



-
萌生现象是高等植物常见的更新方式之一[1-2],也是高等植物高度进化的不稳定特征[3],尤其是在木本被子植物中更普遍[4],是植物重要的生活史特征[5]及独特的功能特征[6]。当植物受到干扰时,萌生更新可成为植物适应各种干扰和胁迫时所采取的一种适应方式[7];同时,萌生更新还是植物更新生态位的重要组成部分,它和实生更新一样是一条植物繁衍后代的途径[8]。现今,植物种子更新的研究相比较萌生更新更深入,但植物的营养生殖[9]和萌生现象产生的原因还有较大的探讨空间[5]。
萌生现象在植物生活史过程中起很重要的作用[5],如萌生更新比实生更新可以为幼苗提供更多的物质基础[10],从而提高幼苗的存活率和生长速度[11];同时,萌生植物树皮的厚度与胸径的比值明显高于实生更新的植物,可以帮助植物抵抗外界干扰[7]。萌生现象为植物提供了侧枝和更大的冠幅来接收阳光占领生态位、增加生物量的累积[12]。另外,萌生现象还可以帮助植物在干扰后更快占据和恢复其所在的生态位,降低生物量损失[13],加快群落演替的过程[14]。
目前,国外关于萌生现象的研究主要集中于萌生在植物群落结构更新中的作用[15]、萌生的生物学特征[16]、个体生活史策略及影响因素[17]和萌生在种群、群落及景观水平上的“驻留生态位”效应等。国内这方面的研究相对较少,仅有对个别样地萌生状态的初步调查[18]、不同垂直结构层次[19]以及对个别树种萌生能力的单独研究[20]-[21],而且这些研究主要集中于物种层面萌生能力的探讨[22],或在干扰[23]后特定植物萌生现象产生的规律上[24]。此外,还有研究从植物自身的基因性状出发或从形态学[25]的角度出发,对植物的萌生现象进行了一定的探讨[26]。萌生现象作为植物生活史的重要组成部分,对于植物产生萌生现象的原因、所在环境中环境因子的影响以及植物个体和物种之间的相互影响,都是未来植物萌生研究的发展方向。
尖峰岭是我国保存最完整的热带原始森林林区之一[27]。2012年,中国林业科学研究院热带林业研究所在海南尖峰岭热带山地雨林典型分布区五分区中建立了一块面积60 hm2的森林动态监测样地,是美国热带林业研究中心CTFS全球森林动态监测样地中,迄今为止已经建立好的、单个面积最大的、单次监测植株数量最多的森林动态监测样地[28]。本文以海南尖峰岭60 hm2大样地内萌生植物为研究对象,分析样地内萌生植物的萌生物种数量特征、空间分布及其影响因素,旨在初步探讨影响热带雨林植物萌生策略选择的内在和外在因素。
-
尖峰岭地区位于海南省西南部乐东黎族自治县和东方市交界处(18°20′~18°57′ N,108°41′~109°12′ E),总面积约640 km2,其林区内的热带雨林是我国现有面积较大、保存较完整的热带原始森林之一。此外,尖峰岭地区属于低纬度热带岛屿季风气候,干湿两季气候明显,雨季为5至10月,旱季为11月至翌年4月,年降水量为1 300~3 700 mm,年均温度为19.8~24.5℃。尖峰岭地区还是我国生物多样性较高的地区之一,林区野生维管植物和习见的栽培植物共2 849种(野生植物2 287种,包括变种、亚种及变形),隶属244科1 250属[27]。该地区面积最广的植被类型为热带山地雨林,其次为热带常绿季雨林,保存有完整的原始生态系统,动植物种类丰富[28]。
-
尖峰岭热带山地雨林60 hm2大样地按照美国史密斯热带研究所热带森林研究中心(Center for Tropical Forest Science, CTFS)的调查技术规范建设,样地东西长1 000 m,南北宽600 m,将整个样地划分为1 500块20 m×20 m样地。大样地海拔范围为866.3~1 016.7 m,样地西南角较平缓,整体地势从西南角向东北角的山脊逐渐过渡抬升。对样地内所有胸径≥1.0 cm的乔灌木逐一挂标签,记录该植物的坐标、名称、胸径、树高和生存状态。首次样地植被调查于2010年年底开始,2012年年底完成。调查结果表明,样地内共记录到439 676株胸径≥1.0 cm的存活乔灌木植株,其中,除61个植株未确定种名外,其他植株分属于62科155属290种[28]。
-
本论文将植物的萌生现象按照萌生发生部位的不同分为:萌条和分支[17]。萌条:植物发生于其主干根部的萌生现象称为萌条,如根萌。分支:植物发生于其主干根部30 cm以上的萌生现象称之为分支[29]。
-
采用萌生数和萌生率2个概念。每棵具有萌生现象的植株上成活的萌生个体数之和称之为萌生数。萌生数和萌生率是衡量物种层面萌生能力的重要指标,萌生数和萌生率高的物种代表发生萌生现象的植株数更多[30-32]。
萌生数=某物种具有萌生的植株个体数
萌生率=某物种具有萌生的植株个体数/该物种在样地总个体数×100%
分支率=某物种具有分支的植株个体数/该物种在样地总个体数×100%
萌条率=某物种具有萌条的植株个体数/该物种在样地总个体数×100%
-
在计算物种重要值时,由于本研究不把植物的萌生现象算做独立植株,因此,主干的相对多度和相对频度与加入萌生现象后的相对多度和相对频度无异;萌生现象的加入直接影响了个体的胸高断面积,因此,这里在讨论萌生现象对物种重要值的计算时只考虑相对显著度的变化。
主干重要值=(主干的相对多度+主干的相对频度+主干的相对显著度)/3×100%
加萌生现象后的重要值=(主干的相对多度+主干的相对频度+主干及萌生现象的相对显著度)/3×100%
-
本研究采用逐步回归的方法,将每个样方的分支率和萌条率同该样方所具有的环境因子之间进行逐步回归,环境因子包括海拔、土壤含水量、土壤密度、土壤pH值。通过逐步回归寻找本次参与逐步回归的环境因子哪些对于样方分支率和萌条率的贡献值最高。
-
尖峰岭大样地海拔最低点(866.3 m)位于样地西南区域,海拔最高点(1 016.7 m)位于东北区域,自西南角向东北角海拔逐渐抬升,海拔跨度150.4 m。根据每个20 m×20 m小样方中心点的海拔高度绘制大样地等高线图。在绘制完成的等高线地形图上,添加各具萌生现象植株的散点分布图。
所有统计分析及作图均在Excel 2010,R 3.1.2 [33]中进行。
-
样地2012年建成时共发现290个物种,隶属62个科155个属。样地物种总个体数为439 676株,独立植株391 686株,29 103个萌条,18 887个分支[28]。其中,具有萌生现象的物种共260种,隶属58个科142个属,分别占样地植物科、属、种的93.5%、91.6%、89.7%,发生萌生的个体总数为33 919株,占样地总个体数的7.7%。样地中具有分支现象的物种共237种,隶属55个科132个属,科、属、种分别占比88.7%、85.2%、81.7%;样地中具有萌条现象的物种共248种,隶属56个科135个属,科、属、种分别占比90.3%、87.1%、85.5%。样地不具有萌生现象的物种有30种,隶属27科25属,占样地物种总数的10.3%。整个样地中,樟科、茜草科、新木姜子属、厚壳桂属、九节属的物种萌生比例高。
图 1表明:在尖峰岭大样地内,每个具有萌生现象的植株所产生的萌生数多集中在2个及以下,单个植株萌生数超过5个的不足200株。从图 2可看出:样地每个20 m×20 m样方的萌生数峰值在20~40个之间呈现单峰分布,即每个样方的萌生数会趋于一个稳定区间;此外,样方数和每个样方的萌生数在整体上呈现反“J”型分布。
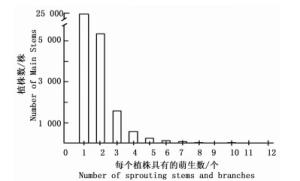
图 1 海南尖峰岭60 hm2大样地不同萌生数的植株数量统计
Figure 1. Number of main stems with different number of sprouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
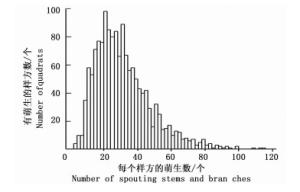
图 2 海南尖峰岭60 hm2大样地每个20 m×20 m样方的萌生数
Figure 2. Number of quadrats with different number of spouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
本文还选取相对频度大于1%的植株个体,对重要值、萌生数、萌生总植株数等因素进行统计,结果(表 1)表明:从重要值来看,白颜树最高为3.01,其次是厚壳桂和油丹,分别为2.62、2.23;从植株相对胸高断面积看,白颜树最大为5.50,其次是油丹和厚壳桂,分别为4.35、2.80;从相对密度看,四蕊三角瓣花最高为4.93,其次是厚壳桂和香果新木姜,分别为3.82、3.58。
表 1 海南尖峰岭60 hm2森林动态监测样地中萌生植物重要值及植株数量
Table 1. Theimportance values and numerical characteristics of sprouting stems in the 60 hm2 plot in Jianfengling, Hainan Island
物种名
Species科
Family重要值
Importance
value/%相对胸高断面积
Relative basal
area/%相对密度
Relative
density/%相对频度
Relative
frequency/%萌生数/个
Number of sprouting
stems and branches总植株数/株
Number of
total stems四蕊三角瓣花Prismatomeri stetrandra Rubiaceae 2.14 0.25 4.93 1.24 643 21 688 厚壳桂Cryptocarya chinensis Lauraceae 2.62 2.80 3.82 1.22 4 573 16 811 香果新木姜Neolitsea ellipsoidea Lauraceae 1.92 1.00 3.58 1.19 1 185 15 747 九节Psychotria rubra Rubiaceae 1.77 0.66 3.44 1.22 3 100 15 124 变色山槟榔Pinanga discolor Arecaceae(Palmae) 1.41 0.11 3.36 0.75 144 14 753 海南韶子Nephelium topengii Sapindaceae 2.10 2.35 2.70 1.23 651 11 878 柏拉木Blastus cochinchinensis Melastomataceae 1.04 0.08 2.65 0.38 1 498 11 692 钮子果Ardisia virens Myrsinaceae 1.25 0.12 2.46 1.16 1 732 10 817 东方琼楠Beilschmiedia tungfangensis Lauraceae 1.83 1.98 2.42 1.10 981 10 647 白颜树Gironniera subaequalis Ulmaceae 3.01 5.50 2.28 1.24 269 10 035 罗伞树Ardisia quinquegona Myrsinaceae 1.05 0.13 1.90 1.13 1 768 8 376 卵叶新木姜Neolitsea ovatifolia Lauraceae 0.76 0.15 1.70 0.44 3 116 7 464 黄叶树Xanthophyllum hainanense Polygalaceae 1.70 2.47 1.51 1.13 112 6 642 海南山矾Symplocos hainanensis Symplocaceae 0.77 0.07 1.49 0.75 1 754 6 556 东方肖榄Platea parvifolia Icacinaceae 1.27 1.21 1.46 1.14 249 6 423 钟萼粗叶木Lasianthus trichophlebus Rubiaceae 0.89 0.20 1.41 1.05 2 198 6 183 香楠Aidia canthioides Rubiaceae 0.82 0.16 1.29 1.00 1 796 5 680 喙果皂帽花Dasymaschalon rostratum Annonaceae 0.81 0.07 1.29 1.06 1 122 5 671 腺叶山矾Symplocos adenophylla Symplocaceae 0.88 0.47 1.24 0.94 1 522 5 445 谷木Memecylon ligustrifolium Melastomataceae 0.80 0.07 1.23 1.09 66 5 406 红磷蒲桃Syzyginm hancei Myrsinaceae 0.88 0.47 1.21 0.97 146 5 323 油丹Alseodaphne hainanensis Lauraceae 2.23 4.35 1.21 1.14 201 5 302 红算盘子Glochidion coccineum Euphorbiaceae 0.88 0.46 1.19 0.99 139 5 213 卵叶桂Cinnamomum rigidissimum Lauraceae 1.17 1.30 1.18 1.03 320 5 194 硬壳桂Cryptocarya chingii Lauraceae 0.86 0.60 1.16 0.83 775 5 106 软荚红豆Ormosia semicastrata Fabaece 0.85 0.64 1.11 0.79 84 4 891 阴香Cinnamomum burmanni Lauraceae 0.86 0.57 1.09 0.91 1 046 4 783 粗毛野桐Mallotus hookerianus Euphorbiaceae 0.61 0.36 1.05 0.43 286 4 638 海岛冬青Ilex goshiensis Aquifoliaceae 1.14 1.44 1.03 0.95 105 4 550 多香木Polyosma cambodiana Saxifragaceae 0.98 0.89 1.02 1.04 112 4 473 -
根据样地萌生物种萌生植株数的多少,由样地中萌生植株数量排前五的物种(表 2)表明:大样地中萌生个体数最多的物种依次是九节(2 423)、厚壳桂(2 317)、卵叶新木姜(1 724)、钟萼粗叶木(1 556)、钮子果(1 477);物种萌生率依次是钟萼粗叶木(25.17%)、卵叶新木姜(23.10%)、九节(16.02%)、厚壳桂(13.78%)、钮子果(13.65%)。
表 2 海南尖峰岭60 hm2森林动态监测样地中萌生能力排前五的物种(按萌生植株数排序)
Table 2. Topfive species with strong sprouting abilities in the 60 hm2 plot in Jianfengling, Hainan Island
种名
Species植株总数/株
Number of
total stems萌生植株数/株
Number of
sprouting stems
and branches萌生率/%
Proportion of sprouting
stems and branches of
total stems萌条数/个
Number of
sprouting
stems萌条率/%
Ratio of
spouting stems
of total stems分支数/个
Number of
branches分支率/%
Proportion of
branches of
total stems厚壳桂Cryptocarya chinensis 16 811 2 317 13.78 346 2.06 4 227 25.14 九节Psychotria rubra 15 124 2 423 16.02 2 869 18.97 231 1.53 钮子果Ardivire virens 10 817 1 477 13.65 820 7.58 912 8.43 卵叶新木姜Neolitsea ovatifolia 7 464 1 724 23.10 134 1.80 2 982 39.95 钟萼粗叶木Lasianthus trichophlebus 6 183 1 556 25.17 1 882 30.44 318 5.14 -
本文选取尖峰岭大样地中,物种加入萌生现象后重要值变化量排前五的物种,来比较各自主干重要值与加入萌生现象后的重要值变化情况,并在后文讨论萌生现象对物种重要值的影响程度。
从表 3可看出:尖峰岭60 hm2大样地中,厚壳桂的主干重要值为2.48%,加入萌生现象后重要值上升到2.62%,在样地具有萌生现象的物种当中重要值的增幅最大达0.14%。此外,除厚壳桂、尖峰岭锥的其他物种,加入萌生现象后该物种的重要值变化都没有超过0.05%。
表 3 海南尖峰岭60 hm2森林动态监测样地中加入萌生现象前后物种重要值的比较
Table 3. Comparison of importance values with or without the addition of spouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
种名
Species植株主干的重要值/% Importance values without spouting stems and branches 植株主干加入萌生现象后的重要值/% Importance values with all stems 厚壳桂
Cryptocarya chinensis2.48 2.62 尖峰岭锥
Castanopsis jianfenglingensis0.52 0.58 米锥Castanopsis chinensis 0.96 0.99 多花五月茶
Antidesma maclurei0.78 0.79 九节Psychotria rubra 1.76 1.77 -
图 3表明:植物的分支现象(绿点)在山脊地区密度更高,在沟谷地带则分布较少。进一步进行多元回归分析表明:样地分支率同地形因子中的海拔因子相关性最高(P < 0.001,r=0.087 7)。同理,将样地萌条率与4个地形因子进行回归发现,样地萌条率同样与地形因子中的海拔因子相关性最高(P < 0.001,r=0.22)。
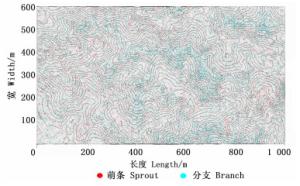
图 3 海南尖峰岭60 hm2大样地萌生现象(萌条、分支)的空间分布
Figure 3. The spatial distribution of sprouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
-
通过对样地萌生现象数据的汇总与分析发现:尖峰岭60 hm2大样地内,具有萌生现象的科、属、种、占样地植株总数的比值均超过80%,这意味着:海南尖峰岭超过8成的物种会产生萌生现象;但从整体样地看,个体萌生现象发生率只有7.7%,这可能与植株所在生境有关,导致个体层面萌生率低下但物种层面萌生率高的现象。
古田山茶湾1 hm2样地内,具有根萌现象(萌条现象)的物种共计68种,占群落总物种数的67.3%,隶属于48个属26个科,分别占群落总科数和总属数的72.2%和70.6%[5]。与古田山样地相比,尖峰岭大样地具有萌条现象物种所占科、属、种的比例分别为90.3%、87.1%、85.5%,均高于古田山样地。
古田山茶湾1 hm2样地内,物种层面萌生现象占比为63.95%[34];天目山自然保护区样地中,物种层面萌生现象占比为55%[35];尖峰岭大样地物种层面萌生现象占比为89.7%。可见,尖峰岭大样地植株在物种层面的萌生比例比古田山茶湾1 hm2样地和天目山自然保护区样地中的高。出现这种现象的原因可能是因为:(1)尖峰岭地区受到的台风干扰较大,造成植株在科、属、种层面上出现较高的萌生率。(2)由于尖峰岭60 hm2大样地为原始丛林,具有较为成熟的生态位分化和相对稳定的生境,导致整个样地层面上植株个体萌生率仅为7.7%的现象。
-
对尖峰岭60 hm2大样地中具有萌生现象的植株进行统计后,将样地中具有萌生现象且萌生植株数排前五的物种分别计算每个物种在样地中的萌生率、分支率、萌条率,对比每个物种在萌生策略选择上的不同。从表 2可看出,萌生数的排序和萌生率的排序略有不同,说明这2个指标虽然都用来表达物种的萌生能力大小,但2个数据之间可能不具有相关性,这与冯辉[36]在研究重庆缙云山长绿阔叶林和刘常幸[6]在研究古田山常绿阔叶林树种的萌生特征中得出的萌生率和萌枝数不存在相关性的结论一致。
在萌生策略选择方面:九节和钟萼粗叶木偏向选择产生萌条;厚壳桂、卵叶新木姜则偏向选择产生分支,其中,物种萌生率最高的是钟萼粗叶木(25.17%)和卵叶新木姜(23.10%),虽然具有相似的植物萌生率,但萌生方式却不同。由此可见,虽同为具有较强萌生能力的树种,但在选择通过不同萌生方式占据生态位时仍然有很大差异。产生这种差异的原因可能有2方面:(1)植物本身基因决定的功能性状,导致植物在萌生策略选择上出现了不同。(2)植物所在生境条件的后天影响,如海拔、光照、物种间的相互作用等因素,导致了植物需要通过选择不同的萌生方式提高自身在当前环境中的生存优势。
在样地萌生能力前五的物种中,只有钮子果的萌条率和分支率接近,分别为:7.58%和8.43%,该物种的萌生能力在样地萌生能力前五的物种中排第五,萌生能力不及偏向产生萌条的九节和钟萼粗叶木,也不及偏向产生分支的厚壳桂和卵叶新木姜。因此,是否偏向选择一种萌生策略更有利于提高物种在样地中的萌生能力,从而占据更多的生态位以便物种在群落中更好的发展?后期应进行相应的统计分析并结合CTFS网络下其他样地数据证明此结论。
-
生境异质性假说认为,植株的萌生情况与生长环境有关[37-38]。因此,通过观察植物萌生现象在空间上的分布,可以初步判断植物萌生现象同植物空间分布之间的关系[7]。
图 1可以清楚的看到植株的萌生现象和植株所在的地理位置的地形因子具有一定相关性,其中,分支现象在山脊地区分布密度明显更高,在山谷地区分布较少。海拔作为热带山地雨林重要的环境影响因子,直接影响植物所在环境的土壤水分[39],从而影响植物的萌生现象和物种多样性,这与黄甫昭等[40]在研究萌生及生物多样性随海拔的变化相吻合,即弄岗样地相对海拔76.3 m以上多样性随相对海拔增高而增大。出现这种现象的原因还可能是尖峰岭地处海南岛西南部,是台风活动的频发区,因此,在山脊地区的植株受到来自台风的外界干扰更大[41],从而迫使植物产生更多的萌生现象以占领生态位。
-
在森林群落中,萌生现象给物种创造了“更新生态位”的条件,是萌生植物占据生存空间、占领生态位并保持竞争优势的机会[5]。根据本文对样地物种重要值的计算,对比加入萌生现象后物种重要值的变化情况,重要值变化量最大的是厚壳桂(+0.14),其次是尖峰岭锥(+0.04),其他物种重要值的变化均小于0.04,因此,本研究认为在尖峰岭大样地植物萌生现象对样地物种重要值的影响不大。
这个结论同刘常幸等[5]在研究古田山常绿阔叶林萌生情况所得出的结论有所不同,在古田山自然无干扰的常绿阔叶林中,个别萌生能力很强的物种如灰白蜡瓣花、石栎等,萌生现象(仅萌条)对其重要值的影响较大,加入萌生现象后重要值的变化量分别为灰白蜡瓣花5.54%、石栎0.49%。与尖峰岭样地所得出不同结论的原因可能在于,古田山地处亚热带,与尖峰岭地区受台风干扰相比,古田山地区受到的干扰较小,植株产生的萌生现象有更大的机会成长并累积生物量,从而在样地生物量计算时,由于萌生现象的胸径过大而对生物量产生较大的影响。反之,在尖峰岭大样地中,因为台风的干扰导致了植物的萌生现象无法长期存活,所以,萌生现象无法为植物个体增加生物量的累积。虽然在长期和频繁的台风干扰下植物会产生大量的萌生现象来占据生态位[42],但相对的植物产生萌生现象的胸径都偏小,在计算重要值时对其产生的影响较小。
目前,国内有关植物萌生现象对物种重要值影响的贡献和处理方法并不统一,如姚良锦等[43]在讨论湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构中,在重要值计算时去除了植物的萌生现象;刘常幸等[5]在分析古田山茶湾样地甜槠—木荷林根萌(萌条)特征时,分开讨论了重要值与萌生现象的2种情况。何兴东等[44]早期在群落层面提出简化重要值和样地指数等概念,通过盖度乘以高度避免乔木萌生现象的统计。在植物萌生现象对物种重要值的贡献量方面后期可以做进一的步探讨。
-
本研究以尖峰岭地区2012年第1次样地调查的数据为基础,分析发现:样地植物在科、属、种水平上的萌生率均超过80%,整体萌生现象发生率只有7.7%;樟科、茜草科、新木姜子属、厚壳桂属、九节属的物种萌生比例高于其他科、属;萌生率较高的物种更偏向选择一种萌生方式(分支/萌条)占领生态位;单个植株更倾向产生的萌生数是2个,单个样方产生的萌生数在20个左右呈现单峰分布,样方数和每个样方的萌生数在整体上呈现反“J”型分布;萌生现象同海拔具有高相关性,分支现象多集中在山脊地区;除厚壳桂和尖峰岭锥外,物种萌生现象对该物种的重要值影响不超过0.05%。
综上所述,海南尖峰岭60 hm2大样地萌生现象的调查明确了样地萌生物种组成、物种萌生能力排序、萌生个体空间分布以及萌生现象对于物种重要值的影响,提出了物种萌生策略选择的猜想以供后期进一步探讨,为下一步深入研究和探讨尖峰岭大样地萌生现象及其影响因子的时空动态变化打下基础。
海南尖峰岭60 hm2大样地萌生植物数量特征
Numerical Characteristics of Sprouting Plants in the 60 hm2 plot, Jianfengling, Hainan Island
-
摘要:
目的 分析海南典型热带山地雨林分布区域尖峰岭60 hm2大样地内萌生植物的物种组成、数量和空间分布特征,探讨其影响因素。 方法 以海南尖峰岭热带山地雨林60 hm2大样地及样地内具有萌生现象的木本植物(胸径≥1 cm)为研究对象,统计样地萌生植物的萌生数和萌生率,并描述其空间分布特征。本文将植物的萌生现象分为两类:萌条和分支。 结果 表明:样地具有萌生现象的木本植物共有260种,隶属58个科142个属,科、属、种数量分别占样地植物总数的93.5%、91.6%与89.7%;萌生个体数最多的前5个物种依次是:九节、厚壳桂、卵叶新木姜、钟萼粗叶木、钮子果;除厚壳桂、尖峰岭锥外的其他植物,萌生现象对物种重要值的影响均不超过0.05%。植物的萌生现象与海拔极显著相关(P < 0.001)。 结论 作为尖峰岭热带山地雨林占优势的更新模式之一,萌生更新不仅是树种对生态位占据的重要方式,同时也对群落物种共存、生物多样性的维持起到重要作用。后期还可以对样地萌生现象受台风干扰后的表现进行分析。 Abstract:Objective To explore the sprouting phenomenon in the tropical mountain rainforest by investigating species composition, spatial distribution, and factors related to the sprouting characteristics. Method Taking the woody plants (DBH ≥ 1cm) with sprouting phenomenon in a montane tropical rainforest plot with an area of 60 hectares plot in Jianfengling of Hainan Province as the objectives of study, the quantitative characteristics of sprouting plants were analyzed and their spatial distribution characteristics were outlined. In this study, the sprouting plants were subdivided into two types:branches and sprouts. Result There are 260 species of woody plants with sprouting phenomenon, belonging to 58 families and 142 genera. The amount of families, genus and species of sprouting plants accounted for 93.5%, 91.6% and 89.7% of the total plants respectively.There are five species with the most sprouting stems, including Psychotria asiatica, Cryptocarya chinensis, Neolitsea ovatifolia, Lasianthus trichophlebus and Ardisia virens. Except Cryptocarya chinensis and Castanopsis jianfenglingensis, the effects of sprouting phenomenon on the changes of importance values were less than 0.05%. The sprouting phenomenon of plants was also significantly correlated with altitude with P-value < 0.001. Conclusion As the dominant regeneration type in the montane tropical rainforest in the plot, the sprouting stem contributes to the maintenance of biodiversity in the forest community and plays a major role for the species coexistence. It is suggested that the responses of the sprouting phenomenon after the typhoon disturbance should be further studied. -
Key words:
- montane tropical rainforest
- / sprouting plant
- / spatial distribution
- / importance value
- / Jianfengling
-
图 1 海南尖峰岭60 hm2大样地不同萌生数的植株数量统计
Figure 1. Number of main stems with different number of sprouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
图 2 海南尖峰岭60 hm2大样地每个20 m×20 m样方的萌生数
Figure 2. Number of quadrats with different number of spouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
图 3 海南尖峰岭60 hm2大样地萌生现象(萌条、分支)的空间分布
Figure 3. The spatial distribution of sprouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
表 1 海南尖峰岭60 hm2森林动态监测样地中萌生植物重要值及植株数量
Table 1. Theimportance values and numerical characteristics of sprouting stems in the 60 hm2 plot in Jianfengling, Hainan Island
物种名
Species科
Family重要值
Importance
value/%相对胸高断面积
Relative basal
area/%相对密度
Relative
density/%相对频度
Relative
frequency/%萌生数/个
Number of sprouting
stems and branches总植株数/株
Number of
total stems四蕊三角瓣花Prismatomeri stetrandra Rubiaceae 2.14 0.25 4.93 1.24 643 21 688 厚壳桂Cryptocarya chinensis Lauraceae 2.62 2.80 3.82 1.22 4 573 16 811 香果新木姜Neolitsea ellipsoidea Lauraceae 1.92 1.00 3.58 1.19 1 185 15 747 九节Psychotria rubra Rubiaceae 1.77 0.66 3.44 1.22 3 100 15 124 变色山槟榔Pinanga discolor Arecaceae(Palmae) 1.41 0.11 3.36 0.75 144 14 753 海南韶子Nephelium topengii Sapindaceae 2.10 2.35 2.70 1.23 651 11 878 柏拉木Blastus cochinchinensis Melastomataceae 1.04 0.08 2.65 0.38 1 498 11 692 钮子果Ardisia virens Myrsinaceae 1.25 0.12 2.46 1.16 1 732 10 817 东方琼楠Beilschmiedia tungfangensis Lauraceae 1.83 1.98 2.42 1.10 981 10 647 白颜树Gironniera subaequalis Ulmaceae 3.01 5.50 2.28 1.24 269 10 035 罗伞树Ardisia quinquegona Myrsinaceae 1.05 0.13 1.90 1.13 1 768 8 376 卵叶新木姜Neolitsea ovatifolia Lauraceae 0.76 0.15 1.70 0.44 3 116 7 464 黄叶树Xanthophyllum hainanense Polygalaceae 1.70 2.47 1.51 1.13 112 6 642 海南山矾Symplocos hainanensis Symplocaceae 0.77 0.07 1.49 0.75 1 754 6 556 东方肖榄Platea parvifolia Icacinaceae 1.27 1.21 1.46 1.14 249 6 423 钟萼粗叶木Lasianthus trichophlebus Rubiaceae 0.89 0.20 1.41 1.05 2 198 6 183 香楠Aidia canthioides Rubiaceae 0.82 0.16 1.29 1.00 1 796 5 680 喙果皂帽花Dasymaschalon rostratum Annonaceae 0.81 0.07 1.29 1.06 1 122 5 671 腺叶山矾Symplocos adenophylla Symplocaceae 0.88 0.47 1.24 0.94 1 522 5 445 谷木Memecylon ligustrifolium Melastomataceae 0.80 0.07 1.23 1.09 66 5 406 红磷蒲桃Syzyginm hancei Myrsinaceae 0.88 0.47 1.21 0.97 146 5 323 油丹Alseodaphne hainanensis Lauraceae 2.23 4.35 1.21 1.14 201 5 302 红算盘子Glochidion coccineum Euphorbiaceae 0.88 0.46 1.19 0.99 139 5 213 卵叶桂Cinnamomum rigidissimum Lauraceae 1.17 1.30 1.18 1.03 320 5 194 硬壳桂Cryptocarya chingii Lauraceae 0.86 0.60 1.16 0.83 775 5 106 软荚红豆Ormosia semicastrata Fabaece 0.85 0.64 1.11 0.79 84 4 891 阴香Cinnamomum burmanni Lauraceae 0.86 0.57 1.09 0.91 1 046 4 783 粗毛野桐Mallotus hookerianus Euphorbiaceae 0.61 0.36 1.05 0.43 286 4 638 海岛冬青Ilex goshiensis Aquifoliaceae 1.14 1.44 1.03 0.95 105 4 550 多香木Polyosma cambodiana Saxifragaceae 0.98 0.89 1.02 1.04 112 4 473  下载: 导出CSV
下载: 导出CSV
表 2 海南尖峰岭60 hm2森林动态监测样地中萌生能力排前五的物种(按萌生植株数排序)
Table 2. Topfive species with strong sprouting abilities in the 60 hm2 plot in Jianfengling, Hainan Island
种名
Species植株总数/株
Number of
total stems萌生植株数/株
Number of
sprouting stems
and branches萌生率/%
Proportion of sprouting
stems and branches of
total stems萌条数/个
Number of
sprouting
stems萌条率/%
Ratio of
spouting stems
of total stems分支数/个
Number of
branches分支率/%
Proportion of
branches of
total stems厚壳桂Cryptocarya chinensis 16 811 2 317 13.78 346 2.06 4 227 25.14 九节Psychotria rubra 15 124 2 423 16.02 2 869 18.97 231 1.53 钮子果Ardivire virens 10 817 1 477 13.65 820 7.58 912 8.43 卵叶新木姜Neolitsea ovatifolia 7 464 1 724 23.10 134 1.80 2 982 39.95 钟萼粗叶木Lasianthus trichophlebus 6 183 1 556 25.17 1 882 30.44 318 5.14
下载: 导出CSV
表 3 海南尖峰岭60 hm2森林动态监测样地中加入萌生现象前后物种重要值的比较
Table 3. Comparison of importance values with or without the addition of spouting stems and branches in the 60 hm2 plot in Jianfengling, Hainan Island
种名
Species植株主干的重要值/% Importance values without spouting stems and branches 植株主干加入萌生现象后的重要值/% Importance values with all stems 厚壳桂
Cryptocarya chinensis2.48 2.62 尖峰岭锥
Castanopsis jianfenglingensis0.52 0.58 米锥Castanopsis chinensis 0.96 0.99 多花五月茶
Antidesma maclurei0.78 0.79 九节Psychotria rubra 1.76 1.77
下载: 导出CSV
-
[1] Randall C K, Duryea M L, Vince S W, et al. Factors influencing stump sprouting by pondcypress (Taxodium distichum var. nutans (Ait.) Sweet)[J]. New Forests, 2005, 29(3):245-260. doi: 10.1007/s11056-005-5652-4 [2] Moreira B, Tormo J, Pausas J G. To resprout or not to resprout:factors driving intraspecific variability in resprouting[J]. Oikos, 2012, 121(10):1577-1584. doi: 10.1111/more.2012.121.issue-10 [3] Bond W J, Midgley J J. The Evolutionary Ecology of Sprouting in Woody Plants[J]. Trends in Ecology & Evolution, 2001, 16(1):45-51. [4] 姚兰, 崔国发, 易咏梅, 等.湖北木林子保护区大样地的木本植物多样性[J].林业科学, 2016, 52(1):1-9. [5] 刘常幸, 金毅, 余建平, 等.古田山茶湾样地甜槠-木荷林根萌特征分析[J].浙江大学学报:理学版, 2014, 41(5):573-582, 592. [6] 刘常幸.古田山常绿阔叶林2007-2012年根萌特征动态分析[D].金华: 浙江师范大学, 2014. [7] 周长宁, 沈有信.半湿润常绿阔叶林植物萌生特征在喀斯特丘陵坡面上的变化[J].云南大学学报:自然科学版, 2012, 34(s1):135-140. [8] Simler A B, Metz M R, Frangioso K M, et al. Novel disturbance interactions between fire and an emerging disease impact survival and growth of resprouting trees[J]. Ecology, 2018, 99(10):2217-2229. doi: 10.1002/ecy.2493 [9] 齐春华.植物组织培养技术发展现状及方向[J].农业科技与装备, 2011(4):10-11. doi: 10.3969/j.issn.1674-1161.2011.04.004 [10] 吴培培, 孙冰, 罗水兴, 等.岭南槭不同种源家系种子形态和幼苗生长变异[J].林业科学研究, 2017, 30(06):1015-1021. [11] 卢群, 石俊慧, 曾小康, 等.深圳湾福田红树林砍伐后萌生更新的初步研究[J].广西师范大学学报:自然科学版, 2013, 31(2):107-112. [12] 曹菁, 杨同辉, 章建红, 等.浙江天童冬青属植物的种群结构及更新类型[J].浙江农林大学学报, 2015, 32(1):76-83. [13] Plotkin A B, Foster D, Carlson J, et al. Survivors, not invaders, control forest development following simulated hurricane[J]. Ecology, 2013, 94(2):414-423. doi: 10.1890/12-0487.1 [14] 陈沐, 曹敏, 林露湘.木本植物萌生更新研究进展[J].生态学杂志, 2007, 26(7):1114-1118. doi: 10.3321/j.issn:1000-4890.2007.07.026 [15] Simões C G, Marques M C M. The Role of Sprouts in the Restoration of Atlantic Rainforest in Southern Brazil[J]. Restoration Ecology, 2010, 15(1):53-59. [16] Tredici P D. Sprouting in temperate trees:A morphological and ecological review[J]. Botanical Review, 2001, 67(2):121-140. doi: 10.1007/BF02858075 [17] Bellingham P J, Sparrow A D. Resprouting as a life history strategy in woody plant communities[J]. Oikos, 2000, 89(2):409-416. doi: 10.1034/j.1600-0706.2000.890224.x [18] 何永涛, 曹敏, 唐勇, 等.云南省哀牢山中山湿性常绿阔叶林萌生现象的初步研究[J].植物科学学报, 2000, 18(6):523-527. doi: 10.3969/j.issn.2095-0837.2000.06.015 [19] 陈沐, 曹敏, 林露湘.木本植物萌生更新研究进展[J].生态学杂志, 2007, 26(7):1114-1118. doi: 10.3321/j.issn:1000-4890.2007.07.026 [20] 王希华, 严晓, 闫恩荣, 等.天童几种常绿阔叶林优势种在砍伐后萌枝更新的初步研究[J].植物科学学报, 2004, 22(1):52-57. doi: 10.3969/j.issn.2095-0837.2004.01.010 [21] 张纪林, 季永华.海岸带杨柳新无性系苗期耐盐性研究[J].南京林业大学学报:自然科学版, 2004, 28(3):45-50. [22] 张琴, 刘利文, 李俊清, 等.云南干热河谷区裸地植被恢复过程中车桑子的种群特征[J].西南农业学报, 2016, 29(9):2234-2238. [23] 赵美玲, 潘勃, 谭运洪, 等.人为干扰下西双版纳地区几种优势树种和濒危树种功能性状的比较(英文)[J].Science Bulletin, 2015, 60(10):916-924, 983. [24] Zhang Y, Chen H Y H, Taylor A R. Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees[J]. Functional Ecology, 2016, 31(2), 419-426. [25] Biagini B, Imazio S, Scienza A, et al. Renewal of wild grapevine (Vitisvinifera L. subsp. sylvestris (Gmelin) Hegi) populations through sexual pathway:Some Italian case studies[J]. Flora, 2016, 219:85-93. doi: 10.1016/j.flora.2016.01.003 [26] Eloy C. Leonardo's rule, self-similarity, and wind-induced stresses in trees[J]. Physical Review Letters, 2011, 107(25):258101. doi: 10.1103/PhysRevLett.107.258101 [27] 李意德.中国生态系统定位观测与研究数据集:森林生态系统卷, 海南尖峰岭站(生物物种数据集)[M].北京:中国农业出版社, 2012. [28] 许涵.海南尖峰岭热带山地雨林-群落特征树种及其分布格局:海南尖峰岭热带山地雨林-群落特征树种及其分布格局[M].北京:中国林业出版社, 2014. [29] Condit R. Tropical forest census plots: methods and results from Barro Colorado Island, Panama and a comparison with other plots.[M].Berlin: SpringerVerlag, 1998. [30] 许涵, 李意德, 林明献, 等.海南尖峰岭热带山地雨林60 hm2动态监测样地群落结构特征[J].生物多样性, 2015, 23(2):192-201. [31] 贺金生, 陈伟烈.神农架地区米心水青冈萌枝过程的研究[J].植物生态学报, 1998, 22(5):385-391. doi: 10.3321/j.issn:1005-264X.1998.05.001 [32] 郭屹立, 王斌, 向悟生, 等.广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析[J].生物多样性, 2015, 23(2):183-191. [33] Team R D C. R Core Team. R A Language and Environment for Statistical Computing 2014[J]. Computing, 2015, 1: 12-21. [34] 叶铎, 钱海源, 王璐瑶, 等.钱江源国家公园古田山常绿阔叶林木本植物的萌生更新特征[J].生态学报, 2018, 38(10):3562-3568. [35] 于硕, 王嵘, 刘敏, 等.天目山米心水青冈群落物种组成特点与萌条更新[J].生态学杂志, 2009, 28(2):182-187. [36] 冯辉.重庆缙云山常绿阔叶林常见树种萌生特征研究[D].重庆: 西南大学, 2012. [37] 朱万泽, 王金锡, 罗成荣, 等.森林萌生更新研究进展[J].林业科学, 2007, 43(9):74-82. [38] 程真, 周光益, 吴仲民, 等.南岭南坡中段不同群落林下幼树的生物多样性及分布[J].林业科学研究, 2015, 28(4):543-550. doi: 10.3969/j.issn.1001-1498.2015.04.014 [39] 时雷雷, 骆土寿, 许涵, 等.尖峰岭热带山地雨林土壤物理性质小尺度空间异质性研究[J].林业科学研究, 2012, 25(3):285-293. doi: 10.3969/j.issn.1001-1498.2012.03.004 [40] 黄甫昭, 丁涛, 李先琨, 等.弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化[J].生态学报, 2016, 36(14):4509-4517. [41] 何增丽, 许涵, 秦新生, 等.海南尖峰岭热带山地雨林2种夹竹桃科植物的空间分布格局与关联性[J].生物多样性, 2017, 25(10):1065-1074. [42] Gao W. Effects of Wind on Species Diversity & Growth of Protection Forest along Different Coastal Gradients[J]. Protection Forest Science & Technology, 2013, 3:4-8 [43] 姚良锦, 姚兰, 易咏梅, 等.湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构[J].生物多样性, 2017, 25(3):275-284. [44] 何兴东, 高玉葆, 刘惠芬.重要值的改进及其在羊草群落分类中的应用[J].植物研究, 2004, 24(4):466-472. doi: 10.3969/j.issn.1673-5102.2004.04.026 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4773
- HTML全文浏览量: 1853
- PDF下载量: 244
- 被引次数: 0
