



-
楸树,泛指梓属(Catalpa Scop.)乔木,包括楸树(Catalpa bungei C. A. Mey.)、灰楸(C. fargesii Bur.)、滇楸(C. fargesii. Bur. f. duclouxii (Dode) Gilmour)及其变种和类型。楸树自然分布于中国华北、西北和西南等地[1]。其材质优良,坚韧致密,具有不翘不裂、耐腐、耐湿、耐磨和不易虫蛀等优点,属我国重要的珍贵用材树种[2]。楸树良种评价和选择工作已开展了20余年,早期的以速生为目的的楸树遗传改良取得了一定的成效[3-4]。如今,为促进楸树新品种和良种选育,其多育种目标改良工作已经开展,如高光效[5-6]和高密度材楸树良种选育[7]。形质改良是材性改良的重要组成部分,在珍贵用材中的地位不亚于生长指标[8]。前人研究表明,尖削度、树皮厚度、分枝角、冠幅等形质性状与木材的出材率密切相关[9-13]。赵阳等[14]采用5年生24个泡桐无性系的4个生长性状(胸径、主干高、总材积、接干高/苗干高)和2个干形性状(主干削度、形数)建立的多性状指数方程选择出了适宜在南方丘陵区栽培的4个优良无性系 。孙晓梅等[15]以12年生落叶松自由授粉家系子代测定林为对象,对生长和形质性状的遗传变异、性状间的相关关系进行研究,并利用选择指数方法对形质性状进行了综合评价,选择出了形质性状优良的家系。由此可见,以形质为目标的林木形质性状改良受到育种研究人员的重视;然而,以形质为主的楸树遗传改良并未开展。
近几十年,随着人民生活水平和文化素质的提高,社会对珍贵用材的需求越来越旺盛,对木材品质要求越来越高。形质改良是提高木材品质的一个重要环节,研究表明,树木的形质性状受中度至较强的遗传控制[16],这意味着形质的遗传改良具有较高的可行性。然而, 不同于生长性状,形质性状的调查更复杂,这方面的育种工作仍处于薄弱阶段。鉴于此,本研究测定了5年生灰楸试验林33个无性系的生长和形质性状,分析了各性状的遗传变异、表型相关和遗传相关,针对不同育种目的利用主成分分析和隶属函数法对灰楸无性系进行综合评定,为灰楸定向培育提供选择策略。
HTML
-
参试的灰楸无性系均种植于甘肃省小陇山林业科学研究所苗圃(105°54′ E,34°28′ N),气候类型属半干旱半湿润气候过渡带,年降水量600800 mm,年平均气温10.7℃,≥10℃积温3 359℃,极端高温39℃,极端低温-19.2℃,无霜期约190 d。
该苗圃地处西秦岭北坡,渭河支流川台区,属典型的黄土高原地貌特征,海拔1 160 m;土壤为黄绵土,其主要性质为:pH值9.07,有机质8.9 g·kg-1,全N 0.93 g·kg-1,速效钾86.77 mg·kg-1,速效磷5.99 mg·kg-1,有效铁1.683 mg·kg-1,有效锌32.665 mg·kg-1,有效锰0.808 mg·kg-1。
-
灰楸试验林为2008年3月初营造,以来自2个优良家系的33个无性系为参试材料,以2个当地推广的楸树良种为对照(表 1),造林苗木为1年生嫁接苗,由小陇山林业科学研究所提供。本试验林营建采用完全随机区组设计,4次重复,每小区4株,株行距2 m×2 m。
来源Source 编号Number 参试无性系
Test clone家系H3-1
Family H3-1H3-1-2、H3-1-3、H3-1-5、H3-1-7、H3-1-8、H3-1-9、H3-1-10、H3-1-15、H3-1-16、H3-1-17、H3-1-18、H3-1-19、H3-1-20、H3-1-21 家系H3-2
Family H3-2H3-2-2、H3-2-3、H3-2-4、H3-2-6、H3-2-8、H3-2-9、H3-2-10、H3-2-11、H3-2-12、H3-2-13、H3-2-15、H3-2-16、H3-2-17、H3-2-19、H3-2-20、H3-2-21、H3-2-22、H3-2-23、H3-2-24 对照无性系
Control clone灰楸H3、楸树9-1 Table 1. Experiment material
-
于2012年年末,进行无性系性状调查。测定各小区中每个单株东西和南北方向的冠幅(单株冠幅以2个方向的平均值代表),伐倒后测定树高(H)、胸径(DBH)。利用国标GB4814-84的方法计算单株材积。分枝度按照等级标准赋值, 具体参见1/2。测定植株最粗侧枝的分枝角度作为单株分枝角。从基部开始,每1 m测定直径,并用生长锥从每个测定直径的部位南向取树皮,用游标卡尺测定树皮厚度。依据李善文等[18]的方法计算平均尖削度。
式中:P11、P2、P3分别代表平均尖削度1、2、3; D0、D2、D4分别代表 0、2、4 m树高的直径; D小头代表的是最高处测得的直径H代表树高。
-
使用Excel2007进行数据分析处理和图表制作。以各性状小区均值为基础,采用SPSS 22. 0对33个参试无性系的树高、胸径、单株材积等10个性状进行方差分析,以检验其是否具有显著差异;其中,遗传参数估算,包括遗传相关、表型相关均使用R语言中的ASReml程序包完成;同时,用R语言中的psych程序包对33个参试无性系进行主成分分析。详细代码及分析步骤见参考1/2。为满足方差分析的独立、正态、方差齐性等条件,对分枝度性状的值经平方根转换后再进行统计分析。
方差分析线性模型为:yij=μ+Ci+Bj+eij
式中:yij为第j个区组第i个无性系的观测值,μ为群体平均效应,Ci为第i个无性系效应,Bi为第j个区组效应,eij为环境误差。
文中具体的参数计算公式如下[20]:
无性系重复力采用公式:R=1-1/F
式中:F为方差分析的F值;
变异系数: CV = S/X×100%
式中:CV为变异系数;S为各性状的标准差;各性状均值。;
表型相关分析采用以下公式:
式中:rp为表型相关系数;σpxpy为性状x与性状y的表型协方差;σpx2为性状x的表型方差;σpy2为性状y的表型方差。
遗传相关分析采用以下公式:
式中:rg为遗传相关系数;σgxgy为性状x与性状y的遗传协方差;σgx2为性状x的遗传方差;σgy2为性状y的遗传方差。
用隶属函数法综合各项指标进行评价[21],隶属函数值计算公式为:
反隶属函数值计算公式为:
式中:i表示某个无性系;j表示某项指标;Tij表示i无性系j指标的隶属函数值;Xij表示i无性系j指标的测定值;Xjmin表示所有无性系j指标的最小值;Xjmax表示所有无性系j指标的最大值。假如某项指标与材积呈负相关,则1减去正相关的隶属函数值就是它的隶属函数值。某一个体某一指标的隶属函数值或反隶属函数值越大,表明该指标越靠近最大值。
2.1. 试验材料
2.2. 性状调查方法
2.3. 数据统计分析方法
-
参试的33个无性系树高、胸径、单株材积等10个性状调查情况(表 2)表明:树高平均6.69 m;胸径平均7.59 cm,最小值仅为4.13 cm,最大值为9.44 cm;单株材积的表型变异系数最大,达34.60%,平均单株材积为0.06 m3。形质性状的表型变异系数均大于15%,其中,平均尖削度P1的变异系数为32.67%。说明5年生灰楸无性系各性状间存在丰富的遗传变异,在无性系水平上具有一定的遗传改良潜力。
性状
Traits平均值
Mean标准差
Standarddeviation表型变异系数/%
Phenotype variable coefficient变幅
Range of Variation树高H/m 6.69 0.90 13.45 3.8~8.71 胸径DBH/cm 7.59 1.10 14.47 4.13~9.44 单株材积V/m3 0.06 0.02 34.60 0.01~0.11 冠幅C/m 2.23 0.42 18.83 1.18~4.78 分枝度BD 1.80 0.43 23.75 1.41~5.75 分枝角BA/(°) 41.51 10.31 24.84 20~65 树皮厚度BT/cm 4.25 0.67 15.76 2.57~7.83 平均尖削度P11 1.69 0.55 32.67 0.63~3.57 平均尖削度P12 1.40 0.34 24.27 0.24~2.37 平均尖削度P13 1.50 0.27 18.08 0.23~2.33 Table 2. Measured value of test clone traits
-
表 3表明:灰楸树高和单株材积在无性系间差异极显著,胸径在无性系间差异显著。树高和单株材积具有中偏上的重复力,达0.592,而胸径具有中等重复力,达0.456。这表明灰楸无性系生长性状遗传变异显著,但遗传控制程度适中。形质性状中,仅平均尖削度P1和P3在无性系间呈显著或极显著差异,其均具有中等大小的重复力,分别为0.479和0.415。冠幅、分枝角、分枝度及树皮厚度在无性系间差异均不显著,且无性系重复力都较小。这意味着灰楸无性系形质性状变异较小,且其遗传控制程度适中,环境因素对灰楸形质性状影响大于生长性状。
变异来源
Variation of source无性系Clone V区组Block 误差Error 自由度df 均方MS F值F value 重复力Repeatability 自由度df 均方MS F值F value 自由度df 均方MS H 32 1.441 2.451** 0.592 3 0.832 1.416 85 0.588 DBH 32 1.790 1.837* 0.456 3 2.085 2.140 85 0.974 V 32 0.001 2.452** 0.592 3 0.000 1.337 85 0.000 C 32 0.184 1.031 0.030 3 0.038 0.213 89 0.178 BD 32 0.182 1.025 0.024 3 0.274 1.538 65 0.178 BA 32 132.826 1.531 0.347 3 363.068 4.185** 75 86.761 BT 32 0.494 1.147 0.128 3 0.226 0.524 79 0.431 P1 32 0.460 1.919 ** 0.479 3 0.555 2.318 85 0.240 P2 32 0.128 1.148 0.129 3 0.043 0.381 84 0.112 P3 32 0.103 1.708* 0.415 3 0.080 1.326 85 0.060 注:**代表P < 0.01,差异极显著;*代表P<0.05,差异显著。下同。
Notes: ** represents P < 0.01, the difference is extremely significant, and * represents P1 < 0.05, the difference is significant. The same below.Table 3. Analysis of variance and estimation of repeatability of Catalpa fargesii clone traits
-
相关分析结果(表 4)表明:灰楸树高、胸径和单株材积的表型相关和遗传相关两两之间均呈极显著正相关。分枝角与其他性状间的遗传和表型相关不显著,说明分枝角性状可能独立于其它性状进行遗传。平均尖削度P1与树高、胸径、单株材积的表型和遗传相关均呈极显著正相关。平均尖削度P13与胸径的表型和遗传相关均呈显著正相关。
H DBH V C BD BA BT P1 P2 P3 H 1 0.786** 0.944** 0.111 0.097 0.087 0.321** 0.203* -0.259** -0.128 DBH 0.901** 1 0.917** 0.165* 0.037 -0.035 0.220** 0.309** 0.052 0.212* V 0.988** 0.943** 1 0.178* 0.046 0.002 0.263** 0.259** -0.125 0.025 C -0.039 -0.123 0.245 1 -0.032 0.291 -0.025 -0.008 0.001 0.129 BD -0.129 -0.255 -0.235 -0.698 1 0.394 0.216* 0.128 0.076 0.062 BA -0.002 0.003 -0.014 0.137 0.064 1 -0.395 -0.071 -0.360 0.200 BT 0.863* 0.847 0.793 0.615 0.825 0.075 1 -0.087 -0.200* -0.126 P1 0.778** 0.777** 0.709** -0.711 0.630 -0.085 0.809 1 0.580** 0.579** P2 -0.053 0.772 0.035 -0.823 0.662 -0.054 0.713 0.863 1 0.700** P3 0.118 0.611* 0.277 0.241 0.844 -0.015 0.993 0.771** 0.940** 1 注:右上角代表遗传相关系数,左下角代表表型相关系数。
Notes: The upper right corner represents the genetic correlation coefficient, and the lower left corner represents the phenotypic correlation coefficient.Table 4. Correlation coefficients and the genetic correlation coefficients of Catalpa fargesii clone traits
-
本研究通过对楸树各无性系生长性状和形质性状的主成分分析,将原来的10个性状降维成相互独立的少数几个能充分反映总体信息的因子, 以累积贡献率达80%为阈值选择出前4个主成分Y1、Y2、Y3、Y4(表 5)。
主成分Main component Y1 Y2 Y3 Y4 H -0.453 -0.295 0.124 DBH -0.492 -0.177 V -0.480 -0.228 -0.152 C -0.740 -0.166 BD -0.144 0.161 0.236 0.745 BA -0.109 -0.554 0.477 BT -0.290 -0.217 0.324 P1 -0.369 0.373 0.143 P2 -0.113 0.598 -0.138 P3 -0.226 0.527 -0.215 特征根Characteristic root 1.881 1.514 1.169 1.091 贡献率Proportion of Variance/% 35.38 22.90 13.67 11.91 累计贡献率Cumulative Proportion/% 35.38 58.27 71.94 83.87 Table 5. Main component analysis of growth traits of Catalpa fargesii clones
表 5表明:Y1主成分具有最大的贡献率35.38%,该主成分中,树高、胸径、单株材积均为负值,但绝对值占比最大,因此,当Y1较小时,树高、胸径等生长量指标大。第2主成分Y2贡献率为22.90%,其中,平均尖削度P11、P2、P3较大,表明第2主成分Y2代表树木的尖削度,Y2越小,尖削度越小。第3主成分Y3贡献率为13.67%,冠幅、分枝角这2个指标均为负值,但绝对值最大,表明Y3越大,冠幅、分枝角越小。第4主成分Y4贡献率为11.91%,分枝度值最大,表明Y4越大,分枝度值越大,树干分叉越少。
-
主成分分析结果表明:Y1主要体现的是生长性状,Y2、Y3、Y4主要体现的是形质指标。根据主成分所代表的生物学意义,同时在实际木材生产中,Y2与出材率具有一定的负相关[22],在此构建参数YZ(YZ=Y3+Y4-Y2)作为形质成分,再结合Y1(生长主成分)进行优良无性系的选择。分别选出Y1和Yz主成分值排名前40%的无性系。图 1中类群A代表在生长方面占优势的无性系;类群B代表形质较优良的无性系;类群C所代表综合性状良好的无性系,不仅生长量大,而且形质较优良。
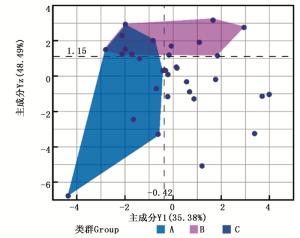
Figure 1. Scatter plot of main component of Catalpa fargesii clones
-
应用隶属函数法对无性系进行综合评价,结果(表 6)表明:33个灰楸无性系中,无性系H3-2-16的10个性状平均隶属函数值(0.667)最高,无性系H3-2-15的平均隶属函数值(0.342)最小。无性系H3-1-18生长(树高、胸径、单株材积)性状的平均隶属函数值最高,为0.945;其次是无性系H3-2-16和H3-1-10,无性系H3-2-15性状的平均隶属函数值(0.041)最小。无性系H3-1-19形质性状的平均隶属函数值最高(0.679),而生长性状平均隶属函数值很低(0.119),综合性状平均隶属函数值排名中等(0.511)。无性系H3-1-9的形质性状平均隶属函数值最低(0.132),而生长性状平均隶属函数值排名第4(0.870),综合性状平均隶属函数值排名倒数第2(0.353)。
综合性状Comprehensive trait 生长性状Growth trait 形质性状Trunk shape 无性系
Clones平均隶属函数值
Average membership function value排序
Order无性系
Clones平均隶属函数值
Average membership function value排序
Order无性系
Clones平均隶属函数值
Average membership function value排序
OrderH3-2-16 0.667 1 H3-1-18 0.945 1 H3-1-19 0.679 1 H3-1-18 0.663 2 H3-2-16 0.905 2 H3-2-8 0.661 2 H3-1-17 0.637 3 H3-1-10 0.904 3 H3-2-9 0.635 3 H3-1-10 0.619 4 H3-1-9 0.870 4 H3-2-3 0.623 4 H3-1-7 0.609 5 H3-1-17 0.853 5 H3-1-16 0.616 5 H3-2-4 0.605 6 H3-1-3 0.729 6 H3-2-12 0.599 6 H3-1-20 0.592 7 H3-1-7 0.688 7 H3-2-4 0.576 7 H3-2-23 0.583 8 H3-2-24 0.675 8 H3-1-7 0.576 8 H3-2-9 0.573 9 H3-2-4 0.672 9 H3-1-20 0.573 9 H3-1-15 0.554 10 H3-2-21 0.636 10 H3-2-16 0.565 10 H3-1-16 0.541 11 H3-1-20 0.635 11 H3-2-23 0.561 11 H3-2-20 0.533 12 H3-2-23 0.635 12 H3-1-15 0.554 12 H3-2-21 0.532 13 H3-2-22 0.623 13 H3-1-17 0.545 13 H3-2-22 0.528 14 H3-1-2 0.563 14 H3-1-18 0.542 14 H3-1-2 0.522 15 H3-1-15 0.554 15 H3-2-20 0.538 15 H3-2-6 0.520 16 H3-2-10 0.549 16 H3-2-6 0.508 16 H3-2-12 0.520 17 H3-1-8 0.548 17 H3-1-2 0.505 17 H3-1-19 0.511 18 H3-2-6 0.547 18 H3-2-13 0.501 18 H3-1-3 0.502 19 H3-2-20 0.521 19 H3-1-10 0.497 19 H3-2-10 0.495 20 H3-2-17 0.473 20 H3-1-21 0.495 20 H3-2-13 0.486 21 H3-1-21 0.459 21 H3-2-19 0.493 21 H3-1-21 0.484 22 H3-2-13 0.451 22 H3-2-21 0.488 22 H3-2-24 0.481 23 H3-2-2 0.444 23 H3-2-22 0.488 23 H3-1-8 0.480 24 H3-2-9 0.430 24 H3-2-17 0.483 24 H3-2-17 0.480 25 H3-1-5 0.386 25 H3-2-10 0.472 25 H3-2-8 0.476 26 H3-1-16 0.369 26 H3-2-15 0.471 26 H3-2-3 0.474 27 H3-2-12 0.335 27 H3-1-5 0.459 27 H3-1-5 0.437 28 H3-2-11 0.300 28 H3-1-8 0.451 28 H3-2-19 0.415 29 H3-2-19 0.232 29 H3-2-11 0.429 29 H3-2-11 0.390 30 H3-2-3 0.125 30 H3-1-3 0.404 30 H3-2-2 0.355 31 H3-1-19 0.119 31 H3-2-24 0.398 31 H3-1-9 0.353 32 H3-2-8 0.046 32 H3-2-2 0.317 32 H3-2-15 0.342 33 H3-2-15 0.041 33 H3-1-9 0.132 33 Table 6. Membership function values of different trait of Catalpa fargesii clones
-
为选育生长量大的无性系,结合主成分分析和隶属函数法筛选出共有无性系H3-1-9、H3-1-18、H3-1-10、H3-1-3、H3-2-16和H3-1-17。从表 7可以看出:与对照灰楸H3相比,筛选出的无性系生长性状都有不同程度的提高,其中,单株材积的增益最大为80%,树高的增益为25.91%,胸径的增益为12.73%。与楸树9-1相比,改良程度较低,单株材积增益为28.57%,树高增益为17.16%。为选育树干分叉少,分支角小,冠幅小,尖削度小的品种,以2种方法筛选出的共有无性系为H3-1-16、H3-1-19、H3-2-12和H3-2-9。表 8显示:与对照无性系相比,筛选出的无性系形质性状都有不同程度的改良,其中,尖削度性状改良效果最佳。
无性系Clone H/m DBH/cm V/m3 优选无性系Optimal clone H3-1-9 7.22±0.46 8.82±0.31 0.09±0.01 H3-1-18 7.64±0.26 8.75±0.66 0.09±0.01 H3-1-10 7.71±0.41 8.46±0.47 0.09±0.01 H3-1-3 7.36±0.47 7.97±0.25 0.08±0.01 H3-2-16 8.01±1.19 8.06±1.18 0.09±0.03 H3-1-17 7.54±0.62 8.39±0.22 0.09±0.01 均值Mean 7.58 8.41 0.09 对照无性系
Control clone灰楸H3 6.02±0.32 7.46±1.37 0.05±0.02 增益Gain/% 25.91 12.73 80.00 楸树9-1 6.47±0.43 8.37±0.22 0.07±0.01 增益Gain/% 17.16 0.48 28.57 Table 7. Growth-improved optimal clone characteristics
性状
Traits优良无性系Optimal clone 对照无性系Control clone H3-1-16 H3-1-19 H3-2-12 H3-2-9 均值Mean 灰楸H3 增益Gain/% 楸树9-1 增益Gain/% C/m 2.02±0.24 1.83±0.30 2.12±0.34 2.25±0.35 2.06 2.31±0.26 -11.04 2.13±0.49 -3.52 BD 1.91±0.15 2.00±0.00 1.65±0.16 1.82±0.15 1.85 1.73±0.00 6.65 1.73±0.00 6.65 BA/(°) 34.25±5.68 40.67±16.77 40.00±14.72 49.33±1.15 41.06 43.75±7.50 -6.14 39.50±9.33 3.96 BT/cm 4.36±0.30 3.83±0.25 4.08±0.44 4.17±0.32 4.11 4.00±0.48 2.75 4.38±0.35 -6.16 P1 1.57±0.42 1.60±0.54 1.49±0.48 1.03±0.39 1.42 2.76±1.83 -48.46 2.35±0.48 -39.47 P12 1.29±0.24 1.20±0.20 1.29±0.13 0.98±0.07 1.19 1.82±0.31 -34.62 2.02±0.30 -41.09 P13 1.50±0.22 1.40±0.15 1.31±0.11 1.23±0.20 1.36 1.58±0.21 -13.92 1.87±0.14 -27.27 Table 8. Shape-modified optimal clone characteristics
为选育兼具高生长和优良形质的无性系,以2种方法选出的共有无性系为H3-1-17、H3-1-10、H3-1-18和H3-2-16(表 9)。与对照灰楸H3相比,单株材积增益最大77.74%,其次是尖削度都有不同程度的改良。与对照楸树9-1相比,尖削度改良得最佳,其次是单株材积(26.96%)。
性状
Traits优良无性系Optimal clone 对照无性系Control clone H3-1-17 H3-1-10 H3-1-18 H3-2-16 均值Mean 灰楸H3 增益Gain/% 楸树9-1 增益Gain/% H/m 7.54±0.62 7.71±0.41 7.64±0.26 8.01±1.19 7.73 6.02±0.32 28.33 6.47±0.43 19.41 DBH/cm 8.39±0.22 8.46±0.47 8.75±0.66 8.06±1.18 8.42 7.46±1.37 12.82 8.37±0.22 0.55 V/m3 0.09±0.01 0.09±0.01 0.09±0.01 0.09±0.03 0.09 0.05±0.02 77.74 0.07±0.01 26.96 C/m 2.10±0.36 2.51±0.41 2.28±0.14 2.28±0.15 2.29 2.31±0.26 -0.80 2.13±0.49 7.59 BD 1.63±0.18 1.80±0.13 1.73±0.00 1.80±0.13 1.74 1.73±0.00 0.52 1.73±0.00 0.52 BA/(°) 35.75±6.50 47.75±15.71 28.75±6.50 40.50±8.23 38.19 43.75±7.50 -12.71 39.50±9.33 -3.32 BT/cm 4.45±0.68 4.80±0.58 4.62±1.03 3.82±0.89 4.42 4.00±0.48 10.56 4.38±0.35 0.97 P11 1.63±0.72 1.29±0.32 2.15±0.97 2.03±1.03 1.77 2.76±1.83 -35.73 2.35±0.48 -24.52 P12 1.41±0.41 1.09±0.31 1.25±0.26 1.23±0.24 1.25 1.82±0.31 -31.54 2.02±0.30 -38.32 P13 1.46±0.30 1.40±0.13 1.45±0.14 1.34±0.11 1.41 1.58±0.21 -10.62 1.87±0.14 -24.48 Table 9. Comprehensive selection of optimal clone characteristics
3.1. 灰楸无性系生长性状和形质性状的变异分析
3.2. 灰楸无性系各性状方差分析及重复力
3.3. 灰楸无性系各性状的相关分析
3.4. 灰楸无性系各性状主成分分析
3.5. 灰楸优良无性系选择方法
3.5.1. 主成分分析法评价结果
3.5.2. 隶属函数法评价结果
3.6. 灰楸优良无性系选择
-
遗传变异是选择的基础,充分了解林木的变异才能准确地评估改良的可靠性。本研究通过对5年生33个灰楸无性系生长进行统计分析发现,树高和单株材积在无性系间差异极显著,胸径在无性系间差异显著,说明在无性系水平具有较大的遗传改良潜力。生长性状重复力估算结果表明,树高、胸径、单株材积受中等的遗传控制(0.456~0.592)。早期研究表明,5年生林木无性系间的生长性状存在显著差异[23],且受到中偏上的遗传控制(0.53~0.78)[24-25], 证明了5年生无性系生长遗传改良具有可行性。
形质性状遗传改良是获得优良形质的主要途径,而形质改良是林木改良的重要部分,优良的形质不仅可以保证林木的良好生长,而且还可以提高木材的材质和出材率[12]。本研究的灰楸形质性状中,平均尖削度P1和P3无性系间存在显著或极显著的差异,且具有中等大小的重复力(0.479和0.415),同样具有一定的改良基础。遗传相关分析结果显示,单株材积与冠幅、树皮厚度均存在显著或极显著较弱的正相关,相关系数分别为0.178、0.263,说明生长性状与形质性状具有独立性,可进行单独定向选择[26-27]。
生长性状和形质性状的平均隶属函数值排名结果表明,无性系在这两方面各有侧重,形质性状平均隶属函数值高的无性系,生长性状平均隶属函数值不一定高,如无性系H3-1-19,因此,可以根据育种目的选择出不同的育种群体。相关分析和评价结果说明,若对灰楸的形质性状进行选择改良,对生长性状的影响极小。然而,这与前人研究结果不一致,如日本落叶松的冠幅、皮厚与生长性状之间存在着中等程度的正相关[15];泡桐的削度与胸径、主干高、接干高/苗干高的遗传正相关关系均达极显著,如果过分强调泡桐生长性状的改良,则有导致干形性状下降的的趋势[14]。
在林木育种过程中,没有任何一个良种能适用于一切用途。林木生长性状的优良与否是保证木材产量的关键因素。另外,对原木加工来说,更需要林木干形通直、圆满、大径级及无节疤等形质性状优良[28]。鉴于此,本研究良种选育了以生长性状为主,形质为辅;以形质为主,生长为辅及生长和形质兼具的多个育种方向的不同类型无性系。
-
5年生灰楸无性系间各个性状存在丰富的遗传变异,且生长性状受到中等的遗传控制,有较好的遗传改良潜力。灰楸无性系生长性状与形质性状具有独立性,可进行单独定向选择。灰楸无性系H3-1-9、H3-1-18、H3-1-10、H3-1-3、H3-2-16和H3-1-17可作为速生丰产良种;H3-1-16、H3-1-19、H3-2-12和H3-2-9可作为优质用材的无性系;无性系H3-1-17、H3-1-10、H3-1-18和H3-2-16可考虑作为生长和形质综合改良的灰楸无性系在当地推广。
 DownLoad:
DownLoad: