



-
植物种群更新和种群中植物个体生活史的完成都依赖于生殖过程。植物生殖过程中的生殖分配格局以及植物如何调节其生殖分配以适应其生活型和年龄变化的动态,是生殖生态学研究的重要任务[1]。植物将有限的资源按一定的比例分配给营养构件(茎、叶)或者生殖构件(花或花序、果实或果序等)[2],投入到某一构件的资源量增加必然会降低投入到其它构件的资源量,即在植物的不同功能构件之间存在着一种权衡关系(trade off)。例如,植物如果增加投入到生殖(花或果)的资源(即生殖投入)时,就会降低投入到营养生长(茎、叶和根)上的资源(即营养投入)。其中,植物生殖构件的生物量占整个植物地上总生物量的比例被定义为生殖分配(reproductive allocation,缩写为RA)[3]。植物的生殖分配通常与植株个体大小、生殖年龄密切相关[4]。由于植物可供生殖的资源是有限的,竞争同一资源库的两个或多个生殖器官各构件或组分之间也应存在权衡关系[5],这种权衡关系可能存在于雄性与雌性功能之间,也可能存在于构件的数量与大小之间,如每果种子数量与大小之间[6-7]。研究植物的生殖分配格局及其构件各组分之间的权衡关系,有助于揭示其生殖与生存的相互关系,对于探讨植物的生活史适应对策具有重要的意义[8]。
水青树(Tetracentron sinense Oliv.)是水青树科(Tetracentranceae)仅有的一种高大落叶乔木。其花序穗状,下垂,生于短枝顶端,与叶对生或互生,花小,黄绿色,直径1~2 mm,无梗,常4朵成一簇;果实为蓇葖果长椭圆形,长3~5 mm, 种子4~6,条形,长2~3 mm;花期6—7月,果期9—10月。主要分布在我国西南和华中地区,多呈零星散布,种群更新困难,被列为国家二级重点保护植物[9]。目前,有关水青树生殖生态学方面的研究主要集中在繁育系统、传粉生态学、种子和幼苗生态学等方面[10-17],而关于水青树种群生殖分配方面的研究还缺乏相关报道,其生殖投入和生殖分配随生殖年龄的变化情况至今尚不清楚。
目前,植物生殖分配的研究大多集中在草本植物和灌木方面[1, 18],对乔木树种的生殖分配研究报道相对较少。基于构件理论,乔木树种1个生殖枝就相当于1株树的基本单元—构件(modules),一棵大树通常被认为是由基本构件单位不断重复而形成的多层次复合结构体[19-20]。在构件水平上研究小枝内部的资源分配与构型特征是乔木植物生活史对策研究的重要内容之一[21-22]。因此,本文采用固定样方法和典型抽样法,通过对四川省美姑大风顶自然保护区水青树不同径级个体标准生殖枝的生殖分配进行研究,在构件水平上探讨以下两个问题:(1)水青树的生殖投入和生殖分配随生殖年龄的变化规律如何?(2)水青树生殖构件和营养构件之间、生殖构件内各组分之间是否存在权衡关系?通过这些研究,探讨水青树的生活史适应对策及其致濒机制,为水青树种质资源的有效保护和管理提供科学参考。
HTML
-
研究地区位于四川省美姑县大风顶自然保护区龙窝保护站(103.20°~103.50° E,28.85°~28.81° N)。该地区属中亚热带季风湿润气候,寒冷、湿润、多雨。年均气温9.6℃,年最高气温在7月,7月平均气温19.5℃。年平均降水量1 100 mm左右,年均相对湿度80%左右。
2014年6月,参照王娟等[23]的研究方法在龙窝保护站海拔2 100~2 200 m区域的水青树种群建立150 m × 250 m的固定样地(3.75 hm2;103.14°~103.14° E,28.77°~28.78° N),坡向为东北坡,坡度30°,群落郁闭度40%,群落高度19.2 m。样地植被类型为落叶阔叶林,其乔木层以水青树为优势种,伴生有连香树(Cercidiphyllum japonicum Sieb. et Zucc.)、珙桐(Davidia involucrata Baill.)和五裂槭(Acer oliverianum Pax)等;灌木层以桦叶荚蒾(Viburnum betulifolium Batal.)为优势种,伴生有银叶杜鹃(Rhododendron argyrophyllum Franch.)等;草本层主要以高山委陵菜(Potentilla contigua Soják)为优势种,伴生有蝎子草(Girardinia diversifolia subsp. suborbiculata (Chen) Chen et Friis)、天名精(Carpesium abrotanoides L.)、东方草莓(Fragaria orientalis Lozinsk.)、蛇莓(Duchesnea indica (Andr.) Focke)、假楼梯草(Lecanthus peduncularis (Wall. ex Royle) Wedd.)和细风轮菜(Clinopodium gracile (Benth.) Matsum.)等。
-
2015年4月对样地内所有胸径(DBH)≥1 cm的水青树个体的胸径、树高、冠幅、枝下高等进行观测。由于缺乏水青树解析木资料,因此参照张远东等[24]、李怀春等[17]的研究方法(以胸径大小代表水青树植株的年龄),以10 cm为间隔划分胸径等级,共分出9级:J1 (DBH<20 cm)、J2(20~29 cm)、J3 (30~39 cm)、J4 (40~49 cm)、J5 (50~59 cm)、J6 (60~69 cm)、J7 (70~79 cm)、J8 (80~89cm)、J9 (DBH≥90 cm)。参照苏智先[1]和董艳红[25]的研究方法,于花期(7月)和果期(10月),每个径级选取3株水青树,从其东北方向(向阳)的树冠中层分别采集3个当年生标准生殖枝(n=9)。分离叶片、花序(或果序)和枝条,装于信封中带回实验室。
-
生殖构件(花序)和营养构件(叶、枝)经80℃烘箱至恒重后,放入干燥锅中冷却,用电子天平称重(精确到0.001 g)。果序构件经自然风干后,细分为果序轴、果皮和种子三部分,并测量果序轴长度、统计种子数;从每个标准枝中,随机选取300粒种子,用精度为0.01 mm的电子数显游标卡尺测量其长度、宽度、厚度;并测定干质量。
-
生殖配置(RA)是指生殖的资源量占总资源或营养资源投入量的比例[3, 22],用下式进行计算:
运用IBM SPSS Statistics 21.0统计软件对数据进行分析。在满足方差齐性的情况下,对不同径级个体水青树各构件生物量、生殖构件各组分生物量、RA和种子千粒质量采用单因素方差分析方法中的Duncan检验进行多重比较;若不满足方差齐性的要求,则采用Dunnett’s进行多重比较;对RA、生殖构件生物量与营养构件生物量之间进行相关性分析和线性回归分析;对种子数量和种子千粒质量进行相关性分析。
1.1. 自然概况
1.2. 样地调查及采样
1.3. 构件观测
1.4. 数据处理
-
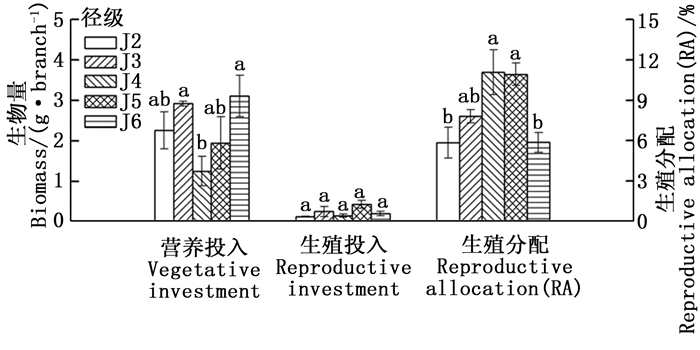
经连续2年调查发现,水青树种群内J1径级的植株均未有生殖现象;J7、J8、J9径级的花序过早脱落,因此分析其生殖与营养投入时缺少上述4个径级的资料。
由图 1可知,水青树的营养投入和生殖投入随径级变化整体上基本呈现出同增同减的趋势,营养投入呈现上升→下降→上升的趋势,生殖投入没有明显的波动趋势。其中,营养投入的最小值出现在J4,其与J3、J6之间具有显著性差异,而与其余径级之间差异不显著;其余径级相互之间差异均不显著。在生殖投入方面,各径级之间均没有显著性差异。

Figure 1. Dynamic changes of vegetative and reproductive investment of T. sinense with the increase of diameter class
-
随着径级的不断增大,水青树生殖枝的RA值呈现先上升后下降的趋势,在J2→J4期间逐渐增加,在J4径级达到最大值,J5径级保持相对稳定,之后迅速减小。其中,J2、J6分别与J4、J5之间差异显著,J3与其余径级之间、J4与J5之间差异均不显著(图 1)。
-
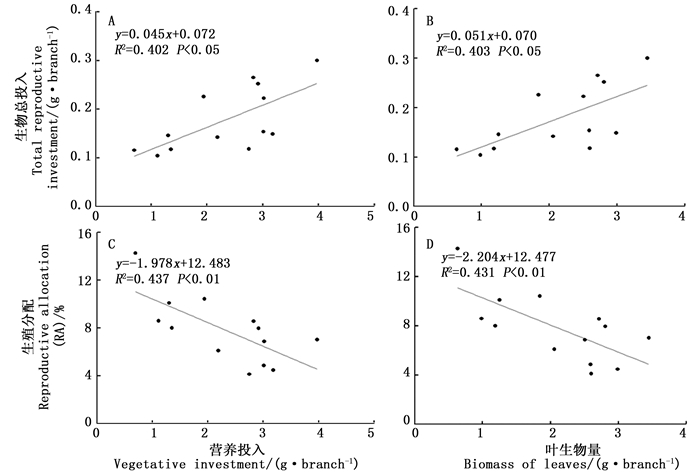
经Pearson相关性和线性回归分析发现,水青树生殖枝的生殖总投入量分别与营养投入、叶生物量之间呈显著直线正相关性(P < 0.05, 图 2-A, B);RA分别与营养投入、叶生物量之间呈极显著直线负相关性(P < 0.01, 图 2-C,D)。

Figure 2. Linear regression analysis between the vegetative moduals biomass and reproductive biomass or RA in T.sinense.
-
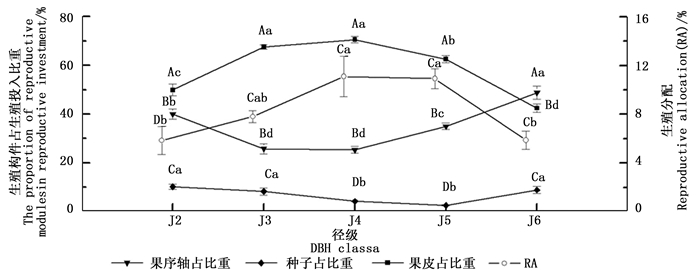
水青树的生殖构件可以分为果序轴、果皮和种子三部分。其果皮生物量所占比重随着径级的增大呈现出“低→高→低”的变化趋势,与RA的变化趋势基本一致;果序轴生物量比重随径级的增大呈现“高→低→高”的趋势,恰好与RA、果皮变化趋势相反;对果皮和果序轴生物量所占比重而言,J3、J4之间差异均不显著,分别与其余径级之间差异显著,其余各径级之间差异均不显著(图 3)。种子生物量所占比重随径级的增大呈现先下降后上升的趋势,与RA、果皮变化趋势基本相反;其中最小值出现在J5径级,仅为2.52%,其与J4之间差异不显著,分别于其余径级之间差异显著,其余各径级之间差异均不显著(图 3)。同一径级下,果序各组分所占生殖投入的比重之间均有显著性差异;其中种子生物量所占比重均为最小,且远远低于生殖的附属结构(果序轴和果皮);除J6外,其果序轴生物量占比重均为最大(图 3)。
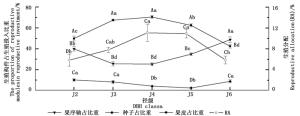
Figure 3. Dynamic changes of the proportion of reproductive modules accounted for reproductive investment and RA with the increase of diameter class
相关性分析结果表明,水青树生殖枝RA值与果序轴干质量之间存在极显著负相关性(P < 0.01),种子生物量与果皮干质量之间存在极显著的正相关性(P < 0.01)。
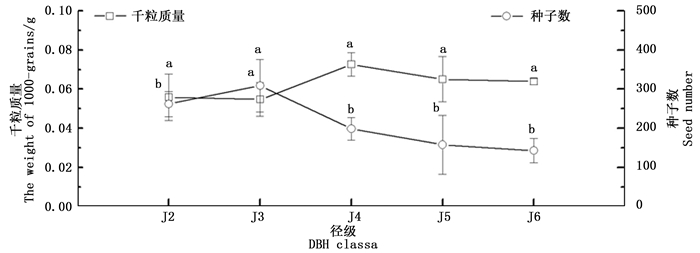
随着径级的增大,水青树生殖枝的种子数量呈现出先缓慢上升有逐级下降的趋势,在J3时达到最大值,在J6时达到最小;其中,J3径级的种子数与其他各径级的种子数具有极显著性差异,其他各径级之间则无显著性差异。
种子的千粒质量随径级的增大呈现出先缓慢下降,然后从J3→J4成明显增加的趋势,J4→J6呈逐渐下降的趋势(图 4),在J4时达到最大值。方差分析表明各径级之间种子千粒质量没有显著性差异。

Figure 4. Dynamic change of seed number and the weight of 1000-grains of T. sinense with the diameter class.
整体上看,种子千粒质量随种子数量的增加有减少趋势,随种子数量的减小而有所增加。相关性分析表明,两者之间存在显著的负相关关系(P < 0.05)。
2.1. 营养和生殖投入随径级的变化动态
2.2. 生殖分配随径级的变化动态
2.3. 生殖构件生物量、RA与营养构件生物量之间的相关性分析
2.4. 水青树生殖构件各组分的变化动态及其权衡关系
-
多年生植物一般需生长到一定大小才能开始生殖,即进行生殖的植株个体大小存在一个阈值[26-28]。研究发现,水青树在J1径级时,不具有生殖现象,直到J2径级才有生殖现象,这表明水青树进行生殖所需的个体大小阈值为DBH=20 cm的个体;在J2径级之前主要进行营养生长,为后期的生殖生长积累营养物质和能量。
对多年生植物而言, 生殖投入量通常随个体的增大而增加。本文研究发现,水青树生殖枝的生殖投入随径级的增大没有明显的变化趋势,营养投入的变化呈现上下波动的趋势,其结果与麻疯树(Jatropha curcas)[29]的变化趋势一致。
个体营养生长与生殖生长之间的对立统一关系一直是生殖生态学研究中的一个经典问题。水青树生殖枝的生殖总投入量与营养投入、叶生物量之间呈显著正相关关系,与麻疯树(Jatropha curcas)[29]、冰草(Agropyron cristatum)[30]的研究结果相似;生殖总投入量也与叶生物量之间呈显著正相关性,这表明水青树生殖枝的营养投入对其生殖投入具有促进效应,这一促进作用主要体现在营养投入和生殖投入基本呈现同增同减的趋势。
-
植物的生殖分配受年龄等植物本身因素的影响[8, 31]。热带棕榈(Astrocaryum mexicanum)和四川大头茶(Gardonia acuminata)等多年生木本植物的生殖分配会随年龄的增长呈直线增加趋势[32, 33],而冷杉(Abies fabri)的生殖分配会随年龄增长呈直线下降趋势[34]。濒危植物疏花水柏枝(Myricaria laxiflora)[35]、马尾松(Pinus massoniana) [18]和四合木(Tetraena mongolica)[4]的生殖分配却随植株年龄的增大呈现出先上升后下降的趋势。本文研究发现,水青树生殖枝的RA值随径级的变化呈现先上升后下降的趋势,其结果与濒危植物疏花水柏枝(Myricaria laxiflora)[35]、四合木(Tetraena mongolica)[4]相似,而不同于热带棕榈(Astrocaryum mexicanum)[29]、四川大头茶(Gardonia acuminata)[33]和冷杉(Abies fabri)[34]。
徐庆等[4]研究发现,四合木种群的生殖期根据其生殖年龄和生殖分配大小可分为生殖起动期、生殖增长期、生殖高峰期和生殖衰退期;其生殖值和生殖分配均随年龄增加呈现先上升后下降的趋势,其最大值出现在生殖高峰期,而在生殖衰退期呈现逐渐下降的趋势。我们研究发现,水青树RA值的最小值出现在J2径级,J2→J3、J3→J4之间均呈逐渐增加的趋势,在J4→J5达到最大值并保持相对稳定,J6径级迅速减小。由此表明,水青树的生活史过程中也可以将其生殖期划分为类似的四个时期:J2(20 ≤DBH﹤30 cm)为生殖起动期,J3(30≤DBH﹤40cm)为生殖增长期,J4、J5(40≤DBH﹤60 cm)为生殖高峰期,J6径级之后(60≤DBH)为生殖衰退期。
在植物的生殖配置研究中,营养生长和生殖分配之间是否存在权衡关系常常是研究的一个热点问题。有研究表明,对多年生植物而言,其幼龄和高龄阶段通常会选择较低的RA以保证其个体的生长和存活[36, 37];在较为合适的胸径或者是年龄阶段时,生殖投入量适度,RA最大,能获得最大的生殖产量而耗用相对较少的资源量,具有较高的生殖效率,以保证其个体繁衍和种群的扩张[29]。我们发现,水青树的生殖分配与其营养投入之间存在显著或极显著负相关关系,在其在生殖起动期和生殖衰退期均具有较低的RA值,在生殖高峰期具有较高的RA值,可能正是水青树为了更好地保证其个体的生长存活和其种群的扩张而形成的生殖适应性机制。
-
Reekie[38]认为:生殖分配随个体大小增加而减少,或许是由于生殖代价随个体大小增大的一个直接结果,代价的增大可以部分地解释为对生殖附属结构的分配增加。研究发现,水青树生殖构件生物量配置的比例始终呈现果皮>果序轴>种子的趋势;随着径级的增大其果皮所占比例呈现与RA值一致的变化趋势,而果序轴和种子所占比例呈现相反的趋势,这表明水青树生殖构件各组件在生物量配置方面存在较为明显的权衡关系。果序轴和果皮作为水青树生殖构件的生殖附属结构,在生殖不同时期均利用了生殖投入的大量资源,尤其是高龄时期将大量的资源投入到附属结构,其投入到种子中真正用于生殖后代的资源将越来越少。有研究表明长期进化形成的小种子虽有利于传播和散布,但不利于种子和幼苗的存活[11]。所有径级的水青树投入到种子的生殖分配比例都是最小的,其生殖附属结构占据了生殖分配大量资源,将导致其种子和幼苗的适合度降低,这可能是限制水青树种群自然更新的一个主要因素。
种子资源投入有两种截然相反的策略,即产生少量的大种子以便在竞争中占优势,和产生大量小种子以占据更多的安全位[39]。水青树生殖构件中,其种子千粒质量和种子数量呈现显著负相关性,与玉簪(Hosta plantaginea)的研究结果相似[5];在生殖增长期(J3),水青树种子千粒质量相对最小,而种子数量在整个生殖期中处于最大值,这表明水青树在此时期采取了产生大量小种子的r-对策,这正好与桃金娘(Rhodomyrtus tomentosa)的研究结果相似[40],通过这种方式可以占据更多的生态位,有助于水青树种群的扩散和增大;在生殖盛期(J4),其千粒质量相对最大,而种子数相对最小,这表明此时水青树采用了产生少量大种子的策略,这也与阴生灌木九节(Psychotria rubra)的研究结果相似[41],可以将大部分的能量用于提高种子的质量,以保证后代在竞争中的优势,符合典型的以“质”取胜的k-对策。水青树种子特征在生殖的不同时期分别采取不同的生活史对策,以分别适应种群的扩散和种群的持续发展这也与Acosta等发现多年生灌木Cistus ladaniferde种子的特征相类似[42],可能是水青树在长期的演化发展过程中所形成的生殖适应性机制。
3.1. 生殖投入的变化规律
3.2. 生殖分配的变化规律
3.3. 生殖构件内各组分之间的权衡关系
-
(1) 水青树生殖枝的营养投入对其生殖投入具有促进作用,两者之间不存在明显的权衡关系。
(2) 水青树生殖枝RA值随径级的增大呈现先增加后减小的趋势,其生殖期可划分为生殖初始期、生殖增长期、生殖高峰期和生殖衰退期等4个时期。
(3) 水青树生殖构件各组分之间存在明显的权衡关系;所有径级投入到种子的生殖分配均为最小,其生殖附属结构占据了生殖分配的大量资源,可能是限制水青树种群自然更新的一个主要因素。
(4) 水青树种子特征在生殖的不同时期分别采取不同的生活史对策,可能是水青树在其演化发展过程中形成的生殖适应性对策。
 DownLoad:
DownLoad: