


-
森林生态系统中,凋落分解对土壤养分周转、能量流动、碳循环等方面发挥着重要的作用[1-2]。森林生态系统一般都是多种植物的复合系统,凋落物主要以混合状态而非孤立的形式存在[3]。这种混合状态可以改变凋落物的分解环境[4]、异质凋落物间的养分互补[5-6],以及微生物群落和土壤动物的栖息生境[7-8],进而改变凋落物分解和养分循环的速率。近年来,混合凋落物的研究逐渐受到重视,混合凋落物分解和养分释放产生的混合效应并无一致的结论,主要表现为正效应[9]、负效应[10]和加和效应[11]。综合30项研究结果发现,70%混合凋落物叶分解表现出混合效应(正效应或负效应)[12],即混合凋落物的实际分解速率偏离于期望分解速率。混合凋落物的分解过程不仅和各物种的特异性质有关,还受其物理、化学性质组分的差异的影响,甚至改变混合凋落物分解的最终结果[13]。因此,在同一生态系统中深入了解凋落物混合分解的效应,对认识不同物种间的相互作用和关系具有重要意义。竹林作为我国一种主要的森林类型,具有分布广、面积大、生长快等特点,高效的固碳能力对调节大气CO2浓度具有重要作用[14]。在亚热带毛竹林中往往伴生一些林下植被[15],这些林下植被的存在对毛竹林凋落物的分解和养分动态的影响如何,以及林下植被本身凋落物的分解动态情况又如何,目前,关于竹林和林下植被复合模式下凋落物分解和养分动态的研究还未见报道。本研究以四川省长宁县典型竹林——毛竹(Phyllostachys edulis (Carr.) H.de Lehaie)和林下植被优势种芒箕(Dicranopteris pedata (Houtt.) Nakaike)为研究对象,对比毛竹和芒箕凋落物的化学特征,分析两种植被单独以及混合凋落分解和养分动态的变化特征,探讨毛竹和芒箕凋落物之间相互作用的潜在机制,为毛竹林林下植被的合理经营管理提供理论参考。
-
研究地点位于四川省长宁县蜀南竹海景区内。中亚热带湿润性季风气候,温暖湿润,年均气温18.3℃,年均降雨量1 114.2 mm,年均空气相对湿度83%,日照数1 148 h。地貌以中低山地和丘陵为主,土壤类型主要为山地黄壤和紫色土。植被覆盖率为42%,主要竹种有毛竹、苦竹(Pleioblastus amarus (Keng) Keng)和硬头黄竹(Bambusa rigida Keng & Keng f.)等,林下植物有芒萁(Dicranopteris pedata (Houtt.) Nakaike)、狗脊(Woodwardia japonica(L. f.) Sm.)、里白(Diplopterygium glaucum (Thunb.) Ching)等[16]。
-
选取环境和林分条件基本一致的毛竹林,随机设置8个20 m×20 m标准样地。2016年4月,采集毛竹和林下植被芒箕的新鲜凋落物叶,65℃烘干至恒质量后装进孔径为1mm的凋落物分解袋,每袋10 g干样品,3种凋落物类型:纯毛竹叶,纯芒箕叶,毛竹和芒箕混合叶(1:1)。2016年6月1日,每个样地内随机放置36个凋落物袋(3种凋落物类型各12个)。2016年6月至2017年6月间,每3个月从每个样地内随机取回9袋(3种凋落物类型各3个),共收集4次。
-
将取回的分解袋内的其它物质(如新长的植物根系等)检出,然后把凋落物放入65℃烘箱中烘干至恒质量,得出剩余干质量。凋落物粉碎过100目筛后待用,全碳含量采用重铬酸外加热氧化法测定,全氮、全磷的含量采用H2S04/H2O2消煮法,用Smartchem 300全自动化学分析仪测定;碳化学组分采用13C固体核磁共振波谱(13C CPMAS NMR)法测定;用Li8100-配备的水分和温度探头测定凋落物袋附近的表层土壤温度和含水量,每半月记录1次数据,连续监测整个凋落物分解周期。
-
凋落物干质量剩余率:MR=(Mt/M0)×100,式中,M0为初始凋落物干质量,Mt是t时凋落物分解剩余干质量。
凋落物分解速率(k):ln(Mt/M0)=-kt,式中,k为凋落物分解系数。半衰期(t0.5)和分解95%所需时间(t0.95)分别为:t0.5=0.693/k, t0.95=3/k。
元素剩余率:NR=100×(Ct×Mt)/(C0×M0),式中,C0为初始元素含量(g·kg-1),Ct为t时凋落物元素含量(g·kg-1)。
期望干质量(养分)剩余率=100×(M1×MR1+ M2×MR2)/(M1 +M2),式中,M表示各组分在混合物中的初始干质量(或养分含量),MR表示t时各组分单一凋落物的干质量剩余率(或养分剩余率),本研究中混合凋落物组分为2个,n=2。
利用SPSS 19.0进行统计分析,采用T-检验法检测毛竹和芒箕凋落物间干质量剩余率、养分剩余率的差异,及混合凋落物干质量剩余率和养分剩余率的实测值与期望值间的差异,采用Pearson法检测凋落物干质量损失量与土壤含水量、土壤温度及初始凋落物质量的相关性;利用Sigmaplot 12.5软件进行制图。
-
毛竹和林下植被芒箕的凋落物叶初始化学组分有着明显差异(表 1)。其中C、N、P 3种元素含量在芒箕凋落物中均显著高于毛竹林凋落物(P < 0.05);C:N和C:P比为毛竹凋落物显著高于芒箕凋落物(P < 0.05),而N:P比无显著差异性。
表 1 凋落物的初始化学组成
Table 1. Initial chemical composition of litter
凋落物类型Litter type C/(g·kg-1) N/(g·kg-1) P/(g·kg-1) C:N C:P N:P 毛竹P. edulis 415.78±0.15b 17.55±0.04b 0.204±0.001b 23.69±0.06a 2 042.40±8.08a 86.21±0.48a 芒箕D. pedata 421.64±0.29a 35.33±0.19a 0.404±0.001a 11.93±0.06b 1 044.06±2.13b 87.49±0.42a 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8.毛竹和芒箕凋落物叶初始有机碳化学结构研究结果显示,两者有机碳化学结构格局不同,毛竹中氧烷基碳最高(62.39%±0.75%),羟基碳最低(7.49%±0.13%),而芒箕中羟基碳最高(52.81%±0.95%),芳香碳最低(9.82%±0.30%);毛竹中烷基碳、氧烷基碳和芳香碳均显著高于芒箕(P < 0.05),而羟基碳及烷基碳/氧烷基碳比显著低于芒箕(P < 0.05)(表 2)。
表 2 凋落物的初始有机碳化学结构相对含量
Table 2. Proportions of the components of initial organic carbon functional group of litter
% 凋落物类型
Litter type烷基碳
Alkyl氧烷基碳
O-alkyl芳香碳
Aromatic羟基碳
Carbonyl烷基碳/氧烷基碳
Alkyl/o-alkyl毛竹P. edulis 15.02±0.12a 62.39±0.75a 15.11±0.21a 7.49±0.13b 24±0.27b 芒箕D. pedata 12.55±0.28b 24.81±0.44b 9.82±0.30b 52.81±0.95a 51±0.43a 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8. -
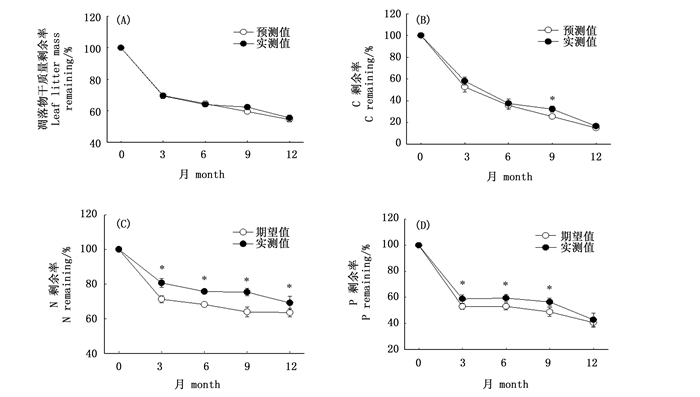
毛竹和芒箕凋落物干质量剩余率随分解进程均呈逐渐下降的趋势,且凋落物分解总体表现为初始阶段分解最快,后期分解逐渐减慢(图 1A)。分解1年后,毛竹和芒箕凋落物干质量剩余率分别为56.10%和48.03%,差异性显著(P < 0.05)(表 3)。从表 3可以看出芒箕凋落物年分解速率(0.73±0.02)明显高于毛竹(0.58±0.03)(P < 0.05),Olson的指数方程能够很好模拟两种植被凋落物的分解过程,毛竹和芒箕相关系数(R2)分别为0.999和0.984。
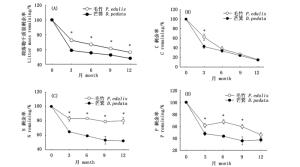
图 1 单一凋落物分解过程中干质量和养分含量的变化(*表示种间差异显著(P < 0.05))
Figure 1. Single litter decomposition rates and nutrient dynamics duiring decomposition (*denote significant difference at P < 0.05 between two species)
表 3 凋落物干质量剩余率与时间的回归分析
Table 3. Regression analysis between dry weight residue of litter and time
类型
Type回归方程
Equation相关系数
R2年残留率
Remaining rate/%分解常数
k半衰期
t0.5/a周转期
t0.95/a毛竹P. edulis y = 0.790 3e-0.341t 0.999 56.10±1.77a 0.58±0.03b 1.21±0.07a 5.25±0.30a 芒箕D. pedata y = 0.630 6e-0.262t 0.984 48.03±0.87b 0.73±0.02a 0.95±0.02b 4.10±0.10b 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8.毛竹和芒箕凋落物C、N、P剩余率全年整体均呈下降趋势(图 1B,C,D),表现为净释放状态。毛竹和芒箕间元素释放格局表现不同,两者C元素释放速率只在分解初期的前3个月有显著差异,表现为芒箕显著高于毛竹(图 1B)(P < 0.05),全年芒箕N元素释放速率显著高于毛竹(图 1C)(P < 0.05),前9个月芒箕P元素释放速率显著高于毛竹(图 1D)(P < 0.05)。
-
凋落物混合干质量剩余率实测值与期望值对比结果表明,毛竹和芒箕凋落物的混合分解无显著混合效应(正效应或负效应)(图 2A)。凋落物混合对分解过程中C、N、P元素释放的影响格局不同(图 2B,C,D)。在整个1年的分解过程中,实测C剩余率只在9个月时与期望值有显著差异(P < 0.05);而混合分解对N元素的释放影响全年表现为负效应,实测N剩余率比期望值平均高15.82%(P < 0.05);混合分解对P元素的释放的前9个月有显著的负效应(P < 0.05)。这说明毛竹和芒箕凋落物混合对C、N、P元素释放整体表现为负效应。
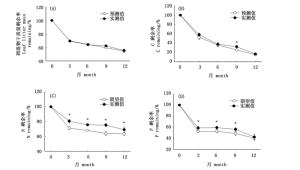
图 2 混合凋落物干质量和养分的实测剩余率与期望剩余率(*表示实测值与期望值差异显著(P < 0.05))
Figure 2. Observed and expected mass and nutrient remaining in mixtures litter(*denote significant differences betweenthe observed and expected values(P < 0.05))
-
在整个凋落物分解过程中,毛竹和芒箕间C、N、P含量整体变化趋势一致(图 3A,B,C),其中C含量整体呈下降趋势,N含量有小幅度的上升趋势,P含量有微弱的下降趋势,毛竹和芒箕凋落物在全年的分解过程中C、N、P含量均差异显著(P < 0.05),分解1年后,毛竹和芒箕C含量分别为291.33 g·kg-1和333.40 g·kg-1,N含量分别为25.11 g·kg-1和38.56 g·kg-1,P含量分别为0.17 g·kg-1和0.32 g·kg-1;毛竹和芒箕间C、N、P化学计量比的特征整体变化趋势一致(图 3D,E,F),C:N比呈整体下降的趋势,C:P比呈波动性变化,而N:P表现出小幅的上升,分解1年后,毛竹和芒箕间N:P由初始的无差异,在末期表现出显著性差异(P < 0.05),N:P比值分别为151.85和125.32。
图 3 凋落物分解过程中化学计量特征(*表示实测值与期望值差异显著(P < 0.05))
Figure 3. Stoichiometric characteristics of litter decomposition(*denote significant differences between the observed and expected values(P < 0.05))
-
相关分析表明,凋落物叶的干质量损失率与土壤温度呈显著正相关(P < 0.01),与土壤含水量无显著相关性;凋落物叶的干质量损失率与与初始凋落物叶的N、P含量呈显著正相关(P < 0.05),与初始凋落物叶的C:N和C:P呈极显著负相关(P < 0.01)(表 4)。
表 4 凋落物干质量损失量与土壤含水量、温度及初始凋落物质量的Pearson相关系数
Table 4. Pearson correlation coefficient between dry mass loss of litter and soil water content and temperature, the initial quality
类型
Type土壤含水量
Soil water content土壤温度
Soil temperatureC N P C:N C:P N:P 毛竹P. edulis -0.247 0.783** 0.154 0.572* 0.521** -0.411* -0.552* 0.178 芒箕D. pedata -0.311 0.809** 0.123 0.504* 0.373* -0.394* -0.379* 0.172 毛竹和芒箕P. edulis&D. pedata -0.290 0.751* 0.253 0.634** 0.437* -0.517* -0.430* 0.204 注:*表示显著水平P < 0.05;**表示显著水平P < 0.01.
Note: *mean sinificant difference(P < 0.05),**mean sinificant difference(P < 0.01). -
理化和生物等因子综合调控凋落物的分解速率[19-21]。凋落物质量主要体现在凋落物的化学组分上,在中小尺度上,凋落物质量在控制自身分解方面具有关键作用[22],凋落物中C:N、C:P比越低,耐分解化合物含量越少,同时,N素可以加强微生物繁殖速率和代谢活性,有利于加快凋落物的分解速率[23],本研究中,芒箕初始N、P元素含量高于毛竹,而C:N、C:P低于毛竹(表 1),分解过程中,凋落物干质量损失量与N、P含量呈现出显著正相关,与C:N和C:P表现为显著负相关(表 4),这与前人的研究结果一致[24-25]结果显示芒箕叶凋落物分解速率比毛竹更快(图 1A)。此外,在13C-NMR波普解析的官能团中,烷基碳和芳香族碳等表征难以被利用的木质素、单宁等,即难分解碳,氧烷基碳表征易被利用的碳水化合物,即易分解碳,研究表明难分解碳含量越高,有机物的分解速率越慢[26]。本研究中芒箕凋落物叶中难分解的烷基碳和芳香碳相对含量均显著低于毛竹,利于凋落物分解速率的加快,同时毛竹凋落物叶中易分解的氧烷基碳相对含量显著高于芒箕,但分解速率并没表现出更快的结果,这可能与毛竹凋落物叶中氮含量相对较低,限制了凋落物分解微生物的活性和繁殖速率,进而影响了毛竹凋落物的分解速率。
此外,环境因子中土壤温度和土壤含水量对凋落物分解速率的影响可通过调节土壤微生物和酶活性来实现[27]。本研究结果发现土壤温度与凋落物分解呈显著正相关(表 4),这支持以往研究中凋落物分解速率随温度升高而加快的结果[28-29]。此外,本研究还发现土壤含水量与凋落分解速率无显著相关性,这可能和本研究区土壤湿度相对稳定有关。可见,在本研究区内土壤温度对凋落物的分解速率比土壤含水量的影响更突出。
-
有研究表明,有些物种在凋落物混合分解过程中分泌的某些产物可对其它物种产生有益(或有害)的影响[30],因各物种的特异性可导致不同物种组合间分解速率产生差异。本研究中,在1年的分解过程中,混合凋落物叶分解表现为加和效应(图 2A),这与前人的有关的研究相一致[30-31]。这可能是因为各物种间产生的正、负效应彼此平衡,导致总体失重率差异不显著,从而忽略了各物种的特异性,未体现出混合效应[32]。此外,混合凋落物不同分解阶段,其混合效应(加和或非加和效应)表现也会有所差异,如通过对红花槭(Acer rubrum L.)、北美乔松(Pinus strobus Linn.)和红云杉(Picea rubens Sarg.)3种凋落物等比例混合研究表明,混合分解失重率第1年表现为加和效应,而第2年却表现出非加和的正效应[33];此外,落叶树凋落物等比例混合后,研究也表现出类似的结果,混合凋落物分解前期的9个月为非加和的正效应,而后期的10个月呈现出非加和的负效应[12]。本研究分解试验只进行了1年,研究结果只能表明短期内毛竹和芒箕凋落物混合对失重率没有显著影响,但混合分解在更长的时间周期内是否会产生非加和效应还有待进一步研究。
不同物种组成、元素间的相互影响、分解阶段等多重因素使凋落物混合分解中养分释放规律十分复杂[13]。前人研究表明,在混合凋落物分解过程中,高质量凋落物可向低质量凋落物进行养分转移,从而阻缓了低质量凋落物的养分释放速率,加快了高质量凋落物的养分释放速率。在一些凋落物分解过程中,C、N养分的释放存在一定的关联性[34],如欧洲赤松(Pinus sylvestris L.)和玉米(Zea mays L.)叶混合凋落物研究发现,欧洲赤松倾向于给玉米转移C元素,而玉米倾向于给欧洲赤松转移N元素,C元素释放在一定程度上受到N元素释放的限制[6]。此外,混合分解的养分释放还表现出不同的阶段性。有研究表明,混合凋落物对C元素的释放在第1年表现为抑制,而在第2年则表现为促进作用[35]。本研究中,混合凋落物叶N元素的释放速率实测值显著低于期望值,这可能是N含量高的芒箕向N含量低的毛竹进行了转移,对芒箕的N元素释放的促进作用低于毛竹的N元素释放的减缓作用,导致结果整体表现出混合分解对N释放的负效应。类似地,本研究中混合分解的部分阶段C元素和P元素未表现出明显的混合效应,这可能与凋落物混合对毛竹和芒箕C元素或P元素释放的作用(促进或减缓)相当有关,从而使整体未表现出混合效应。
-
单独分解过程中,毛竹凋落物叶分解速率低于林下植被芒箕,养分释放特征均表现为直接释放;混合分解过程中,毛竹和芒箕凋落物叶分解速率无显著混和效应,但抑制了N、P养分的释放,影响作用整体较为明显,而对C养分释放速率只在中末期有一定的抑制作用,影响作用相对较弱;毛竹和芒箕凋落物叶的C、N、P及其化学计量比的变化格局有所不同,呈现出C含量和C:N比整体下降,N含量和N:P比小幅上升,P含量微弱下降和C:P比波动性变化的趋势;毛竹和芒箕凋落物叶的分解速率与土壤温度呈极显著正相关,与土壤含水量无显著相关性,说明水热环境因子中土壤温度是影响本研究区域中凋落物分解速率的关键因素之一。
毛竹和林下植被芒箕凋落物分解特征研究
Decomposition Characteristics of Litter of Phyllostachys edulis and Dicranopteris pedata
-
摘要:
目的 探讨竹林与其林下植被凋落物叶之间相互影响的潜在机制,为合理经营管理毛竹林林下植被提供理论参考。 方法 采用原位分解袋法研究了四川长宁毛竹与林下植被芒箕凋落物叶分解和养分释放过程。 结果 (1)芒箕凋落物叶初始C、N、P含量和羟基碳高于毛竹(P < 0.05),而C:N、C:P、烷基碳、氧烷基碳和芳香碳低于毛竹(P < 0.05)。(2)凋落物叶分解和养分释放速率芒箕整体高于毛竹,芒箕和毛竹分解常数(k)分别为0.58±0.03和0.73±0.02,C、N、P养分释放均表现为净释放。(3)凋落物叶混合对分解速率没有显著影响,但抑制了N、P元素整个分解周期和C元素中后期的释放。(4)凋落物叶分解过程中元素含量变化格局表现为C含量和C:N比整体呈下降趋势,N含量和N:P比有小幅上升,P含量有微弱的下降趋势,C:P比呈波动性变化。(5)凋落物叶分解速率与土壤温度、初始凋落物叶N和P含量呈显著正相关(P < 0.01),与初始凋落物叶的C:N和C:P呈极显著负相关(P < 0.01),与土壤含水量相关不显著。 结论 单独分解过程中,毛竹凋落物叶分解速率低于林下植被芒箕,养分释放特征均表现为直接释放;混合分解过程中,毛竹和芒箕凋落物叶分解速率无显著混和效应,但养分释放的混合效应表现出一定负效应和不同阶段性。 Abstract:Objective To explore the potential mechanisms of interaction and function between bamboo forest and its understory vegetation's leaf litter, and provide a theoretical reference for the rational management of understory vegetation in Phyllostachys edulis forest. Method The litter in situ decomposition and nutrient release process of Ph. edulis and understory vegetation (Dicranopteris pedata) in Changning of Sichuan Province were investigated using litter bag. Result (1) The initial C, N, P contents and Carbonyl C of D. pedata were higher than those of Ph. edulis (P < 0.05), while C:N, C:P and Alkyl C, O-alkyl C and Aromatic C of D. pedata were all lower than those of P. edulis (P < 0.05). (2) The rate of litter decomposition and nutrient releasing of D. pedata were higher than that of Ph. edulis. The decomposition coefficient of Ph. edulis and D. pedata were 0.58±0.03 and 0.73±0.02, respectively. C, N and P nutrient release all showed net release. (3) The litter mixture of Ph. edulis and D. pedata had no significant effect on the decomposition rate, however, it significantly inhibited the release of N and P elements throughout the decomposition cycle and C elements in the late. (4) The change pattern of element content in litter decomposition process of Ph. edulis and D. pedata showed that C content and C:N ratio turned out a downward trend, and N content and N:P ratio increased slightly. P content showed a slight downward trend, and C:P ratio fluctuated. (5) Litter decomposition rate was positively correlated with soil temperature, initial litter N and P content (P < 0.01), and negatively correlated with C:N and C:P of initial litter (P < 0.01), but had no significant correlation with soil water content. Conclusion There was no significant mixed effect of decomposition rate, but the mixed effect of nutrient release showed certain negative effects and different periodic features. -
Key words:
- Phyllostachys edulis
- / Dicranopteris pedata
- / litter decomposition
- / nutrient release
- / mixing effects
-
图 1 单一凋落物分解过程中干质量和养分含量的变化(*表示种间差异显著(P < 0.05))
Figure 1. Single litter decomposition rates and nutrient dynamics duiring decomposition (*denote significant difference at P < 0.05 between two species)
图 2 混合凋落物干质量和养分的实测剩余率与期望剩余率(*表示实测值与期望值差异显著(P < 0.05))
Figure 2. Observed and expected mass and nutrient remaining in mixtures litter(*denote significant differences betweenthe observed and expected values(P < 0.05))
图 3 凋落物分解过程中化学计量特征(*表示实测值与期望值差异显著(P < 0.05))
Figure 3. Stoichiometric characteristics of litter decomposition(*denote significant differences between the observed and expected values(P < 0.05))
表 1 凋落物的初始化学组成
Table 1. Initial chemical composition of litter
凋落物类型Litter type C/(g·kg-1) N/(g·kg-1) P/(g·kg-1) C:N C:P N:P 毛竹P. edulis 415.78±0.15b 17.55±0.04b 0.204±0.001b 23.69±0.06a 2 042.40±8.08a 86.21±0.48a 芒箕D. pedata 421.64±0.29a 35.33±0.19a 0.404±0.001a 11.93±0.06b 1 044.06±2.13b 87.49±0.42a 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8. 下载: 导出CSV
下载: 导出CSV
表 2 凋落物的初始有机碳化学结构相对含量
Table 2. Proportions of the components of initial organic carbon functional group of litter
% 凋落物类型
Litter type烷基碳
Alkyl氧烷基碳
O-alkyl芳香碳
Aromatic羟基碳
Carbonyl烷基碳/氧烷基碳
Alkyl/o-alkyl毛竹P. edulis 15.02±0.12a 62.39±0.75a 15.11±0.21a 7.49±0.13b 24±0.27b 芒箕D. pedata 12.55±0.28b 24.81±0.44b 9.82±0.30b 52.81±0.95a 51±0.43a 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8.
下载: 导出CSV
表 3 凋落物干质量剩余率与时间的回归分析
Table 3. Regression analysis between dry weight residue of litter and time
类型
Type回归方程
Equation相关系数
R2年残留率
Remaining rate/%分解常数
k半衰期
t0.5/a周转期
t0.95/a毛竹P. edulis y = 0.790 3e-0.341t 0.999 56.10±1.77a 0.58±0.03b 1.21±0.07a 5.25±0.30a 芒箕D. pedata y = 0.630 6e-0.262t 0.984 48.03±0.87b 0.73±0.02a 0.95±0.02b 4.10±0.10b 注:同列不同字母表示差异显著(P < 0.05),n=8。
Note: Values followed with the different letters within a column mean sinificant difference(P < 0.05),n=8.
下载: 导出CSV
表 4 凋落物干质量损失量与土壤含水量、温度及初始凋落物质量的Pearson相关系数
Table 4. Pearson correlation coefficient between dry mass loss of litter and soil water content and temperature, the initial quality
类型
Type土壤含水量
Soil water content土壤温度
Soil temperatureC N P C:N C:P N:P 毛竹P. edulis -0.247 0.783** 0.154 0.572* 0.521** -0.411* -0.552* 0.178 芒箕D. pedata -0.311 0.809** 0.123 0.504* 0.373* -0.394* -0.379* 0.172 毛竹和芒箕P. edulis&D. pedata -0.290 0.751* 0.253 0.634** 0.437* -0.517* -0.430* 0.204 注:*表示显著水平P < 0.05;**表示显著水平P < 0.01.
Note: *mean sinificant difference(P < 0.05),**mean sinificant difference(P < 0.01).
下载: 导出CSV
-
[1] Wardle D A, Hörnberg G, Zackrisson O, et al. Long-term effects of wildfire on ecosystem properties across an island area gradient[J]. Science, 2003, 300(5621):972-975. doi: 10.1126/science.1082709 [2] Wardle D A, Nilsson M C, Zackrisson O, et al. Determinants of litter mixing effects in a Swedish boreal forest[J]. Soil Biology & Biochemistry, 2003, 35(6):827-835. [3] 詹鸿振, 刘传照, 刘吉春, 等.小兴安岭阔叶红松林主要树种凋落物的分解研究[J].东北林业大学学报, 1990, 18(3):1-8. [4] Hättenschwiler S, Tiunov A V, Scheu S. Biodiversity and Litter Decomposition in Terrestrial Ecosystems[J]. Annual Review of Ecology Evolution & Systematics, 2005, 36(36):191-218. [5] Lummer D, Scheu S, Butenschoen O. Connecting litter quality, microbial community and nitrogen transfer mechanisms in decomposing litter mixtures[J]. Oikos, 2012, 121(10):1649-1655. doi: 10.1111/more.2012.121.issue-10 [6] S Linnea Berglund, GÅran I ögren, Alf Ekblad. Carbon and nitrogen transfer in leaf litter mixtures[J]. Soil Biology and Biochemistry, 2013, 57(341):341-348. [7] 胡亚林, 汪思龙, 黄宇, 等.凋落物化学组成对土壤微生物学性状及土壤酶活性的影响[J].生态学报, 2005, 25(10):2662-2668. doi: 10.3321/j.issn:1000-0933.2005.10.030 [8] Kubartová A, Ranger J, Berthelin J, et al. Diversity and decomposing ability of saprophytic fungi from temperate forest litter[J]. Microbial Ecology, 2009, 58(1):98-107. [9] Malosso E, English L, Hopkins D W, et al. Use of 13C-labelled plant materials and ergosterol, PLFA and NLFA analyses to investigate organic matter decomposition in Antarctic soil.[J]. Soil Biology & Biochemistry, 2004, 36(1):165-175. [10] Liu P, Huang J, Han X, et al. Litter Decomposition in Semiarid Grassland of Inner Mongolia, China[J]. Rangeland Ecology & Management, 2009, 62(4):305-313. [11] Tardif A, Shipley B. Using the biomass-ratio and idiosyncratic hypotheses to predict mixed-species litter decomposition[J]. Annals of Botany, 2013, 111(1):135. doi: 10.1093/aob/mcs241 [12] Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter[J]. Oikos, 2004, 104(2):230-246. doi: 10.1111/oik.2004.104.issue-2 [13] 李宜浓, 周晓梅, 张乃莉, 等.陆地生态系统混合凋落物分解研究进展[J].生态学报, 2016, 36(16):4977-4987. [14] 江泽慧.世界竹藤[M].沈阳:辽宁科学技术出版社, 2002. [15] 高平珍, 陈双林.毛竹林下植被管理与利用研究综述[J].竹子研究汇刊, 2017, 36(2):44-48. doi: 10.3969/j.issn.1000-6567.2017.02.007 [16] 刘广路, 范少辉, 蔡春菊, 等.撑绿杂交竹和硬头黄竹克隆生长特性比较[J].植物学报, 2013, 48(3):288-294. [17] Olson J S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems[J]. Ecology, 1963, 44(2):322-331. doi: 10.2307/1932179 [18] 陈瑾, 李扬, 黄建辉.内蒙古典型草原4种优势植物凋落物的混合分解研究[J].植物生态学报, 2011, 35(1):9-16. [19] Jhc C. An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types.[J]. Journal of Ecology, 1996, 84(4):573-582. doi: 10.2307/2261479 [20] 杨玉盛, 郭剑芬, 陈银秀, 等.福建柏和杉木人工林凋落物分解及养分动态的比较[J].林业科学, 2004, 40(3):19-25. doi: 10.3321/j.issn:1001-7488.2004.03.003 [21] Tian X J, Takahiro T. Relative roles of microorganisms and soil animals on needel litter decomposition in a subalpine coniferous forest J]. Acta Phytoecologica Sinica, 2002, 26(3):257-263. [22] Garcia-Pausas J, Casals P, Romanyà J. Litter decomposition and faunal activity in Mediterranean forest soils:effects of N content and the moss layer[J]. Soil Biology & Biochemistry, 2004, 36(6):989-997. [23] Sanchez F G.Loblolly pine needle decomposition and nutrient dynamics as affected by irrigation, fertilization, and substrate quality[J]. Forest Ecology & Management, 2001, 152(1):85-96. [24] Mendonça E S, Stott D E. Characteristics and decomposition rates of pruning residues from a shaded coffee system in Southeastern Brazil[J]. Agroforestry Systems, 2003, 57(2):117-125. doi: 10.1023/A:1023900822261 [25] 施妍, 陈芳清.大老岭自然保护区日本落叶松林凋落物分解及养分释放研究[J].林业科学研究, 2016, 29(3):430-435. doi: 10.3969/j.issn.1001-1498.2016.03.019 [26] Preston C M, Trofymow J A. Variability in litter quality and its relationship to litter decay in Canadian forests.[J]. Canadian Journal of Botany, 2000, 78(10):1269-1287. doi: 10.1139/b00-101 [27] 杨万勤, 邓仁菊, 张健.森林凋落物分解及其对全球气候变化的响应[J].应用生态学报, 2007, 18(12):2889-2895. [28] Berg B, Mcclaugherty C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration[M]. Berlin:Springer-Verlag, 2008. [29] 李正才, 徐德应, 杨校生, 等.北亚热带6种森林类型凋落物分解过程中有机碳动态变化[J].林业科学研究, 2008, 21(5):675-680. doi: 10.3321/j.issn:1001-1498.2008.05.015 [30] Einhellig F A, Rasmussen J A, Hejl A M, et al. Effects of root exudate sorgoleone on photosynthesis[J]. Journal of Chemical Ecology, 1993, 19(2):369-375. doi: 10.1007/BF00993702 [31] Knops J M H, Wedin D, Tilman D. Biodiversity and decomposition in experimental grassland ecosystems.[J]. Oecologia, 2001, 126(3):429-433. doi: 10.1007/s004420000537 [32] Rustad L E, Cronan C S. Element loss and retention during litter decay in a red spruce stand i.[J]. Canadian Journal of Forest Research, 1988, 18(7):947-953. doi: 10.1139/x88-144 [33] Hansen R A, Coleman D C. Litter complexity and composition are determinants of the diversity and species composition of oribatid mites (Acari:Oribatida) in litterbags.[J]. Applied Soil Ecology, 1998, 9(1-3):17-23. doi: 10.1016/S0929-1393(98)00048-1 [34] Tlalka M, Bebber D P, Darrah P R, et al. Emergence of self-organised oscillatory domains in fungal mycelia[J]. Fungal Genetics & Biology, 2007, 44(11):1085-1095. [35] 蒋云峰.长白山针阔混交林主要凋落物分解及土壤动物的作用[D].长春: 东北师范大学, 2013. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4027
- HTML全文浏览量: 1805
- PDF下载量: 402
- 被引次数: 0
