°左右,坡向正南,海拔高度169 m。建立标准样地20 m×20 m,采用每木检测的方法分别测定样地内竹株的胸径和竹高。样地内竹林密度为3 875株·hm-2,郁闭度0.95,竹株胸径分布在4.0~13.6 cm之间,平均胸径为9.8 cm,平均竹高13.2 m。林下几乎没有灌木和草本,但地表覆盖一定的凋落物,平均厚度约2.0 cm。每2年劈山1次,不施肥,不翻耕,只采挖春笋。
-
于2014年7月下旬(毛竹停止发笋,新竹高生长完成后)在研究区人工毛竹林中选择具有代表性、大小为10 m×10 m的地块6个,对照样地和截雨处理干旱样地各3块,干旱样地和对照样地成对出现,并分别进行了样地本底调查。将干旱样地上稀疏的灌木去除,然后用顶棚法模拟截雨。具体做法:在截雨处理干旱样地内,为去除边缘效应、保证干旱面积为10 m×10 m,在距离地面上方1.5 m高度搭建PVC防水板材的截雨大棚(面积为11 m×11 m)进行截雨,用胶水粘合塑料布与毛竹结合处缝隙;为保证坡度、地形和林分条件与对照样地尽可能一致,大棚一边与等高线平行。在样地周围开挖深约50 cm、宽约20 cm的壕沟,并沿着壕沟埋人50 cm深的铁皮,壕沟内部同时铺上塑料薄膜,用于阻止水分从旁边渗入,并能更好地排水。对照样地只在周围开挖相似壕沟,不作其它处理。本次试验人工截雨干旱于2014年7月下旬开始,试验仍在持续中。
-
本研究于人工截雨干旱1 a后开始叶片水势测定,即分别于2015年8月、9月、10月、12月、2016年3月每月上旬选择晴朗无云的天气测定1次。在每块试验样地中分别选择长势一致、胸径大小大致相同的1年生至7年生的不同年龄毛竹各1株。借助野外临时搭建的梯子,分别针对各竹龄毛竹上、中、下3个冠层部位的向阳位置,用镰刀勾取阳生枝条,迅速将枝条插入水中,并在水下在距切口末端约3 cm处用修枝剪再次快速一次性剪切,以防止枝条失水,保证离体叶片尽可能少失水。然后根据测定时间,依次选取水中每个枝条的中部位置长势基本相同,且形状完整、健康成熟、叶龄一致的叶片8~10片迅速带回室内进行凌晨水势的测定。其中最上部8~10档为上冠层,基部第1~2档枝条为下冠层。为保证叶龄一致,尽量摘取色泽一致和在枝上着生位置尽可能相同的叶片来测定。
毛竹叶片水势具体测定方法如下:选取晴朗无云的天气,于每天凌晨5:00—5:30太阳升起之前取样,迅速用冰壶带回实验室,采用PSYPRO水势测量系统(美国Wescor公司)进行毛竹离体叶片凌晨水势测定。每个处理取3片叶片分别测定,作为3个重复测定值,最后取平均值。叶水势测定时平衡时间为25 min。
-
试验期间,于每天测定凌晨叶水势结束后,取土壤样品测定土壤含水量。在截雨处理干旱样地和对照样地内分别采集4个土壤剖面作为重复,采样深度为0~20 cm,土壤装在铝盒中带回,用电子天平(0.01 g)称取20 g鲜土,在80℃下烘干至恒质量。计算公式为:土壤含水量=[(鲜质量-干质量)/鲜质量] ×100%。图 1表示对照样地和和截雨处理干旱样地内0~20 cm深度土壤含水量季节变化。

图 1 毛竹试验样地土壤含水量动态变化
Figure 1. Dynamic change of soil water moisture
-
不同年龄毛竹叶片凌晨水势生长季节间和冠层间的差异,分别采用Excel 2007软件进行数据处理,各参数以平均值加减1个标准误(mean ±SE)表示。
-
清晨植物的水势值反映根系周围土壤的水分状况,同时也反映植物水分的恢复状况。图 2显示了不同年龄毛竹个体叶片凌晨水势随生长季节的变化。结果表明,自然生长条件下(对照),毛竹叶片凌晨水势随竹龄的变化因季节不同而不同。除了5龄竹叶片凌晨水势高于4龄竹外,8月份叶片凌晨水势基本随着竹龄增加而下降,1年生至3年生竹叶片凌晨水势显著高于老竹。说明鞭根生长旺盛期,毛竹幼龄竹夜间更容易水分恢复,且水分有从幼竹向老竹输送的趋势。12月份和翌年3月份正好相反,随着竹龄增加其叶片凌晨水势增加,新竹凌晨水势最小。说明冬季和春季毛竹老竹水分在夜间更容易得到恢复,且水分具有从老竹高水势向幼竹低水势输送的趋势,表明孕笋期,一天当中上午早些时段内,幼龄壮龄竹子生长需要从老竹得到水分补给。在9月份和10月份,毛竹叶片凌晨水势随着竹龄增加表现出先降后升的趋势。上述研究结果表明水分在毛竹不同竹龄个体之间输送方向因生长季节不同而存在差异。

图 2 自然条件下不同生长季节毛竹个体叶片水势随竹龄的变化
Figure 2. The change of leaf water potential with ages for different growth period of Ph. edulis under natural condition
研究还发现,同龄毛竹个体叶片凌晨水势随生长季节变化也会呈现出峰值变化。其中1年生和7年生毛竹叶片凌晨水势随季节变化呈现“单峰”趋势,以9月份最高;2年生至5年生毛竹个体叶片凌晨水势随季节变化为“双峰”趋势,分别在9月份和12月份出现峰值,以12月份最高。一般而言,植物经过夜间的水分平衡过程,叶片凌晨水势已经与土壤水分达到平衡状态,能够很好地反映土壤水分状况的差异。本试验结果说明叶片凌晨水势与土壤水分状况的一致性。从毛竹试验样地不同处理土壤含水量季节变化曲线可知(图 1),对照样地土壤含水量随季节变化的总体趋势均表现为春季(3月)>冬季(12月)>秋季(9月)>夏季(8月)。由此可见,2年生至5年生毛竹个体叶片凌晨水势在12月份最高,与其土壤含水量季节差异密切相关,土壤含水量高时其叶片凌晨水势也较高。
竹龄与季节水平的双因素方差分析结果表明,同一竹龄毛竹叶片凌晨水势存在较显著的季节间差异(P=0.012 < 0.05),但不同竹龄个体之间叶片水势无显著差异,这表明由于毛竹无性克隆生长的生物学特性,地下相连的复杂鞭根系统,使得不同年龄毛竹个体叶片凌晨水势维持一个相对稳定范围,水分可以在不同年龄毛竹个体之间相互补给,补给方向一定程度受毛竹所在关键生长季节的影响。
-
从图 3可知,模拟截雨干旱后,8月份新竹叶片凌晨水势最高,随着竹龄增加叶片凌晨水势下降;9月份叶片凌晨水势同样以新竹最高,其它竹龄之间差异较小。说明8、9月份鞭根生长旺盛期,水分是从新竹或者小龄竹向母竹输送,即下一级幼小竹向母竹输送水分。10月至翌年3月份,对于4龄以下毛竹,随着竹龄增加其叶片凌晨水势增加,新竹叶片凌晨水势最低,但差异不明显。由此可知,模拟截雨干旱与自然生长下毛竹凌晨水势在不同生长季节随竹龄大小的变化较为相似。方差分析结果表明,截雨干旱后,毛竹叶片凌晨水势在季节间差异极显著(P=0.000 < 0.01),而在竹龄之间无显著差异。

图 3 干旱处理下不同生长季节毛竹个体叶片水势随竹龄的变化
Figure 3. The change of leaf water potential with ages for different growth period of Ph. edulis under drought stress
截雨干旱后,毛竹叶片凌晨水势随季节变化因竹龄而异,但与自然生长毛竹情况不同。1年生和2年生毛竹呈“双峰型”,3年生至5年生毛竹随季节变化呈现“单峰型”,7年生老竹随季节增加而持续增加。
-
图 4显示不考虑毛竹年龄影响情况下,自然生长(对照)与干旱处理毛竹个体叶片凌晨水势随生长季节的变化。结果表明,无论竹龄大小,在自然生长条件下,毛竹个体叶片凌晨水势随生长季节呈现“双峰”趋势,分别在9月和12月份出现峰值;干旱处理下,毛竹叶片凌晨水势随季节呈现“单峰”趋势,在12月出现峰值。结合图 1和图 4可知,毛竹个体叶片凌晨水势随季节变化趋势与其样地土壤含水量的季节变化趋势较为一致。除了12月和翌年3月冬春季节,其它季节干旱处理的毛竹叶片凌晨水势低于自然生长的毛竹。

图 4 自然生长与干旱处理下毛竹叶片凌晨水势随季节的变化
Figure 4. The change of leaf predawn water potential with the different growth period between control and drought stress for different ages of Ph. edulis
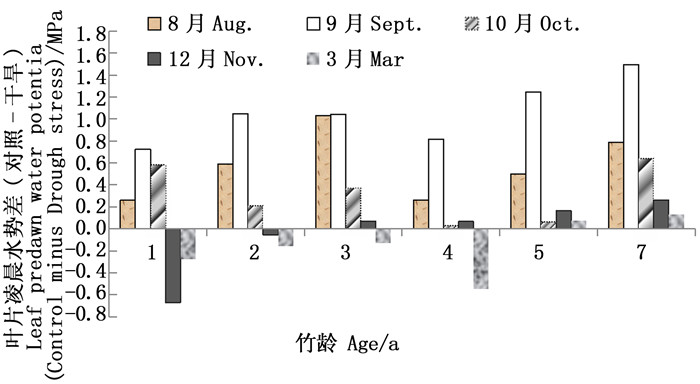
图 5显示不同年龄毛竹个体自然生长(对照)与干旱处理之间的叶片凌晨水势差(对照-干旱)随生长季节的变化。自然生长与干旱处理后毛竹叶片凌晨水势的差异随季节变化而变化,同时受毛竹竹龄大小的影响。主要表现在8月至10月份毛竹生长旺盛期,干旱截雨后毛竹叶片凌晨水势显著低于自然生长的毛竹,且随着竹龄增加,叶片凌晨水势差异幅度增大。而在12月至翌年3月份孕笋期,幼龄竹干旱处理后叶片凌晨水势有接近对照处理毛竹的趋势,这种适应性变化以2年生以下的毛竹个体表现最为明显,5年生、7年生老竹干旱处理后叶片凌晨水势均低于对照处理毛竹。

图 5 不同年龄毛竹个体叶片凌晨水势差随季节的变化
Figure 5. The change of leaf predawn water potential variance with the different growth period between control and drought stress for different ages of Ph. edulis
-
图 6显示毛竹叶片凌晨水势冠层差异随生长季节的变化。研究表明,无论干旱与否,毛竹叶片凌晨水势的冠层差异随季节变化较为明显。自然生长条件下,毛竹叶片凌晨水势冠层差异随生长季节呈现“双峰”趋势,峰值分别在9月份和12月份出现;干旱处理下,毛竹叶片凌晨水势冠层差异随季节变化则呈明显的“单峰”趋势,峰值在12月份出现。方差分析显示,无论干旱与否,毛竹叶片凌晨水势的冠层差异不显著(P>0.05),但各冠层叶片凌晨水势季节间差异极显著(P < 0.001)。

图 6 不同生长季节毛竹叶片凌晨水势冠层差异
Figure 6. The canopy difference of leaf predawn water potential between control (a) and drought stress (b) for different growth stage of Ph. edulis
图 7显示9月份生长旺盛期不同年龄毛竹各冠层叶片凌晨水势在干旱处理前后的变化。自然生长条件下(对照),毛竹叶片凌晨水势冠层差异受竹龄大小影响,2年生和5年生毛竹叶片凌晨水势以冠层中部最高,其次冠层下部,冠层上部较低(图 7-a)。1年生和3年生冠层下部叶片凌晨水势最低,而4年生毛竹冠层下部叶片凌晨水势迅速上升较高水平,符合水分移动规律。在9月份生长旺盛期,3年生以下毛竹,干旱处理后各龄竹叶片凌晨水势基本都为冠层下部最小,冠层中部和冠层上部差异较小(图 7-b)。

图 7 不同年龄毛竹叶片凌晨水势冠层差异
Figure 7. The canopy difference of leaf predawn water potential between control (a) and drought stress (b) for different ages of Ph. edulis
毛竹叶片水势冠层和竹龄的双因素方差分析结果表明(表 1),在9月份生长旺盛期,自然生长条件下毛竹叶片凌晨水势在冠层之间和竹龄之间均无显著差异;而干旱处理后毛竹叶片凌晨水势在不同冠层之间无显著差异,但相同冠层不同年龄个体之间存在极显著差异(P=0.000 9 < 0.001)。可知9月份生长旺盛期,无论干旱与否,毛竹叶片凌晨水势冠层差异均不显著,仅干旱处理后的叶片凌晨水势存在显著的年龄差异。
表 1 毛竹叶片凌晨水势的冠层和竹龄间差异方差分析
Table 1. The variance analysis of predawn water potential among different canopies and ages for Ph. edulis
月份Month 差异源Variance 对照Control 干旱处理Drought stress P-value P-value 8 冠层间Canopy 0.228 7 0.833 0 竹龄间Age 0.015 0 0.278 6 9 冠层间Canopy 0.598 1 0.448 4 竹龄间Age 0.300 3 0.000 9 10 冠层间Canopy 0.878 2 0.067 9 竹龄间Age 0.066 5 0.000 0 12 冠层间Canopy 0.860 8 0.253 4 竹龄间Age 0.000 9 0.733 9 3 冠层间Canopy 0.430 8 0.996 2 竹龄间Age 0.002 1 0.145 9 针对毛竹不同生长发育阶段,通过对不同竹龄个体的各冠层叶片凌晨水势进行方差分析(表 1),结果表明,无论干旱与否,各生长季节毛竹叶片凌晨水势的冠层差异不显著。自然生长条件下,在8月、12月和3月份,毛竹各冠层叶片凌晨水势在不同竹龄个体之间存在极显著差异;干旱处理下,仅9月和10月份毛竹各冠层叶片凌晨水势在不同竹龄个体之间存在极显著差异。
-
叶水势由于能够更直接地反映土壤、植物和大气环境对植物体内水分可利用性的综合影响,在很多研究中作为衡量植物水分状况的指标[4, 6, 9, 13]。树木叶片水势的变化受蒸腾作用和土壤水分状况的影响,且蒸腾耗水对叶片水势的影响较土壤水分状况对叶片水势的影响更快速而直接[6]。植物经过夜间的水分平衡过程,叶片凌晨水势已经与土壤水分达到平衡状态,与白天蒸腾作用对叶片水分状况的影响相比,叶片凌晨水势能够很好地反映土壤实际水分状况的差异。
本研究通过毛竹叶片凌晨水势的测定,定量分析叶片凌晨水势随毛竹生长季节和竹龄的变化趋势,阐明不同竹龄叶片凌晨水势的季节差异性。结果发现,毛竹叶片凌晨水势随竹龄变化幅度较小,随生长季节变化幅度较大,且季节差异又同时受竹龄大小的影响。研究表明,毛竹叶片凌晨水势的显著差异主要表现在同龄竹的不同生长季节之间,不同竹龄之间有一定差异,但不显著。王丁等[9]研究得出干旱胁迫下6种喀斯特造林树种苗木叶片凌晨水势均以生长旺盛期的下降幅度最大,生长末期次之,生长初期最小。本研究毛竹叶片凌晨水势的季节变化趋势与王丁等[9]的研究结果较为一致。与袁佳丽等[8]对毛竹从笋长成竹的快速生长期叶片凌晨水势的研究不同的是,本试验测定了毛竹生长旺盛期、孕笋期和出笋期等不同生长季节叶片凌晨水势的动态变化。毛竹叶片凌晨水势大小随季节变化整体反映了不同生长季节土壤水分状况,而不同竹龄个体之间的叶片凌晨水势相对稳定,这可能从另一个方面反映了毛竹作为克隆植物,其发达的地下鞭根系统彼此相连,使得水分在幼龄竹和母竹之间相互补给,且输送方向因生长季节不同而有差异,这说明叶片凌晨水势可能是毛竹不同生长季节水分有效利用的驱动因素。如本试验研究发现,无论干旱与否,毛竹叶片凌晨水势在8月份至10月份生长旺盛期,新竹叶片凌晨水势最高,即随着竹龄增加叶片水势下降,此时水分从幼龄竹向母竹输送,这可能反映了毛竹长鞭季节幼龄竹发达的鞭根从土壤中吸水的能力较强;12月份孕笋期至翌年3月出笋期,老竹叶片凌晨水势较高,即随着竹龄增加叶片凌晨水势增加,此时水分从母竹向幼龄竹输送,反映了幼龄竹在孕笋期和出笋期需要消耗更多水分,其通过保持较低的水势从水势较高的母竹获得水分补给。袁佳丽等[8]研究认为,毛竹从笋长成竹过程中不同竹龄毛竹叶水势基本都表现为随年龄增长,叶水势不断增大,且竹龄对水势的影响达到显著水平。本研究发现翌年3月份(出笋期)随着竹龄增加,毛竹叶片凌晨水势亦增加,这与袁佳丽等[8]毛竹从笋长成竹期间的凌晨水势变化研究结果一致。
-
在干旱胁迫条件下植物如何保持有效的水分运输显得尤为重要[14]。罗永忠等[4]对水分胁迫下紫花苜蓿(Medicago sativa L.)叶水势随生育期的变化规律研究发现,随土壤含水量的不同,叶水势出现的峰值在各生育阶段也不一样,轻度、中度水分胁迫下,叶水势最低值出现在苗期,充分供水时最高值出现在开花期。与该研究结果相似,本研究发现毛竹自然生长状态和截雨模拟干旱状态下叶片凌晨水势随生长季节的变化也不一样,其凌晨水势峰值出现季节不同,且受毛竹竹龄大小影响。
有学者研究表明,短期干旱胁迫后,水稻(Oryza.sativa L.)叶片凌晨水势明显低于对照处理的[5]。黎明前叶水势和相对含水量均随着干旱胁迫的增加而降低,这种响应有利于植物更有效地从土壤中吸收水分[9]。本研究也发现毛竹经历截雨干旱1a后,干旱处理与自然生长(对照)毛竹叶片凌晨水势差异较大,并且受生长季节影响。在生长旺盛期,干旱处理后毛竹叶片凌晨水势显著低于自然生长下的毛竹,且随着竹龄增加,叶片凌晨水势差异幅度增大,说明干旱胁迫下毛竹体内水分出现亏缺,且随着竹龄增加水分亏缺程度加深,因此老竹鞭根生长受限。而在孕笋期,5年生以上老竹干旱处理后叶片凌晨水势低于自然生长(对照)毛竹,2年生以下幼龄竹干旱处理后叶片凌晨水势出现接近自然生长(对照)毛竹的趋势。从试验样地的土壤含水量季节变化趋势可知,在12月份至翌年3月份的孕笋期,其土壤含水量较高,这与该季节降水较多有关,因此干旱样地幼龄毛竹叶片凌晨水势接近自然生长(对照)毛竹,是否与不同年龄毛竹叶片对林中露水的吸收等有关还有待进一步研究。这也可能是干旱胁迫后,毛竹个体在土壤水分供应受限情况下,通过生理生态适应产生干旱忍耐或逃避策略。如调查发现随着干旱时间推进毛竹产生大量落叶,以保证在一定的光合作用时,尽可能减少叶面蒸腾失水。其中自然生长(对照)和干旱处理下样地内毛竹半年凋落物总产量分别为0.287 0 kg·m-2和0.256 5 kg·m-2,与对照样地相比,干旱处理样地凋落物产量降低10.64%;无论干旱与否,试验样地内毛竹均是11月份凋落物产量最高,对照处理比干旱处理高19.06%。另外,毛竹孕笋期土壤水分含量、空气湿度相比夏季旺盛生长期有所增加,太阳辐射相比减弱,所以蒸腾减弱的同时保证了叶片凌晨水势的夜间恢复。
本研究发现,各生长季节5年生以上老竹干旱处理后叶片凌晨水势均显著低于自然生长(对照)下的毛竹,这也进一步说明老竹与幼龄竹相比,鞭根系统活力下降,所以随着干旱胁迫时间的推进,老竹根系从土壤中吸水的能力下降,更易遭受土壤水分胁迫的危害,抗旱能力有所下降。
植物叶片的水分状态对植物叶片光合作用有直接的影响,尽管植物在旱季水势降低,植物可通过提高渗透调节,减少叶面积来维持水分平衡和基本的光合生理功能,进而适应干旱环境[14]。对不同土壤水分条件下紫花苜蓿叶水势、蒸腾速率和气孔导度的变化规律研究发现,当叶水势下降一定程度时会引起气孔关闭,而水分充分或者轻度胁迫时,蒸腾速率越高则叶水势越低,有利于水分的吸收;且叶水势的变化除受土壤水分影响外,还与叶片生长发育密切相关[4]。杨晓东等[15]研究毛竹生理生态特征对不同土壤水分的响应,发现土壤水分变化影响毛竹叶片水势,白天水势下降,傍晚均可恢复到凌晨水势。
-
冠层叶片功能性状的梯度分布是林木冠层基本特征之一。同一植株不同部位的叶片的水势是不同的,这种变化与处于不同位置的叶片的叶龄及其周围的环境有关。水分移动的规律一般认为水分由水势高处往水势低处移动,植物的水分经由土壤到达根部,通过根系吸水进入植物体内,再通过植物的茎枝,进入叶片,最后扩散至植物周围的空气中[16]。叶片水势冠层分布梯度因树种自身特性、因时间而异。胡杨(Populus euphratica Oliv.)上、中、下3个部位的叶水势日变化测定结果表明,胡杨叶片水势在9:00之前表现为上高下低,9:00以后为上低下高[17]。袁佳丽等[8]研究发现,从纵向水分传输上看,毛竹不同林冠层对水势的影响均达到显著水平,不同生长期林冠层的叶水势均为下层水势>中层水势>上层水势,表明毛竹在从笋到竹快速生长期内水分传输符合从冠层下部向上输送的水分移动规律。
有关冠层高度对毛竹生长的影响已有报道[18-20],但对叶片水势的影响研究较少[8]。本研究发现不同生长季节不同冠层部位的毛竹叶片凌晨水势有差异,且叶片凌晨水势随冠层分布趋势受竹龄大小影响。与司建华等[17]研究不同,本研究在自然生长下,各季节5年生以上老竹冠层下部叶片凌晨水势大于冠层上部,这符合水分从冠层下部到上部的输送规律;而3年生以下毛竹叶片凌晨水势以冠层下部较低。干旱处理后,除了2年生、4年生毛竹外,各龄竹毛竹叶片凌晨水势基本为冠层下部 < 冠层中部 < 冠层上部的梯度变化。叶水势低,有利于树木从土壤中吸取水分。毛竹冠层水势分布出现反常趋势,可能是其通过调节较低的水势以实现从土壤中获取水分补给来满足正常生长需要,从一个侧面反映毛竹适应环境能力的大小。
一般植物上层叶片的蒸腾强度最高,叶水势最低;下层叶片的蒸腾强度最低,叶水势最高[16]。作者前期研究发现不同生长季节不同年龄毛竹其叶片蒸腾速率日均值存在显著的冠层差异,在出笋期,2年生和3年生毛竹个体叶片蒸腾速率日均值表现出冠层上部 < 冠层下部的垂直变化趋势;在孕笋期,3年生毛竹个体叶片蒸腾速率日均值随冠层增加略微下降[21]。这可能也佐证了本文研究发现自然生长3年生以下毛竹叶片凌晨水势冠层下部小于冠层上部的现象。
-
清晨植物的水势值反映植物水分的恢复状况。自然生长下,毛竹个体叶片凌晨水势随年龄变化受季节影响,表明水分在毛竹不同竹龄个体之间输送方向因生长季节不同而存在差异,这也体现水势可能是毛竹不同生长季节水分有效利用和增加抗旱的驱动因素。在毛竹生长旺盛期,幼龄毛竹夜间更容易水分恢复,水分有从幼竹向老竹输送的趋势。秋冬季孕笋期间,老竹夜间更容易水分恢复,水分有从老竹高水势向幼竹低水势输送的趋势。毛竹叶片凌晨水势与土壤水分状况具有一致性,土壤含水量高时其叶片凌晨水势也较高。因此,同一竹龄毛竹叶片凌晨水势存在较显著的季节间差异。截雨干旱后,毛竹凌晨水势在不同生长季节随竹龄大小的变化与自然生长下较为相似。无论竹龄大小,在自然生长条件下,毛竹个体叶片凌晨水势随生长季节呈现“双峰”趋势;干旱处理下,毛竹叶片凌晨水势则随季节呈现“单峰”趋势。干旱处理的毛竹叶片凌晨水势低于自然生长的毛竹。无论干旱与否,毛竹各冠层叶片凌晨水势在不同竹龄个体之间的显著程度因生长季节而异。本研究定量分析了毛竹叶水势随生长季节和竹龄的梯度变化,拟结合毛竹不同生长季节叶片光合速率、蒸腾速率和气孔导度的季节变化和日变化相关性分析,进一步开展毛竹叶水势变化及其气孔响应机制研究。
模拟干旱下毛竹叶片水势的动态变化
The Dynamic Change of Leaf Water Potential for Moso Bamboo Under Throughfall Exclusion
-
摘要:
目的 定量分析毛竹叶片凌晨水势随生长季节和竹龄的梯度变化趋势及其对截雨干旱的响应特征,阐明毛竹不同年龄个体水分供给关系的季节差异。 方法 通过人工截雨模拟干旱试验,采用PSYPRO水势测量系统动态监测毛竹叶片凌晨水势的变化。 结果 (1)无论干旱与否,同一竹龄毛竹叶片凌晨水势在季节间差异较显著(P < 0.05),而在竹龄之间无显著差异。(2)同龄毛竹叶片凌晨水势随生长季节变化呈现峰值,且因竹龄而异。自然生长(对照)1年生和7年生毛竹叶片凌晨水势随季节变化呈现"单峰"趋势,以9月份最高;2年生至5年生个体叶片凌晨水势随季节变化为"双峰"趋势,分别在9月份和12月份出现峰值。截雨干旱后,1年生和2年生毛竹叶片凌晨水势呈"双峰型",3年生至5年生随季节变化呈"单峰型"。叶片凌晨水势与土壤水分状况随季节变化趋势具有一致性。(3)截雨干旱与自然生长下毛竹叶片凌晨水势于不同生长季节随竹龄大小的变化趋势较为相似。干旱与否,在8月份至10月份生长旺盛期,叶片凌晨水势随着竹龄增加而下降,水分有从幼竹向老竹输送的趋势。12月份至翌年3月份孕笋期正好相反,随着竹龄增加其叶片凌晨水势增加,水分具有从老竹高水势向幼竹低水势输送的趋势。表明水分在毛竹不同竹龄个体之间输送补给方向因生长季节不同而存在差异。(4)在生长旺盛期,干旱处理毛竹叶片凌晨水势显著低于自然生长毛竹,且随着竹龄增加,水势差异幅度增大;在孕笋期,5年生以上老竹干旱后叶片凌晨水势低于对照毛竹。(5)不同生长季节不同冠层部位叶片凌晨水势存在差异,但均不显著,叶片凌晨水势随冠层分布趋势也受竹龄大小影响。 结论 同一竹龄毛竹叶片凌晨水势存在较显著的季节间差异,毛竹叶片凌晨水势与土壤水分状况的季节变化趋势一致。水分在毛竹不同竹龄个体之间输送方向因生长季节而异,表明水势是驱动毛竹适应干旱胁迫的重要因素。 Abstract:Objective The objective of this paper is to analyze quantitatively the leaf predawn water potential change with the growth season and age along canopy height for moso bamboo (Phyllostachys edulis) in natural and drought stress environment, and to clarify the seasonal differences of water supply for individuals of different ages of Ph. edulis, lay a theoretical basis for the sustainable and efficient management of Ph. edulis plantations in the global climate background. Method The throughfall exclusion was set to simulate the drought stress, and the PSYPRO water potential measurement system was used to quantitatively analyze the trend of leaf water potential change in the predawn with the growth season and bamboo age. Result (1) There was significant difference among different seasons for leaf predawn water potential for the same age of bamboo (P < 0.05), regardless of throughfall exclusion or not. (2) The leaf predawn water potential changing with the growth season showed a peak trend for the same age of bamboo, and was different due to bamboo age. Under natural growth environment (the control), the predawn water potential for 1-and 7-year-old bamboo changing with the seasonal showed a single peak with the highest value in September, and for 2-to 5-year-old individual it showed a bimodal change trend with the highest values in September and December respectively. The seasonal change trend of leaf predawn water potential was consistent with the soil moisture. After the throughfall exclusion, the leaf predawn water potential was "double peak" type for 1-and 2-year-old individual, and was "single peak" type for 3-to 5-year-old individual, which changed with the seasons. (3) The change with increasing age of bamboo for leaf predawn water potential under throughfall exclusion environment was similar to that of the control during different growth season. Regardless of throughfall exclusion or not, the leaf predawn water potential decreased with the increase of bamboo age during the rhizome vigorous growth season from August to October, and showed the trend of the water transport from young bamboos to old ones, while the leaf predawn water potential increased with the increase of bamboo age during the shoot development period from October to March the following year, and the water transported from the high water potential of old bamboo to low water potential of young ones, indicating that the water supply direction between different age of bamboo individuals changed in different season. (4) During the vigorous growth of rhizome, the leaf predawn water potential under throughfall exclusion was significantly lower than that under natural growth environment, and the difference in leaf predawn water potential became more significant with the increase of bamboo age. Under throughfall exclusion, only the leaf predawn water potential of the bamboo with the age over 5-years-old was lower than the control during the shoot development period. (5) There were no significant differences in predawn water potential of different canopy leaves in different growth seasons, but the change trend of leaf water potential was also affected by the bamboo age. Conclusion There was a significant seasonal difference of leaf predawn water potential for same aged individual. Both the leaf predawn water potential and the soil moisture follow same seasonal change trend. The transport direction of water between different age of bamboo individuals varied with season, indicating that the leaf predawn water potential is an important factor in driving the bamboo to adapt to drought stress. -
Key words:
- Phyllostachys edulis
- / throughfall exclusion
- / leaf water potential
- / growth period
- / age
- / canopy height
-
图 2 自然条件下不同生长季节毛竹个体叶片水势随竹龄的变化
Figure 2. The change of leaf water potential with ages for different growth period of Ph. edulis under natural condition
图 3 干旱处理下不同生长季节毛竹个体叶片水势随竹龄的变化
Figure 3. The change of leaf water potential with ages for different growth period of Ph. edulis under drought stress
图 4 自然生长与干旱处理下毛竹叶片凌晨水势随季节的变化
Figure 4. The change of leaf predawn water potential with the different growth period between control and drought stress for different ages of Ph. edulis
图 5 不同年龄毛竹个体叶片凌晨水势差随季节的变化
Figure 5. The change of leaf predawn water potential variance with the different growth period between control and drought stress for different ages of Ph. edulis
图 6 不同生长季节毛竹叶片凌晨水势冠层差异
Figure 6. The canopy difference of leaf predawn water potential between control (a) and drought stress (b) for different growth stage of Ph. edulis
图 7 不同年龄毛竹叶片凌晨水势冠层差异
Figure 7. The canopy difference of leaf predawn water potential between control (a) and drought stress (b) for different ages of Ph. edulis
表 1 毛竹叶片凌晨水势的冠层和竹龄间差异方差分析
Table 1. The variance analysis of predawn water potential among different canopies and ages for Ph. edulis
月份Month 差异源Variance 对照Control 干旱处理Drought stress P-value P-value 8 冠层间Canopy 0.228 7 0.833 0 竹龄间Age 0.015 0 0.278 6 9 冠层间Canopy 0.598 1 0.448 4 竹龄间Age 0.300 3 0.000 9 10 冠层间Canopy 0.878 2 0.067 9 竹龄间Age 0.066 5 0.000 0 12 冠层间Canopy 0.860 8 0.253 4 竹龄间Age 0.000 9 0.733 9 3 冠层间Canopy 0.430 8 0.996 2 竹龄间Age 0.002 1 0.145 9  下载: 导出CSV
下载: 导出CSV
-
[1] 金鹰, 王传宽.植物叶片水力与经济性状权衡关系的研究进展[J].植物生态学报, 2015, 39(10):1021-1032. doi: 10.17521/cjpe.2015.0099 [2] Mcdowell N G, Beerling D J, Breshears D D, et al. The interdependence of mechanisms underlying climate-driven vegetation mortality[J]. Trends in Ecology & Evolution, 2011, 26(10):523-532. [3] Martinez V J, Poyatos R, Aguade D, et al. A new look at water transport regulation in plants[J]. New Phytologist, 2014, 204(1):105-115. doi: 10.1111/nph.12912 [4] 罗永忠, 成自勇.水分胁迫对紫花苜蓿叶水势、蒸腾速率和气孔导度的影响[J].草地学报, 2011, 19(2):215-221. [5] 胡继超, 曹卫星, 姜东, 等.小麦水分胁迫影响因子的定量研究Ⅰ干旱和渍水胁迫对光合、蒸腾及干物质积累与分配的影响[J].作物学报, 2004, 30(4):315-320. doi: 10.3321/j.issn:0496-3490.2004.04.004 [6] 张迎辉, 王华田, 亓立云, 等.水分胁迫对3个藤本树种蒸腾耗水性的影响[J].江西农业大学学报, 2005, 27(5):723-728. doi: 10.3969/j.issn.1000-2286.2005.05.018 [7] 应叶青, 郭璟, 魏建芬, 等.干旱胁迫对毛竹幼苗生理特性的影响[J].生态学杂志, 2011, 30(2):262-266. [8] 袁佳丽, 温国胜, 张明如, 等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报, 2015, 32(5):722-728. [9] 王丁, 姚健, 杨雪, 等.干旱胁迫条件下6种喀斯特主要造林树种苗木叶片水势及吸水潜能变化[J].生态学报, 2011, 31(8):2216-2226. [10] 陈志成, 陆海波, 刘晓静, 等.宝天曼三桠乌药对降雨减少后的生理生态响应[J].林业科学研究, 2017, 30(3):430-435. [11] Brando P M, Nepstad D C, Davidson E A, et al. Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest:results of a throughfall reduction experiment[J]. Philosophical Transactions Royal Society B, 2008, 363(1498):1839-1848. doi: 10.1098/rstb.2007.0031 [12] Misson L, Rocheteau A, Rambal S, et al. Functional changes in the control of carbon fluxes after 3 years of increased drought in a Mediterranean evergreen forest[J]. Global Change Biology, 2010, 16(9):2461-2475. [13] 杨显基, 杜建会, 张楚杰, 等.平潭岛典型海岸草丛沙堆植物群落水势日变化特征及其影响因素[J].生态学报, 2016, 36(9):2614-2619. [14] 王爱英, 姜艳娟, 郝广友, 等.季节性干旱胁迫对石灰山三种常绿优势树种的水分和光合生理的影响[J].云南植物研究, 2008, 30(3):325-332. doi: 10.3969/j.issn.2095-0845.2008.03.006 [15] 杨晓东, 罗艾滢, 黄玉清, 等.漓江上游毛竹生理生态特征对不同土壤水分的响应[J].广西植物, 2017, 37(2):211-219. [16] 付爱红, 陈亚宁, 李卫红, 等.干旱、盐胁迫下的植物水势研究与进展[J].中国沙漠, 2005, 25(5):744-749. doi: 10.3321/j.issn:1000-694X.2005.05.021 [17] 司建华, 冯起, 张小由.极端干旱区胡杨水势及影响因子研究[J].中国沙漠, 2005, 25(4):505-510. doi: 10.3321/j.issn:1000-694X.2005.04.009 [18] Cao K F, Yang S J, Zhang Y J, et al. The maximum height of grasses is determined by roots[J]. Ecology Letters, 2012, 15(7):1-7 [19] Wen G S, Zhang L Y, Zhang R M, et al. Temporal and spatial dynamics of carbon fixation by moso bamboo (Phyllostachys pubescens) in subtropical China[J]. Botanical Review, 2011, 77(3):271-277. doi: 10.1007/s12229-011-9068-x [20] Cao Y H, Zhou B Z, Wang X M, et al. The Photosynthetic characteristics of moso bamboo (Phyllostachys pubescens) for different canopy leaves[J]. Advanced Materials Research, 2013, 726-731:4274-4279. doi: 10.4028/www.scientific.net/AMR.726-731 [21] 曹永慧, 周本智, 王小明, 等.冠层高度对毛竹叶片光合生理特性的影响[J].西北植物学报, 2016, 36(11):2256-2266. doi: 10.7606/j.issn.1000-4025.2016.11.2256 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4287
- HTML全文浏览量: 1572
- PDF下载量: 368
- 被引次数: 0
