



-
油橄榄(Olea europaea L.)原产于地中海沿岸,为木犀科(Oleaceae)、木犀榄属(Olea L.)植物,是著名的木本油料兼果用树种,其果实含油率高,鲜榨油即可直接食用,并具有极高的营养价值。于20世纪60年代开始,我国大量引种油橄榄,并开展了栽培技术和品种选育等研究[1-2]。已有研究表明,多数油橄榄品种具有自交不亲和性或部分自交不亲和性,且品种间杂交亲和性也有一定的差异[3-9]。‘巴尼亚’(Barnea)是以色列油橄榄密植园主栽品种,与‘科拉蒂’(Coratina)和皮瓜尔’(Picual)杂交的2年平均座果率分别高达5.3%和7.3%,而与‘豆果’(Arbequina)杂交的平均座果率仅为0.22 %[9]。因此,合理配置授粉品种可以提高产量[7]。传统方法主要依据杂交授粉的座果率来进行授粉树的选配[5, 10-11],然而, 在授粉过程中不可避免的花粉污染会影响结果的准确性,而基于分子标记的父本分析则可解决这一问题。对已知母本的种子进行遗传标记分析而确定每一个子代(种子)的亲本,鉴定出大多数种子的花粉来源,推算出不同父本的贡献率,从而用于研究品种间亲和性,确定品种授粉组合[12]。由于SSR标记因具有多态性高和共显性的特点,被广泛应用于油橄榄子代的父本分析[7-8, 12-13]。
目前,国内基于人工授粉的座果率来研究油橄榄品种间亲和性,确定适宜的授粉树[10, 14-17],而利用父本分析研究油橄榄亲和性尚未见报道。在国外虽有相关报道,但研究结论存在一定差异,而且由于环境因素对研究油橄榄的亲和性有一定影响,如Gharibzadeh等[7]发现,‘克罗莱卡’(Koroneiki)在伊朗戈勒斯坦省是自交亲和的,而Mookerjee等[18]发现,‘克罗莱卡’在澳大利亚南部地区是自交不亲和的。此外,不同地区油橄榄主栽品种亦不同。因此,本文利用SSR标记对四川省西昌市油橄榄自由授粉子代进行父本分析,研究油橄榄品种间亲和性,确定适宜授粉品种,为凉山州油橄榄建园品种配置提供依据。
HTML
-
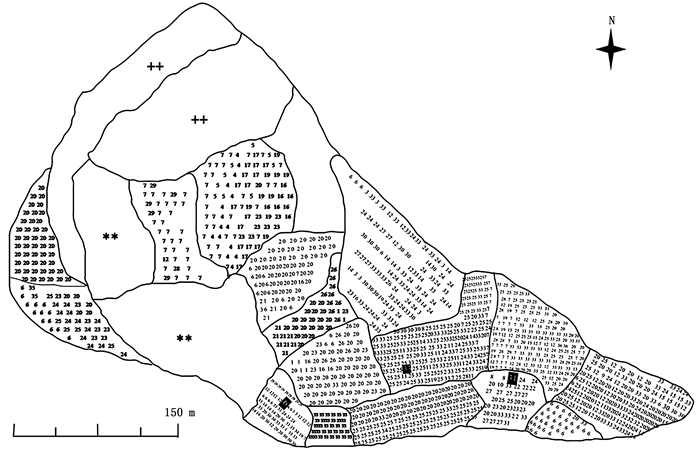
试验地处于四川省凉山彝族自治州西昌市中泽新技术开发有限公司北河基地(102°14′33″ E,27°44′40″N)。凉山州属于暖温带湿润气候区,干湿分明,冬半年日照充足,少雨干暖;夏半年云雨较多,气候凉爽。日温差大,年温差小,年均气温16~17℃。年日照时数约1 600~2 000 h,最高达2 400 h。于2007—2012年期间陆续以2年生扦插苗和嫁接苗定植建园,常规管理。园内开花品种共36个(表 1),均视为候选父本。这些品种生长表现良好,均已进入成熟期并盛花盛果。试验园为孤立园,品种配置见图 1,周围3 km范围内无开花的油橄榄树。选取‘豆果’、‘九峰4号’、‘鄂植8号’3个品种作为母树。
编号
No.品种Cultivar 来源Origin 用途
Use编号 品种Cultivar 来源Origin 用途
Use1 卡拉马达Kalamata 希腊Greece O/T 19 莱星Leccino 意大利Italy O 2 美佳伦Megaron 希腊Greece O/T 20 克罗莱卡Koroneiki 希腊Greece O 3 坦彩Tanche 法国France O/T 21 阿布桑娜Arbosana 西班牙Spain O 4 奥托卡Ottobratica 意大利Italy O 22 贝翠丽Patrim 希腊Greece O 5 沙龙奎Salonenqne 法国France O/T 23 巴尼亚Barnea 以色列Israel O/T 6 豆果Arbequina 西班牙Spain O 24 皮瓜尔Picual 西班牙Spain O 7 佛奥Frantoio 意大利Italy O 25 科拉蒂Coratina 意大利Italy O 8 切姆拉尔Chemlal de Kabylie 阿尔及利亚Albania O 26 小苹果Manzanilla de Sevilla 西班牙Spain O 9 长叶卡林Kaliniot Changye 阿尔巴尼亚Albania O/T 27 戈达尔Gordal 西班牙Spain O/T 10 圆果卡林Kaliniot Yuanguo 阿尔巴尼亚Albania O/T 28 配多灵Pendolino 意大利Italy O 11 尖叶卡林Kaliniot Jianye 阿尔巴尼亚Albania O/T 29 张林Zhanglin 国内选育Domesticated selection O/T 12 贝拉特Berat 阿尔巴尼亚Albania O/T 30 云台Yuntai 国内选育Domesticated selection O/T 13 哥郎米扎Mixaj Dukat 阿尔巴尼亚Albania O 31 九峰4号Jiufeng-4 国内选育Domesticated selection O/T 14 阿斯Ascolano Tenera 意大利Italy T 32 中山24Zhongshan-24 国内选育Domesticated selection O/T 15 白橄榄Ullir I bardhe 阿尔巴尼亚Albania O 33 鄂植8号Ezhi-8 国内选育Domesticated selection O 16 皮削利Picholine 法国France O/T 34 城固32Chenggu-32 国内选育Domesticated selection O/T 17 科新佛奥Frantoio de Corsini 意大利Italy O/T 35 巴尼亚实生苗Seedling of Barnea 国内选育Domesticated selection O 18 北庆米扎Mixaj Peginit 阿尔巴尼亚Albania O 36 未知1号Unknown-1 Unknown O 注:表中“O”为油用,“T”为餐用。‘O’ is oil olive and ‘T’ is table olive. Table 1. Name and origin country of olive cultivars

Figure 1. Field plan of the olive orchard showing the location of cultivars
于2014年3—4月对园内各品种花期进行调查,以开花量1%~25%为初花期,26%~75%为盛花期,76%~100%为末花期[10]。于2014年10月,采集园内所有品种的嫩叶,硅胶干燥保存,用于DNA提取,并采集3株母树自由授粉种子共334个(‘豆果’117个,‘九峰4号’103个,‘鄂植8号’114个),播种于中国林业科学研究院温室。2015年9月,采集所有子代播种苗嫩叶(共204株,其中‘九峰4号’80株,出苗率77.67%;‘豆果’64株,出苗率54.7%;‘鄂植8号’60株,出苗率52.63%),用于DNA提取。
-
采用改良CTAB法[19]提取亲本及子代嫩叶DNA,并于-20℃冰箱保存。
-
SSR-PCR反应体系为20 μL,包含150 ng样本DNA,1×PCR缓冲液(Tris-HCl 20 mmol·L-1 pH值8.0,KCl 100 mmol·L-1,MgCl2 1.5 mmol·L-1),1.5 mmol·L-1 Mg2+,0.225 mmol·L-1 dNTP,正反引物各0.2 μmol·L-1,Taq聚合酶1.25 U(TaKaRa)。PCR反应程序为:94℃ 5 min,35个循环(94℃30 s,Tm30 s,72℃30 s),72℃延伸7 min,4℃保温,不同引物的Tm值不同。SSR-PCR产物在8%的聚丙烯酰胺凝胶上电泳,并银染检测。
共筛选出多态性高的12对引物(DCA3、DCA11、DCA18、GAPU59、GAPU89、GAPU103A、GAPU71B、UDO99-6、UDO99-11、UDO99-19、EMO90和OLEAGEN-H6),并合成带有FAM和HEX荧光基团的荧光引物(上海捷瑞),用于SSR-PCR反应及毛细电泳检测。试验所用引物来自于相关文献[19-23]。
-
取FAM和HEX荧光标记扩增后的PCR产物2 μL,混合后加入6 μL ddH2O,混匀。取1 μL混合产物,加入9 μL Hi-Di和0.5 μL ROX-500荧光分子量内标混合均匀,95℃变性5 min,上样于ABI3730 DNA分析仪上,进行毛细管电泳和自动荧光检测,并收集数据[24]。
-
利用Gene MarkerV1.95软件分析收集数据,输出Allele和Bin数据。利用Cervus 3.0软件对204个自由授粉子代进行父本分析,并计算等位基因数(NA)、等位基因频率(Pi)、观测杂合度(Ho)、期望杂合度(He)和多态性信息量(PIC)。
2.1. 总DNA提取
2.2. SSR-PCR体系和引物筛选
2.3. 毛细电泳检测
2.4. 数据统计与分析
-
园内36个品种花期观测结果见表 2。所有品种花期持续时间为31 d,其中, 花期持续时间最长的品种是‘阿斯’、‘城固32’和‘皮削利’,均21 d;‘长叶卡林’和‘张林’的花期持续时间最短, 仅11 d。本文所选3个品种‘豆果’、‘九峰4号’、‘鄂植8号’,均与园内多个品种的花期重叠。
品种
Cultivar持续时长
Time of
bloom
duration /d花期(月-日) 品种
Cultivar持续时长
Time of bloom duration/d花期(月-日) 初花期
Time of start bloom盛花期
Time of full bloom末花期
Time of end bloom初花期Time
of start bloom盛花期
Time of start
bloom末花期
Time of end
bloom阿布桑娜Arbosana 16 03-27—31 04-01—06 04-07—11 科拉蒂Coratina 15 03-31—04-04 04-05—09 04-10—14 阿斯
Ascolano Tenera21 03-25—30 03-31—04-07 04-08—14 科新佛奥
Frantoio de Corsini16 03-31—04-06 04-07—11 04-12—15 奥托卡Ottobratica 18 03-28—04-01 04-02—07 04-08—14 克罗莱卡Koroneiki 18 03-26—04-01 04-02—07 04-08—12 巴尼亚Barnea 17 03-30—04-03 04-04—08 04-09—15 配多灵Pendolino 16 03-30—04-04 04-05—10 04-11—14 巴尼亚实生苗
Seedling of Barnea15 03-28—04-01 04-02—04 04-05—11 皮瓜尔Picual 16 03-29—04-02 04-03—04-07 04-08—13 贝翠丽Patrim 12 04-01—05 04-06—08 04-09—12 皮削利Picholine 21 03-25—30 03-31—04-05 04-06—14 贝拉特Berat 15 03-31—04-03 04-04—09 04-10—14 切姆拉尔
Chemlal de Kabylie16 03-29—04-04 04-05—07 04-08—13 长叶卡林
Kaliniot Changye11 04-06—08 04-09—12 04-13—16 沙龙奎
Salonenqne14 03-26—29 03-30—04-05 04-06—08 城固32
Chenggu—3221 03-24—04-02 04-03—06 04-07—13 坦彩Tanche 18 03-25—31 04-01—05 04-06—11 豆果Arbequina 19 03-23—31 04-01—06 04-07—10 美佳伦Megaron 18 03-26—29 03-30—04-04 04-05—12 鄂植8号Ezhi—8 18 03-31—04-03 04-04—09 04-10—17 云台Yuntai 15 03-28—31 04-01—06 04-07—11 佛奥Frantoio 15 03-30—04-04 04-05—08 04-09—13 白橄榄Ullir I bardhe 19 03-22—27 03-28—31 04-01—09 哥朗米扎
Mixaj Dukat19 03-21—28 03-29—04-01 04-02—08 小苹果
Manzanilla de Sevilla19 03-25—30 03-31—04-04 04-05—12 九峰4号Jiufeng—4 17 03-29—04-03 04-04—08 04-09—14 戈达尔Gordal 16 03-31—04-03 04-04—08 04-09—15 未知1号
Unknown—112 03-28—31 04-01—04 04-05—08 北庆米扎
Mixaj Peginit17 03-24—29 03-30—04-03 04-04—09 尖叶卡林
Kaliniot Jianye17 03-23—28 03-29—04-03 04-04—08 圆果卡林
Kaliniot Yuangua17 03-29—04-04 04-05—04-08 04-09—14 卡拉马达Kalamata 18 03-21—26 03-27—03-01 04-01—07 莱星Leccino 18 03-31—04-04 04-05—11 04-12—17 张林Zhanglin 11 04-10—13 04-14—15 04-16—20 中山24
Zhongshan—2418 03-30—04-06 04-07—09 04-10—16 Table 2. Flowering times of the cultivars for the flowering survey in 2014
-
由表 3可知:12个SSR位点共扩增出109个等位基因,每个SSR位点等位基因数为5~12,平均数为9.08个,其中,DCA11和GAPU103A的等位基因数最多,为12个,UDO99-19和EMO90的等位基因数最少,为5个。试验群体中的平均观测杂合度(Ho)为0.732,平均期望杂合度(He)为0.709,平均多态性信息量(PIC)为0.675。
SSR位点
SSR primer等位基因数
Observed number
of alleles(NA)/个片段长度
Size range/bp基因频率
Allelic frequency
(Pi)观测杂合度
Observed
heterozygosity(Ho)期望杂合度Expected
heterozygosity
(He)多态性信息量
Polymorphism information
content(PIC)DCA3 11 232~268 0.002~10.229 2 0.925 0.843 0.822 DCA11 12 126~179 0.002~10.483 3 0.546 0.710 0.680 DCA18 11 160~183 0.002~10.316 7 0.950 0.818 0.795 UDO19 5 100~168 0.002~10.739 6 0.354 0.429 0.401 GAPU59 8 208~224 0.002~10.662 5 0.392 0.522 0.484 GAPU89 10 158~208 0.002~10.443 7 0.842 0.704 0.658 GAPU71B 6 118~144 0.016~70.418 8 0.821 0.723 0.679 GAPU103A 12 136~196 0.002~10.193 8 0.933 0.863 0.845 UDO6 10 148~197 0.002~10.389 6 0.621 0.757 0.723 UDO11 11 100~129 0.002~10.204 2 0.938 0.854 0.835 EMO90 5 184~196 0.002~10.652 1 0.583 0.486 0.408 OLEH6 8 140~189 0.004~20.318 7 0.883 0.796 0.766 平均Average 9.08 0.732 0.709 0.675 Table 3. Polymorphism in olive cultivars with SSR primers
-
204个自由授粉子代的父本分析结果见表 4。
父本
Pollen donor母本Mother tree 豆果Arbequina 鄂植8号Ezhi-8 九峰4号Jiufeng-4 子代个数
合计Total
number of
progeny /个平均繁殖贡
献率Average
reproductive
contribution /%子代个数
Number of progeny/个繁殖贡献率
Reproductive contribution/%子代个数
Number of progeny/个繁殖贡献率
Reproductive contribution/%子代个数
Number of progeny/个繁殖贡献率
Reproductive contribution/%克罗莱卡Koroneiki 0 0.00 20 42.55 33 45.83 53 32.52 佛奥Frantoio 0 0.00 18 38.30 20 27.78 38 23.31 城固32 Chenggu-32 20 45.45 0 0.00 1 1.39 21 12.88 贝拉特Berat 9 20.45 0 0.00 0 0.00 9 5.52 巴尼亚Barnea 0 0.00 3 6.38 6 8.33 9 5.52 鄂植8号Ezhi-8 4 9.09 0 0.00 0 0.00 4 3.07 豆果Arbequina 0 0.00 1 2.13 4 5.56 5 2.45 阿斯Ascolano Tenera 1 2.27 0 0.00 3 4.17 4 2.45 圆果卡林Kaliniot Yuanguo 0 0.00 2 4.26 1 1.39 3 1.84 皮瓜尔Picual 3 6.82 0 0.00 0 0.00 3 1.84 中山24 Zhongshan-24 0 0.00 0 0.00 3 4.17 3 1.84 坦彩Tanche 2 4.55 0 0.00 0 0.00 2 1.23 莱星Leccino 0 0.00 1 2.13 1 1.39 2 1.23 北庆米扎Mixaj Peginit 1 2.27 0 0.00 0 0.00 1 0.61 歌朗米扎Mixaj Dukat 1 2.27 0 0.00 0 0.00 1 0.61 皮削利Picholine 1 2.27 0 0.00 0 0.00 1 0.61 小苹果Manzanilla
de Sevilla1 2.27 0 0.00 0 0.00 1 0.61 尖叶卡林Kaliniot Jianye 1 2.27 0 0.00 0 0.00 1 0.61 配多灵Pendolino 0 0.00 1 2.13 0 0.00 1 0.61 云台Yuntai 0 0.00 1 2.13 0 0.00 1 0.61 总计 44 68.75 47 78.33 72 90.00 163 Table 4. Paternity analysis of open-pollinated progeny from olive cultivars
在95%置信区间下检测出父本的子代163个(占子代总数的79.9%),其余子代的可信度未达到要求,未被视为子代的真实父本。36个候选父本中只检测出20个,其余16个未在子代中检测出。20个候选父本的贡献率为0.61%~32.52%,平均为5 %,其中, 平均繁殖贡献率最高的品种为‘克罗莱卡’(32.52 %),以‘克罗莱卡’为父本的子代共有53个;其次是‘佛奥’和城固32,繁殖贡献率分别为23.31%和12.88 %。平均繁殖贡献率大于5%的品种为‘克罗莱卡’、‘佛奥’、‘城固32’、‘贝拉特’和‘巴尼亚’。
-
表 4表明: 在‘豆果’64个子代中,在95%置信区间检测出父本的子代有44个(占68.75%)。共检测出父本品种11个,仅2个品种‘城固32’和‘贝拉特’的繁殖贡献率高于10 %,其中以城固32为父本的子代数量最多(20个),繁殖贡献率为45.45 %,‘贝拉特’为父本的子代数量为9,繁殖贡献率为20.45%。其余9个品种为父本的子代个数为1~4个,繁殖贡献率均小于10 %。由此可知,与‘豆果’亲和性较强的品种仅2个,为‘城固32’和‘贝拉特’,并且这3个品种的花期重叠,因此‘城固32’和‘贝拉特’可以作为‘豆果’的授粉树。此外,‘豆果’44个子代中未发现自交现象。
-
在‘鄂植8号’60个子代中,在95%置信区间检测出父本的子代有47个(占78.33%)。共检测出父本品种8个,‘克罗莱卡’和‘佛奥’2个品种的繁殖贡献率高于10 %,以‘克罗莱卡’为父本的子代数量最多(20个),繁殖贡献率为42.55 %,其次是‘佛奥’,子代数量为18,繁殖贡献率为38.30 %。其余6个品种为父本的子代数量为1~3个,繁殖贡献率均小于10%。由此可知,与‘鄂植8号’亲和性较强的品种为‘克罗莱卡’和‘佛奥’。由于‘克罗莱卡’和‘佛奥’与‘鄂植8号’花期重叠,因此‘克罗莱卡’和‘佛奥’可作为‘鄂植8号’的授粉树。此外,‘鄂植8号’47个子代中未发现自交现象。
-
’在‘九峰4号’80个子代中,95%置信区间检测出父本的子代有72个(占90%)。共检测出父本品种9个,仅2个品种(‘克罗莱卡’和‘佛奥’)的繁殖贡献率高于10 %,其中,以‘克罗莱卡’为父本的子代数量最多(33个),繁殖贡献率为45.83 %,其次是‘佛奥’,其子代数量为20,繁殖贡献率为27.78 %。其余7个品种繁殖贡献率均小于10%,子代数6个。由此可知,与‘九峰4号’亲和性较强的品种有2个, 为‘克罗莱卡’和‘佛奥’。由于‘克罗莱卡’和‘佛奥’与‘九峰4号’花期重叠,因此均可作为‘九峰4号’的授粉树。此外,‘九峰4号’72个子代中未发现自交现象。
3.1. 花期分析
3.2. SSR多态性分析
3.3. 自由授粉子代的父本分析
3.4. 油橄榄品种的亲和性
3.4.1. ‘豆果’
3.4.2. ‘鄂植8号’
3.4.3. ‘九峰4号
-
本文父本分析研究结果表明,‘克罗莱卡’和‘佛奥’与‘九峰4号’和‘鄂植8号’之间、‘城固32’和‘贝拉特’与‘豆果’之间的亲和性均较强,但‘克罗莱卡’和‘佛奥’与‘豆果’之间、‘城固32’和‘贝拉特’与‘鄂植8号’之间的亲和性均未被检测出,仅检测出‘九峰4号’的1个子代,这与Mookerjee等[8]利用父本分析方法的研究结果相同,即油橄榄不同品种间亲和性有差异。湖北省林科院[17]在湖北武昌测定了‘九峰4号’人工授粉的座果率发现, ‘中山24’与‘九峰4号’有一定亲和性,这与本文研究结果相同。姜成英等[10]和史艳虎等[25]在甘肃武都测定‘鄂植8号’与‘莱星’间人工授粉座果率,发现‘鄂植8号’与‘莱星’之间正反交授粉座果率均较高,二者间可以互为授粉树;而本文在四川凉山州油橄榄品种子代父本分析的研究发现,在‘鄂植8号’47个子代中,‘莱星’为父本的子代仅有1个,繁殖贡献率仅为2.13%,表明‘莱星’与‘鄂植8号’之间有亲和性,但亲和性较低。由此可见,相同品种在不同地点的亲和性具有一定的差异,可能是由于气候条件或研究方法的不同。同时油橄榄园内品种配置也会影响亲和性研究结果,在本研究的园内,‘城固32’和‘贝拉特’与‘豆果’之间定植距离较近,‘克罗莱卡’和‘佛奥’与‘九峰4号’和‘鄂植8号’之间定植距离较近,可能会对研究试验结果有一定的影响。根据3个品种与候选父本之间的亲和性,结合品种间花期重叠性的观测结果,可为四川省凉山州油橄榄建园时的品种最适授粉树配置提供依据。以‘鄂植8号’和‘九峰4号’为主栽品种时,可配置‘克罗莱卡’或‘佛奥’为授粉树,以‘豆果’作为主栽品种时,‘城固32’和‘贝拉特’可作为授粉树进行品种配置。
本文对‘豆果’、‘鄂植8号’和‘九峰4号’自由授粉子代的父本分析,均未发现自交现象,说明这3个品种在四川凉山州油橄榄园是自交不亲和。Diaz等[8]和Marchese[26]利用父本分析研究,也发现‘豆果’是自交不亲和;然而,De la Rosa等[12]利用自花授粉和杂交授粉子代的父本分析法研究‘豆果’自交亲和性,发现在西班牙不同地区油橄榄园,‘豆果’自花授粉20个子代中,有2个是自交产生的,而在其杂交授粉的30个子代中,有1个是由于‘豆果’自身花粉污染产生的,说明‘豆果’具有一定的自交亲和性。姜成英等[10]在甘肃陇南测定了不同年份‘鄂植8号’自花授粉座果率, 发现‘鄂植8号’坐果率均较低;湖北省林科院[17]于1987—1990年在湖北武昌测定了‘九峰4号’授粉座果率,发现‘九峰4号’自花授粉的座果率高于自由授粉和杂交授粉的。这些研究结果与本文研究结果有一定差异,是由于亲和性研究的测定方法不同。利用测定人工授粉座果率的方法,研究油橄榄品种亲和性,具有简单、直观、易操作等优点,但在授粉过程中,可能会存在花粉污染、授粉袋内外环境差异、花粉过多导致柱头堵塞等问题,还有树体营养竞争引起的落果,也会影响座果率。父本分析法具有准确性高、不受环境影响的优点,但只能利用子代进行父本分析,环境、营养竞争及树体自身状况对座果的影响均被忽略,所以会对品种亲和性研究结果产生一定的影响,甚至出现截然不同的研究结论[7, 12, 27]。
气候条件、树体营养状况、品种自身遗传特性等许多因素,都会影响油橄榄品种亲和性,因而不同油橄榄品种亲和性有一定差异[9]。如Gonzolez等[27]在西班牙不同地点油橄榄园,对不同年份‘豆果’授粉座果率进行测定,以研究其自交不亲和性,发现‘豆果’在地点La Caada的油橄榄园是自交不亲和的,而在地点Tabernas的油橄榄园是自交亲和的,并且2002年和2003年,‘豆果’自交亲和性亦有一定差异。因此,油橄榄品种亲和性因种植地点的不同,其自交亲和性存在差异,甚至同一地点测定年份的不同,其自交亲和性也有差异。由此可见,本文对油橄榄品种亲和性的研究结果,与他人的研究结果有一定差异,可能是由于测定年份、种植地点或试验方法不同造成的。由此可知,在不同种植地点对油橄榄品种亲和性,宜采用多种试验方法进行多年份测定,从而确定油橄榄品种间及品种内的亲和性。
-
利用12个SSR标记对四川省西昌市3个油橄榄品种自由授粉子代进行父本分析,结果表明,‘城固32’和‘贝拉特’与‘豆果’亲和性较强,‘克罗莱卡’和‘佛奥’与‘鄂植8号’和‘九峰4号’亲和性均较强,且品种间花期重叠。‘城固32’或‘贝拉特’可以作为‘豆果’的最适授粉树;‘克罗莱卡’和‘佛奥’可以作为‘鄂植8号’和‘九峰4号’的授粉树。SSR标记和父本分析是研究油橄榄亲和性的有效工具,可以用于油橄榄品种亲和性的研究,从而为油橄榄建园时的品种配置提供依据,同时也为今后油橄榄育种时亲本的选择提供了依据。
 DownLoad:
DownLoad: