

-
林窗式疏伐指通过砍伐特定的冠层林木形成林冠空窗,是一种模拟林窗干扰的近自然林业措施,对森林可持续经营管理具有重要意义。林窗式疏伐降低了余留木的竞争,改变了资源可利用性,进而影响其生长[1-2]。叶作为林木主要的同化器官,其形态和生理特性对林窗式疏伐的响应是决定疏伐后余留木生长响应的关键因素。常绿树种包含从几天到几年不等叶龄的叶片,因此同化过程比落叶树种更为复杂[3-4]。之前的研究多关注于当年生叶光合特性对疏伐的响应,对多年生叶了解较少。然而多年生叶在常绿树种叶总生物量中占有很大的比例[3],对整株林木生长发挥重要作用[5]。因此,了解余留木各个叶龄叶光合特性对林窗式疏伐的响应对全面评估其响应机制和准确预估余留木生长动态具有重要意义。
林窗式疏伐后余留木叶光合特性的改变主要是由其光照、水分和养分条件的改变导致的。光照强度、光照时长和光合有效辐射均随着林窗面积增大而升高[6-8],温度、土壤水分和养分可利用性也与林窗大小紧密相关[9-10]。通常大林窗对余留木生长的促进效应高于小林窗[10]。因此,研究叶光合特性对不同大小林窗疏伐的响应差异可为林业经营管理措施的制定提供理论依据。
叶光合特性对林窗式疏伐的响应主要分为两种方式。一是通过改变叶的形态或者解剖特征更大程度的增加光捕获,提高光合速率。比叶重(LMA)是反映叶光捕获能力的一个重要指标。前人研究表明疏伐通常会增加叶的厚度、密度、叶肉细胞的数量或改变叶肉细胞的排列等[1-2, 11],导致LMA升高。另外一种是通过增加叶氮含量或者改变氮在捕光组分、1, 5-二磷酸核酮糖羧化酶和生物力能学组分中的分配比例[12-13],提高羧化速率和电子传导速率[14]。然而,不同年龄叶形态、化学组成和生理生态特性具有明显差异[15-16]。例如,LMA随着叶龄增大而升高[3-4, 17],但是叶形态可塑性、氮含量和光合氮利用效率随着叶龄增大而逐渐降低[4]。因此,不同叶龄叶形态和生理特性对林窗式疏伐的响应可能存在很大差异。Li等[18]利用双同位素方法发现疏伐导致当年生和1年生叶光合速率的增加,但是对2~4年生叶影响不显著。但是有研究却发现疏伐后最大净光合速率(Amax)的提高主要表现在成熟叶(<2年叶)和老龄叶上(>2年生叶),而对当年生叶影响不明显[1, 19]。
华山松(Pinus armandii Franch.)和油松(Pinus tabuliformis Carr.)是我国青藏高原东缘亚高山地区常见的栽培树种,叶寿命2~4年[20]。各叶龄针叶光合特性对林窗式疏伐的响应,是解释华山松和油松生长响应差异的重要方面。本研究通过对比研究华山松和油松当年生、1年生、2年生叶LMA、叶绿素含量、叶氮含量、Amax、光合氮利用效率(PNUE)对林窗式疏伐的响应差异,探讨林窗大小、叶龄和树种对林窗式疏伐后叶光合特性响应的影响。
-
试验样地选在四川省阿坝州茂县凤仪镇的中国科学院茂县山地生态定位研究站(103°54′ E, 31°42′ N)。地处青藏高原东缘、横断山系北段高山峡谷地带的岷江上游大沟流域。暖温带气侯,年均气温9.3℃,最热月7月份平均气温18.6℃,最冷月1月份平均气温-0.9℃。年均日照时数为1 373.8 h,年均降水量825.2 mm,降水多集中在4—10月,约占全年降水量的70%。土壤类型主要为褐土和棕壤[21]。
-
选择30年生的华山松和油松混交人工林为研究对象。2008年7月中旬对林分进行调查,林分郁闭度92%,密度为2 321株·hm-2,平均胸径15.4 cm,平均树高11.2 m。林下灌木、草本较少,盖度约为8%,其中主要优势种有黄鹤菜(Youngia japonica (L.) DC.)、紫花堇菜(Viola grypoceras A.Gray)、假升麻(Aruncus sylvester Kostel.)、四川鹿蹄草(Pyrola szechuanica H. Andr.)、小叶蔷薇(Rosa willmottiae Hemsl.)等。
试验采用完全随机区组设计。共分为3个区组,每个区组设置3个20 m×15 m的样方,随机布设对照(CK,不疏伐)、小林窗(SG,80 m2)、中林窗(IG,110 m2)处理。林窗面积是根据森林中自然形成的、常见的林窗大小确定[22]。林窗式疏伐在2008年11月进行,从距离地面50 cm处砍伐提前标记的样木,砍伐下来的干、枝叶全部移出样地,林下灌草不处理。
-
2013年,在对照样地和林窗北侧边缘各选择2棵长势较好的华山松和油松作为研究对象。将每棵树树冠沿树干平均分为上、下两层。之前研究表明树冠下层叶对疏伐的响应比上层叶更明显[22-24],为了更好的对比林窗效应,本研究选择树冠中下层位置面向林窗一侧枝条上的叶进行取样和测定。根据叶着生特点、颜色及特性将叶分为3个叶龄:当年生、1年生、2年生[17]。
光合参数测定是在2013年8月12—16日09:00—12:00,14:00—16:00进行。测定方法为离体标准枝法。每棵树每个叶龄取3个标准枝,标准枝取下立即放入盛水的容器内,并尽快测定,减少离体对测定的影响[23]。利用Li-6400便携式光合系统测定仪选用针叶叶室(Li-6400;Li-Cor Inc., NE, USA),温度设定22℃(接近大气温度),相对湿度40%,CO2浓度380 mg·L-1,流速为500 μmol ·s-1。光照强度1 600(μmol ·m-2 ·s-1)下稳定后记录最大净光合速率(Amax)。光合速率测定后的叶取下,用扫描仪LiDE120(CanoScan,Japan)结合Image J软件(National Institutes of Health, Bethesda, MD, USA)计算用于气体交换的植物叶片叶面积。然后将叶放入烘箱,60℃下烘48 h,测定其干质量。针叶干质量与其叶面积的比值计算得出LMA。
另外,在华山松和油松所取标准枝靠近光合测定部位再收集各叶龄叶50 g。其中5 g低温保存(4℃)带回实验室,用于测定单位面积叶绿素a(Chl a)和叶绿素b含量(Chl b)。采用纯丙酮和无水乙醇按2:1配成混合提取液,分光光度计下测定[24]。叶绿素比值(Chl a:b)是叶绿素a和叶绿素b的比值。余下45 g放入烘箱,60℃下烘48 h。然后进行粉碎、过筛,用元素分析仪(Vario Macro Cube, Elementar, Germany)测定氮含量。光合氮利用效率(PNUE)是单位面积叶最大净光合速率(Amax)与单位面积氮含量(N)的比值。
-
采用线性混合模型分析林窗大小、叶龄、树种对叶LMA、Chl a、Chl b、Chl a+b、Chl a:b、N、Amax、PNUE的影响。林窗大小、叶龄、树种及其交互作用为固定效应参数,区组为随机效应参数。某特定年龄叶某一性状在不同大小林窗式疏伐间的差异用单因素分析方法(ANOVA,多重比较选用Duncan, P < 0.05)。所有的统计和图在SPSS 18.0和OriginPro 8.0完成。
-
随着叶龄增大,叶LMA逐渐升高,Amax和PNUE逐渐降低(表 1,图 1和图 2)。不同叶龄叶光合特性对林窗疏伐的响应具有明显差异:华山松、油松当年生叶LMA、Chl a、Chl b、Chl a+b、Chl a:b、N、Amax、PNUE在不同林窗疏伐处理间差异均不显著,但是其1年生和2年生叶光合特性对林窗疏伐响应明显(表 1和图 1~4)。
表 1 叶光合特性线性混合模型分析的F值
Table 1. F values based on linear mixed-effect model analyses for leaf photosynthetic characteristics
项目
Items比叶重
LMA叶绿素a
Chl a叶绿素b
Chl b叶绿素a+b
Chl a+b叶绿素a:b
Chl a:b氮含量
N最大净光合速率Amax 光合氮利用效率PNUE 树种Species 28.097** 44.412** 55.007** 46.437** 5.865* 14.948** 5.616* 1.315 叶龄Age 27.315** 3.870* 5.205** 4.574* 14.558** 0.684 33.365** 12.770** 林窗大小Gap size 0.804 5.135* 6.770* 5.748* 4.215* 1.379 4.026* 1.554 树种×叶龄Species×Age 0.518 2.075 2.739 2.047 0.772 1.836 0.342 0.828 树种×林窗大小
Species×Gap size0.474 1.202 0.956 1.112 0.612 0.212 0.165 1.425 年龄×林窗大小
Age×Gap size1.091 3.457* 3.920** 3.951** 0.154 1.954 1.323 1.935 树种×年龄×林窗大小
Species×Age×Gap size1.093 1.394 1.320 1.396 1.248 1.034 0.896 1.531 *P < 0.05, **P < 0.01 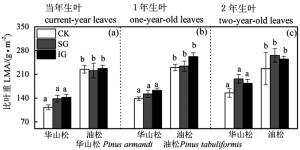
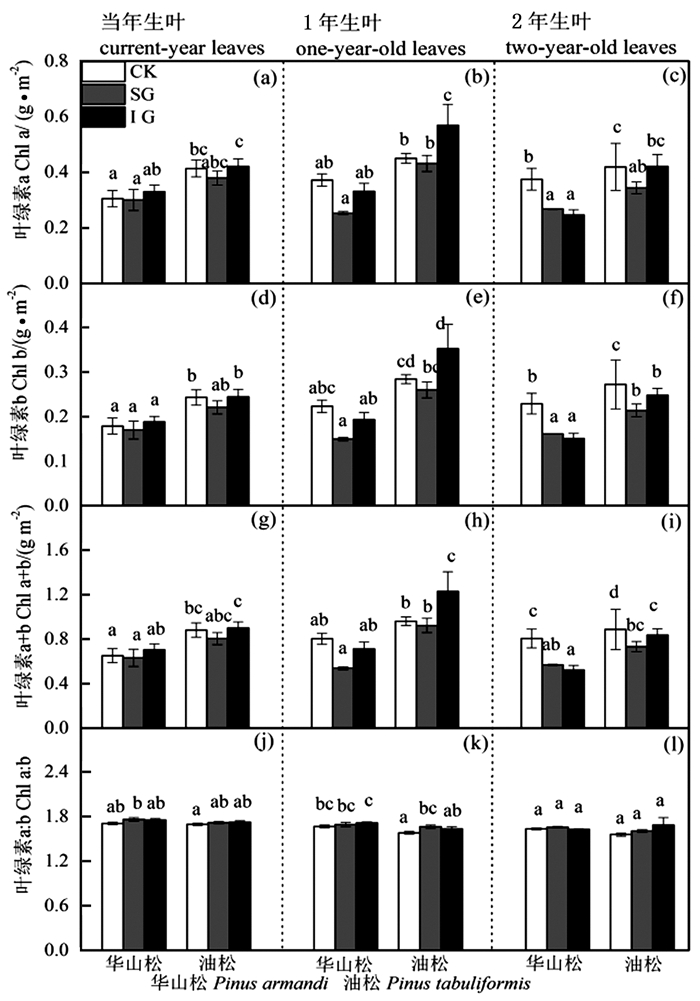
图 1 华山松和油松比叶重LMA对林窗式疏伐的响应
Figure 1. LMA responses of Pinus armandii and P. tabuliformis to gap-model thinning
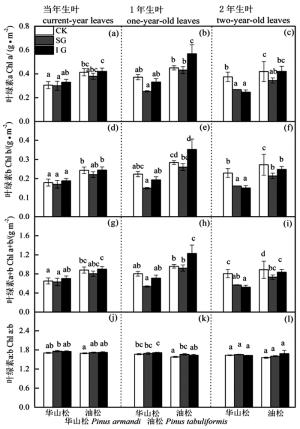
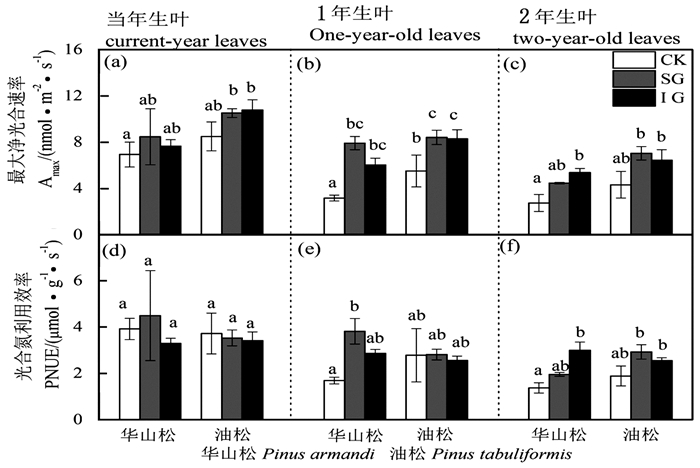
图 2 华山松和油松最大净光合速率Amax和光合氮利用效率PNUE对林窗式疏伐的响应
Figure 2. Leaf Amax and PNUE responses of Pinus armandii and P. tabuliformis to gap-model thinning

图 3 华山松和油松叶绿素含量对林窗式疏伐的响应
Figure 3. Leaf chlorophyll responses of Pinus armandii and P. tabuliformis to gap-model thinning

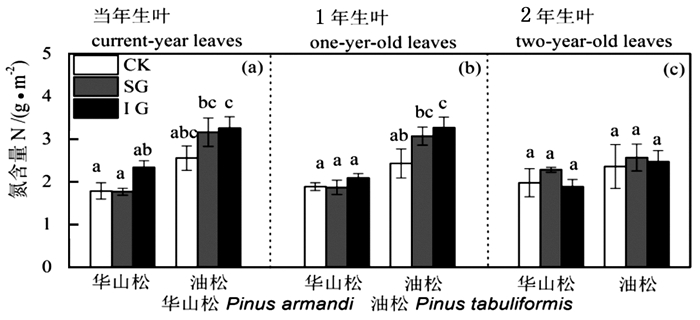
图 4 华山松和油松叶氮含量N对林窗式疏伐的响应
Figure 4. Leaf nitrogen content responses of Pinus armandii and P. tabuliformis to gap-model thinning
华山松1年生叶LMA、Chl a、Chl b、Chl a+b、Chl a:b、N含量在不同林窗疏伐处理间均无显著差异(图 1、3、4),SG显著提高了其Amax和PNUE(图 2b)。IG和SG边缘华山松2年生叶LMA、N含量与对照林木差异不显著(图 1c和图 4c),但其叶绿素含量显著低于对照(图 3),Amax和PNUE高于对照(图 2c)。
油松叶LMA、叶绿素含量、N、Amax高于华山松,油松1年生和2年生叶对林窗疏伐的响应也与华山松存在很大差异。油松1年生叶LMA和PNUE在对照样地和林窗样地差异不显著(图 1b和图 2b),但IG边缘油松1年生叶叶绿素和N含量显著高于CK(图 3和图 4),IG和SG边缘叶Amax显著高于CK(图 2b)。油松2年生叶LMA、N含量在不同林窗疏伐处理间没有显著差异(图 1c和图 4c),但SG边缘油松1年生叶绿素含量低于CK(图 3),Amax和PNUE略高于CK(图 2c和图 2f)。
-
林窗式疏伐后IG(110 m2)和SG(80 m2)边缘木叶Amax显著高于对照(图 2),叶绿素含量、N含量和PNUE也与对照林木有显著差异。显然林窗式疏伐改变了边缘木叶光合特性,影响了其光合潜力。这跟前人的研究结果一致,疏伐导致了叶Amax的升高[2, 25-27]。尽管叶Amax的提高并不直接代表生长效应,但Amax可以反映边缘木是否能有效利用疏伐后增加的辐射[2]。该结果表明华山松和油松混交人工林中80~110 m2的林窗疏伐就能明显改善林木光合潜力,影响林木生长。因此,林业经营管理中要根据树种和林分特性制定合适的管理措施。
IG边缘木叶Amax、LMA、叶绿素含量、N含量、PNUE与SG均无显著差异,表明本林分中叶光合特性对林窗式疏伐的响应没有表现出随着林窗面积增大而增强的趋势。叶光合特性的改变本质上是对林窗式疏伐后光照、水分和养分条件的适应性结果[1]。本研究设置的110 m2与80 m2林窗边缘木叶光合特性差异不明显可能是由于两个林窗间环境异质性不大,未来需要拉大林窗面积差距,研究更小或者更大的林窗面积对树木生长的影响,更好的指导林业实践。
-
本研究发现随着叶龄增长,LMA升高,Amax和PNUE降低(图 1和图 2)。这是因为老叶气孔导度小、电子传递速率和羧化速率低,光组分氮分配比例低[28],是由自身生物学特性决定的。
不同叶龄叶对林窗式疏伐响应具有明显差异。当年叶LMA、叶绿素含量、N、Amax、PNUE对林窗式疏伐响应均不显著,而林窗边缘木1年生叶、2年生叶Amax、叶绿素、N、PNUE与对照林木有显著差异。Medhurst等[1]和Goudiaby等[17]研究结果也发现当年生叶光合速率对疏伐响应不明显。这可能主要是因为当年生叶生理活性高,Amax和PNUE高,且多生于枝的顶端,生长受到资源的限制较少,因此疏伐后光照和养分条件的改善对其影响不显著。然而随着叶龄增长,光合参与酶活性降低[29],光合速率受到电子传递速率和羧化速率的限制[28]。林窗疏伐后多年生叶氮含量或者氮的利用效率增加,光合潜力提高。不同年龄叶对疏伐差异性响应结果表明在研究某些常绿树种对疏伐或者其他林业管理措施响应时仅关注当年生叶是片面和不合理的,应该综合考虑各个叶龄叶,结合各个叶龄叶所占的比例,全面评估生长响应。
-
华山松叶LMA、叶绿素含量、N、Amax均低于油松(图 1、3、4),之前研究也发现华山松光饱和点和Amax低于油松[20-31]。表明华山松单位面积叶光捕获能力和光合潜力低于油松。华山松与油松叶对林窗式疏伐的响应也明显不同:林窗边缘华山松1年生、2年生叶Amax和PNUE高于对照,而LMA和N含量与对照没有显著差异(图 2、4)。该结果表明对于华山松多年生叶来说,林窗式疏伐处理并没有改变氮含量,而是提高了光合氮利用效率,促进光合速率的升高。光合氮利用效率的提高可能主要是通过改变氮在在捕光组分、1, 5-二磷酸核酮糖羧化酶和生物力能学组分中的分配比例,华山松林窗边缘木2年生叶叶绿素含量显著低于对照(图 3)也间接证明华山松减少了对补光组分氮的分配。这可能是因为疏伐后边缘木光照条件改善[1, 26],光捕获不再是限制其光合速率的首要因素,而多年生叶暗反应阶段低的羧化酶生理活性和低的光量子传导速率[28-29],限制其同化速率。因此疏伐后华山松减少对光捕获的投入,将更多的氮投入到暗反应阶段,从而提高光合氮利用效率和光合潜力。
林窗边缘油松1年生叶N、叶绿素含量和Amax显著高于对照。这与Medhurst等[2]结论一致,疏伐后叶氮含量提高,光合速率增加。疏伐后光照和养分条件的改善,加速了叶绿素、光合反应酶、羧化酶等的合成,进而提高光合潜力。不同树种对林窗式疏伐的响应差异跟其生物学特性及资源利用能力紧密相关。
-
80 m2和110 m2的林窗疏伐显著改变叶光合特性,提高其Amax,但两个林窗对叶特性的影响差别不显著。林窗疏伐对叶光合特性的影响主要集中在1年生和2年生叶,对当年生叶影响不显著。华山松和油松1年生和2年生叶对林窗式疏伐的响应存在差异:华山松主要是提高氮利用效率,提高光合潜力,而油松则主要提高叶氮含量,促进光合速率的加快。
华山松、油松不同叶龄针叶光合特性对林窗式疏伐的响应
Differential Responses of Age-related Leaf Photosynthetic Characteristics of Pinus armandii and Pinus tabuliformis to Gap-model Thinning
-
摘要:
目的 通过对比研究华山松和油松不同叶龄针叶形态、光合速率、叶绿素和叶氮含量对林窗式疏伐的响应,探讨林窗大小、叶龄和树种对林窗式疏伐后叶光合特性响应的影响。 方法 2008年在30年生华山松和油松混交人工林中,设置对照、小林窗(80 m2)和中林窗(110 m2)处理,2013年以对照样地内和林窗边缘华山松和油松为研究对象,测定其当年生、1年生和2年生叶比叶重(LMA)、单位叶面积叶绿素a(Chl a)、叶绿素b含量(Chl b)、氮含量(N)、最大净光合速率(Amax)、光合氮利用效率(PNUE)等指标。 结果 小林窗和中林窗疏伐对叶光合特性影响一致,均对当年生叶影响不显著,但显著改变了1年生和2年生叶绿素含量、N和PNUE,提高了其Amax。华山松叶LMA、叶绿素含量、N、Amax均低于油松,两树种多年生叶对林窗式疏伐的响应也明显不同:林窗边缘华山松1年生和2年生叶LMA和N与对照差异不显著,但叶绿素含量低于对照,PNUE和Amax显著高于对照;林窗边缘油松1年生和2年生叶LMA和PNUE与对照差异不显著,但N和Amax显著高于对照。 结论 80~110 m2林窗疏伐显著改变叶光合特性,提高其光合潜力;林窗疏伐影响1年生和2年生叶光合特性,对当年生叶影响不显著;华山松和油松对林窗式疏伐的响应特征存在差异。 Abstract:Objective To study the responses of age-related leaf morphology, photosynthetic rate, photosynthetic pigments, and leaf nitrogen of Pinus armandii and P. tabuliformis to gap-model thinning, intending to explore the influences of gap size, leaf age, and species on leaf characteristics after thinning. Method The small gaps (80 m2), intermediate gaps (110 m2) and a control were set in a 30-year-old mixed P. tabuliformis and P. armandii plantation in 2008. The P. armandii and P. tabuliformis in control plots and gap-edged pines were chosen as test materials, and the leaf mass per area (LMA), area-based chlorophyll a, chlorophyll b, nitrogen content (N), maximum photosynthetic rate (Amax), and photosynthetic nitrogen use efficiency (PNUE) of current year, one-year-old, and two-year-old leaves were measured in 2013. Result Small and intermediate gaps had no significant influences on current-year leaves of P. armandii and P. tabuliformis, while significantly altered chlorophyll, N content, and PNUE, enhanced Amax of one-year-old and two-year-old leaves. P. armandii had lower LMA, chlorophyll content, N, and Amax. Moreover, older leaves of P. armandii and P. tabuliformis showed differentiated responses to gap-model thinning:gap-edged P. armandii showed lower chlorophyll content, higher Amax and PNUE than the control, while the LMA and N were no significantly different with the control; gap-edged P. tabuliformis had higher Amax and N than the control, while LMA and PNUE had no obvious difference with the control. Conclusion Gaps in size of 80-110m2 can alter the leaf characteristics and improve its photosynthetic capacity. Gap-model thinning has significant influences on the older leaves, while has no influence on current leaves. P. armandii and P. tabuliformis have different responding strategies to gap-model thinning. -
图 1 华山松和油松比叶重LMA对林窗式疏伐的响应
Figure 1. LMA responses of Pinus armandii and P. tabuliformis to gap-model thinning
图 2 华山松和油松最大净光合速率Amax和光合氮利用效率PNUE对林窗式疏伐的响应
Figure 2. Leaf Amax and PNUE responses of Pinus armandii and P. tabuliformis to gap-model thinning
图 3 华山松和油松叶绿素含量对林窗式疏伐的响应
Figure 3. Leaf chlorophyll responses of Pinus armandii and P. tabuliformis to gap-model thinning
图 4 华山松和油松叶氮含量N对林窗式疏伐的响应
Figure 4. Leaf nitrogen content responses of Pinus armandii and P. tabuliformis to gap-model thinning
表 1 叶光合特性线性混合模型分析的F值
Table 1. F values based on linear mixed-effect model analyses for leaf photosynthetic characteristics
项目
Items比叶重
LMA叶绿素a
Chl a叶绿素b
Chl b叶绿素a+b
Chl a+b叶绿素a:b
Chl a:b氮含量
N最大净光合速率Amax 光合氮利用效率PNUE 树种Species 28.097** 44.412** 55.007** 46.437** 5.865* 14.948** 5.616* 1.315 叶龄Age 27.315** 3.870* 5.205** 4.574* 14.558** 0.684 33.365** 12.770** 林窗大小Gap size 0.804 5.135* 6.770* 5.748* 4.215* 1.379 4.026* 1.554 树种×叶龄Species×Age 0.518 2.075 2.739 2.047 0.772 1.836 0.342 0.828 树种×林窗大小
Species×Gap size0.474 1.202 0.956 1.112 0.612 0.212 0.165 1.425 年龄×林窗大小
Age×Gap size1.091 3.457* 3.920** 3.951** 0.154 1.954 1.323 1.935 树种×年龄×林窗大小
Species×Age×Gap size1.093 1.394 1.320 1.396 1.248 1.034 0.896 1.531 *P < 0.05, **P < 0.01  下载: 导出CSV
下载: 导出CSV
-
[1] Jones T A, Thomas S C. Leaf-level acclimation to gap creation in mature Acer saccharum trees[J]. Tree Physiology, 2007, 27(2):281-290. doi: 10.1093/treephys/27.2.281 [2] Medhurst J L, Beadle C L. Photosynthetic capacity and foliar nitrogen distribution in Eucalyptus nitens is altered by high-intensity thinning[J]. Tree Physiology, 2005, 25(8):981-991. doi: 10.1093/treephys/25.8.981 [3] Li M H, Krauchi N, Dobbertin M. Biomass distribution of different-aged needles in young and old Pinus cembra trees at highland and lowland sites[J]. Trees-Structure and Function, 2006, 20(5):611-618. doi: 10.1007/s00468-006-0076-0 [4] Sellin A. Morphological and stomatal responses of Norway spruce foliage to irradiance within a canopy depending on shoot age[J]. Environment Experiment and Botony, 2001, 45(2):115-131. doi: 10.1016/S0098-8472(00)00086-1 [5] Drenkhan R, Kurkela T, Hanso M, et al. The relationship between the needle age and the growth rate in Scots pine (Pinus sylvestris): a retrospective analysis by needle trace method (NTM)[J]. European Journal of Forest Research, 2006, 125(4):397-405. doi: 10.1007/s10342-006-0131-9 [6] Canhanm C D, Denslow J S, Plantt W J, et al. Light regimes beneath closed canopies and tree-fall gaps in tmperate and tropical forests[J]. Canadian Journal of Forest Research, 1990, 20(5):620-631. doi: 10.1139/x90-084 [7] Gray A N, Spies T A, Easter M J, et al. Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests[J]. Canadian Journal of Forest Research, 2002, 32(2):332-343. doi: 10.1139/x01-200 [8] 陈梅, 朱教君, 闫巧玲, 等.辽东山区次生林不同大小林窗光照特征比较[J].应用生态学报, 2008, 19(12):2555-2560. [9] 刘聪, 朱教君, 吴祥云, 等.辽东山区次生林不同大小林窗土壤养分特征[J].东北林业大学学报, 2011, 39(1):79-81. doi: 10.3969/j.issn.1000-5382.2011.01.025 [10] Zhao Q X, Pang X Y, Bao W K, et al. Effects of gap-model thinning intensity on the radial growth of gap-edge trees with distinct crown classes in a spruce plantation[J]. Tree-Structure and Function, 2015, 29(6):1861-1870. doi: 10.1007/s00468-015-1267-3 [11] Rabelo G R, Vitória Ậ P, da Silva M V A, et al. Structural and ecophysiological adaptations to forest gaps[J]. Trees, 2013, 27(1):259-272. doi: 10.1007/s00468-012-0796-2 [12] Chen J W, Kuang S B, Long G Q, et al. Steady-state and dynamic photosynthetic performance and nitrogen partitioning in shade-demanding plant Panax notoginseng under different levels of growth irradiance[J]. Acta Physiologiace Plantarum, 2014, 36(9):2409-2420. doi: 10.1007/s11738-014-1614-9 [13] 唐敬超, 史作民, 罗达, 等.遮荫处理对灰木莲幼苗叶片光合氮利用效率的影响[J].生态学报, 2017, 37(22):1-10. [14] Han Q, Chiba Y. Leaf photosynthetic responses and related nitrogen changes associated with crown reclosure after thinning in a young Chamaecyparis obtusa stand[J]. Journal of Forest Research, 2009, 14(6):349-355. doi: 10.1007/s10310-009-0146-4 [15] Puchalska E, Czajkowska B, Kielkiewicz M. Morphological, anatomical and chemical characterization of white spruce (Picea glauca 'Conica') differently aged needles and hypotheses on their influence on Oligonychus ununguis infestation[J]. Acta Physiologiae Plantarum, 2008, 30(2):225-232. doi: 10.1007/s11738-007-0111-9 [16] Li C, Wu C, Duan B, et al. Age-related nutrient content and carbon isotope composition in the leaves and branches of Quercus aquifolioides along an altitudinal gradient[J]. Trees, 2009, 23(5):1109-1121. doi: 10.1007/s00468-009-0354-8 [17] Mediavilla S, Gonzalez-Zurdo P, Garcia-Ciudad A, et al. Morphological and chemical leaf composition ofMediterranean evergreen tree species according to leaf age[J].Trees, 2011, 25(4):669-677. doi: 10.1007/s00468-011-0544-z [18] Li R S, Yang Q P, Zhang W D, et al. Thinning effect on photosynthesis depends on needle ages in a Chinese fir (Cunninghamia lanceolata) plantation[J]. Science of the Total Environment, 2017, 580:900-906. doi: 10.1016/j.scitotenv.2016.12.036 [19] Goudiaby V, Brais S, Grenier Y, et al. Thinning effects on Jack Pine and Black Spruce photosynthesis in Eastern Boreal Forests of Canada[J]. Silva Fennica, 2011, 45(4):595-609. [20] 衣宁, 赵文倩, 刘倩, 等.油松新生叶与老叶光合功能的比较[J].林业科技, 2014, 39(6):10-14. [21] 赵庆霞, 包维楷.华山松和油松冠型结构对林窗式疏伐的响应[J].四川农业学报, 2016, 34(1):24-28. [22] 谭辉, 朱教君, 康宏樟, 等.林窗干扰研究[J].生态学杂志, 2007, 26(4):587-594. doi: 10.3321/j.issn:1000-4890.2007.04.025 [23] 唐艳, 王传宽.东北主要树种光合作用可行的离体测定方法[J].植物生态学报, 2011, 35(4):452-462. [24] 刘秀丽, 宋平, 孙成明, 等.植物叶绿素测定方法的探讨[J].江苏农业研究, 1999, 20(3):46-47. [25] Gravatt D A, Chambers J L, Barnett J P, et al. Temporal and spatial patterns of net photosynthesis in 12-year-old loblolly pine five growing seasons after thinning[J]. Forest Ecology and Management, 1997, 97(1):73-83. doi: 10.1016/S0378-1127(97)00055-8 [26] Peterson J A, Seiler J R, Nowak J, et al. Growth and physiological responses of young loblolly pine stands to thinning[J]. Forest Science, 1997, 43(4):529-534. [27] Tang Z M, Chambers J L, Sword M A, et al. Seasonal photosynthesis and water relations of juvenile loblolly pine relative to stand density and canopy position[J]. Trees-Structure and Function, 2003, 17(5):424-430. doi: 10.1007/s00468-003-0256-0 [28] Niinemets ü, Cescatti A, Rodeghiero M. Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species[J]. Plant Cell and Environment, 2010, 28(12):1552-1566. [29] Ethier G J, Livingston N J, Harrison D L, et al. Low stomatal and internal conductance to CO2 versus Rubisco deactivation as determinants of the photosynthetic decline of ageing evergreen leaves[J]. Plant Cell and Environment, 2010, 29(12):2168-2184. [30] 李勇, 韩海荣, 唐峰峰, 等.油松人工林冠层光合生理特性的空间异质性[J].东北林业大学学报, 2013, 41(4):32-35. doi: 10.3969/j.issn.1000-5382.2013.04.008 [31] 梁军生, 陈晓鸣, 杨子祥, 等.云南松与华山松人工混交林针叶光合速率对光及CO2浓度的响应特征[J].林业科学研究, 2009, 22(1):21-25. doi: 10.3321/j.issn:1001-1498.2009.01.004 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4257
- HTML全文浏览量: 1580
- PDF下载量: 317
- 被引次数: 0
