

-
檫木(Sassafras tzumu(Hemsley)Hemsley)隶属樟科(Lauraceae)檫木属(Sassafras J. Presl),又名檫树、黄楸木、桐梓树、鹅脚板,落叶大乔木,自然分布于我国长江流域及其以南地区海拔100~1 900 m的森林中,是山地次生林的主要先锋树种和优势树种[1-2]。檫木早期生长快,树冠常居于林冠上层,早春开花,花色明黄,先花后叶,秋季叶色转红或黄,色彩醒目,是重要的彩叶景观树种之一[3-4];檫木树干通直,木材花纹明显美观,耐腐有香气,也是优良的用材树种[5-7],2017年被国家林业局列入《中国主要栽培珍贵树种参考名录》。
檫木属包括3个种,分别为檫木、台湾檫木(S. randaiense (Hayata) Rehder)和美洲檫木(S. albidum (Nuttall) Nees),为典型的东亚-北美间断分布属[2, 8],研究表明檫木属3个种属于单系发生类群,东亚的2种组成与北美种是相对的姊妹群,并认为台湾檫木源于大陆檫木[9]。不同于美洲檫木明确的花性别特征,台湾檫木和檫木在花性别描述上一直存在争议[10-11]。早期文献记载檫木为雌雄异株[1],沈卓君等[10]认为这种记载是由于早期样本采集时出现了错误,并总结了前人对檫木花性别的描述,包括单性雌雄异株、两性或功能上雌雄异株、杂性异株、花两性以及花近乎两性等,后续通过对大量花的观察认为檫木花为两性花。谷淑芳等[12]和王馨等[13]的研究在一定程度上支持了该观点,并认为檫木花为雌蕊先熟的两性花结构。但到目前为止,包括修订后的《中国植物志》等权威著作尚无对檫木的花性别进行界定[2]。在果实研究方面,仅见沈卓君等[10]与谷淑芳等[12]对果实结构进行过初步观察,对果实整个发育过程规律尚缺乏系统的研究。为深化对檫木花和果实的生物学认知,本试验进一步观察檫木开花与果实发育过程,解析檫木开花与果实发育规律,为檫木种质资源收集评价、保存利用及育种等研究提供参考依据。
-
取样点位于浙江省杭州市富阳区城市森林公园(119°57′ E, 30°03′ N),属亚热带季风性湿润气候,冬冷夏热,春季平均气温19.1℃,夏季平均气温26.5℃,降水充沛。试验共选取6株无病害且树龄超过10 a的檫木并编号,于2018年1—7月取样并观察。
-
自2018年1月起,每周观察并取样1~2次,样品用50% FAA固定48 h后转入70% FAA中保存备用,其中檫木花的采样时间为1—3月,果实采样时间为4—7月,至果实成熟为止。
檫木开花规律:观察并记录檫木混合芽芽鳞开裂期、初花期(开花数 ≥ 5%时记为起点)、盛花期(开花数 ≥ 50%时记为起点)及末花期(开花数 ≥ 95%时记为起点)起始时间[14];用高枝剪于盛花期自每株檫木上剪取30个花枝,统计剪取的花枝上每个混合芽包含的花序数、每个花序花数量,测量花梗长和花径大小。
檫木果实发育规律:自檫木幼果形成开始,每周取样1次,利用游标卡尺测定果实横径与果实纵径(每重复50个单果),测定果实鲜质量(称量50个果实总质量后取平均值),105℃杀青30 min后于70℃烘干至恒质量,试验设置3次重复。计算果形指数、果实含水率和果实干质量比例并绘制果实生长发育曲线,其中:果形指数 = 果实纵径/果实横径;果实含水率 =(果实鲜质量 − 果实干质量)/果实鲜质量;果实干质量比例 = 果实干质量/果实鲜质量。
檫木花与果实结构:样品采集固定后,利用MDG33型体视显微镜(徕卡公司)和石蜡切片观察檫木花和果实结构。其中石蜡切片制作方法参照王馨等[15]方法,经脱水、透明处理后用石蜡包埋,切片厚度6~8 μm,利用Imager A2型研究级显微镜(蔡司公司)观察并拍照。
采用Excel2003软件进行数据处理,利用Origin2018软件作图。
-
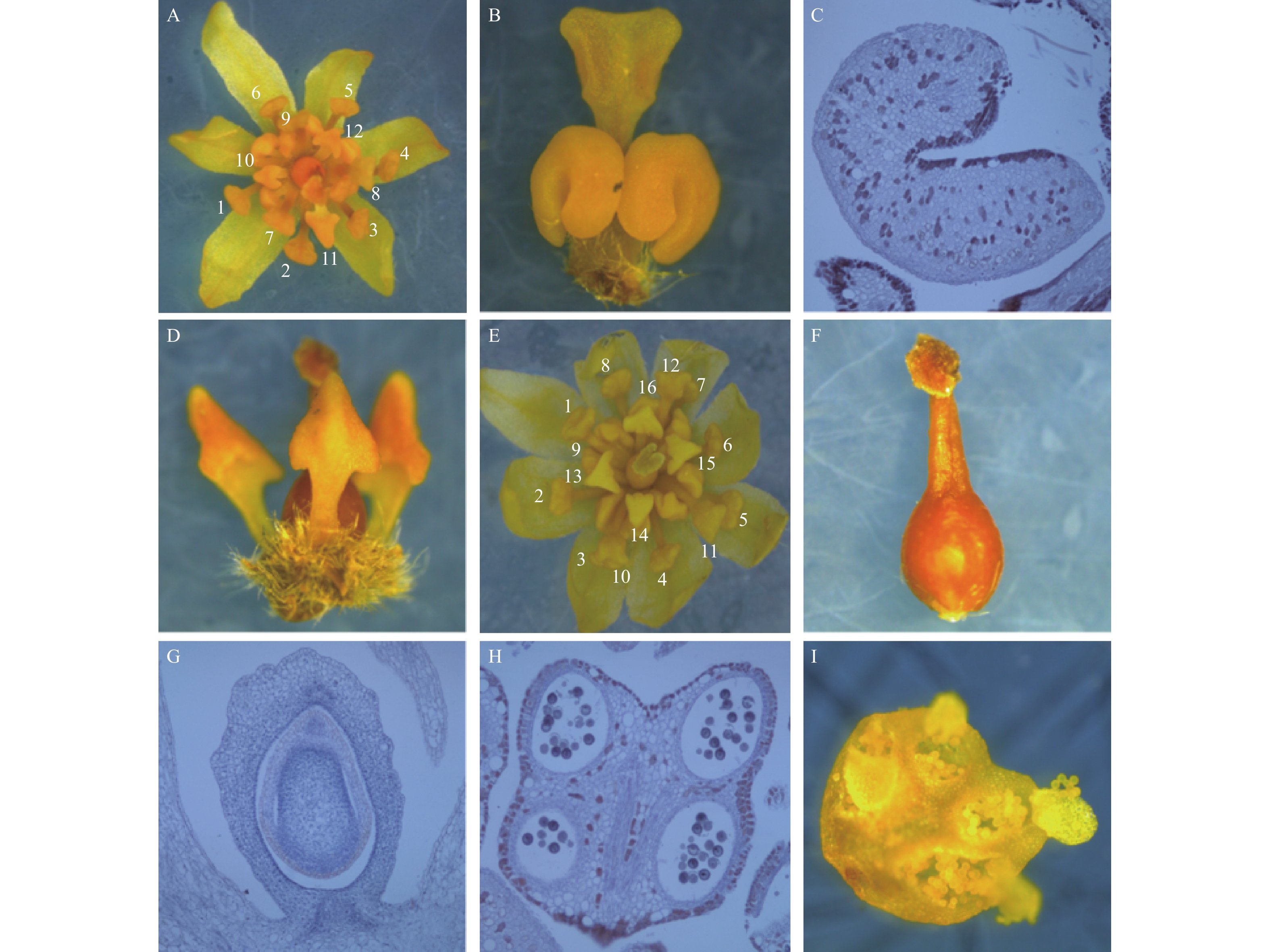
利用体视显微镜和石蜡切片对檫木花整体及组成部位进行观察,主要结果如下:檫木花由花被片6枚、雄蕊12枚和雌蕊1枚组成(图1-A)。花被片呈披针形,长约3 mm,分2轮排列,每轮3枚;12枚雄蕊分4轮排列,最外2轮每轮雄蕊3枚,由花药和花丝组成,长约3 mm;第3轮雄蕊3枚,在其基部着生2枚腺体,腺体成熟时呈杏黄色或橘红色,可吸引昆虫授粉,体式显微镜下观察到腺体外部呈马蹄形(图1-B),石蜡切片观察发现其含有大量油性细胞(图1-C);第4轮雄蕊3枚,外形呈倒三角形,围绕雌蕊排列,形态退化无育性且较前3轮体积小,长约1.5 mm(图1-D),试验中观察到的花多为该结构。
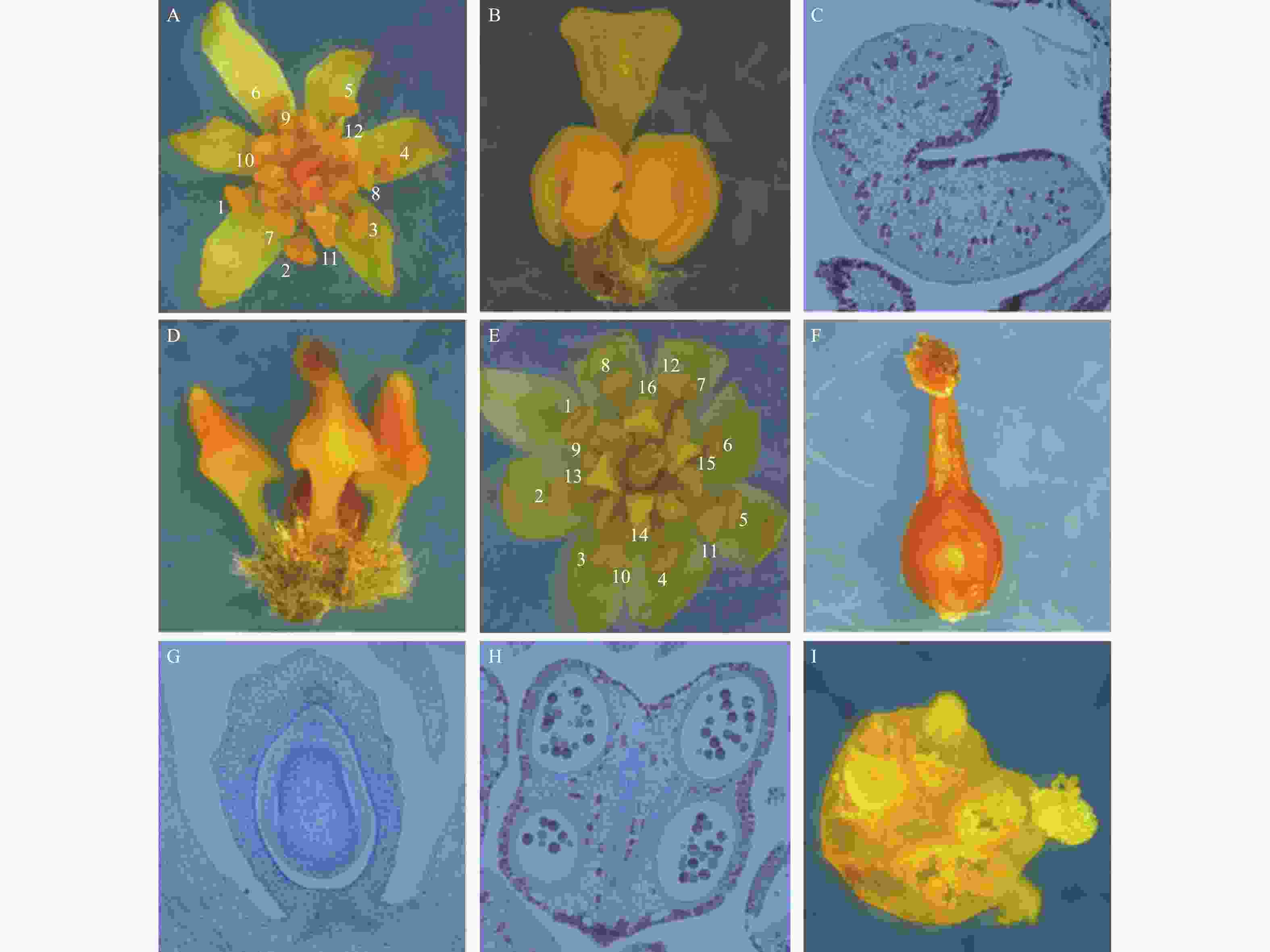
图 1 檫木花结构特征
Figure 1. The flower structure characteristics of S. tzumu
作者还观察到另一种檫木花结构类型,其由花被片8枚、雄蕊16枚和雌蕊1枚组成(图1-E),花被片8枚分2轮排列,16枚雄蕊分4轮排列,具体表现最外2轮雄蕊每轮4枚,第3轮雄蕊4枚,第4轮雄蕊4枚,其中前3轮雄蕊可育,第4轮雄蕊退化;檫木花雌蕊位于整个花的中央,由柱头、花柱和子房3部分组成,柱头具有乳头状突起(图1-F),子房上位,包含1室1胚珠(图1-G);可育雄蕊花药4室(图1-H),花药囊朝向雌蕊,授粉时花粉囊破裂花粉外散(图1-I),雄配子体经花柱进入胚囊与卵结合并最终完成授粉。
-
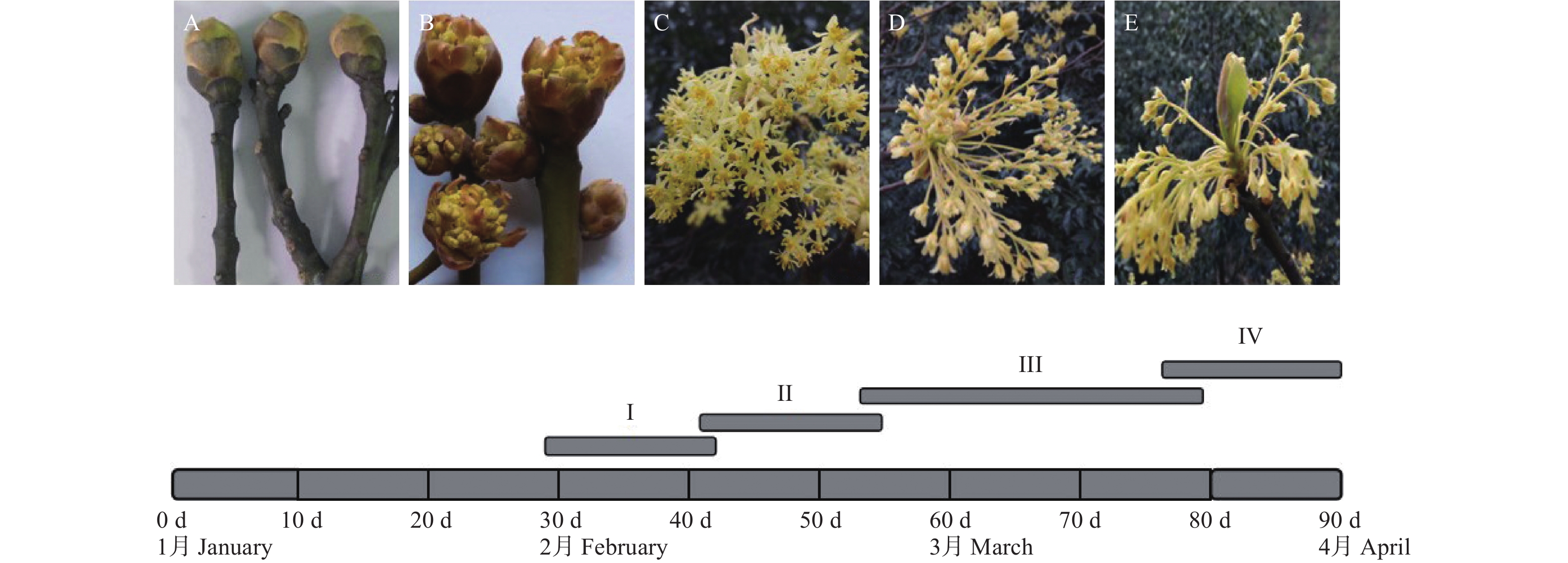
檫木开花过程可分为4个阶段,分别为I-芽鳞开裂期、Ⅱ-初花期、Ⅲ-盛花期、Ⅳ-末花期,整个过程持续约60 d。檫木成花的芽位于枝条顶端(图2-A),是由簇生的花序与新枝生长锥组成的混合芽,芽外部覆盖数目不等的黄褐色芽鳞,其大小与所包含的花序数目有关。

图 2 檫木开花动态
Figure 2. Floral dynamic of S. tzumu
混合芽于1月底至2月初体积达到最大后芽鳞开裂,簇生状总状花序裸露(图2-B),之后花序不断伸长,视气温条件及发育情况逐渐开花。2月下旬至3月中旬为盛花期(图2-C),每个混合芽平均可形成花序4.75~9.50个,每个花序平均成花8.25~14.50朵,每个混合芽可成花约68~92朵,且每个混合芽成花量在同株不同混合芽间差异大,平均花径8.76~11.01 mm,花梗长6.95~7.65 mm(表1)。3月中旬开始授粉,该时期腺体呈杏黄色或橘红色,并分泌香味物质,花粉囊破裂,花粉外散,吸引昆虫授粉,完成授粉后花被片闭合、柱头萎蔫(图2-D)。3月下旬进入末花期,该时期子房开始膨大,之后花被片及雄蕊逐渐脱落并形成幼果,位于花序中央的新枝生长锥开始生长(图2-E)。
表 1 檫木花部结构数量特征
Table 1. The quantitative characteristic of flower structure of S. tzumu
编号
Number每芽花序数
Number of inflorescence每花序花数
Number of flowers per inflorescence每芽总花量
Total number of flowers花梗长
Length of peduncle /mm花径
Flower diameter/mmCM1 4.75 ± 0.50 14.50 ± 0.42 68.75 ± 9.25 6.95 ± 1.68 8.76 ± 1.01 CM2 7.40 ± 2.30 10.93 ± 1.00 82.20 ± 28.67 7.42 ± 2.26 9.62 ± 0.76 CM3 8.20 ± 1.30 11.20 ± 0.45 92.20 ± 15.29 6.97 ± 1.78 9.35 ± 0.71 CM4 7.00 ± 1.09 12.83 ± 1.17 89.17 ± 14.37 7.34 ± 1.52 9.67 ± 0.70 CM5 7.14 ± 0.89 9.29 ± 2.49 66.86 ± 23.52 7.65 ± 2.07 8.83 ± 0.67 CM6 9.50 ± 1.73 8.25 ± 0.50 77.75 ± 15.35 6.75 ± 1.68 11.01 ± 0.83 -
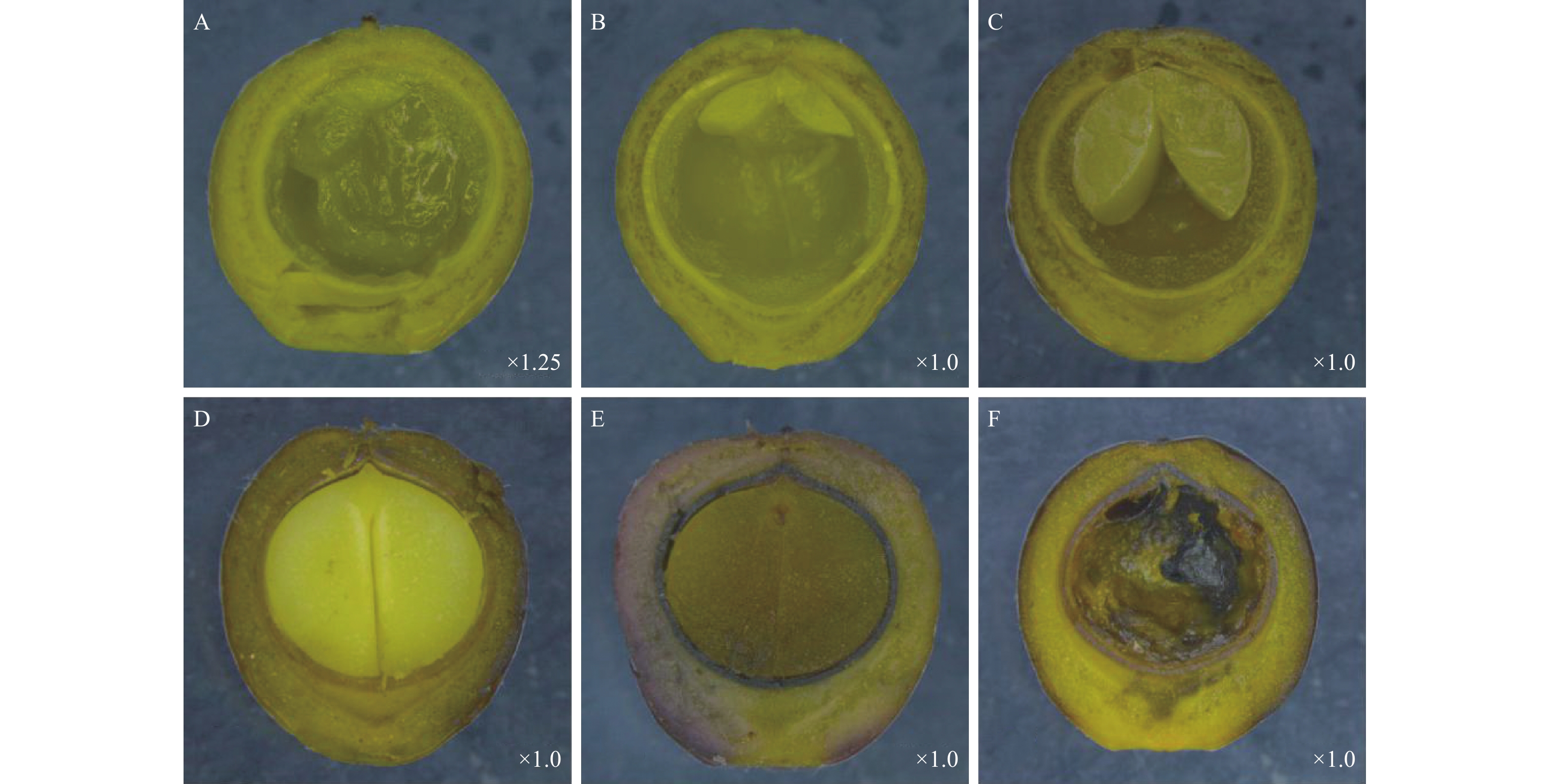
檫木果实由果皮和种子两部分组成,按照果实发育特征,可将果实发育分为幼果期、果实膨大期、果实变色期和果实成熟期。檫木花3月中下旬完成授粉后子房膨大,4月初可观察到幼果,幼果果皮呈绿色,解剖可见内部物质呈白色絮状,种皮呈白色,果皮厚度约0.22 mm(图3-A)。5月初解剖新鲜果实(第5周),可观察到对称的两片子叶平伏于胚乳上,两片子叶中间为胚芽,位于果腔上方(图3-B),之后子叶吸收胚乳营养不断生长,体积逐步增大(图3-C);至6月中旬,胚乳被子叶完全吸收,2片子叶充满整个果腔(图3-D),此时种皮已由白色变为黄色,生长1~2周后种皮硬化并由黄色变为黑色,果皮厚度约为0.93 mm,并开始由绿色向黑色转变,果皮细胞介质逐渐软化并最终呈浆状,至果皮成紫黑色时果实完全成熟(图3-E),整个发育过程持续12周,最终形成浆果状核果,种子黑色呈圆形,直径约5.12 mm,为双子叶无胚乳种子。此外,试验还观察到8.53%~27.22%不等的非正常发育果实,该类果实体积与正常果实无差异,具有正常发育的果皮和种皮,但未观察到种胚(图3-F)。
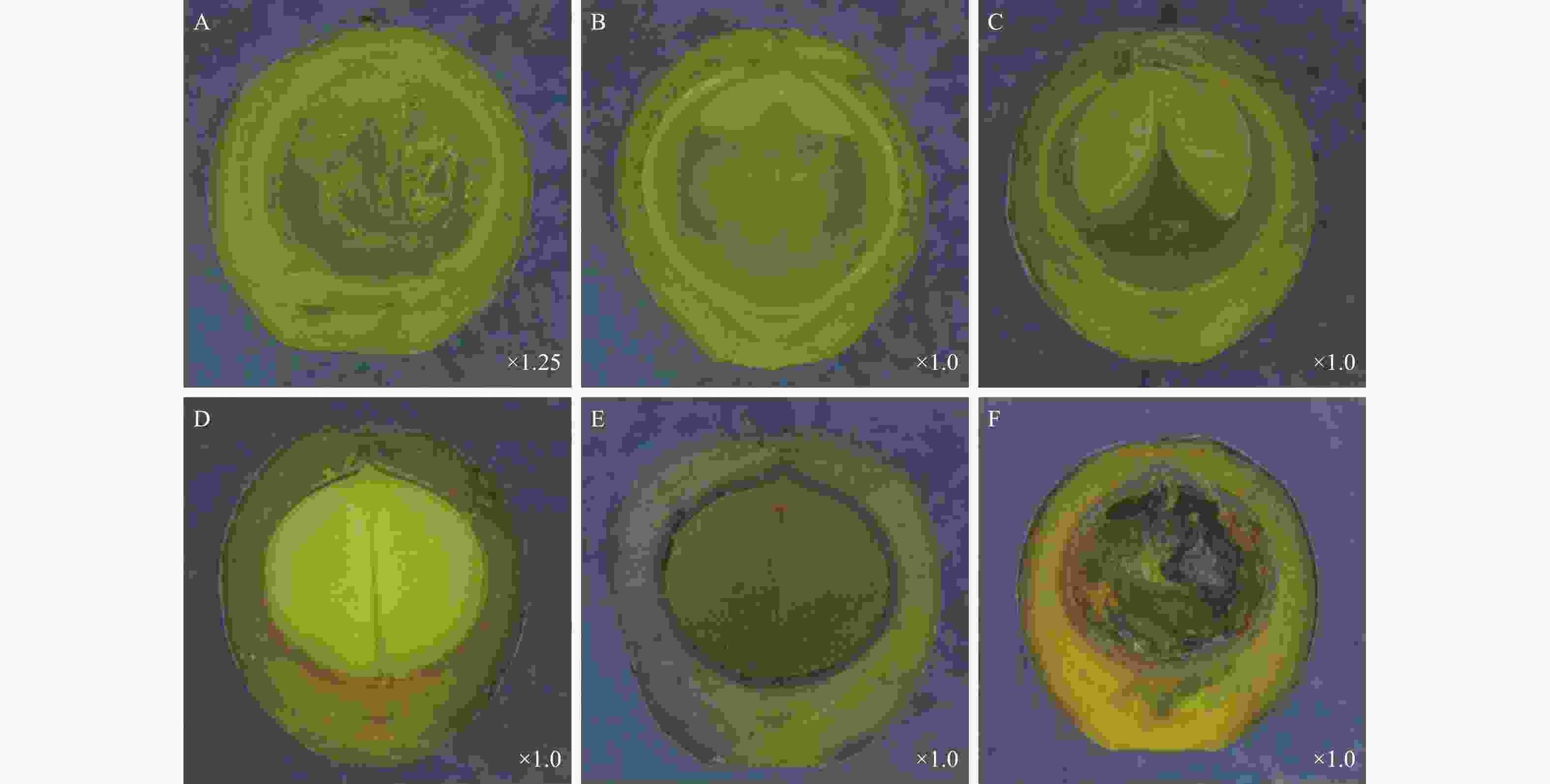
图 3 不同发育时期檫木果实结构特征
Figure 3. Fruit structure characteristics of S. tzumu at different developmental stages
-
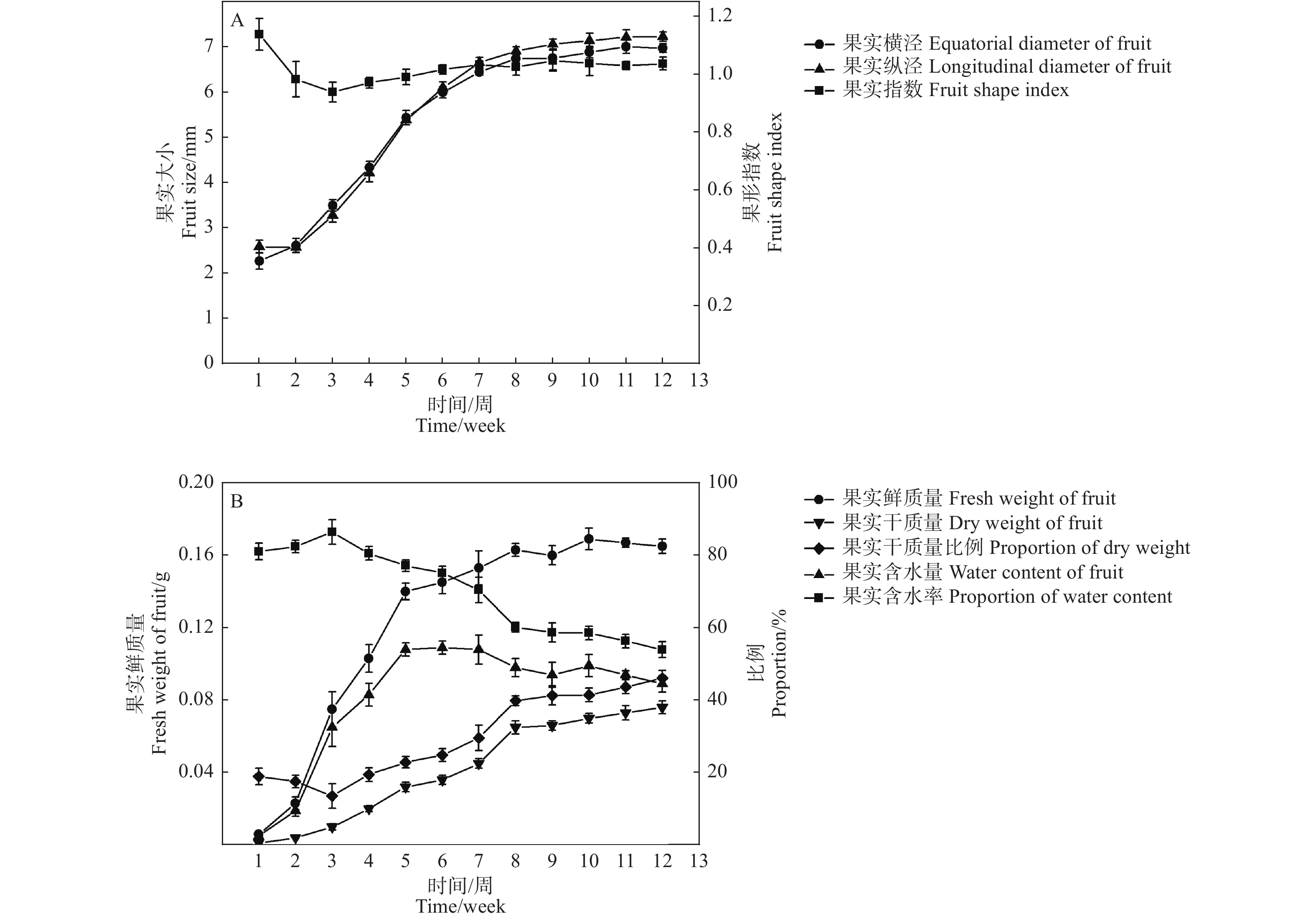
在檫木果实发育初期(第1—2周),果实鲜质量和体积缓慢增加,果实纵径大于果实横径,第3—8周果实横径与果实纵径快速增加,8周后果实生长速度降低,果实体积缓慢增加,至第10周果实体积基本达到最大,此时果实横径与纵径分别为6.89 mm和7.14 mm,果形指数1.04(图4-A),果实外形呈圆形;第3—5周果实鲜质量快速增加,之后生长速度减缓,至第10周果实鲜质量达到最大,单果质量0.17 g,在整个发育过程中果实体积和鲜质量均表现为“慢-快-慢”的变化规律。
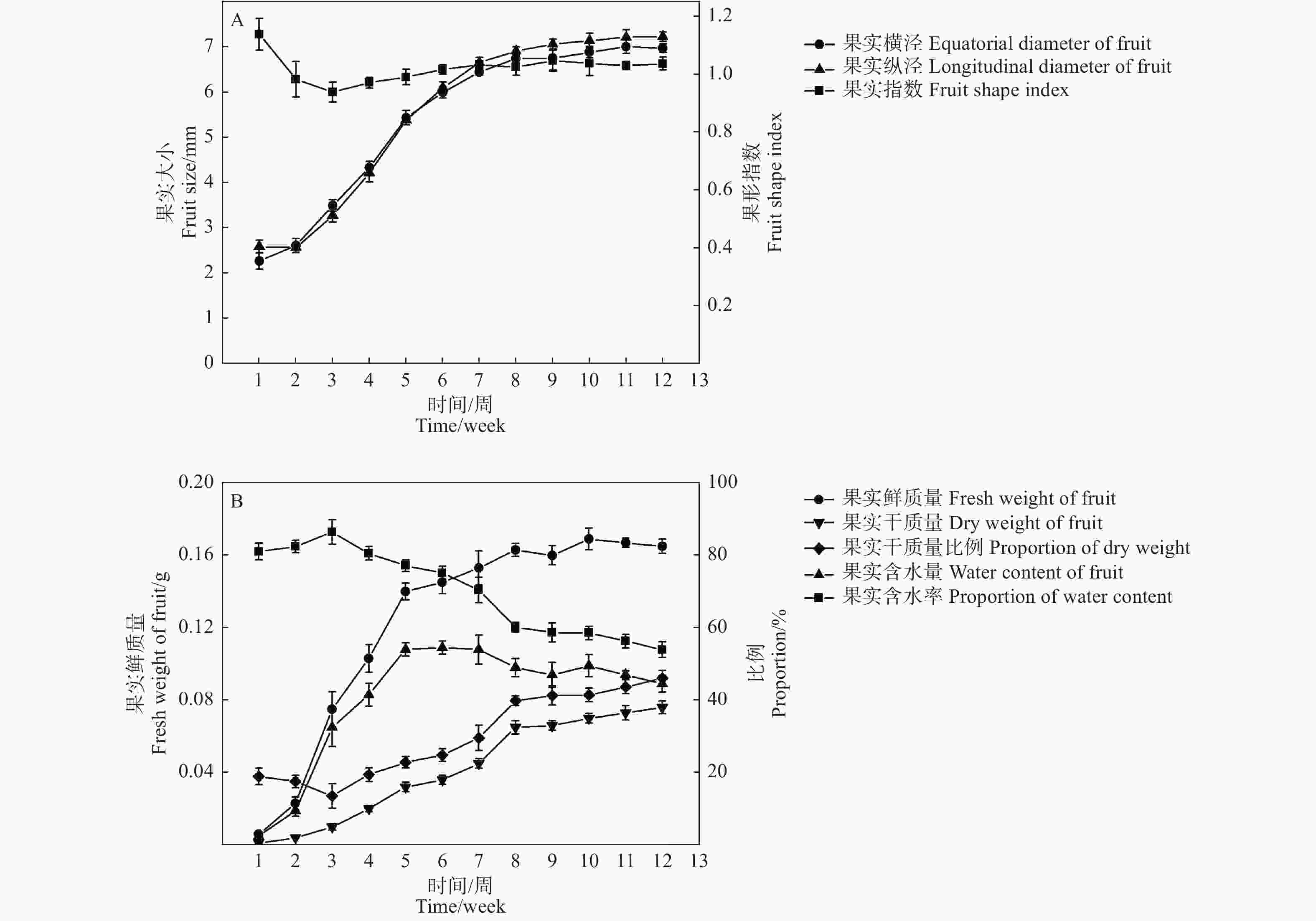
图 4 檫木果实大小与质量变化规律
Figure 4. Changes of fruit size and weight of S. tzumu
檫木果实发育早期含水率达80%以上(第1—3周),果实干质量低;随着种胚的形成与生长,果实含水率降低,果实干质量呈线性增加,果实含水量则随着果实生长先升高后降低的变化特征,其中第5周时含水量最高为0.11 g,占该时期果实鲜质量的77.15%,至果实成熟时含水量降为0.09 g,占果实鲜质量的53.93%,单果干质量0.08 g,占果实鲜质量的46.07%(图4-B)。以果实发育周期为横坐标(x),分别以果实鲜质量、果实含水量、果实干质量、果实横径和果实纵径为纵坐标(y)绘制散点图,数学模型拟合显示相关指标均与果实发育周期存在一定的函数关系,其R2值均达到0.900以上(表2),表明模型可以较准确地反映檫木果实生长发育的动态变化规律。
表 2 檫木果实性状与果实发育周期函数关系
Table 2. The relationship between fruit characteristics and develpment periods
果实性状
Fruit characteristics函数关系式
Functional relationR2 果实鲜质量
Fresh weight of fruity = 0.074 4ln(x) − 0.001 7 0.943 果实含水量
Water content of fruity = −0.002 1x2 + 0.033 9x − 0.025 5 0.909 果实干质量
Dry weight of fruity = 0.007 7x − 0.008 5 0.967 果实横径
Transverse diameter of fruity = 2.279 0ln(x) + 1.623 3 0.946 果实纵径
Transverse diameter of fruity = 2.360 7ln(x) + 1.601 9 0.914 -
明确植物开花规律和结构特征是研究植物生活史、了解植物传粉机制、繁殖系统以及资源收集利用等工作的前提[16-17]。檫木成花的芽1月底2月初芽鳞开裂,群体水平上花期自2月中旬至3月下旬,持续约45 d;在花结构方面,不同于美洲檫木单性花、雄花雄蕊9枚、无退化雄蕊和退化雌蕊、雌花具6枚退化雄蕊的分类特征[18],东亚分布的姐妹种——台湾檫木和檫木则以花具有正常雌雄蕊结构以及内轮具3枚形态退化雄蕊为主要特征,以至于曾将东亚种作为不同属或亚属[2, 9]。但关于东亚2个种的花性别长期存在争议,研究表明台湾檫木花药2~4室,花为雌蕊先熟的雌雄异熟两性结构,应称为同步或异步雌雄异熟[11, 19],檫木具正常两性花结构,雄蕊花药4室,雌蕊先于雄蕊成熟[12-13]。调查发现檫木开花量大,每混合芽可成花68~92朵,主要依赖昆虫授粉,观察到的花均具有雄蕊和雌蕊结构,表现出两性花的特征,试验中未发现单性花,可育雄蕊花药4室,且供试檫木植株均能正常结实;试验观察到2种不同花结构类型,一种由花被片6枚、雄蕊12枚和雌蕊1枚组成,这与多数文献中所描述的花结构特征一致[2, 12],另一种由花被片8枚、雄蕊16枚和雌蕊1枚组成,但该类型花数量较少,关于该类型的花能否正常授粉结实,以及两种类型的花是否同时存在于同一花序上等问题有待进一步观察。
檫木果实生长呈现“慢-快-慢”的变化规律,4月初幼果形成,此时新枝生长锥开始生长,与幼果生长形成竞争,造成檫木果实初期生长缓慢[12];4月中下旬檫木叶发育完成并开始提供光合产物,果实进入快速生长期,果实鲜质量快速增加,种胚体积增大,果实含水率降低,果实干质量增加;至6月底进入成熟期,该时期细胞介质成浆汁状,形成浆果状核果,果托浅杯状具长柄,果皮紫黑色被白粉,果核近球形,种子为双子叶无胚乳种子[12, 20]。试验观察到檫木果实中存在8.53%~27.22%的无种胚果实,其可能与胚囊仅少数正常发育、雌配子体很少出现数目齐全的胚囊核以及珠心组织存在解离等现象有关[19, 21],另一方面,檫木依赖昆虫传粉,花雌蕊先于雄蕊熟,且柱头可授期仅为8 d,由于檫木花中存在花粉败育现象且花粉活力仅能维持32 h[21],若遇早春严寒霜冻或授粉期阴雨天气等因素,极易造成檫木授粉失败[10-11],进而导致檫木虽然开花量大,但结实率低甚至不结实现象的产生。
-
檫木开花与果实发育过程观察结果表明:供试檫木植株群体水平上花期自2月中旬持续至3月下旬,开花过程可分为芽鳞开裂期、初花期、盛花期、末花期4个阶段;观察到少见的花被片8枚、雄蕊16枚及雌蕊1枚的花结构;檫木花具有正常雌雄蕊结构,能够正常授粉并结果,进一步支持檫木花为两性花的观点;檫木果实发育可分为幼果期、果实膨大期、果实变色期以及果实成熟期4个阶段,发育过程持续12周,果实为浆果状核果,种子为双子叶无胚乳种子。研究结果进一步深化了对檫木生物学特性的认知,并为檫木种质资源收集、保存利用及育种等研究提供了参考依据。
檫木开花与果实发育规律初步研究
A Study on Flowering and Fruit Development of Sassafras tzumu
-
摘要:
目的 观察檫木开花与果实发育过程,为檫木资源收集利用等研究提供参考。 方法 以杭州市富阳区城市森林公园内树龄超过10 a的檫木为调查对象,于2018年1—7月对其开花习性与果实发育过程进行观测,利用石蜡切片和体视显微镜观察花和果实结构。 结果 (1)檫木成花的芽是由簇生花序和新枝生长锥构成的混合芽,自混合芽芽鳞开裂至花期结束约60 d。(2)观察到2种结构类型的花,分别由花被片6枚、雄蕊12枚、雌蕊1枚和花被片8枚、雄蕊16枚、雌蕊1枚组成,前者具退化雄蕊3枚,后者具退化雄蕊4枚。(3)檫木果实发育呈现“慢-快-慢”的变化规律,随着果实生长,果实含水率下降,而果实干质量呈线性增加,5月初种胚开始发育,至6月下旬果皮呈紫黑色时果实成熟。 结论 檫木开花过程可以分为芽鳞开裂期、初花期、盛花期、末花期4个阶段,群体水平上花期约45 d,且檫木花能够正常授粉结实;檫木果实发育12周,可分为幼果期、果实膨大期、果皮变色期和果实成熟期,果实为浆果状核果,种子为双子叶无胚乳种子。 Abstract:Objective To study the flowering and fruit development of Sassafras tzumu so as to further understand the biological characters of S. tzumu and provide references for its collection and utilization. Method Taking the S. tzumu trees with the age over 10-year-old in city forest park of Fuyang District, Hangzhou City as trial materials, the flowering habits and fruit growth process were observed from January to July 2018, and the flower and fruit structure of the trees were observed by paraffin section and stereomicroscope. Result (1) The mixed buds of S. tzumu were composed of raceme and new branch growth cones. The time form buds scales break to the end of flowering was about 60 days. (2) Two types of flower were observed, one type had 6 perianth segments, 12 pistils, 1 stamen and the other one had 8 perianth segments, 16 pistils, and 1 stamen. The former one had 3 staminodes and the latter one had 4 staminodes. (3) The growth of fruit followed a "slow -fast -slow" curve. The water content of the fruits decreased while the dry matter accumulation linearly increased with the growth of the fruits. The embryo began to grow in early May, and the fruit was fully mature when the skin was purple black in the end of June. Conclusion The flowering includes buds scales break, early florescence, blossom period, and final one. The flowering period is about 45 days at the population level, and can be pollinated and fruited normally. The growth of fruit can be divided into young fruit, expanding stage, peel discoloration and maturity, which lasts for 12 weeks. The fruits are berrylike drupe and dicotyledonous seeds without endosperm. -
Key words:
- Sassafras tzumu
- / flower
- / fruit development
-
图 3 不同发育时期檫木果实结构特征
Figure 3. Fruit structure characteristics of S. tzumu at different developmental stages
表 1 檫木花部结构数量特征
Table 1. The quantitative characteristic of flower structure of S. tzumu
编号
Number每芽花序数
Number of inflorescence每花序花数
Number of flowers per inflorescence每芽总花量
Total number of flowers花梗长
Length of peduncle /mm花径
Flower diameter/mmCM1 4.75 ± 0.50 14.50 ± 0.42 68.75 ± 9.25 6.95 ± 1.68 8.76 ± 1.01 CM2 7.40 ± 2.30 10.93 ± 1.00 82.20 ± 28.67 7.42 ± 2.26 9.62 ± 0.76 CM3 8.20 ± 1.30 11.20 ± 0.45 92.20 ± 15.29 6.97 ± 1.78 9.35 ± 0.71 CM4 7.00 ± 1.09 12.83 ± 1.17 89.17 ± 14.37 7.34 ± 1.52 9.67 ± 0.70 CM5 7.14 ± 0.89 9.29 ± 2.49 66.86 ± 23.52 7.65 ± 2.07 8.83 ± 0.67 CM6 9.50 ± 1.73 8.25 ± 0.50 77.75 ± 15.35 6.75 ± 1.68 11.01 ± 0.83  下载: 导出CSV
下载: 导出CSV
表 2 檫木果实性状与果实发育周期函数关系
Table 2. The relationship between fruit characteristics and develpment periods
果实性状
Fruit characteristics函数关系式
Functional relationR2 果实鲜质量
Fresh weight of fruity = 0.074 4ln(x) − 0.001 7 0.943 果实含水量
Water content of fruity = −0.002 1x2 + 0.033 9x − 0.025 5 0.909 果实干质量
Dry weight of fruity = 0.007 7x − 0.008 5 0.967 果实横径
Transverse diameter of fruity = 2.279 0ln(x) + 1.623 3 0.946 果实纵径
Transverse diameter of fruity = 2.360 7ln(x) + 1.601 9 0.914
下载: 导出CSV
-
[1] 中国科学院中国植物志编辑委员会. 中国植物志: 第三十一卷 樟科[M]. 北京: 科学出版社, 1982: 238. [2] Wu Z Y, Raven P H. Flora of China: Vol. 7 Lauraceae[M]. Beijing: Science Press, 2008: 159-161. [3] 蒋艾平, 姜景民, 刘 军. 天目山不同海拔檫木群体遗传多样性和遗传结构[J]. 应用生态学报, 2016, 27(6):1829-1836. [4] Li Y J, Sun Y, Jiang J M, et al. Spectroscopic determination of leaf chlorophyll content and color for genetic selection on Sassafras tzumu[J]. Plant Methods, 2019, 15: 73. doi: 10.1186/s13007-019-0458-0 [5] 中国森林编委会. 中国森林: 第三卷 阔叶林[M]. 北京: 中国林业出版社, 1999: 1429-1430. [6] 中国树木志编委会. 中国主要树种造林技术[M]. 北京: 中国林业出版社, 1983: 545-551. [7] 孙洪刚, 姜景民, 万志兵. 海拔和坡向对北亚热带檫木天然次生林生长、空间结构和树种组成的影响[J]. 东北林业大学学报, 2017, 45(4):8-13. [8] 李 捷, 李锡文. 世界樟科植物系统学研究进展[J]. 植物分类与资源学报, 2004, 26(1):1-11. [9] Nie Z L, Wen J, Sun H. Phylogeny and biogeography of Sassafras (Lauraceae) disjunct between eastern Asia and eastern North America[J]. Plant Systematics and Evolution, 2007, 267(1): 191-203. [10] 沈卓群, 盛 哲, 毛玉先, 等. 檫树开花结实生物学特性的研究初报[J]. 湖南农学院学报, 1980,(3):13-15. [11] Chung K F, Werff H V D, Peng C I. Observations on the floral morphology of Sassafras randaiense (Lauraceae)[J]. Missouri Botanical Garden, 2010, 97(1): 1-10. doi: 10.3417/2008029 [12] 谷淑芳, 陈建国, 韩 红, 等. 檫木花果形态构造及其发育初步观察[J]. 浙江林业科技, 1991, 11(4):16-22. [13] 王 馨, 于 芬, 季春峰. 檫木花粉发育过程的解剖学研究[J]. 甘肃农业大学学报, 2014, 49(2):116-119. [14] 潘丽琴, 郝 建, 徐建民, 等. 灰木莲花期物侯观测及生殖构件分布[J]. 林业科学研究, 2018, 31(2):96-101. [15] 王 馨, 于 芬, 季春峰, 等. 樟科檫木的大孢子发生和雌配子体发育[J]. 江西农业大学学报, 2016, 38(1):42-47. [16] 刁松锋, 姜景民, 伊 焕, 等. 浙江低山地区多用途植物无患子的开花物候特征[J]. 生态学报, 2016, 36(19):6226-6234. [17] Lambert A M, Millerrushing A J, Inouye D W. Changes in snowmelt date and summer precipitation affect the flowering phenology of Erythronium grandiflorum (glacier lily; Liliaceae)[J]. American Journal of Botany, 2010, 97(9): 1431-1437. doi: 10.3732/ajb.1000095 [18] Flora of North America Editorial Committee. Flora of North America: North of Mexico, Vol. 3: Magnoliophyta: Magnoliidae and Hamamelidae[M]. New York: Oxford University Press, 1997: 26-36. [19] 蔡永信. 台湾檫树开花行为与微卫星体基因坐特性分析[D]. 台北: 台湾大学森林环境与资源学研究所, 2011: 1-5. [20] 国家林业局国有林场和林木种苗工作总站. 中国木本植物种子[M]. 北京: 中国林业出版社, 2000: 211-213. [21] 王 馨, 于 芬, 季春峰, 等. 檫木花粉活力和柱头可授性研究[J]. 江西林业科技, 2013(5):5-6. -

 点击查看大图
点击查看大图
图(4) / 表(2)
计量
- 文章访问数: 5003
- HTML全文浏览量: 2809
- PDF下载量: 72
- 被引次数: 0
