




-
混交是促进珍贵树种干材生长、塑造良好干形及提高林分生产力和生物多样性的重要育林措施[1-2]。由于林窗、林隙等冠层干扰的存在,混交树种处于一个复杂的光环境中,各树种对异质光环境的适应,主要是通过种间形态和生理特性的差异性响应,表现出不同的表型可塑性,诸如形态塑性[3]、生理塑性[4]和生物量分配塑性[5]等,进而影响到混交树种生产力的高低。除树种基因型差异外[6-7],树木对光环境的响应模式还与其个体的发育阶段密切相关[9]。现有针对性的研究树木某个阶段的生理特性得出的结论难以全面地说明其光适应性能力及生长差异机制。因此,研究混植状态下不同树木基因型对光照的塑性反应及发育阶段特征,有助于全面了解林木适应异质光环境的差异和机制,进而为选育觅光效率高的林木新品种和人工林持续高产提供新的突破口。
木荷(Schima superba Gardn. et Champ.)是我国亚热带地区重要的珍贵阔叶用材、生物防火和生态防护树种,具有速生、丰产、材质优异和适应性强等特点[10]。自2001年以来,本研究组开展了我国木荷的多目标育种研究,发现其存在丰富的种源和家系变异,并初选出一批优良种源和家系[11-13]。研究表明,在异质养分环境中,杉荷混交可显著提高木荷和杉木(Cunninghamia lanceolata (Lamb.) Hook.)的生产力水平,木荷生长优势尤为明显[14]。研究还发现,使用不同地理区域的木荷种源营建的混交林生产力差异很大,作者已从根系形态、根系空间构型、氮磷效率等觅养塑性方面基本阐明了异质养分环境中杉荷混交增产原因及木荷种源间生长竞争能力差异机制[10, 14-15]。除觅养行为不同外,木荷种源生产力差异还与其觅光能力有关。为此,本研究选择3个不同生产力的木荷种源,模拟森林异质分布养分和不同光环境,通过1年生和2年生两个发育阶段的木荷/杉木幼苗混植盆栽试验,持续观测木荷种源幼苗生长和光合生理特性,进一步揭示木荷种源生长竞争能力及生产力差异机制,为选育竞争能力强和高效利用光能的木荷新品种以及制定高效育林措施提供科学依据。
HTML
-
试验在浙江省淳安县富溪林场苗圃(119.13°E,29.48°N)遮荫大棚内进行,该地所处海拔高度130 m,属亚热带季风气候,温暖湿润,光照充足,雨量充沛。年均降水量1 701.2 mm,全年降水集中在5—9月,年平均气温17.5℃,1月平均气温3.2℃,7月平均气温28.5℃,无霜期241 d,年日照时数1 850.3 h。土壤为红壤,pH值5.12。
-
试验以浙江龙泉(ZJLQ)、江西信丰(JXXF)和福建建瓯(FJJO)3个木荷种源幼苗为材料,与其混植的杉木幼苗种子来自浙江省龙泉市林科院第3代杉木无性系种子园。采用直径25 cm、高度40 cm的无纺布袋作为盆栽容器。盆栽基质取自浙江省淳安县富溪林场的贫瘠酸性红壤,其全氮和全磷含量分别为0.34、0.35 g·kg-1,水解氮、速效钾和有效磷含量分别为45.3、40.9、1.08 mg·kg-1,有机质含量6.11 g·kg-1。构建异质养分环境所用控释肥委托山东金正大集团专门生产,控释氮肥(速效氮含量440 mg·kg-1)、控释磷肥(有效磷含量440 mg·kg-1)和控释钾肥(速效钾含量440 mg·kg-1)的肥效均为2 a。
-
基于文献[16]报道的森林土壤数据,模拟土壤垂直空间养分分布的异质性,即表层养分丰富,向深层迅速降低的森林土壤养分含量自然分布状况。利用风干过筛后贫瘠酸性红壤为基质,将盆栽容器从上而下分为4层,每层高度10 cm,每层加入控释肥构建异质养分环境。第1层每千克基质中加入控释氮肥372.93 g、控释磷肥60.81 g和控释钾肥402.93 g,相当于有效N、P、K含量分别为300、30、300 mg·kg-1,第2层加入1/3第1层养分含量控释肥,第3、4层均不添加控释肥。
-
以自然光照为基础,通过搭建遮阳网控制光环境,分别设置全光照(100%光照,L0)、轻度遮阳(50%光照,L1)和强度遮阳(25%光照,L2)3个遮阳处理,以模拟全光照、林缘和林窗光环境[17]。光照控制从2015年5月上旬开始,到2016年12月初结束。
-
盆栽试验在浙江省淳安县富溪林场钢构自控荫棚内进行,荫棚四周放置林场培育的普通容器苗,而试验各处理盆栽苗摆放在靠近中心位置,以确保试验处理环境一致。3个木荷种源及杉木种子于2015年1月10日播种,5月7—8日选用生长整齐一致的芽苗移栽。混植的2株芽苗分别栽植在盆中线的正中点两侧5 cm处。每试验处理20盆,即重复20次。苗木按正常管理2 a直至采收。
-
于2015年9月上旬和2016年8月下旬,随机选取不同光环境木荷种源5株,每株选择顶芽下第3~4片完全展开的1片成熟叶,使用Li-6400便携式光合作用测定系统(LI-COR Inc6400,Nebraska,USA)进行光响应曲线的测定,并采用非直角双曲线模型进行光响应曲线拟合求得最大净光合速率(Pmax)、光补偿点(LCP)、光饱和点(LSP)及暗呼吸速率(DR)[13]。光合作用参数测定后,立即将所测定的叶片剪下带回实验室测量叶绿素含量[18]。
-
叶绿素荧光用PAM-2500型便携式叶绿素荧光仪(德国Walz公司生产)测定。叶片暗适应30 min后测量最小荧光(Fo)和最大荧光(Fm),每种光环境下木荷种源随机选取5株,测量5片叶。通过计算获得可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)等参数[19]。
-
试验于2015年12月中旬和2016年12月上旬分2期收获。每期每处理随机选取6株生长正常的木荷植株测量其苗高和地径,然后按根、茎和叶分开,经105℃杀青30 min,80℃烘干至恒质量,测定各部分的干质量,并计算每一植株根冠比(地上生物量/根系生物量)。
比叶面积的测量在试验收获时进行,从每个处理中随机选择4株正常生长的木荷,每株随机选取其中上部10片完全展开、健康的成熟叶片,带回实验室用扫描仪扫描,测量和计算比叶面积[20]。
-
用SPSS19.0统计软件的ANOVA程序按种源、光环境进行单因素和双因素的方差分析,以检验种源、光照及其互作效应的显著性(α=0.05)。采用Excel 2007软件计算和整理所得试验数据,并进行图表绘制。
1.1. 试验地概况
1.2. 试验材料
1.3. 试验设计
1.3.1. 异质养分环境构建
1.3.2. 不同光环境的构建
1.3.3. 盆栽试验
1.4. 形态及生理指标测定
1.4.1. 光合生理指标测定
1.4.2. 叶绿素荧光参数测定
1.4.3. 植株形态和生物量测定
1.5. 数据分析
-
经过1 a的生长,福建建瓯种源木荷幼苗表现出较强的生长势,其苗高和地径分别比浙江龙泉种源的高113.6%和41.1%,比江西信丰种源的高76.6%和39.8%(P<0.05),而浙江龙泉种源和江西信丰种源的苗高和地径无显著差异(P>0.05)(表 1)。3个木荷种源对遮阳处理的响应有所差异,福建建瓯种源的苗高和地径在遮阳后呈显著下降趋势,与全光照(L0)相比,在轻度遮阳(L1)和强度遮阳(L2)下其苗高分别下降20%和55%,地径降幅也有类似趋势,但2种遮阳处理对浙江龙泉种源和江西信丰种源的苗高和地径的生长影响则较小,表明福建建瓯种源对遮阳更为敏感。长期遮阳(2 a)导致3个木荷种源幼苗高径生长不同程度的降低,其中,福建建瓯种源在轻度遮阳(L1)和强度遮阳(L2)条件下的苗高和地径均明显低于全光照(L0)(P<0.05)。木荷不同种源间2年生幼苗高径生长规律与1年生时类似,福建建瓯种源的最高,说明在遮阳条件下福建建瓯种源生长仍保持一定优势。
种源
Provenance光环境
Light environment1年生One year old 2年生Two years old 苗高
Seedling height/cm地径
Ground diameter/mm苗高
Seedling height/cm地径
Ground diameter/mm浙江龙泉ZJLQ L0 11.4a 3.4a 67.3a 9.5a L1 13.4a 3.2a 62.2b 8.4b L2 13.1a 3.2a 60.5b 8.7b 江西信丰JXXF L0 14.8a 3.6a 99.7a 11.9a L1 14.1a 3.4a 89.0b 10.5a L2 16.9a 2.9b 76.0c 10.7a 福建建瓯FJJO L0 36.0a 5.7a 115.0a 12.9a L1 28.8b 5.2a 101.3b 11.2b L2 16.2c 3.0b 79.0c 10.0b 注:不同小写字母表示0.05水平存在显著差异(P<0.05)。L0:100%光照, L1:50%光照,L2:25%光照。下同。
Note: Different letters indicated significant difference under 0.05 level. L0: 100% of natural illumination, L1: 50% of natural illumination, L2: 25% of natural illumination. The same below.Table 1. Differences of three S. superba provanances in seedling height and ground diameter in different light environments
-
图 1显示:3个木荷种源幼苗的全株干质量福建建瓯种源的最高,其1年生幼苗分别较浙江龙泉种源和江西信丰种源高3.4倍和2.1倍,2年生时分别高1.2倍和0.3倍。3个木荷种源幼苗的叶干质量差异也有相似的规律。方差分析结果表明:无论1年生还是2年生幼苗,3个木荷种源的全株干质量和叶干质量的遗传差异均达极显著差异(表 2)。不同木荷种源幼苗的干质量对光照的响应差异明显。福建建瓯种源幼苗的全株干质量和叶干质量在2个发育阶段均随光照强度的减弱而明显降低,浙江龙泉种源和江西信丰种源干质量降幅则相对较小。如1年生时,福建建瓯种源幼苗全株干质量在L1光环境下较全光照(L0)处理显著降低46.4%,L2光环境下下降幅度达76.3%,但仍显著高于同水平下的其它种源幼苗全株干质量。
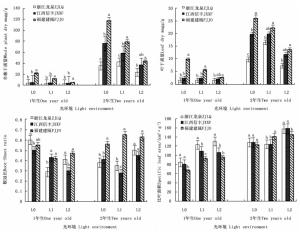
Figure 1. Differences of three S.superba provenances in seedling growth in different light environments
时间
Period因素
Factor全株干质量
WDM根冠比
RSR叶干质量
LDM比叶面积
SLA最大净光合速率
Pmax光饱和点
LSP光补偿点
LCP暗呼吸速率
DR1年生
One year old种源Provenance 18.21** 1.06 13.42** 0.94 0.35 10.16** 3.74* 5.54** 光照Light 5.79** 8.80** 3.48* 5.49* 4.37* 2.41 3.62* 3.43* 种源×光照Provenance ×Light 4.34** 0.67 3.16* 2.28 1.67 0.79 0.83 1.47 2年生
Two years old种源Provenance 25.70** 1.17 14.15** 0.94 6.44** 4.21** 3.68** 8.49** 光照Light 20.74** 0.11 26.17** 8.43** 0.17 5.01** 11.66** 9.13** 种源×光照Provenance ×Light 4.15** 0.69 5.61** 1.25 1.27 1.94 0.25 1.58 注:*表示在0.05水平上差异显著,**表示在0.01水平上差异显著。下同。
Note: * Means the significant difference at 0.05 level. ** Means the significant difference at 0.01 level.The same below.Table 2. ANOVA analysis of seeding growth and photosynthetic parameters of three provenances of S. superba in different light environments
遮阳处理后,3个木荷种源的1年生幼苗在干质量分配上表现出较一致的光响应模式,即降低根冠比,增加比叶面积,且均与全光照(L0)处理差异显著(P<0.05),如L1和L2遮阳处理下,福建建瓯种源幼苗的根冠比较全光照(L0)分别降低23.6%和14.5%,而比叶面积分别增加39.1%和43.6%,表明各种源幼苗干物质较多地分配至地上部分尤其叶片,增加了整株叶面积,从而更有效地捕获光能,以适应弱光环境。然而长期遮阳(2 a)下,参试木荷种源幼苗却表现出不同的弱光适应方式,如在轻度遮阳(L1)下,浙江龙泉种源和江西信丰种源幼苗通过加大叶干物质分配,降低根冠比,而福建建瓯种源幼苗通过增加比叶面积来维系较高的光吸收效率(图 1)。
-
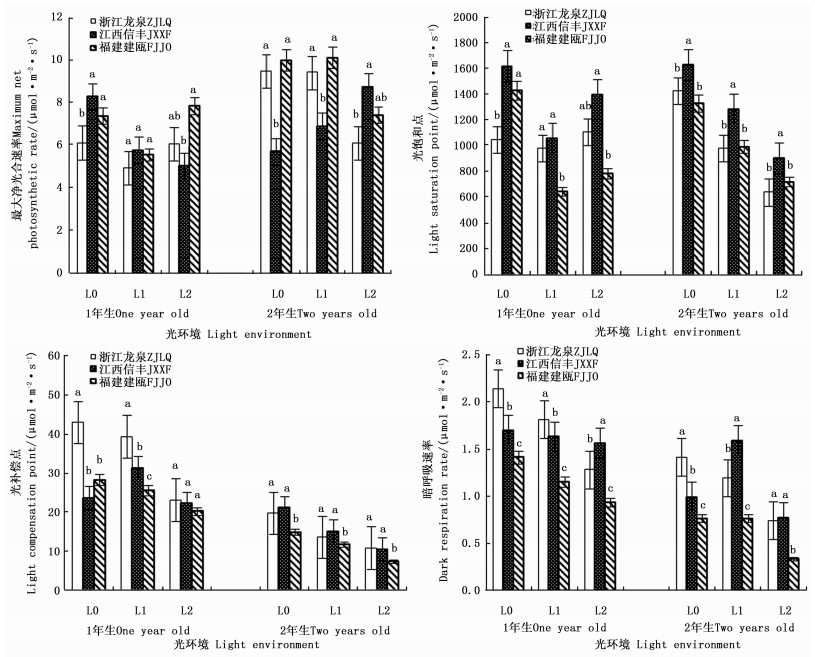
遮阳处理不仅影响了木荷幼苗的生长性状,而且影响了其光合生理反应,且不同种源间差异显著(表 2)。1年生时,随着遮阳强度的增大,3个木荷种源幼苗的最大净光合速率、光饱和点均呈先降低再升高的趋势,其中,轻度遮阳(L1)下,生长势较强的福建建瓯种源幼苗的最大净光合速率和光饱和点分别较全光照(L0)光环境下降24.7%和17.9%。在强度遮阳(L2)下,其光补偿点、暗呼吸速率也下降明显,分别降低19.5%和28.3%。不同遮阳环境中,福建建瓯种源幼苗的光饱和点、光补偿点和暗呼吸速率均显著低于浙江龙泉种源和江西信丰种源。2年生时,3个木荷种源幼苗的光饱和点和光补偿点对光强的响应趋势较一致,即随光照强度减弱而下降,其中,福建建瓯种源的2个光合指标值最小,其暗呼吸速率也显著低于其它种源。推测遮阳条件下,2个发育阶段的福建建瓯种源幼苗较高的生长势不仅与其光补偿点和光饱和点大幅降低,提高利用弱光能力有关,而且还与其维持较低的呼吸消耗,积累更多干物质相关(图 2)。
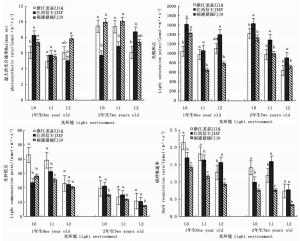
Figure 2. Differences of three S. superba provanances in photosynthetic parameters in different light environments
-
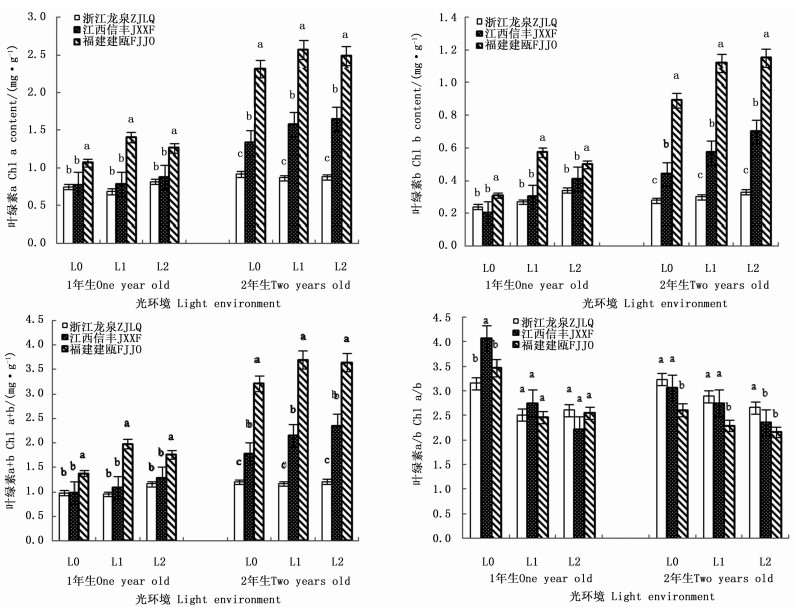
在2个发育阶段,木荷幼苗叶绿素含量对光照的响应因种源而异(表 3)。干物积累量较大的福建建瓯种源1年生和2年生幼苗,其叶绿素a、叶绿素b和叶绿素a+b含量均随遮阳强度的增加先明显升高后保持稳定,且各叶绿素含量均显著高于同一遮阳环境下的另外2个种源(图 3)。与之不同的是,浙江龙泉种源和江西信丰种源1年生幼苗的叶绿素a和叶绿素a+b含量受遮阳的影响较小,各光环境间差异不显著,但江西信丰种源2年生幼苗3种叶绿素含量在遮阳后也显著增加。在叶绿素组成比例上,遮阳使得2个发育阶段各种源幼苗的叶绿素a/b值均显著下降,导致L1和L2遮阳处理的3个木荷种源1年生幼苗的叶绿素a/b值差异不明显,但福建建瓯种源2年生幼苗叶绿素a/b值却显著低于同遮阳环境下的其它种源,表明福建建瓯种源幼苗的耐阴性较浙江龙泉种源和江西信丰种源高。
时间
Period因素
Factor叶绿素
aChl a叶绿素
bChl b叶绿素总量
Chl a+b叶绿素
a/bChl a/b最小荧光
Fo最大荧光
FmPSⅡ最大光化学效率
Fv/FmPSⅡ潜在活性
Fv/Fo1年生
One year old种源Provenance 5.67** 6.35** 3.58* 0.38 0.10 1.53 3.99* 3.90* 光照Light 1.79 5.85* 3.18* 8.62** 0.51 0.47 0.10 0.25 种源×光照Provenance ×light 0.65 0.42 0.52 0.91 1.29 2.66* 1.59 1.66 2年生
Two years old种源Provenance 8.16** 16.21** 12.53** 7.53** 0.97 2.30 0.20 0.12 光照Light 2.11 8.54** 2.52 6.36** 6.41** 9.70** 1.51 1.27 种源×光照Provenance ×light 1.15 5.02* 3.08 2.47 4.90** 7.50** 0.88 0.74 Table 3. ANOVA analysis of leaf chlorophyll content and chlorophyll fluorescence parameters of three provenances of S. superba in different light environments
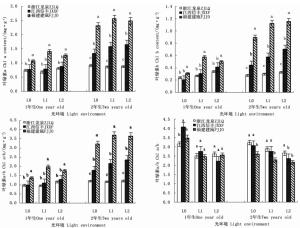
Figure 3. Differences of three S. superba provenances in leaf chlorophyll content in different light environments
-
表 2显示:遮阳处理对3个木荷种源幼苗叶绿素荧光参数的影响也较大。由图 4可知:在2个发育阶段,福建建瓯种源幼苗的最小荧光(Fo)受遮阳影响显著降低,在L1和L2遮阳处理下,该种源1年生幼苗的Fo值分别较全光照(L0)处理降低13.6%和15.0%,2年生幼苗的Fo值分别降低5.1%和30.2%。浙江龙泉种源1年生幼苗的Fo受遮阳影响略有降低,而江西信丰种源1年生幼苗的Fo值则在遮阳后升高明显。随着遮阳时间的延长,轻度遮阳(L1)导致浙江龙泉种源2年生幼苗的Fo值明显下降,在强度遮阳(L2)时其Fo值又明显升高,而江西信丰种源2年生幼苗的Fo变化趋势恰与浙江龙泉种源相反(图 4)。Fo值升高意味着PSII反应中心受到破坏或失活[21],因此,推断3个种源中福建建瓯种源幼苗弱光适应的能力最强。
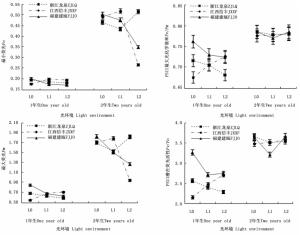
Figure 4. Differences of three S. superba provenances in chlorophyll fluorescence parameters in different light environments
Fv/Fm和Fv/Fo分别用于度量植物叶片PSⅡ原初光能转换效率和PSⅡ潜在活性[8, 22]。遮阳后福建建瓯和浙江龙泉种源1年生幼苗的Fv/Fm值均显著低于全光照(L0)处理,并随着遮阳强度的增加而降低,但江西信丰种源1年生幼苗的Fv/Fm值却随遮阳强度的增加而提高。在不同光环境下,浙江龙泉种源和江西信丰种源幼苗的Fv/Fm值均小于福建建瓯种源,尤其在全光照(L0)和L1条件下差异显著;Fv/Fo值的变化也如此,这表明福建建瓯种源1年生幼苗具有较高的光合电子传递活性,其光合器官能把所捕获的光能较高效地转化为三磷酸腺苷(ATP)和还原型辅酶Ⅱ(NADPH),从而为光合同化提供充足的能量和还原力。2年生时,3个木荷种源幼苗的Fv/Fm和Fv/Fo值均较1年生幼苗有明显升高,其中,不同光环境各种源幼苗的Fv/Fm值均处于0.75~0.85正常范围之内[23],且各光环境间及相同光环境种源间差异不明显,表明长期遮阳后3个木荷种源幼苗均对弱光环境有了一定的适应性(图 4)。
2.1. 遮阳处理对3个木荷种源幼苗生长指标的影响
2.2. 遮阳处理对3个木荷种源幼苗干物质积累与分配的影响
2.3. 遮阳处理对3个木荷种源幼苗光合生理指标的影响
2.4. 遮阳处理对3个木荷种源幼苗叶绿素的影响
2.5. 遮阳处理对3个木荷种源幼苗叶绿素荧光参数的影响

-
木荷天然分布广泛,种内变异丰富,根据种源试验优选木荷优良种源造林,可获得较高的遗传增益[24-25]。本文针对混植条件,设置不同光环境的盆栽试验,研究不同发育阶段木荷种源幼苗生长对光照响应的差异,结果表明:无论1年生还是2年生幼苗,参试木荷种源的苗高、干物质积累量、光合生理等主要性状指标均存在显著的种源变异。这种种源差异可能与木荷长期适应产地地理环境的结果有关。福建建瓯种源位于木荷地理分布中心,当地水热资源丰富,光照适宜,立地条件良好,促成了木荷优良基因型的形成,植株能适应不同光环境,光合能力较强,呼吸消耗最低,同化物最易积累[13];江西信丰种源位于木荷二级产区,年均温度较高,降水偏少,日照时数和强度均较大,植株光合能力和呼吸消耗适中,生长状况中等;北部浙江龙泉种源地理纬度较高,日照时数较短,水热资源和立地条件一般,植株光补偿点和暗呼吸速率较高,光合能力较低,同化物不易积累,生长较差[13, 24]。
处于竞争中的优势植物往往通过功能性状的改变,即不同表型可塑性,积极获取和利用资源来应对异质性环境条件,更大程度地促进生长,维护自身优势地位。作者前期研究表明,在异质养分环境中,与杉木混植时,木荷细根大量增生,且占据富养表层,从而增强养分获取,提高了竞争力[14]。同时研究还发现,不同木荷种源可塑性能力差异导致其生长竞争能力不同,福建建瓯种源根系形态和生理可塑性较强,其生长竞争优势明显高于其它参试种源[10, 15]。除根系觅养可塑性外,植物对光环境变化的适应能力也是影响其生长和竞争能力差异的主要因素[17, 26]。光饱和点和光补偿点是评价植物耐荫性的重要指标,已有研究表明这2项指标的降低是树种适应林下弱光环境的一种表现[27]。本研究结果表明,遮阳后3个木荷种源幼苗在2个发育阶段的光饱和点和光补偿点显著降低,表明3个木荷种源幼苗对遮阳环境均有一定的适应能力。但相同遮阳环境下福建建瓯种源幼苗较浙江龙泉种源和江西信丰两种源具有更低的光饱和点和光补偿点,单位叶面积的叶绿素含量明显提高,使得福建建瓯种源幼苗在弱光环境下可有效利用较低的光量子密度,获得较大的碳收益。除此之外,遮阳环境下较低的暗呼吸率也减少了福建建瓯种源幼苗自身的碳损耗,保持其干物质的积累在较高水平,这一结果与杨莹等[28]研究结论相似。从植株整体碳收益和损耗考虑,弱光环境下福建建瓯种源幼苗较其它2个种源幼苗具有较高的光合效率和较低的暗呼吸效率,因而具有明显的生长优势。
叶片是植物对环境变化最敏感的器官,其形态特征和叶绿素含量是植物适应和利用环境因子的重要指标[29-30]。在本研究中,遮阳后2个发育阶段生长势均较高的福建建瓯种源幼苗其比叶面积、叶绿素a、叶绿素b和叶绿素a+b含量均显著高于全光照环境,叶绿素a/b值则明显低于全光照环境,表明福建建瓯种源幼苗通过扩大叶面积、增加光吸收,特别增加叶绿素b含量(叶绿素a/b值减小),提高其对蓝紫光的利用效率来增强弱光适应能力[18]。同时,不同光环境下福建建瓯种源幼苗的叶绿素含量也远高于浙江龙泉种源和江西信丰种源,其叶片干物质量是这两种源的1.3~3.5倍,意味着福建建瓯种源幼苗全株总叶面积更大,光合能力及适应不同光环境的能力更强。此外,光强变化导致生物量分配模式的改变也是植物幼苗适应不同光环境的重要策略[28]。本研究中,遮阳后3个木荷种源1年生幼苗根冠比均显著降低,将更多的生物量分配给叶片进行光合作用以获得更多的能量,这种生物量分配模式对早期植物幼苗在森林冠层下不利的光环境中生存和生长非常有利,这与王暾等[31]研究结果类似。然而,与浙江龙泉种源和江西信丰种源不同,在遮阳环境下福建建瓯种源2年生幼苗的根冠比增加明显,分析原因可能是与本研究设置的异质养分环境和混植条件有关,随着混植苗木年龄和根系的增大,其地下竞争也趋于激烈,福建建瓯种源幼苗将更多的生物量向根系部分转移,提高了对土壤养分的吸收和利用,进而促进其整株苗木的生长及与混植杉木的竞争。推测福建建瓯种源具有较高的生物量分配塑性也是其生长优势形成的主要原因之一。
-
叶绿素荧光与光合作用效率密切相关,外界环境因子对光合作用的影响均可通过叶绿素荧光反映出来[21]。在本研究中,第1个生长期福建建瓯种源和浙江龙泉种源幼苗在遮阳后Fv/Fm值下降并同时伴随Fo值的下降,意味着两种源叶片叶绿分子捕获激发能的效率及光系统Ⅱ(PSⅡ)潜在活性降低,减弱了植株正常的光合作用,推测在遮阳处理下其叶片的结构和生理特征产生了相应的适应性变化[23],这表明福建建瓯种源幼苗能适应多种光环境,光照生态幅较宽,具有较好的光适应性和可塑性。江西信丰种源幼苗在遮阳后Fo值增加,意味着其光合器官受到一定程度的破坏或可逆失活,光合作用原初反应过程受抑制,光量子产量和光合电子传递速率降低。何森等[32]研究表明,Fo、Fm值较低,Fv/Fm、Fv/Fo值较高的植物,其PSⅡ反应中心的光量子电子传递和转换效率也较高。也有学者研究证实,Fv/Fm与光合作用的净产率和生长量呈正相关[33]。在本试验所设置的2个遮阳环境下,福建建瓯木荷种源1年生幼苗的Fo值最低,Fv/Fm值最高,这与其较高的干物质生产能力相符。然而,值得注意的是,2年生时3个参试木荷种源幼苗的Fv/Fm值较1年生幼苗有明显的增加,但不同遮阳环境的Fv/Fm值与全光照之间及种源间的差异较小,且处于未受环境胁迫的0.75~0.85正常范围之内,说明长期遮阳后,3个木荷种源幼苗均适应了弱光环境。木荷种源1年生与2年生幼苗对弱光环境适应性差异可能与树木生活史较长,其表型塑性在不同发育阶段会发生显著变化有关[9, 34]。因此,在评价木荷种源对不同光照响应能力时,要重视其所处的发育阶段,避免得出相悖或混淆的结论。随着木荷林龄的增长,木荷种源生长和表型塑性差异是否会持续发生变化,有待于进一步研究。
3.1. 不同木荷种源幼苗植株形态及光合生理特征对光照响应的差异
3.2. 不同木荷种源幼苗的叶绿素荧光对光照响应的差异
-
本试验模拟森林不同光环境,通过木荷/杉木幼苗混植盆栽试验,研究1年生和2年生两个发育阶段的木荷种源幼苗生长和光合生理特性,认为不同木荷种源混交林生产力差异与其对光环境的塑性反应能力有关。2个生长阶段,福建建瓯种源幼苗均表现出较高的形态、生理和生物量分配塑性,觅光能力强,因此其干物质量生产能力较浙江龙泉和江西信丰种源幼苗高。随着遮阳处理时间的延长,福建建瓯种源1年生幼苗的Fv/Fm值显著高于浙江龙泉种源和江西信丰种源,但2年生时各种源幼苗的Fv/Fm值均处于0.75~0.85正常范围之内,3个种源间无显著差异,反映出3个木荷种源幼苗在不同发育阶段的不同调整能力。
 DownLoad:
DownLoad: