




-
相较于大自然的广阔,蝴蝶在觅食及求偶时必须要精确定位或缩小寄主植物和异性的寻找范围,在此期间,视觉及嗅觉的利用扮演了极为重要的角色。
蝴蝶具有比人类更为宽广的色域,大部分可感知紫外线及可见光波段[1],部分蝴蝶甚至可感知近红外光的部分区域[2-3],因此蝴蝶的色彩辨别能力较强。一般来说,具有访花习性的蝴蝶在觅食时会依靠视觉信息(花冠的颜色、形状和大小等)定位蜜源植物,嗅觉信息(花香挥发物等)则起到近距离识别的作用[4-13],但每种蝴蝶对颜色和气味的趋性有较大的区别[14],视觉及嗅觉的利用权重也因种而异[13, 15]。如白带锯蛱蝶(Cethosia cyane)和金斑蝶(Danaus chrysippus),觅食以视觉为主,在访花过程中对红色、橙色及黄色的花冠偏好较强,对嗅觉的依赖较小[15];红锯蛱蝶(Cethosia biblis)对白色、黄色及粉色花显示出一定的偏好,但觅食时更依赖于嗅觉,对花香挥发物有明显的趋性,最喜访水杨酸甲酯,对视觉的依赖较小[7]。也有例外,如虎斑蝶(Danaus genutia)是专性访花的蝶种,其在觅食时却仅依靠嗅觉[12]。食腐型的蝴蝶则一般嗅觉较为发达,主要依靠挥发性物质寻找食物[12],有些仅依靠嗅觉即可完成觅食,如枯叶蛱蝶(Kallima inachus)[15]。传粉昆虫的访花行为和偏好不仅受花的颜色、形状、气味等性状的影响,还会受到天敌和其它访花昆虫的影响[16],同时森林中物种繁多,有许多其他来源的化学噪音(chemical noise)[17],而蝴蝶维持嗅觉所消耗的能量是巨大的[18],为了减少能量消耗和规避被捕食的风险,它们必须节省觅食的成本,将视觉及嗅觉信号合理的利用起来。唐宇翀[12]则根据觅食时嗅觉和视觉的利用权重将蝴蝶划分为以下4种类型:(1)以视觉信号为主,嗅觉信号为辅;(2)嗅觉信号为主,视觉信号为辅;(3)视觉和嗅觉信号同等重要;(4)只依赖嗅觉信号。
蝴蝶在求偶的过程中,一般通过视觉(翅的色彩、斑纹、形状及大小等)及嗅觉(虫体挥发物等)信号来寻找和辨识配偶 [10, 13, 19-21]。通常情况下,雄蝶会主动发起求偶,并利用视觉确定雌蝶的位置[22],而雌蝶在择偶时,为提高繁殖成功率,也会选择具有特定特征的雄蝶[23]。性二型的蝴蝶多通过自然光下的翅膀色彩及斑纹来识别异性[24],如白带锯蛱蝶(C. cyane)[21]和菜粉蝶欧洲亚种(Pieris rapae rapae)[25],前者具有明显的性二色性,后者雌、雄蝶前翅背面具有数目不同的黑色斑点,因此能利用视觉识别雌雄。有些可见光下色彩相同的蝴蝶,如菜粉蝶日本亚种(Pieris rapae crucivora),其雌蝶通过后翅腹面反射紫外线来引诱雄蝶求偶,且在紫外线充足的环境中雄蝶求偶行为表现的更积极[26],达摩凤蝶(Papilio demoleus)[13]及一种灰蝶(Lycaena rubidus)[27]也可以通过紫外光下色彩辨识雌雄。蝴蝶在识别雌雄时,同样可以利用化学信号。如大帛斑蝶(Idea leuconoe),在求偶时会先利用共有挥发物识别同类,后利用雌、雄特有挥发物识别同性,但不能通过挥发物直接辨别异性[10];虎斑蝶(Danaus genutia)对异性的色彩不敏感,可能仅利用嗅觉识别雌雄[13];酢浆灰蝶(Zizeeria maha)和黑纹粉蝶(Pieris melete)的种内识别也主要依靠嗅觉[28]。
玉带凤蝶(Papilio polytes)是一种雌雄异型的蝴蝶,隶属凤蝶科(Papilionidae)凤蝶属(Papilio)美凤蝶亚属(Menelaides),是一种较为常见的蝶种,主要分布于亚洲及东欧。其幼虫寄主主要为柑橘(Citrus reticulata Blanco)、花椒(Zanthoxylum bungeanum Maxim.)等芸香科作物。其成虫寿命在一个月左右[29],羽化当天即可交配[30],以花蜜补充营养。蜜源植物主要为马缨丹(Lantana camara L.)、龙船花(Ixora chinensis Lam.)和茉莉(Jasminum sambac(L.)Ait)等,具有一定的传粉生态价值;同时玉带凤蝶外形大方美丽,姿态翩跹,可用于景观建设和制作标本,具有很高的观赏价值和工艺价值[31]。
玉带凤蝶的生态学和行为学研究较少,本研究通过玉带凤蝶访花和求偶过程中的颜色识别和选择行为观察,解析玉带凤蝶的视觉和嗅觉在觅食和求偶中的识别和选择规律,讨论玉带凤蝶的视觉和嗅觉在觅食和求偶过程中的生态学意义。
-
试验在云南省昆明市盘龙区中国林业科学研究院资源昆虫研究所进行(25°3′48″ N,102°3′48″ E),海拔1 954 m,年平均气温14.9℃,年平均降水量1000.5 mm,属于亚热带高原季风气候。用白色尼龙纱构建8 m × 8 m × 5 m网室,网室内阳光充足,通风状况良好。
-
供试蝴蝶选用未交配的玉带凤蝶(P. polytes),选取求偶行为最为活跃的3日龄健康成虫用于试验。
-
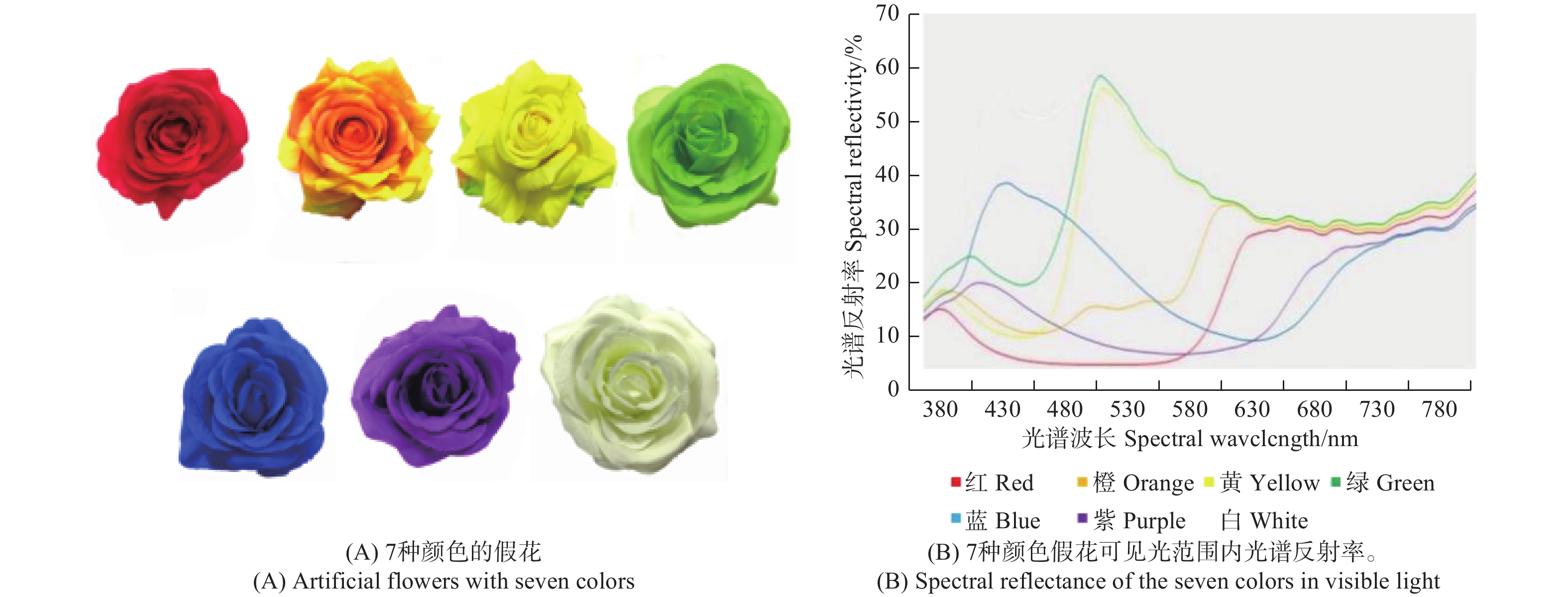
为绢布制成的假花,选用红、橙、黄、绿、蓝、紫和白色7种花,其具有基本相同的花瓣形状和大小(图1A)。花冠直径相近(8.23 ± 0.11 cm),花冠深度相近(2.63 ± 0.05 cm)。用光谱仪测量花色的反射光谱,该光谱仪针对MgO涂覆的表面作为参考进行校准,在自然光(380~780 nm)波长范围内测各色假花的光谱反射率(图1B)。
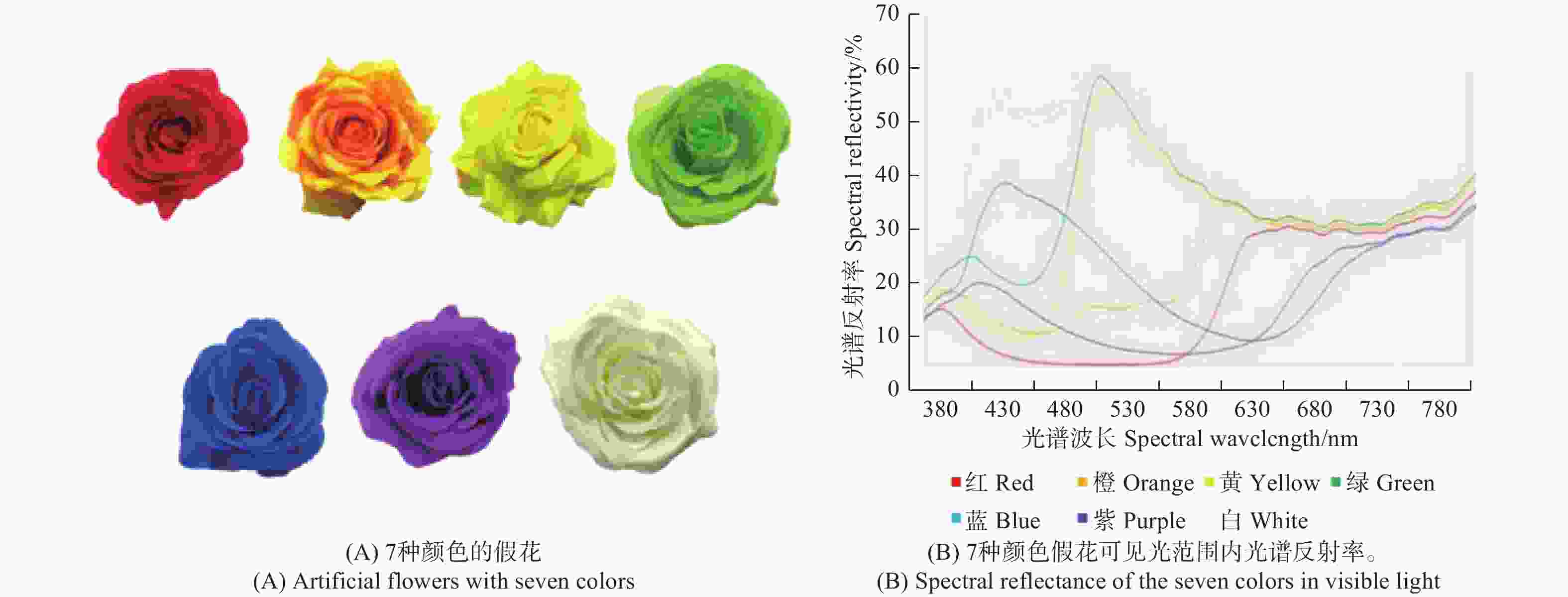
图 1 7种假花和可见光范围内光谱反射率
Figure 1. Artificial flowers with seven colors and their spectral reflectance in visible light
-
使用玉带凤蝶雌、雄蝶各5只,用PVC塑料塑封,制作成隔绝雌、雄蝴蝶的塑封模型(图2),用于蝴蝶求偶模型试验。

图 2 玉带凤蝶塑封模型
Figure 2. An artificial P. polytes model
-
试验在网室中进行,选择红、橙、黄、绿、蓝、紫和白色7种假花,将5朵相同颜色的花组成一束,插入透明的塑料瓶中固定,将瓶子放置在80 cm的高度,每种颜色的花相距200 cm,间隔30 min移动轮换各色花的位置。在天气晴朗、光照充足,温度15~25℃,相对湿度70%~80%的环境条件下,放入雌、雄玉带凤蝶各20只,于上午8:00开始不间断的人工观察,至19:00点结束,观察3日,记录它们对不同颜色花朵的访问次数(下同)。
-
以洋槐花蜂蜜为原料,用水稀释配制为10%浓度的蜂蜜水,在红、橙、黄、绿、蓝、紫、白色7种假花束上各喷洒10 mL的蜂蜜水,间隔30 min喷一次蜜水。观察并记录上述20对玉带凤蝶对不同颜色花朵的访问次数。
-
用10种常见的花香挥发物[32]对玉带凤蝶做触角电位(EAG)分析(使用PRG-2触角电位装置,将蝴蝶触角完整剪下,触角顶端切下约0.5 mm,使用导电液将触角黏合在电极上,参考电极连在触角基部,记录电极连在触角端部)。选择玉带凤蝶触角电位EAG值较高的花香挥发物,使用正己烷(分析纯,并作为对照)做溶剂[13],配成10%的挥发物溶液。将假花花冠去掉,剩下茎秆,在茎秆上喷撒配好的挥发物溶液,放入20对玉带凤蝶,观察并记录玉带凤蝶对花柄的访问次数。
选取玉带凤蝶访花最多的红色花,将5朵扎成一束并在花冠上喷洒配好的花香挥发物溶液,观察并记录玉带凤蝶对花冠+挥发物溶液的访问次数。
-
自然种群:在网室内放置进入求偶期的雌、雄玉带凤蝶各20只,观察并记录自然种群雌、雄蝶的求偶追逐行为。
蝴蝶模型:在求偶期,在网室内垂直悬挂雌、雄蝶塑封模型各5只及1只空模型,模型距地垂直高度约1.8 m。分别放入雌、雄玉带凤蝶各20只,每1小时交换一次模型位置,观察并记录雌、雄蝶追逐模型的行为次数。
2个试验用10%浓度的洋槐花蜂蜜水为玉带凤蝶提供营养。
-
取样方法:在无挥发性化学试剂污染的室内,将300 mL干净无污染的锥形瓶用塑料膜封口,将PDMS/DVB固相微萃取头(65 μm,Supelco,Bellefonte,PA,USA)扎穿塑料并固定,吸附时间为40 min,作为空白对照。取5只羽化3天左右且未交配的玉带凤蝶雄蝶,置入另一干净无污染的300 mL锥形瓶,用上述塑料封口,用老化后的萃取头吸附40 min,雌蝶同上。
样品分析仪器:结果分析使用气质联用仪(GC-MS)(Thermo Fisher TRACE GC ULTRA和Thermo Fisher ITQ 900 MS),每次进样前需在GC进样口老化萃取头(250℃, 10 min)。使用TR-5MS色谱柱(内径:0.25 mm,厚度:0.25 μm,长度:30 m)。
程序设定:40℃保持2 min,升温至120℃保持2 min (4℃·min−1),再升温至230℃保持5 min (5℃·min−1),进样口温度为 250℃,载气He的气压为69 kPa[13]。
化合物定性方法:以美国国家标准技术研究所(National Institute of Standards and Technology,NIST,http://webbook.nist.gov/chemistry/)谱库检索为主,人工解析质谱图。
-
将雌、雄玉带凤蝶标本及模型按背、腹面顺序平展开,用光谱仪(USB200+,Ocean Optics,Inc,USA)测量自然光波长范围内的反射光谱,利用Excel绘制色谱图。用紫外仪(北京六一紫外仪,型号WD-9403C,反射波长365 nm)拍摄标本和蝴蝶模型紫外图像。
-
使用SPSS25.0统计分析软件,对玉带凤蝶访问不同颜色假花和自然种群求偶的数据分别采用卡方检验;对玉带凤蝶访问喷洒蜂蜜水及花香挥发物前后的数据分别采用独立样本t检验。
-
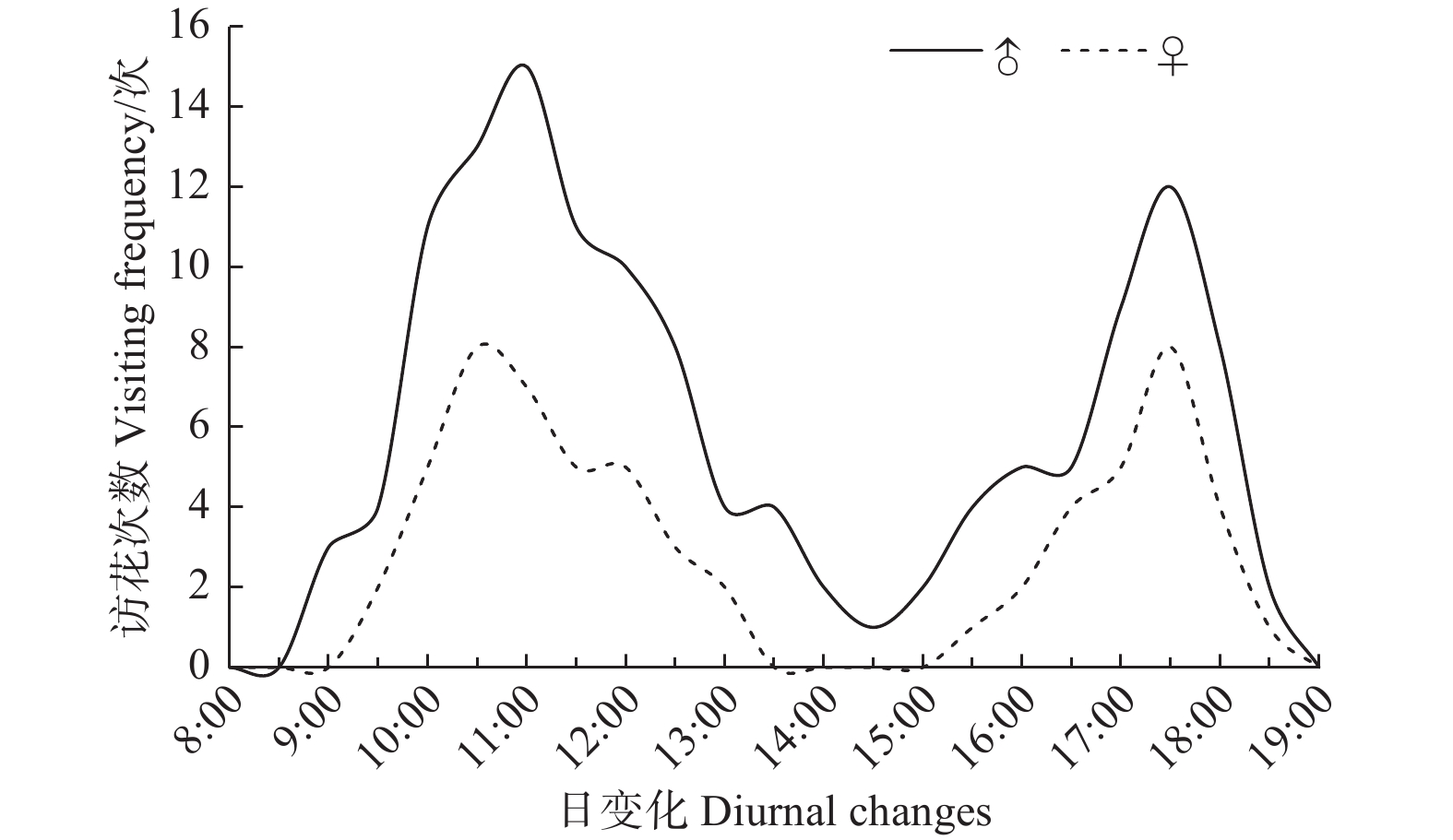
玉带凤蝶飞行活动最早出现在上午8:30,从9:00后开始访花,在下午13:30—15:00达到白昼访花最低谷,于下午19:00访花结束。雌蝶的访花起始时间晚于雄蝶,而结束时间与雄蝶相同。玉带凤蝶有2个明显访花高峰期,第一次高峰期出现在10:00—11:30,第二次高峰期出现在17:00—18:00(图3)。
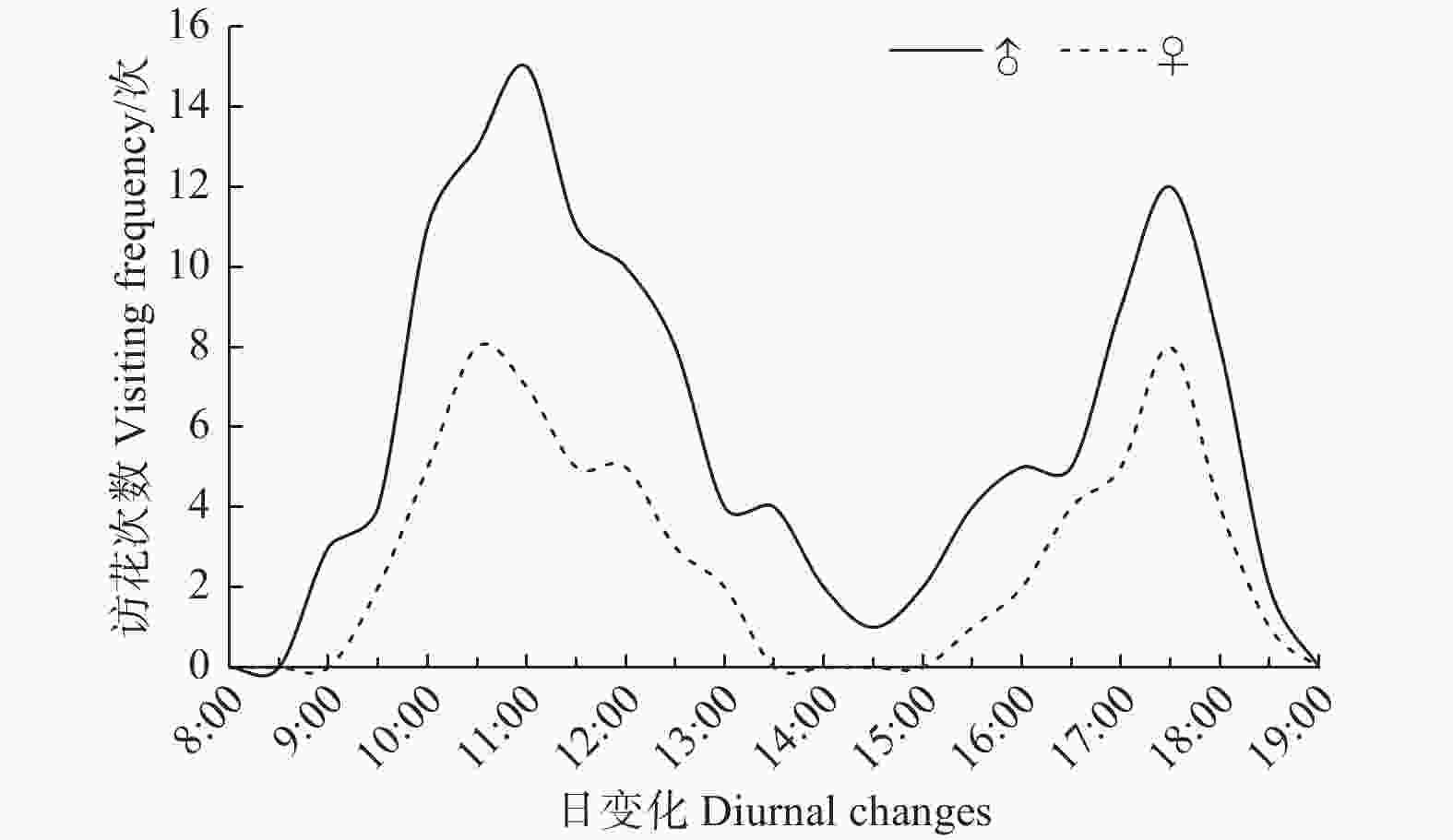
图 3 玉带凤蝶日访花活动规律
Figure 3. Diurnal changes in the frequency of flower visits of P. polytes
-
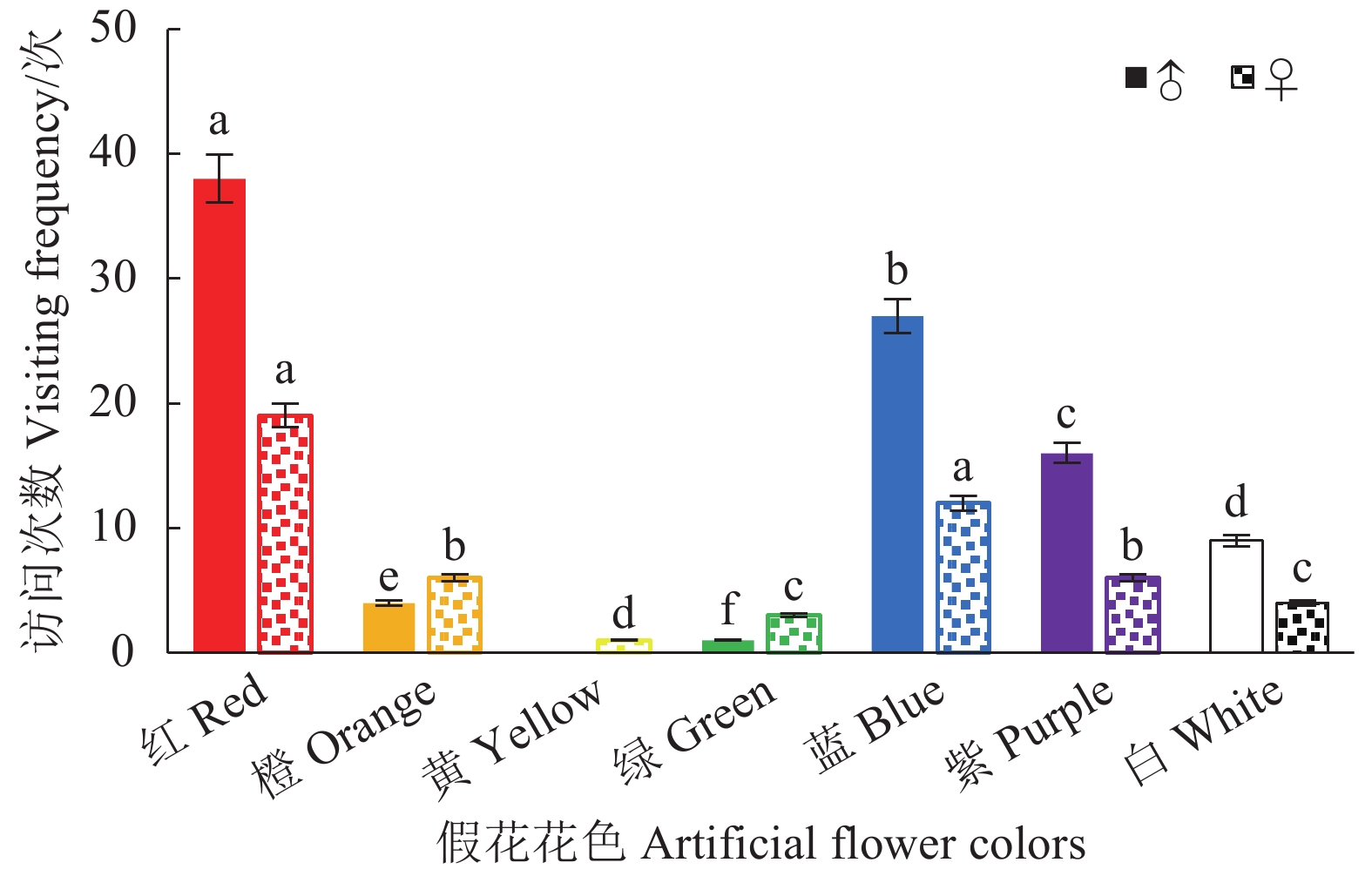
在玉带凤蝶对无气味的假花访花行为试验中发现,雄蝶访问除黄色花外的其他6种颜色的花,对红色花的访问次数最多,占雄蝶总访花次数的40%;蓝色花访问次数第二(28.42%),紫色花访问次数次之(16.84%);对白色(9.47%)、橙色(4.21%)和绿色花(1.05%)的访问次数显著低于红、蓝、紫3种花色(χ2, P < 0.01)。雌蝶对全部颜色的假花均有访问,对红色花的访问次数最多,占雌蝶总访花次数的37.25%;访蓝色花的次数其次(23.53%);对紫色和橙色花的访问次数均占11.76%;对白色、绿色、黄色花访问次数分别占7.84%、5.88%,1.96% (图4)。雌蝶对红色、蓝色、紫色和橙色的访问次数显著高于其他颜色(χ2, P < 0.01)。说明玉带凤蝶对红色花趋性最强,对蓝色和紫色花也有一定的趋性。雌、雄蝶对颜色的访问有差异,雄蝶对颜色的响应高于雌蝶(χ2, P < 0.01)。
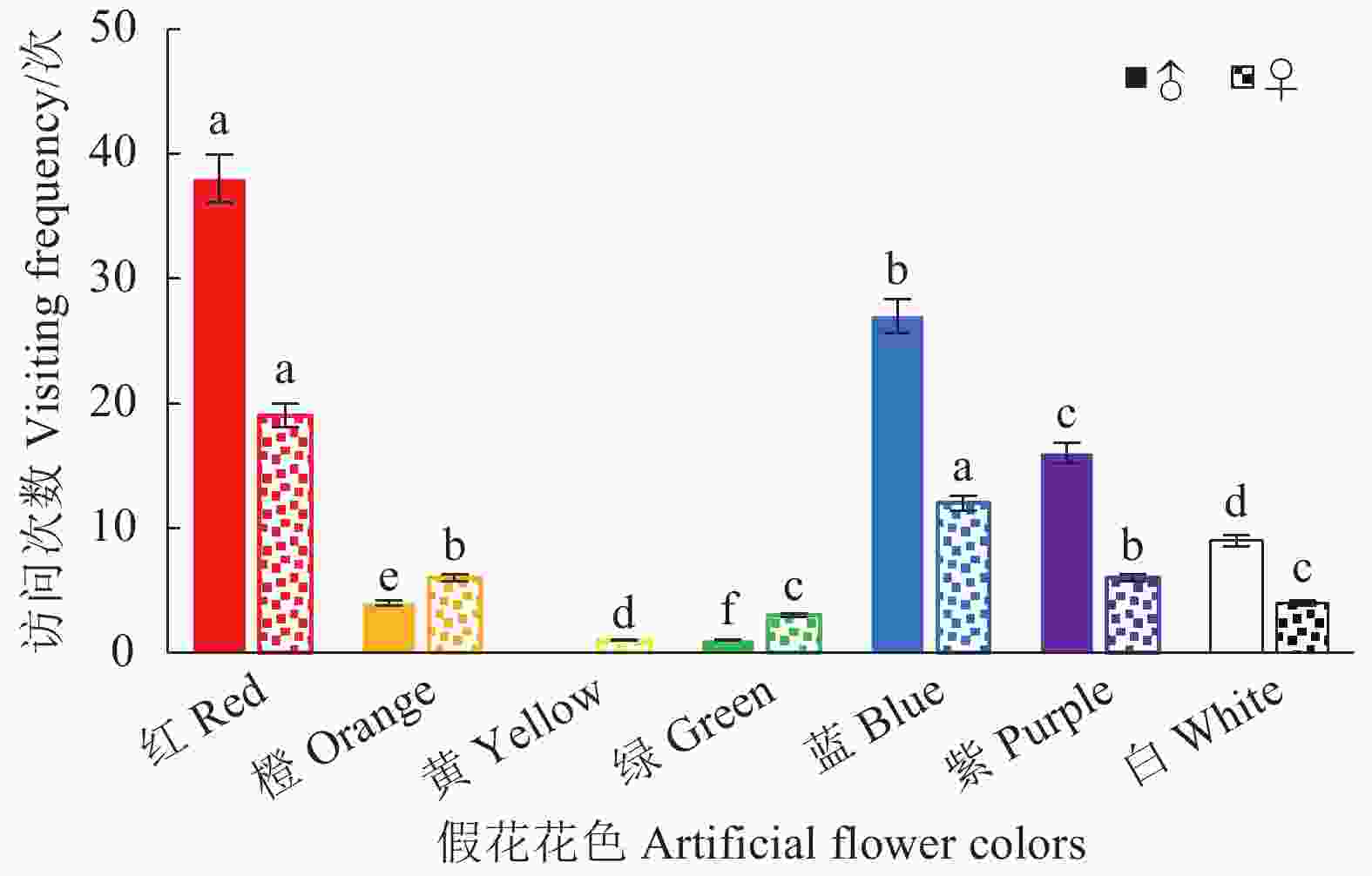
图 4 玉带凤蝶访7种颜色假花次数
Figure 4. Visiting frequencies of P. polytes to artificial flowers of seven colors
-
对10种常见的花香挥发物进行了触角电位试验,发现玉带凤蝶雄蝶EAG反应相对值最高的花香挥发物是β-石竹烯,而雌蝶EAG反应相对值最高的花香挥发物为苯甲醛(图5A)。选择β-石竹烯和苯甲醛在无颜色条件下进行单一花香挥发物诱集试验,结果显示,玉带凤蝶对单一花香挥发物反应不敏感(图5B)。
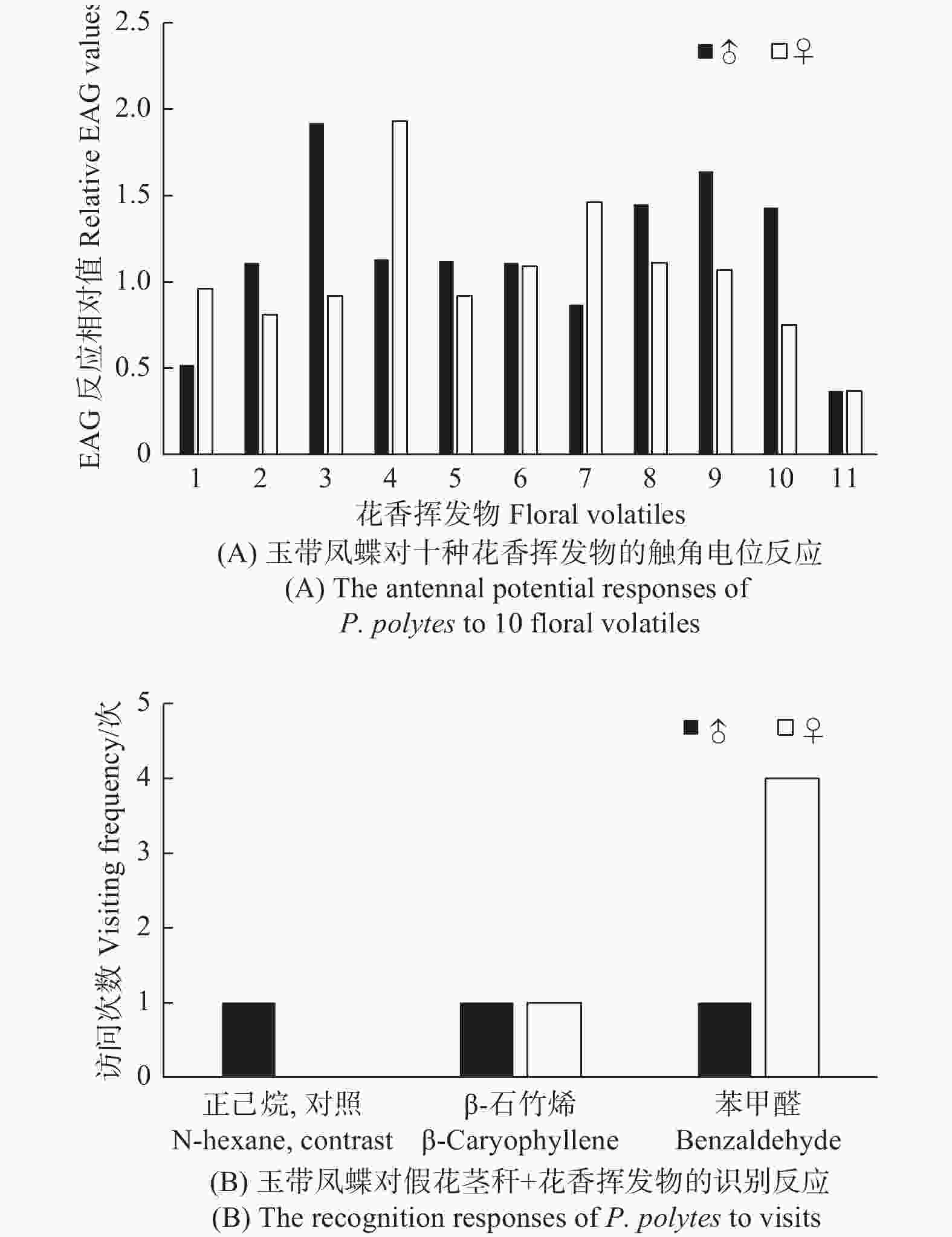
图 5 玉带凤蝶对花香挥发物的识别反应
Figure 5. The recognition responses of P. polytes to visits floral volatile
-
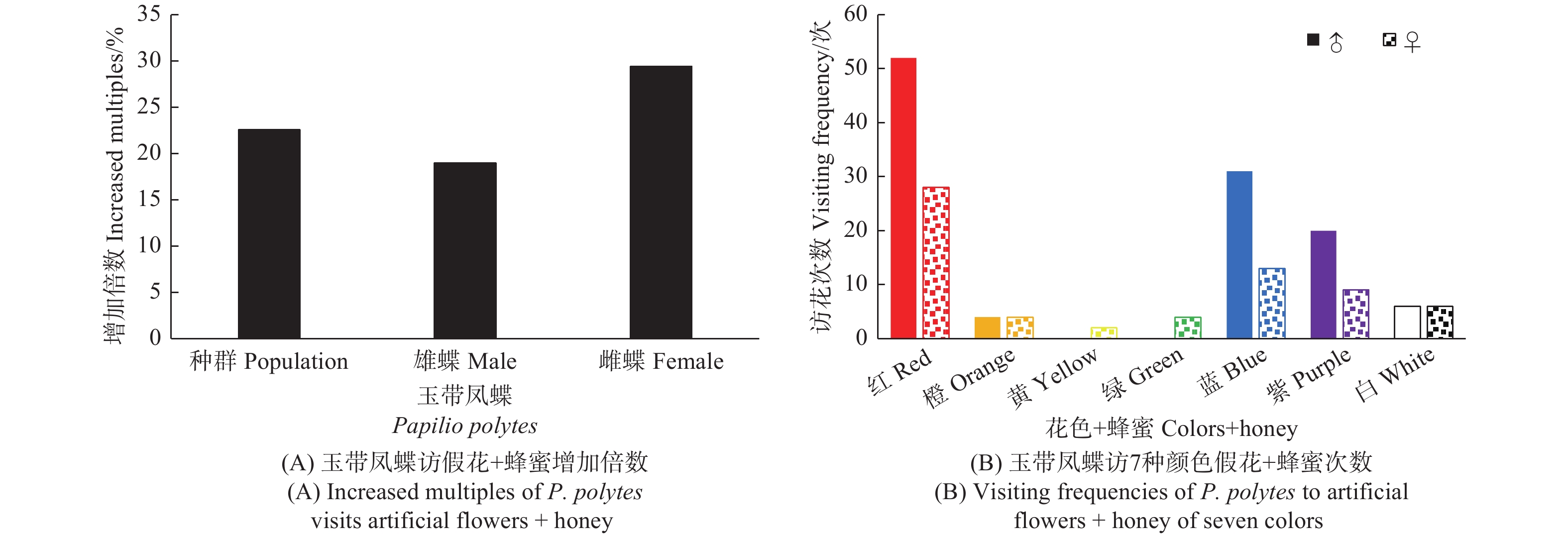
给7种颜色的假花喷洒蜂蜜水后,发现该蝶总体访花次数增加了22.6%,雄蝶增加了18.95%,雌蝶增加了29.41%,较喷洒蜂蜜水前增加显著(t, P < 0.01)(图6A)。访问红色花增加了40%,蓝色花增加了12.82%,紫色花增加了31.82%,而其它颜色的花被访问次数不增加甚至减少(黄色假花访问数据太少,存在偶然性,忽略不计)。雌、雄蝶访花次数有明显区别,雄蝶访花数量明显大于雌蝶,雌蝶访花占整体访花的比例(36.87%)与喷洒蜂蜜水前(34.93%)基本持平,访问红色花增加47.37%,蓝色花增加了8.33%,紫色花增加了50%;雄蝶访问红色花增加36.84%,蓝色花增加了14.81%,紫色花增加了25%(图4、图6B)。
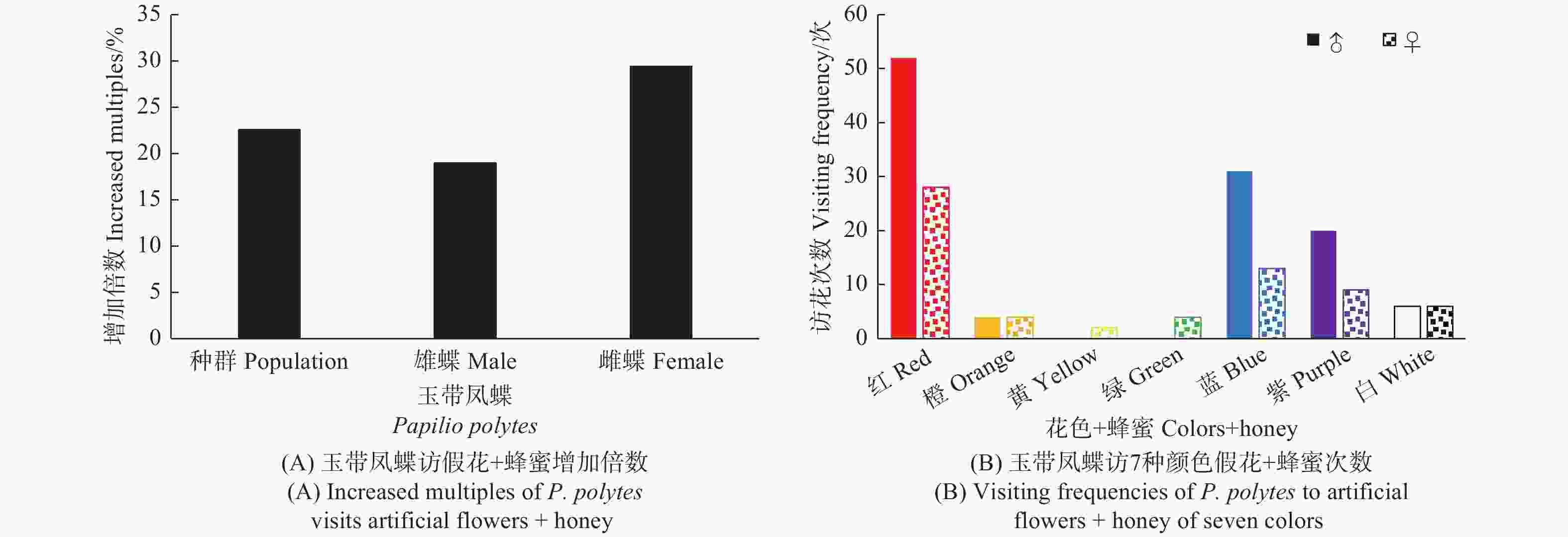
图 6 玉带凤蝶访7种颜色假花+蜂蜜行为
Figure 6. Behavior of P. polytes to visits artificial flowers of seven colors + honey
-
在单一花香挥发物基础上加上红色花冠后,蝴蝶诱集量大大增加(图7A~B),玉带凤蝶访问红色花冠+β-石竹烯后访问量增加了90倍,其中雄蝶增加了120倍,雌蝶增加了60倍,诱集效果显著(t, P < 0.01)(图7A);苯甲醛添加红色花冠后增加了29.8倍,其中雄蝶增加了62倍,雌蝶增加了21.75倍,诱集效果同样显著(t, P < 0.01)(图7B)。玉带凤蝶种群访问红色花冠+β-石竹烯较单一红色花冠只增加了3.16倍,雌、雄蝶均增加了3.16倍(图7C);玉带凤蝶种群对红色花冠+苯甲醛访花次数较仅红色花冠增加了2.61倍,其中雄蝶增加了1.63倍,雌蝶增加了4.58倍(图7D)。试验结果表明,单一颜色诱集蝴蝶远比单一花香挥发物诱集的蝴蝶多(t, P < 0.01),说明颜色与花香挥发物比较,颜色在觅食过程中扮演着更重要的作用。
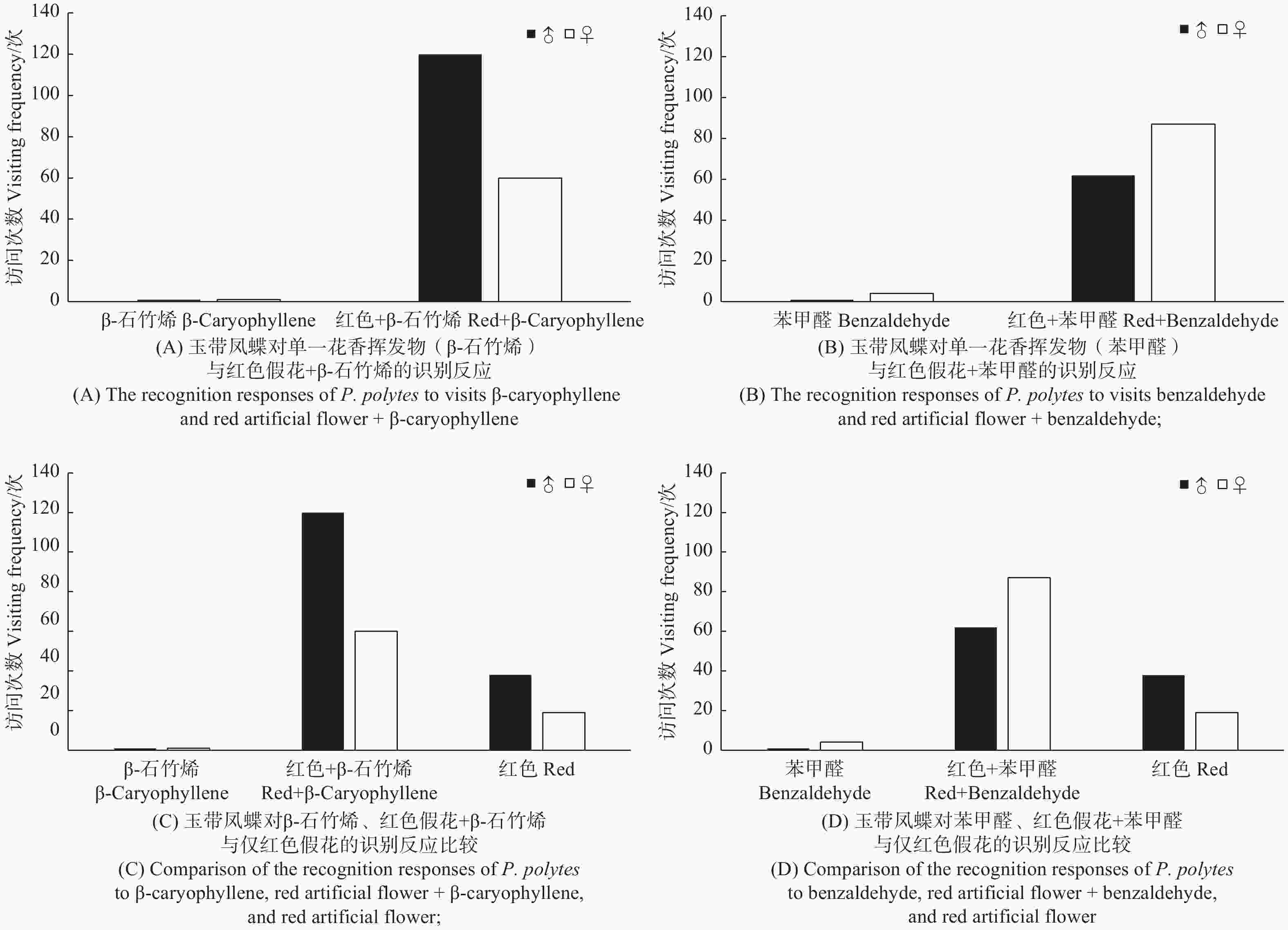
图 7 玉带凤蝶访问花香挥发物行为
Figure 7. Behavior of P. polytes to visits floral volatiles
-
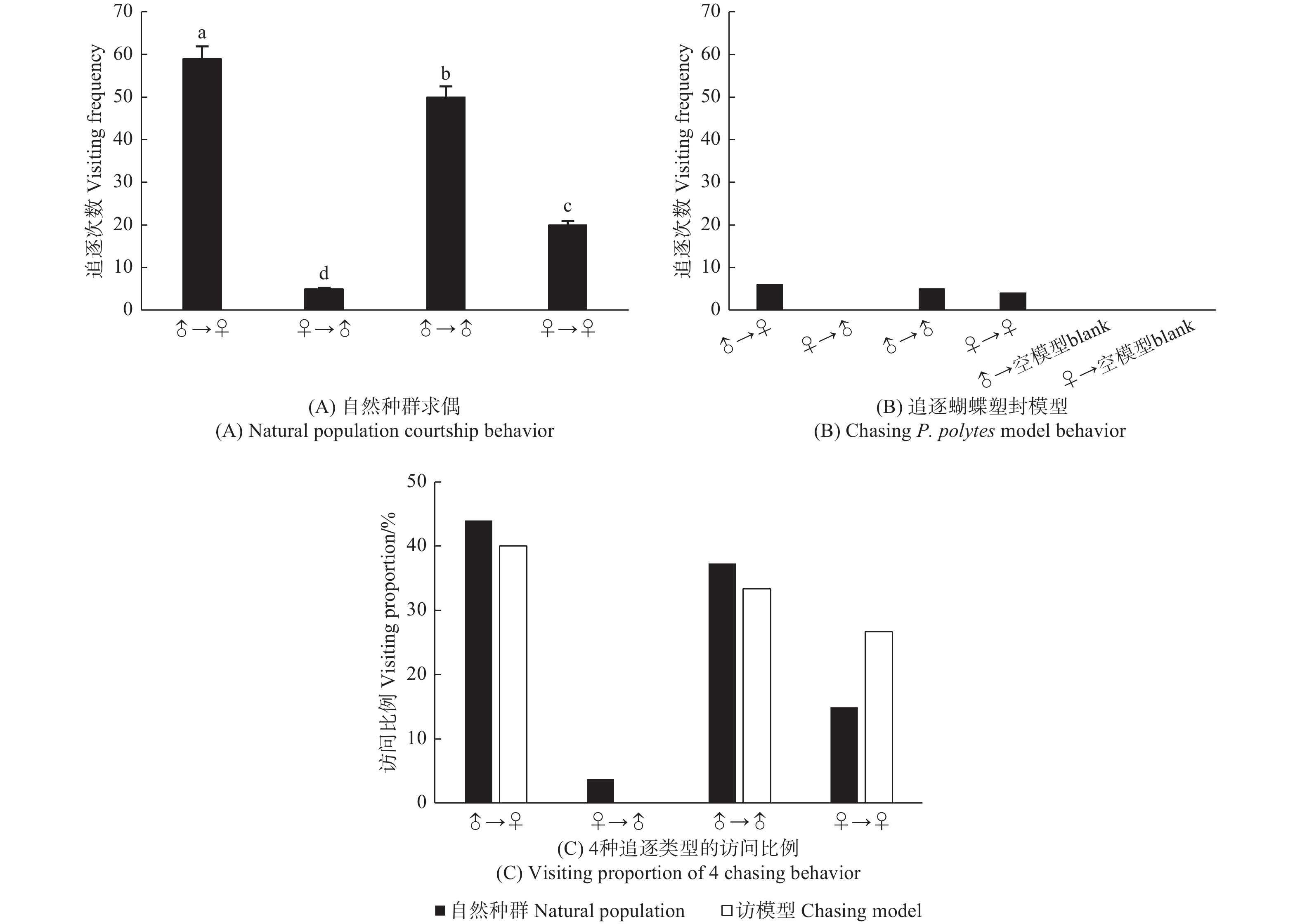
玉带凤蝶自然种群求偶有雄蝶追雄蝶、雄蝶追雌蝶、雌蝶追雌蝶和雌蝶追雄蝶的4种现象。一天中发生求偶追逐134次,其中,雄蝶追逐雌蝶59次(44.03%),雄蝶追逐雄蝶50次(37.31%),雌蝶追逐雌蝶20次(14.93%),雌蝶追逐雄蝶5次(3.73%)。玉带凤蝶雄蝶在求偶过程中占据主动地位,追逐雌蝶次数显著高于其它3种追逐行为(χ2, P < 0.01),雌蝶追逐雄蝶最少,还有同性之间追逐行为(图8A)。
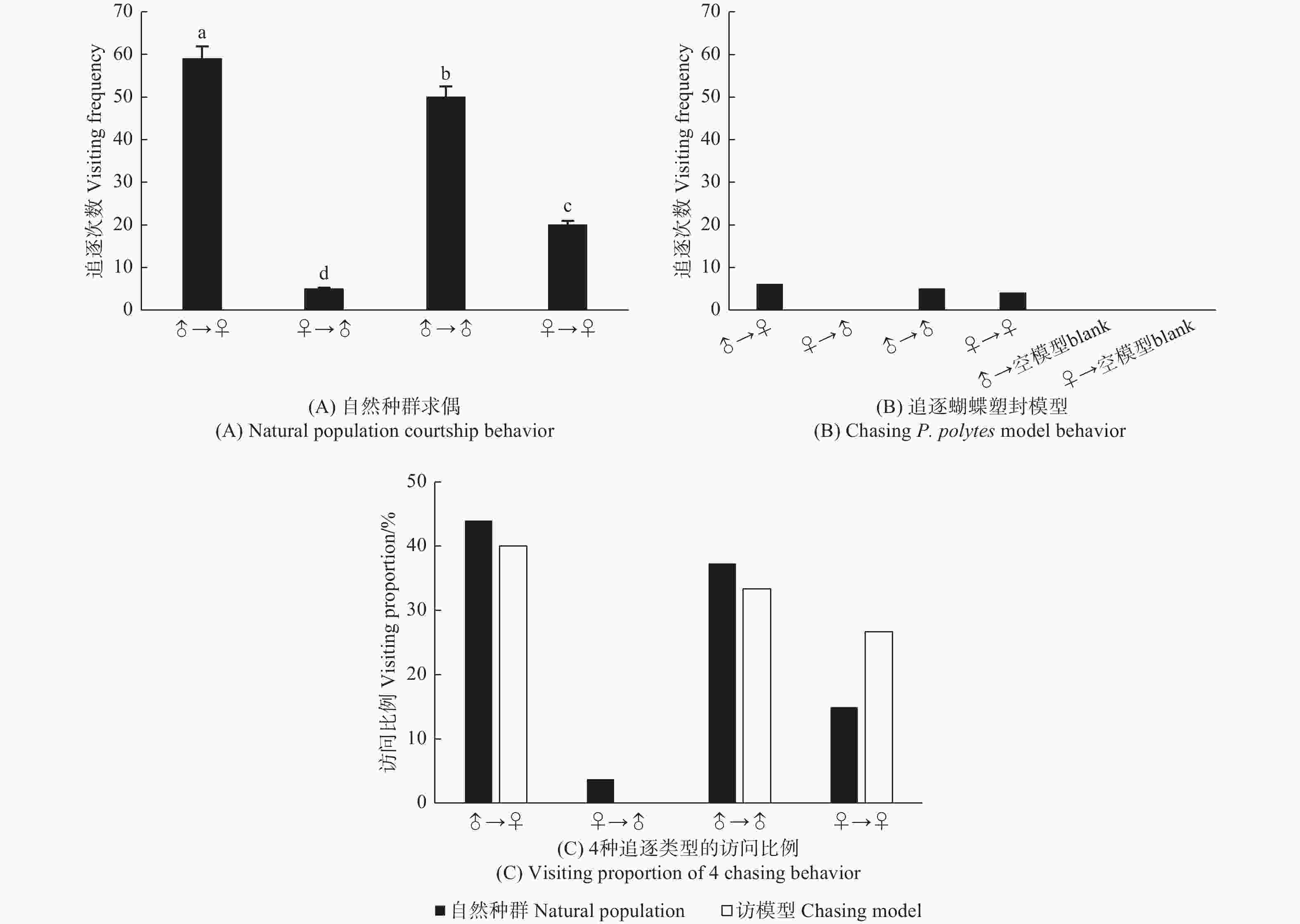
图 8 玉带凤蝶自然种群求偶和追逐模型行为
Figure 8. Natural population courtship and chasing model behavior of P. polytes
-
当在网室内悬挂雌、雄蝶翅的模型和透明空模型时,玉带凤蝶对模型的追逐有3种现象,雄蝶追逐雌蝶模型(40%)、雄蝶追逐雄蝶模型(33.33%)和雌蝶追逐雌蝶模型(26.67%),与自然种群相似(图8B,C)。试验结果表明,玉带凤蝶仅通过颜色基本上可以识别配偶。玉带凤蝶一天中对模型追逐次数只有15次,显著低于自然种群求偶时的134次,说明在求偶时,化学信号起到重要的作用。
-
分析了玉带凤蝶的虫体挥发物,发现雌、雄蝶的总挥发物数量为14种,其中,雌、雄蝶有7种挥发物为共有挥发物,雄蝶有4种独有的挥发物,雌蝶有3种独有挥发物(表1)。
表 1 玉带凤蝶虫体挥发物
Table 1. Volatile organic compounds of P. polytes
挥发物名称 (CAS编号)
Volatile organic compounds(CAS number)雄蝶
Male雌蝶
Female反式-2-戊烯醛 (1576-87-0) 29.86% 37.06% 7,7-dimethyl-2-methylene-norbornane (471-84-1) 49.53% 2.94% 7-methylidenebicyclo[4.1.0]heptane (54211-14-2) 1.17% − (15232-88-9) − 1.55% 烯丙基醚 (557-40-4) 1.34% − 乙烷 (74-84-0) − 1.39% (40801-29-4) 0.89% 2.84% 巴伦西亚橘烯 (4630-07-3) 1.01% 1.95% 二苯甲烷 (101-81-5) 0.25% 1.69% 正十五烷 (629-62-9) 0.78% − 邻苯二甲酸二丁酯 (84-74-2) 0.90% 1.24% 3-(4-羟基苯基)-7-甲氧基-4H-色烯-4-酮 (486-63-5) − 1.87% 1-二十醇 (629-96-9) 10.02% 47.47% 正二十六烷 (630-31-3) 4.24% − -
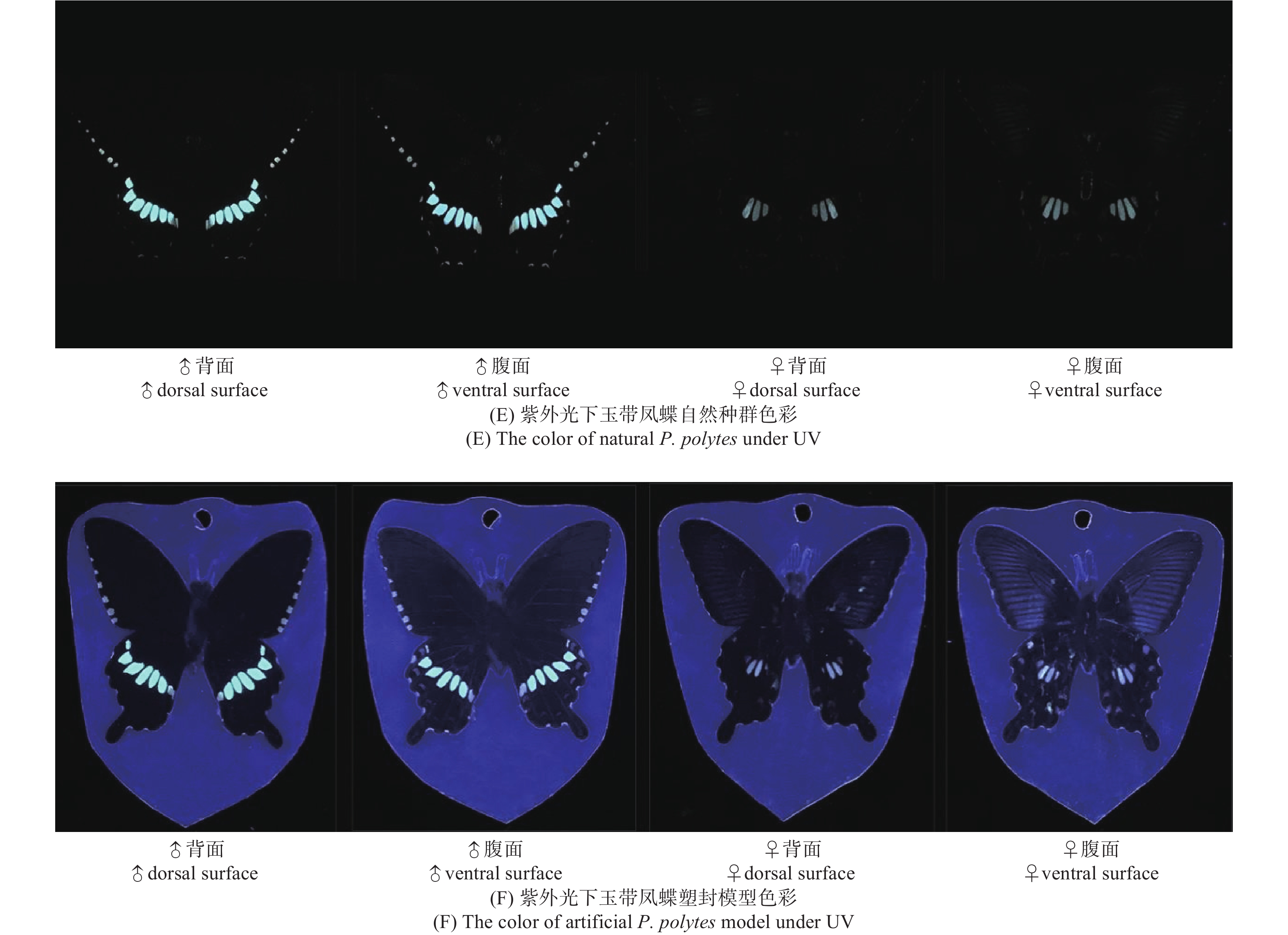
玉带凤蝶雌、雄蝶的翅膀颜色和图案在自然光下有显著的区别(图9A)。玉带凤蝶与塑封模型在视觉上差异不大,能够清晰地反映出雌、雄蝶间的差异(图9B)。进一步通过光谱分析可知,雌、雄蝶自然种群光谱反射率在379~529 nm波段内基本一致,且均在379~429 nm波段形成反射率高峰;雄蝶在529 nm波段后基本呈平滑走势,雌蝶在529 nm后反射率逐渐上升,最终在734~770 nm形成第二次高峰。光谱分析指出,在自然种群与塑封模型的光谱分析走势类似的情况下,雄蝶塑封模型的光谱反射率虽低于自然种群,但在重要的反射率高峰阶段相似。光谱分析同时指出,塑封蝴蝶模型与标本比较,在可见光和紫外光范围内,有一定差异,但塑封蝴蝶模型反射率更强,更易识别(图9C~D)。说明塑封模型对蝴蝶寻找配偶的干扰较少。在紫外光范围内,玉带凤蝶雌、雄蝶的翅膀图案有显著的区别,塑封模型中的蝴蝶的紫外翅膀图案区别更为显著,易于识别雌雄(图9E~F)。
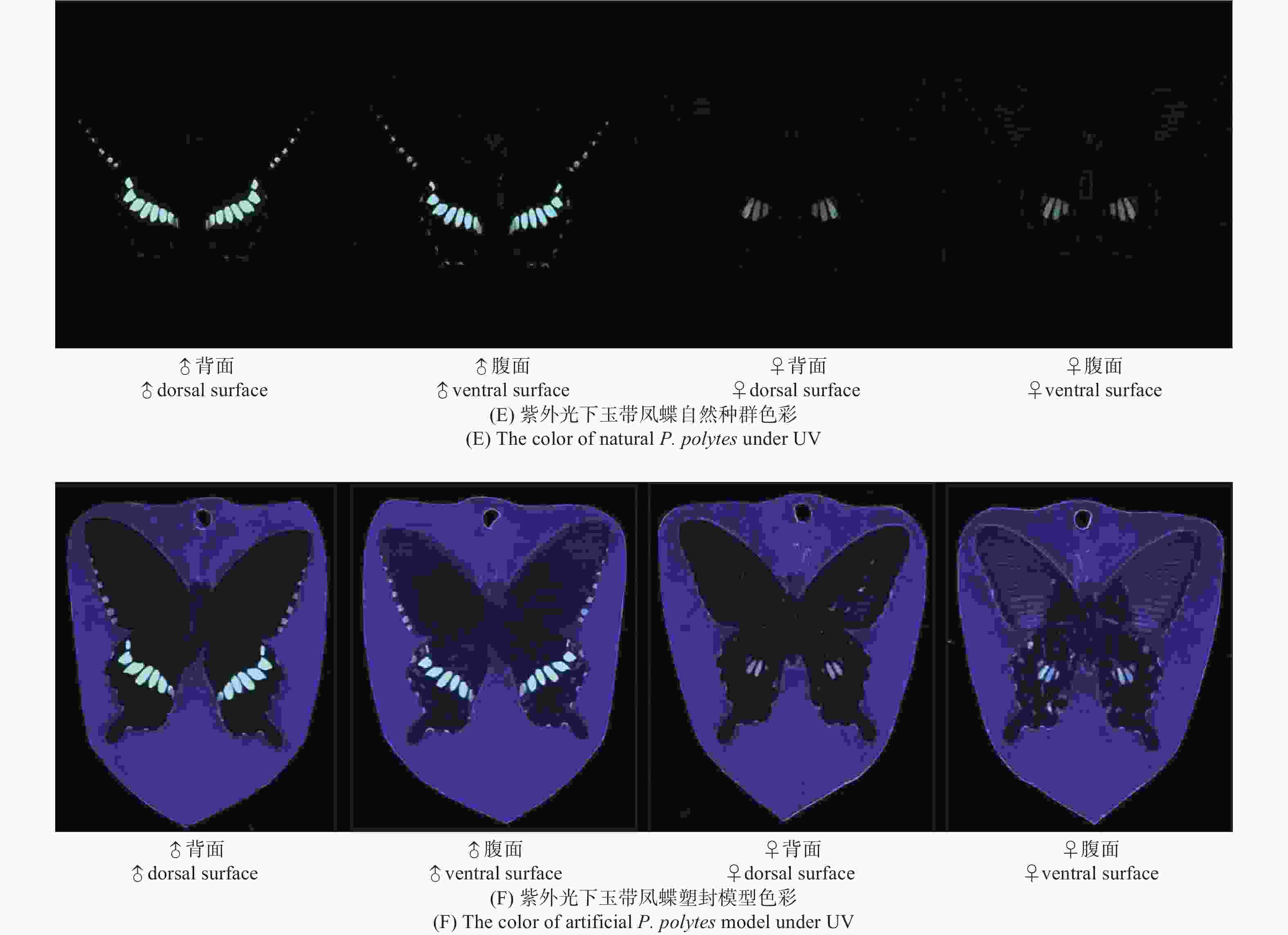
图 9 玉带凤蝶雌、雄蝶自然种群及塑封模型色彩
Figure 9. The color of females and males of natural P. polytes and artificial P. polytes model
-
玉带凤蝶在觅食过程中,对红色(622~770 nm)、蓝色(440~485 nm)和紫色花(380~440 nm)显示出了明显的颜色趋性,颜色加蜂蜜水诱集了更多的蝴蝶访花,说明颜色和气味都是玉带凤蝶觅食的指示因子。颜色和气味在玉带凤蝶觅食中,哪一个占有更高的权重?我们将两者进行了分析比较,玉带凤蝶访问单一花香挥发物时,数量很少,而访问单一颜色时,数量远高于访问单一花香挥发物,从访问单一花香挥发物与花香挥发物加颜色的蝴蝶数量分析,加上颜色后,增加的诱集倍数达到60%~120%,说明颜色对玉带凤蝶的诱集超过气味;而访问单一颜色与加上花香挥发物后比较,增加倍数只有1%~5%,说明气味对玉带凤蝶的诱集作用较小。
在求偶过程中,玉带凤蝶对塑封模型的访问与自然种群求偶的追逐类型及比例相似,说明玉带凤蝶仅依靠视觉即可较准确地识别雌雄。视觉上的信号包括了翅的色彩、斑纹、及形状等特征[10, 13, 19-21],但具体是哪一种,还需进一步研究。玉带凤蝶雌、雄蝶翅膀在自然光及紫外光下色彩及斑纹区别明显,可能是该蝴蝶识别配偶的一个重要指标。玉带凤蝶追逐模型的次数远远低于自然种群之间的相互追逐,说明气味在求偶过程中同样发挥着作用。我们分析了玉带凤蝶雌、雄蝶的挥发物,发现雄蝶有4种独有的挥发物,雌蝶有3种独有挥发物,挥发物的差异可能是雌、雄蝶识别配偶的重要信息。
玉带凤蝶在觅食和求偶的过程中,颜色和(或)斑纹的诱导作用高于气味,这可能是颜色和(或)斑纹的指示作用更显著或者在检测视觉信号时的成本更低等因素造成的。自然界中挥发物种类比较复杂,由多种植物和动物的挥发物组成,而且易受环境影响,在搜寻化学信号时会花费较大的成本[18]。在长期的进化过程中,玉带凤蝶演化出以视觉为主的识别系统,可能通过颜色或斑纹发现目标,后通过花蜜和雌、雄蝶特有的气味精确识别食物和配偶。
-
本研究表明,在觅食时,玉带凤蝶对花色及气味均有反应;在利用权重上,对花色的趋性显著高于气味,觅食以视觉信号为主,嗅觉信号为辅。在求偶时,玉带凤蝶仅利用视觉即可辨别雌雄,同时虫体挥发物也起到了不可或缺的作用;在利用权重上,更偏向于视觉信号为主,嗅觉信号为辅。
玉带凤蝶访花及求偶过程中的视觉及嗅觉识别行为
Visual and Olfactory Response of Papilio polytes During Foraging and Courtship
-
摘要:
目的 解析玉带凤蝶的视觉和嗅觉在觅食和求偶过程中的识别和选择规律,讨论玉带凤蝶的视觉和嗅觉在觅食和求偶过程中的生态学意义。 方法 通过玉带凤蝶自然种群访问颜色、气味和颜色加气味的试验,自然种群求偶和访塑封模型试验,分析玉带凤蝶觅食及求偶时的视觉及嗅觉利用权重。 结果 在无气味的7种颜色的假花条件下,玉带凤蝶对红、蓝和紫色显示出了明显的颜色趋性。在假花上喷洒蜂蜜水后,雌、雄蝶访花次数均增加3.16倍;在无颜色的花香挥发物(β-石竹烯、苯甲醛)试验中,玉带凤蝶访问数量很少,增加颜色后,雌、雄蝶访花次数增加了60~120倍,说明颜色的诱集作用超过气味;而访问单一颜色与加上花香挥发物后比较,增加倍数只有1%~5%,说明气味对玉带凤蝶的诱集作用较小。在求偶过程中,玉带凤蝶对塑封模型的访问与自然种群求偶的追逐类型及比例相似,说明玉带凤蝶可利用视觉识别雌雄。在自然光和紫外光下,玉带凤蝶雌、雄蝶翅膀颜色和图案区别明显,可能是该蝶识别配偶的一个指标。 结论 玉带凤蝶在觅食及求偶时均以视觉为主、嗅觉为辅。 Abstract:Objective To analyze the recognition and selection rules of Papilio polytes' visual and olfactory response during foraging and courtship, and to further discuss their ecological significance. Method Through color-visit, odor-visit, color plus odor-visit, courtship and artificial models-visit experiments in natural population butterflies to analyze the visual and olfactory weight factor of P. polytes in foraging and courtship. Result Under the condition of odorless 7-color artificial flowers, the P. polytes showed obvious color tendency to red, blue, and purple. After spraying honey on the artificial flowers, the frequency of female and male butterfly foraging increased by 3.16 times. In the colorless floral volatiles foraging experiment (β-caryophyllene, benzaldehyde), the frequency of butterfly foraging was less; after add odor to color, the frequency of female and male butterfly foraging increased by 60 to 120 times, indicating that the attractant effect of color exceeds odor. However, compared with the single color and floral volatiles, the increase ratio was only 1%~5%, indicating that the effect of scent on the collection of P. polytes was small. In the process of courtship, the visit to the plastic seal model was similar to that of the natural population in the type and proportion of courtship, indicating that the butterfly could use color and stripes to identify male and female. Conclusion P. polytes is mainly visual and supplemented by olfaction when foraging and courtship. -
Key words:
- Papilio polytes
- / foraging behavior
- / courtship behavior
- / visual response
- / olfactory response
-
图 1 7种假花和可见光范围内光谱反射率
Figure 1. Artificial flowers with seven colors and their spectral reflectance in visible light
图 3 玉带凤蝶日访花活动规律
Figure 3. Diurnal changes in the frequency of flower visits of P. polytes
图 4 玉带凤蝶访7种颜色假花次数
Figure 4. Visiting frequencies of P. polytes to artificial flowers of seven colors
图 5 玉带凤蝶对花香挥发物的识别反应
Figure 5. The recognition responses of P. polytes to visits floral volatile
图 6 玉带凤蝶访7种颜色假花+蜂蜜行为
Figure 6. Behavior of P. polytes to visits artificial flowers of seven colors + honey
图 8 玉带凤蝶自然种群求偶和追逐模型行为
Figure 8. Natural population courtship and chasing model behavior of P. polytes
图 9 玉带凤蝶雌、雄蝶自然种群及塑封模型色彩
Figure 9. The color of females and males of natural P. polytes and artificial P. polytes model
表 1 玉带凤蝶虫体挥发物
Table 1. Volatile organic compounds of P. polytes
挥发物名称 (CAS编号)
Volatile organic compounds(CAS number)雄蝶
Male雌蝶
Female反式-2-戊烯醛 (1576-87-0) 29.86% 37.06% 7,7-dimethyl-2-methylene-norbornane (471-84-1) 49.53% 2.94% 7-methylidenebicyclo[4.1.0]heptane (54211-14-2) 1.17% − (15232-88-9) − 1.55% 烯丙基醚 (557-40-4) 1.34% − 乙烷 (74-84-0) − 1.39% (40801-29-4) 0.89% 2.84% 巴伦西亚橘烯 (4630-07-3) 1.01% 1.95% 二苯甲烷 (101-81-5) 0.25% 1.69% 正十五烷 (629-62-9) 0.78% − 邻苯二甲酸二丁酯 (84-74-2) 0.90% 1.24% 3-(4-羟基苯基)-7-甲氧基-4H-色烯-4-酮 (486-63-5) − 1.87% 1-二十醇 (629-96-9) 10.02% 47.47% 正二十六烷 (630-31-3) 4.24% −  下载: 导出CSV
下载: 导出CSV
-
[1] Sison-Mangus M P. Beauty in the eye of the beholder: the two blue opsins of lycaenid butterflies and the opsin gene-driven evolution of sexually dimorphic eyes[J]. Journal of Experimental Biology, 2006, 209(16): 3079-3090. doi: 10.1242/jeb.02360 [2] Arikawa K, Iwanaga T, Wakakuwa M, et al. Unique temporal expression of triplicated long-wavelength opsins in developing butterfly eyes[J]. Frontiers in Neural Circuits, 2017, 11: 96. doi: 10.3389/fncir.2017.00096 [3] Chen P J, Awata H, Matsushita A, et al. Extreme spectral richness in the eye of the common bluebottle butterfly, Graphium sarpedon[J]. Frontiers in Physiology, 2016, 4(10): 107. [4] Perry M, Kinoshita M, Saldi G, et al. Molecular logic behind the three-way stochastic choices that expand butterfly colour vision[J]. Nature, 2016, 535(7611): 280. doi: 10.1038/nature18616 [5] Mcculloch K J, Yuan F, Zhen Y, et al. Sexual dimorphism and retinal mosaic diversification following the evolution of a violet receptor in butterflies[J]. Molecular Biology & Evolution, 2017, 34(9): 9. [6] Arikawa K. The eyes and vision of butterflies[J]. The Journal of Physiology, 2017, 595(16): 5457-5464. doi: 10.1113/JP273917 [7] 王 华, 李承哲, 陈晓鸣, 等. 红锯蛱蝶觅食过程中的视觉和嗅觉反应[J]. 林业科学研究, 2016, 29(5):759-763. [8] Balkenius A, Rosén W, Kelber A. The relative importance of olfaction and vision in a diurnal and a nocturnal hawkmoth[J]. Journal of Comparative Physiology a-Neuroethology Sensory Neural and Behavioral Physiology, 2006, 192(4): 431-437. doi: 10.1007/s00359-005-0081-6 [9] Pohl N B, Wyk J V, Campbell D R. Butterflies show flower color preferences but not constancy in foraging at four plant species[J]. Ecological Entomology, 2011, 36(3): 290-300. doi: 10.1111/j.1365-2311.2011.01271.x [10] 王翻艳. 大帛斑蝶成虫行为学观察及其求偶机制研究[D]. 北京, 中国林业科学研究院, 2015. [11] Ômura H. Foraging behavior of adult butterflies and its semiochemicals as olfactory signals[J]. Comparative Physiology and Biochemistry, 2006, 23(3): 134-142. [12] 唐宇翀. 蝴蝶觅食过程中的嗅觉和视觉行为反应研究[D]. 北京, 中国林业科学研究院, 2013. [13] 李承哲. 基于蝴蝶成虫行为学的两性求偶识别机制及蝴蝶飞舞景观构建[D]. 北京, 中国林业科学研究院, 2017. [14] Honda K, Mura H, Hayashi N. Identification of floral volatiles from Ligustrum japonicum that stimulate flower-visiting by cabbage butterfly, Pieris rapae[J]. Journal of Chemical Ecology, 1998, 24(12): 2167-2180. doi: 10.1023/A:1020750029362 [15] Tang Y C, Zhou C L, Chen X M, et al. Visual and olfactory responses of seven butterfly species during foraging[J]. Journal of Insect Behavior, 2013, 26(3): 387-401. doi: 10.1007/s10905-012-9358-0 [16] Yuya F, Yosuke T, Ibrahim F S, et al. Flower-visiting butterflies avoid predatory stimuli and larger resident butterflies: testing in a butterfly pavilion[J]. Plos One, 2016, 11(11): e0166365. doi: 10.1371/journal.pone.0166365 [17] Salcedo C. Pollen preference forPsychotria sp. is not learned in the passion flower butterfly, Heliconius erato[J]. Journal of Insect Science, 2011, 11(25): 25. [18] Maciasmuñoz A, Smith G, Monteiro A, et al. Transcriptome-wide differential gene expression in Bicyclus anynana butterflies: female vision-related genes are more plastic[J]. Molecular Biology & Evolution, 2015, 33(1): 1. [19] Rutowski R L, Rajyaguru P K. Male-specific iridescent coloration in the pipevine swallowtail (Battus philenor) is used in mate choice by females but not sexual discrimination by males[J]. Journal of Insect Behavior, 2013, 26(2): 200-211. doi: 10.1007/s10905-012-9348-2 [20] Li C Z, Wang H, Chen X M, et al. Role of visual and olfactory cues in sex recognition in butterfly Cethosia cyane cyane[J]. Scientific Reports, 2017, 7(1): 5033. doi: 10.1038/s41598-017-04721-6 [21] 李承哲, 王 华, 陈晓鸣, 等. 白带锯蛱蝶雌蝶利用视觉和嗅觉信息识别两性[J]. 林业科学研究, 2017, 30(4):648-652. [22] Dinesh A S, Venkatesha M G. Analysis of the territorial, courtship and coupling behavior of the Hemipterophagous butterfly, Spalgis epius, (Westwood) (Lepidoptera: Lycaenidae)[J]. Journal of Insect Behavior, 2013, 26(2): 149-164. doi: 10.1007/s10905-012-9341-9 [23] Pinzari M. A comparative analysis of mating recognition signals in graylings: Hipparchia statilinus vs. H. semele (Lepidoptera: Nymphalidae, Satyrinae)[J]. Journal of Insect Behavior, 2009, 22(3): 227-244. doi: 10.1007/s10905-008-9169-5 [24] Stavenga D G, Arikawa K. Evolution of color and vision of butterflies[J]. Arthropod Structure & Development, 2006, 35(4): 307-318. [25] Giraldo M A, Stavenga D G. Sexual dichroism and pigment localization in the wing scales of Pieris rapae butterflies[J]. Proceedings Biological Sciences, 2007, 274(1606): 97. doi: 10.1098/rspb.2006.3708 [26] Obara Y, Koshitaka H, Arikawa K. Better mate in the shade: enhancement of male mating behaviour in the cabbage butterfly, Pieris rapae crucivora, in a UV-rich environment[J]. Journal of Experimental Biology, 2008, 211(23): 3698-3702. doi: 10.1242/jeb.021980 [27] Bernard G D, Remington C L. Color vision in Lycaena butterflies: spectral tuning of receptor arrays in relation to behavioral ecology[J]. Proceedings of The National Academy of Sciences of The United States of America, 1991, 88(7): 2783-2787. doi: 10.1073/pnas.88.7.2783 [28] Kan E, Hidaka T. Role of male scent in the mating behavior ofPieris melete Ménétriès (Lepidoptera: Pieridae)[J]. Journal of Ethology, 1997, 15(2): 87-93. doi: 10.1007/BF02769393 [29] 尹小刚, 程 方, 刘良源. 南昌地区玉带凤蝶饲养及生物学特性研究[J]. 江西科学, 2015, 33:75-78. [30] 廖 宇. 玉带凤蝶生物学特性研究[D]. 福州, 福建农林大学, 2009. [31] 陈晓鸣, 周成理, 史军义, 等. 中国观赏蝴蝶[M]. 北京: 中国林业出版社, 2008.67-68. [32] Zheng W Y, Chen Y H, Zhang X T, et al. Analysis of volatiles in Paeonia obovata flowers by HS-SPME-GC-MS[J]. Chemistry of Natural Compounds, 2016, 52(5): 922-923. doi: 10.1007/s10600-016-1817-y -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5987
- HTML全文浏览量: 3561
- PDF下载量: 79
- 被引次数: 0
