




-
崖柏属(Thuja L.)为柏科(Cupressaceae)的一个小属群,属下仅有5个现存种,间断分布在东亚和北美东西部。东亚3个种分布范围均较狭窄,如崖柏(T.sutchuenensis Franch)为我国特有的极度濒危物种,仅见于渝东北的石灰岩山地(城口县和开县交界处);朝鲜崖柏(T. koraiensis Nakai)分布于我国长白山地区和朝鲜半岛,日本香柏(T. standishii L.)则为日本所特有。北美2个种分布较广,北美乔柏(T. plicata D. Don)分布可北至亚北极泰加林,南至落叶林区;北美香柏(T. occidentalis L.)分布于北美西部太平洋海岸山地与落基山脉[1]。
崖柏属植物的细胞学研究国内外已有一些报道[2-3],本文对属内5种植物(崖柏、朝鲜崖柏、日本香柏、北美乔柏和北美香柏)进行了核型比较分析,其中,崖柏和朝鲜崖柏的核型均为首次报道,旨在为崖柏属的系统发育研究及资源利用提供细胞学资料。
HTML
-
研究材料(崖柏、朝鲜崖柏、日本香柏、北美乔柏和北美香柏)取自中国林业科学研究院科研温室的2年生实生苗,种子依次来源于大巴山国家级自然保护区、长白山国家级自然保护区、日本本州、美国蒙太拿州和美国缅因州。取5个树苗的幼嫩根尖(约1 cm)置于0.002 mol·L-1 8-羟基喹啉和0.05%秋水仙素(1∶1)的混合溶液中,20℃下预处理2 h,然后用卡诺固定液(冰醋酸∶纯酒精=1∶3)固定备用。制片前用1 mol·L-1的盐酸溶液在60℃水浴锅中软化12 min,水洗3~4次。取根尖的分生区部分置载玻片上,用小镊子涂碎,滴少许PICCH(丙酸-铁-水合三氯乙醛-苏木精)染色液染色,盖上盖玻片,用吸水纸吸干,压平。Olympus BH-2光学显微镜10×100倍数下观察染色体分裂中期状况,选择染色体分裂较好的照相,每个材料至少观察10个细胞,选择染色体分散较好的照相。每个种核型的平均值为5个细胞各项指标的平均值。
核型分析的项目、标准和方法参照李懋学等[4]的核型分析标准。臂比(AR)=按长臂/短臂;染色体长度比(LR)=最长染色体长度/最短染色体长度;核型不对称系数(As.K.C)=(长臂总长/全组染色体总长)×100%;染色体总形态百分比(TM)=(短臂总长/全组染色体总长)×100%;染色体相对长度指数(I.R.L)=染色体长度/全组染色体平均长度。
-
分别从崖柏、朝鲜崖柏、日本香柏、北美乔柏和北美香柏的5个细胞分裂相确定体细胞染色体数目,5个树苗的体细胞染色体数目均为2n=22,染色体基数x=11,未见染色体非整倍性变异和加倍现象。核型特征和数据结果见图 1和表 1。

Figure 1. Chromosome morphology in species of genus Thuja L.
植物
Species核型公式
Formula of karyotypes染色体相对长度指数I.R, L 染色体长度比LR 染色体总形态百分比TM/% 核型不对称系数As.K.C/% 崖柏T. sutchuenensi 2n=2x=22=18m (2SAT)+4sm 2L+8M2+12M1 1.76 43.22 56.78 朝鲜崖柏T. koraiensis 2n=2x=22=20m(2SAT)+2sm 2L+8M2+12M1 1.68 43.92 56.08 日本香柏T. standishii 2n=2x=22=18m (2SAT)+4sm 2L+8M2+12M1 1.75 43.42 56.58 北美乔柏T. plicata 2n=2x=22=20m(2SAT)+2sm 2L+8M2+12M1 1.64 43.38 56.62 北美香柏T.occidentalis 2n=2x=22=20m(2SAT)+2sm 2L+6M2+14M1 1.60 43.81 56.19 Table 1. The karyotype data of five species of genus Thuja L.
崖柏的根尖染色体数目为22,核型公式为2n=2x=22=18m (2SAT)+4sm,有9对中部着丝粒染色体和2对近中部着丝粒染色体,只有1对随体位于第11对染色体上(表 1、2,图 1A)。染色体相对长度为6.95~12.23,染色体长度比1.76,平均臂比为1.36。不对称系数为56.78,核型类型为1A。
染色体 崖柏T. sutchuenensis 朝鲜崖柏T. koraiensis 日本香柏T. standishii 北美乔柏T. plicata 北美香柏T.occidentalis 相对长度AL/% 臂比AR 着丝点位置PC 相对长度AL/% 臂比AR 着丝点位置PC 相对长度AL/% 臂比AR 着丝点位置PC 相对长度AL/% 臂比AR 着丝点位置PC 相对长度AL/% 臂比AR 着丝点位置PC 1 12.23 1.04 m 11.99 1.09 m 12.05 1.16 m 12.03 1.12 m 11.85 1.14 m 2 11.09 1.32 m 11.30 1.11 m 11.12 1.08 m 11.31 1.21 m 11.31 1.24 m 3 10.81 1.28 m 10.45 1.12 m 10.56 1.08 m 10.31 1.07 m 10.84 1.07 m 4 10.23 1.40 m 9.70 1.40 m 9.59 1.79 sm 9.47 1.42 m 9.24 1.53 m 5 9.22 1.11 m 9.35 1.32 m 9.21 1.07 m 9.26 1.25 m 9.03 1.34 m 6 8.79 1.04 m 8.91 1.20 m 8.86 1.15 m 8.91 1.52 m 8.58 1.08 m 7 8.14 1.61 m 8.08 1.46 m 8.33 1.47 m 8.36 1.14 m 8.37 1.53 m 8 7.76 1.79 sm 7.89 1.58 m 8.15 1.45 m 7.95 1.53 m 8.08 1.47 m 9 7.54 1.47 m 7.74 1.11 m* 7.74 1.71 sm 7.71 1.67 m 7.76 1.54 m 10 7.25 1.83 sm 7.46 1.34 m 7.52 1.49 m 7.34 1.09 m* 7.51 1.72 sm 11 6.95 1.12 m* 7.13 1.76 sm 6.87 1.10 m* 7.34 1.84 sm 7.42 1.08 m* 注:*次缢痕, Note: secondary constriction. Table 2. The chromosome parameter of five species of genus Thuja L.
朝鲜崖柏的根尖染色体数目为22,核型公式为2n=2x=22=20m(2SAT)+2sm,有10对中部着丝粒染色体和1对近中部着丝粒染色体,只有1对随体位于第9对染色体上(表 1、2,图 1B)。染色体相对长度为7.13~11.99,染色体长度比1.68,平均臂比为1.32。不对称系数为56.08,核型类型为1A。
日本香柏的根尖染色体数目为22,核型公式为2n=2x=22=18m (2SAT)+4sm,有9对中部着丝粒染色体和2对近中部着丝粒染色体,只有1对随体位于第11对染色体上(表 1、2,图 1C)。染色体相对长度为6.87~12.05,染色体长度比为1.75,平均臂比为1.32。不对称系数为56.58,核型类型为1A。
北美乔柏的根尖染色体数目为22,核型公式为2n=2x=22=20m(2SAT)+2sm,有10对中部着丝粒染色体和1对近中部着丝粒染色体,只有1对随体位于第10对染色体上(表 1、2,图 1D)。染色体相对长度为7.34~12.03,染色体长度比1.64,平均臂比为1.35。不对称系数为56.62,核型类型为1A。
北美香柏的根尖染色体数目为22,核型公式为2n=2x=22=20m(2SAT)+2sm,有10对中部着丝粒染色体和1对近中部着丝粒染色体,只有1对随体位于第11对染色体上(表 1、2,图 1E)。染色体相对长度为7.42~11.85,染色体长度比1.60,平均臂比为1.34。不对称系数为56.19,核型类型为1A。
结果显示:崖柏和日本香柏有9对中部着丝粒染色体和2对近中部着丝粒染色体,朝鲜崖柏、北美乔柏和北美香柏有10对中部着丝粒染色体和1对近中部着丝粒染色体,近中部着丝粒的数目不是完全相同,但平均臂比与染色体长度比比较接近,均没有臂比大于2的染色体。因此,按照Stebbins[5]的核型分类标准,崖柏属5种植物的核型均属于1A类型,这是一种最为整齐的对称核型。对崖柏属5种植物的染色体相对长度指数按Kuo等[6]提出的分类方法进行分类发现,染色体相对长度组成中除M1、M2染色体外都有1对L染色体;且均只具有一对随体,但不同种次缢痕的位置有所不同,如崖柏位于第11对染色体上,而朝鲜崖柏位于第9对染色体上。
-
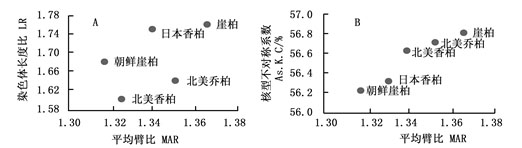
平均臂比(MAR)和染色体长度比(LR)可以表示不同种间核型的不对称性,二者愈大,其核型愈不对称。从MAR和LR的二维进化趋势图(图 2A)发现,崖柏和日本香柏沿MAR和LR方向进化较快,较为进化;而朝鲜崖柏和北美香柏则较为原始。

Figure 2. Evolution trend map of karyotype in species of genus Thuja L. (A: Two-dimensional representation of evolutionary trend of MAR and LR; B: Two-dimensional representation of evolutionary trend of MAR and As.K.C)
平均臂比(MAR)和核型不对称系数(As.K.C)则从另一个角度反映了核型的不对称性,二者愈大,核型愈不对称。从二维进化图(图 2B)可以看出:崖柏属5种植物均按MAR和As.K.C向右上角方向进化,崖柏MAR及As.K.C均最大,表明其在崖柏属中分化最晚;相比之下,朝鲜崖柏则较为原始。
2.1. 染色体及核型分析
2.2. 崖柏属植物的进化趋势分析
-
李林初等[2]曾报道北美乔柏和北美香柏的核型公式分别为2n=2x=22=16m+6sm(2SAT)和2n=2x=22=20m(2SAT)+2sm;Kuroki等[3]曾报道日本香柏的核型公式为2n=2x=22=16m+6sm。其中,北美乔柏和日本香柏的核型公式与本文报道的结果不尽相同,这可能是由于取材、实验方法的处理不同或测量误差所致。
染色体核型特征研究有助于理解植物属种间[7-9]或种内水平[10-11]的进化趋势。本文对崖柏属5种植物核型研究发现,核型中除大多为中部着丝粒染色体外,还有1~2对不等的近中部着丝粒染色体。前苏联核型比较形态学派的代表Levitzky[12]曾指出:所谓对称的核型就是其中所有染色体大小近乎一致,而且都具有中部和近中部着丝粒,可见崖柏属5种植物的核型均为对称的核型。核型不对称系数(As.K.C)越接近50,表明着丝粒越接近中点,染色体越对称;相对而言,As.K.C越接近0,染色体越不对称。臂比和染色体长度比同样可以反映核型的对称性,二者越接近1,核型越对称。根据Stebbins[13]的观点,在系统演化上处于较古老或原始地位的植物,往往具有较对称的核型,而不对称的核型则往往出现在较进化或特化的植物中。本研究结果发现,崖柏属5种植物的平均臂比为1.32~1.36,染色体长度比为1.60~1.76、As.K.C为56.08~56.78,与柏科的柏木属(Cupressus L.)[14]、扁柏属(Chamaecyparis Spach)[15]、侧柏属(Platycladus Spach)[16]相比都较大,表明该属具有较为不对称的核型,在柏科中可能处于比较进步的进化地位。这与李林初[3]对侧柏亚科10个属植物核型分析的结果一致,即侧柏属较为原始,而崖柏属较进化。
McIver等[17]根据化石记录、分布范围及形态特征,尤其是球果结构,对崖柏属现存种和化石种进行进化史分析,认为崖柏属植物在第三纪早期曾广泛分布于北半球,随后的剧烈气候变化和地质变迁是导致该属植物形成现有间断分布格局的主要原因,但对于该属植物的起源,尚存不同观点。崖柏的成熟球果为5~8 mm,较属内其它种(7~18 mm)小,而苞鳞数量为4对,也少于其他种(4~6对),而一度被认为是该属最古老的种[17-18]。Li等[19]利用核糖体DNA ITS序列分析推断该属为东亚起源,且划为两大分支,其一为北美乔柏和朝鲜崖柏,其他3种归为另一分支,现有分布格局是经过2次向北美的迁移而形成的,这2次迁移大致发生在渐新世晚期至中新世早期;但Peng等[20]利用5套叶绿体基因、2套低拷贝核基因片段和核基因ITS序列,辅以最新化石标本诊断的时间和地点作为佐证,推断崖柏属物种之间极有可能存在网状进化,认为该属可能起源于古新世或更早的北美高纬度地区,通过白令大陆桥扩散到东亚,后来又在中新世由东亚扩散至北美。本文通过二维进化趋势图分析发现,崖柏属植物呈现出东亚(朝鲜崖柏、日本香柏)和北美(北美乔柏、北美香柏)两大分支,崖柏处于最进化地位,但其起源尚需进一步研究。
-
本研究发现,崖柏和日本香柏的核型公式为2n=2x=22=18m(2SAT)+4sm,朝鲜崖柏、北美乔柏和北美香柏的核型公式为2n=2x=22=20m(2SAT)+2sm,5种植物的核型均属于1A类型。崖柏属处于柏科较进化的地位,而崖柏为崖柏属最后分化种。
 DownLoad:
DownLoad: