


-
树干液流是树木蒸腾耗水的一个重要参数,可以反映出植物生理特性和环境因素对树木水分利用的综合调节作用[1-2]。然而,树干液流受到生物因素(如树种和树形)以及环境要素(如地域、时间、气象因子和土壤含水量)的影响较大,导致关于树干液流与气象因子(例如日照、温度、湿度、气压、风速、降雨)之间关系的研究得出了不同的结论。一般而言,太阳辐射、饱和水汽压差、气温和湿度被认为是驱动树木蒸腾的主要气象因子。其中,树干液流与太阳辐射、饱和水汽压差和气温呈正相关,与湿度呈负相关[3-12]。目前对于影响树干液流的气象因子的主次顺序存在争论:买尔当·克依木等[13]研究认为太阳辐射是影响植物光合作用和蒸腾作用的主要外界因子,在其直接或间接影响下气温、湿度才发生变化;孙旭等[14]研究认为太阳辐射、饱和水汽压亏缺是影响日尺度油松蒸腾速率的主要因子;张雷等[15-16]研究认为影响树干液流的主控气象因子因区域、时间和物种不同而有差异。另外,由于研究的时间、地点、对象和方法的不同,树干液流与气象因子之间时滞长短也存在差异[17-22]。目前相关研究主要是针对单一或少数几种树种进行分析[23],如松树[3, 14, 24]、杉树[10]、杨树[18, 20]、相思树[5]、核桃树[17]、侧柏[6]和刺槐[25]等。而不同树种的蒸腾耗水能力不一,这可能是上述研究结果不一致的原因。因此,本文选择了哈尼梯田水源区的3种林分中的10种乔木树种作为研究对象,以消除树种因素的限制。从生理生态层面上研究乔木树干液流与气象因子的关系,对进一步研究乔木的蒸腾作用具有重要意义。
-
全福庄小流域位于云南省红河州元阳县新街镇全福庄小寨(图1a),是元阳哈尼梯田的核心区域,是哈尼梯田“森林-村庄-梯田-河流”垂直景观结构[26]的典型代表,占地面积约为76.87 hm2。研究区气候属亚热带山地季风气候,干湿季分明,平均气温16.6 ℃,年平均降雨量约为1 398 mm,其中约78%的降水发生在雨季(5月至10月),11月至次年4月为旱季,3月至10月为植被生长季,11月至2月为非生长季。年平均日照时数1 550 h,相对湿度85%,土壤以黄棕壤为主,土壤剖面较为完整,土层大约有100 cm[27]。哈尼梯田水源区森林分布主要有原始林和次生林,本文在全福庄小流域水源区选取了3个植被类型具有代表性的试验点(图1a,表1)。
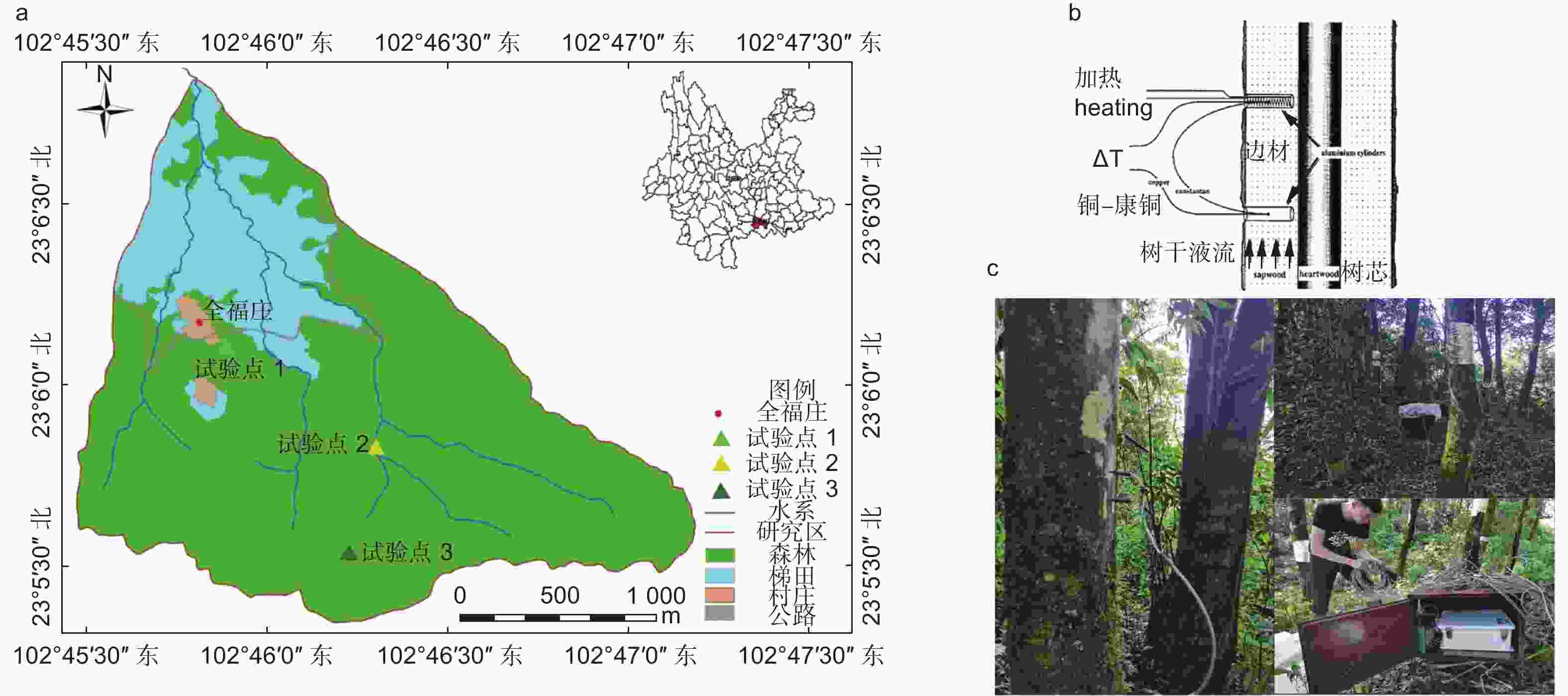
图 1 研究区位置及Granier热扩散测量树干液流速率的原理[28]与现场测定
Figure 1. Schematic diagram of study area location and Granier thermal diffusion measurement of SAP flow rate
表 1 树干液流速率监测样木基本参数
Table 1. Basic parameters of sample wood were monitored by SAP flow rate
试验点
Study sites林分类型
Forest type样树编号
Sample tree number胸径
DBH/cm树高
HT/m冠幅(东西,南北)
RTC/(m*m)试验点1
Study plot1次生常绿阔叶林
Secondary evergreen broad-leaved forest液1-尖齿木荷1 Schima khasiana 10.7 11.5 2.1,1.6 液1-尖齿木荷2 Schima khasiana 10.8 13.5 1.8,1.5 液1-元江锥1 Castanopsis orthacantha 11.3 14.5 2.0,1.4 液1-元江锥2 Castanopsis orthacantha 11.2 15.0 2.4,1.6 液1-喜树1 Camptotheca acuminata 12.5 13.0 1.8,1.6 液1-喜树2 Camptotheca acuminata 11.5 12.2 1.6,1.5 试验点2
Study plot2次生落叶阔叶林
Secondary deciduous broad-leaved forest液2-桤木1 Alnus cremastogyne 33.5 23.0 6.3,4.2 液2-桤木2 Alnus cremastogyne 25.2 20.3 4.2,3.2 液2-桤木3 Alnus cremastogyne 24.8 20.2 4.2,1.0 液2-喜树3 Camptotheca acuminata 15.3 15.6 2.1,2.0 液2-喜树4 Camptotheca acuminata 28.4 22.1 4.5,4.7 试验点3
Study plot3原生常绿阔叶林
Native evergreen broad-leaved forest液3-云南樟1 Cinnamomum glanduliferum 22.0 17.2 4.2,3.1 液3-云南樟2 Cinnamomum glanduliferum 24.0 18.1 4.5,3.4 液3-西南红山茶1 Camellia pitardii 18.7 14.5 4.2,4.1 液3-西南红山茶2 Camellia pitardii 20.0 15.2 5.7,4.6 液3-西南红山茶3 Camellia pitardii 31.0 18.3 6.9,5.1 液3-刺栲1 Castanopsis hystrix 48.0 25.0 6.2,5.3 液3-刺栲2 Castanopsis hystrix 12.5 13.2 2.3,1.5 液3-猴樟1 Cinnamomum bodinieri 27.8 22.3 5.3,4.2 液3-猴樟2 Cinnamomum bodinieri 23.0 19.2 4.2,3.6 液3-三桠苦 Evodia lepta 31.0 25.3 7.3,6.5 注:DBH为胸径,HT为树高,RTC为冠幅
Notes: DBH, diameter at breast height; HT, height of tree; RTC, range of tree-crown -
Granier 热扩散探针安装在样树树干离地面1.3 m处,监测期间使用两块12 v蓄电池轮流为其持续供电,每10 s 读取 1 次数据,每30 min 记录1 次平均值,定期维护仪器并下载数据(图1c)。Granier热扩散探头[28]由两根探针组成,上部探针内含有加热器和热电偶,可以恒定连续加热;下部探针只有热电偶,作为参考端(图1b)。通过测定两根探针在边材的温差值计算液流速率。当液流速率等于0时,两根探针的温差最大,当液流速率增大,温差值减小。液流速率计算公式:
$ J_s=3\;600 \times 0.011\;9 \times K^{1.231} $
(1) $ k=\frac{\Delta T_m-\Delta T}{\Delta T} $
(2) 式中:Js 为液流速率/(mL·cm−2·h−1);∆Tm为日尺度所记录的最大温度差值/℃;∆T为上下探头之间瞬时的温度差值/℃。
-
2020年8月至2021年9月,采用 Granier热扩散探针技术对哈尼梯田水源区次生常绿阔叶林、次生落叶阔叶林、原生常绿阔叶林的测验样树(表1)进行树干液流速率测定。在保证所有实验设备正常工作的前提下,选择供研究的同步异常数据。本文根据树干液流速率呈现“昼高夜低”的单峰或双峰曲线的特点,先找出每棵样树在所有观测期间正常的最高峰值,再把非峰值时出现的高于此值的间断性跳跃数据作为每棵样树的异常数据。在同一观测时间,所有监测样树都出现了“异常数据”的树干液流异常数据才最终作为同步异常数据,所有监测样树都满足了相应条件的数据作为同步数据。
-
在试验区林外空旷地安装一台WertherHawk-232自动气象站(WertherHawk,美国)记录气象条件,自计频率为5 min,测定指标包括空气气温(Ta)/℃、空气相对湿度(RH)/%、风速(SP)/(m·s−1)、大气压(P)/kPa、降雨量(Pn)/mm和太阳辐射(Rn)/(W·m−2),饱和水汽压差(VPD)/kPa[29]通过计算求得,计算公式如下:
$ V P D=(1-R H) \times 0.611 \mathrm{e}^{[17 \cdot 502 T_a /(T_a + 240.97)]} $
(3) -
$ \text { 变化幅度 }=\text { (本期数 }- \text { 上期数 }) / \text { 上期数 } \times 100 {\text{%}} $
(4) -
使用Origin 2018和photoshop 2022软件对数据进行计算和绘图;使用SPSS 25.0进行各树种树干液流与气象因子的Pearson相关性分析,p<0.01表示极显著相关,p< 0.05表示显著相关;采用错位相关法分析各月树干液流与气象因子的Pearson相关系数;使用Excel 2010筛选功能找出增幅超过100%的77个上升同步数据和增幅小于−50%的63个下降同步数据。
-
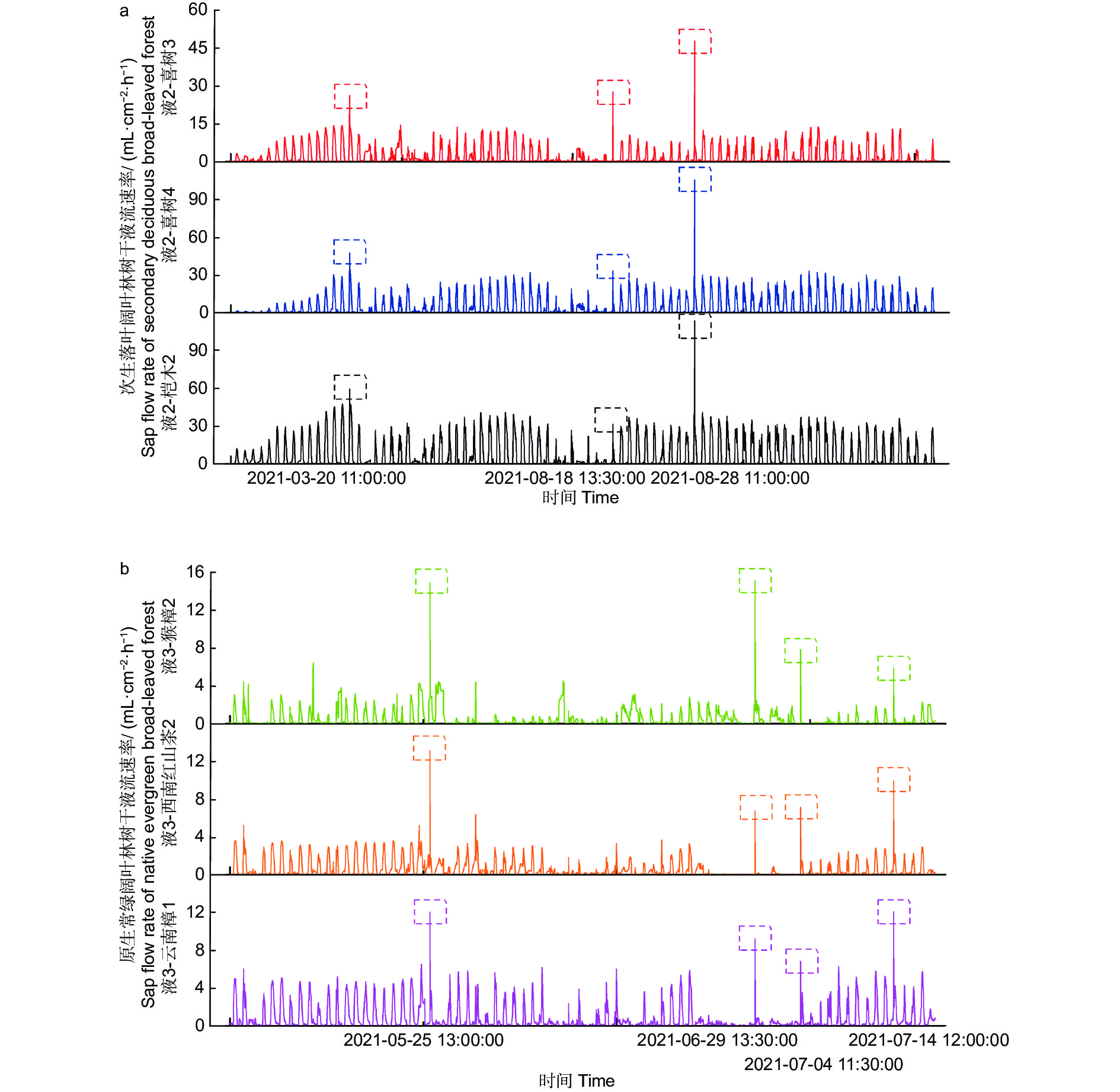
通过分析监测期间树干液流,2021年3月至9月次生落叶林和原生常绿阔叶林的树干液流数据中发现7个同步异常数据(图2)。在试验点2次生落叶阔叶林树干液流速率监测中,样树液2-桤木2、液2-喜树3、液2-喜树4最大的正常峰值分别为47.25 mL·cm−2·h−1、13.33 mL·cm−2·h−1和32.9 mL·cm−2·h−1,非峰值时间出现高于最大正常峰值的同步异常数据有3条,它们分别出现在3月20日11:00,8月18日13:30,8月28日11:00(图2a)。次生落叶阔叶林中发现的这3条同步异常数据,在不同树种(桤木或喜树)、不同径级、不同树形,相同环境条件下都表现出极为明显的时间同步性。在试验点3原生常绿阔叶林中,样树液3-云南樟1、液3-西南红山茶2、液3-猴樟2最大的正常峰值分别为 12.48 mL·cm−2·h−1、7.14 mL·cm−2·h−1和8.36 mL·cm−2·h−1,每棵监测样树非峰值时间出现高于最大正常峰值的数量和出现时间不同。能够满足同步异常条件的数据有4条,分别出现在5月25日13:00、6月29日13:30、7月4日11:30和7月14日12:00(图2b)。原生常绿阔叶林中找出的这4条同步异常数据,也表现出明显的同步一致性。利用这7个同步异常数据分析该地区树干液流与气象因素的关系,可以消除树种因素对其的影响,反映一个或多个气象因素引起的树干液流的变化。

图 2 次生落叶阔叶林和原生常绿阔叶林出现的7个同步异常数据
Figure 2. Seven synchronous anomalies in secondary deciduous broad-leaved forest and primary evergreen broad-leaved forest
-
为了探究引起树干液流同步异常的气象原因,图3罗列出了7条同步异常数据出现日对应的太阳辐射、气温、湿度、饱和水汽压差等主要气象因子从6:00至18:00的变化和增幅。7月14日12:00当太阳辐射快速升高到达最大值时,气温也随之快速升高,湿度快速减少,饱和水汽压差变化幅度最大,所以树干液流在此时出现了同步异常数据。5月25日13:00当太阳辐射值接近最大值时,气温也接近最高值,湿度接近最小值,饱和水汽压差接近最大值,此时样树树干液流出现了同步异常数据。3月20日、8月28日11:00当太阳辐射快速升高到达第一次最大值时,尽管气温没有快速升高,湿度没有快速减少,树干液流在此时出现了同步异常数据。6月29日、8月18日13:00太阳辐射值到达最大值后快速下降,但是气温、湿度和饱和水汽压差变化不大,同步异常数据出现在30 min后的13:30。7月4日11:00太阳辐射快速升高,气温也随之快速升高,湿度快速减少,饱和水汽压差快速变大,同步异常数据出现在太阳辐射增幅最大后30 min的11:30。如图3a,5月25日、7月14日同步异常数据出现在全日太阳辐射最大的点上,3月20日、8月28日同步异常数据出现在太阳辐射快速升高到达第一次最大的点上,6月29日、8月18日同步异常数据出现在太阳辐射最大后的30 min,7月4日同步异常数据出现在太阳辐射增幅最大后30 min。如图3b,5月25日、6月29日和8月18日异常数据出现在气温接近最高的中午13:00至13:30,7月14日异常数据出现在气温增幅最大的点上,3月20日、7月4日和8月28日异常数据出现在气温增幅最大前的30~60 min。如图3c,7月14日异常数据出现在湿度变化幅度最大的点上,8月18日同步异常数据出现在湿度最小的30 min前,3月20日、7月4日和8月28日同步异常数据出现在湿度变化幅度最大的30~60 min前。如图3d,5月25日异常数据出现在饱和水汽压差值接近最大值的点上,7月14日异常数据出现在饱和水汽压差变化幅度最大的点上,6月29日和8月18日同步异常数据出现在饱和水汽压差最大的30 min前,3月20日、7月4日和8月28日同步异常数据出现在饱和水汽压差变化幅度最大的30~60 min前。从以上分析可以看出,气象因子的瞬时大变化是产生同步异常数据的原因,而且太阳辐射是产生同步异常树干液流的直接驱动力,同步异常数据容易出现在太阳辐射最大或增幅最大后的0~30 min,气温、湿度和饱和水汽压差最大或者增幅最大前0~60 min,而且气温、湿度和饱和水汽压差三者表现出高度的一致性。

图 3 同步异常树干液流与气象因子关系分析
Figure 3. Relationship analysis diagram of synchronous abnormal SAP flow and meteorological factors
-
根据7个同步异常数据,气象因子的瞬时变化会改变树干液流速率。本文进一步分析了次生落叶林和原生常绿阔叶林在监测期内每日6:00至18:00树干液流的增幅情况,如表2所示。不同树种的液流变化幅度不同,但大部分液流增幅都在−50%~50%之间,约占78%,增幅大于100%的约占9%,增幅小于−50%的约占7%。树干液流增幅超过100%和低于−50%可能是由于某些气象因素的突变,导致树干液流的起伏。本文从这些数据中找出增幅超过100%的77个同步数据作为液流快速上升的代表,增幅小于−50%的63个同步数据作为液流快速下降的代表。
表 2 样树树干液流变化幅度最大值和频次
Table 2. Table of frequency and maximum values of trunk sap flow variation of sample trees
样树编号
Sample tree number变化幅度频次 Frequency of change amplitude 最大值/%
MaximumR<−50% −50%≤R<0 0%≤R<50% 50%≤R<100% 100%≤R 液2-桤木2 Alnus cremastogyne 109 822 862 132 224 1 568 液2-喜树4 Camptotheca acuminata 100 890 817 120 222 2 355 液2-喜树3 Camptotheca acuminata 194 740 865 132 218 9 632 液3-云南樟1 Cinnamomum glanduliferum 163 692 723 128 194 2 750 液3-西南红山茶2 Camellia pitardii 167 717 856 45 115 6 520 液3-猴樟2 Cinnamomum bodinieri 164 701 819 87 129 55 127 注:R表示每日6:00至18:00时间内每30 min的变化幅度
Note: R represents the range of change every 30 minutes between 6:00 and 18:00 per day -
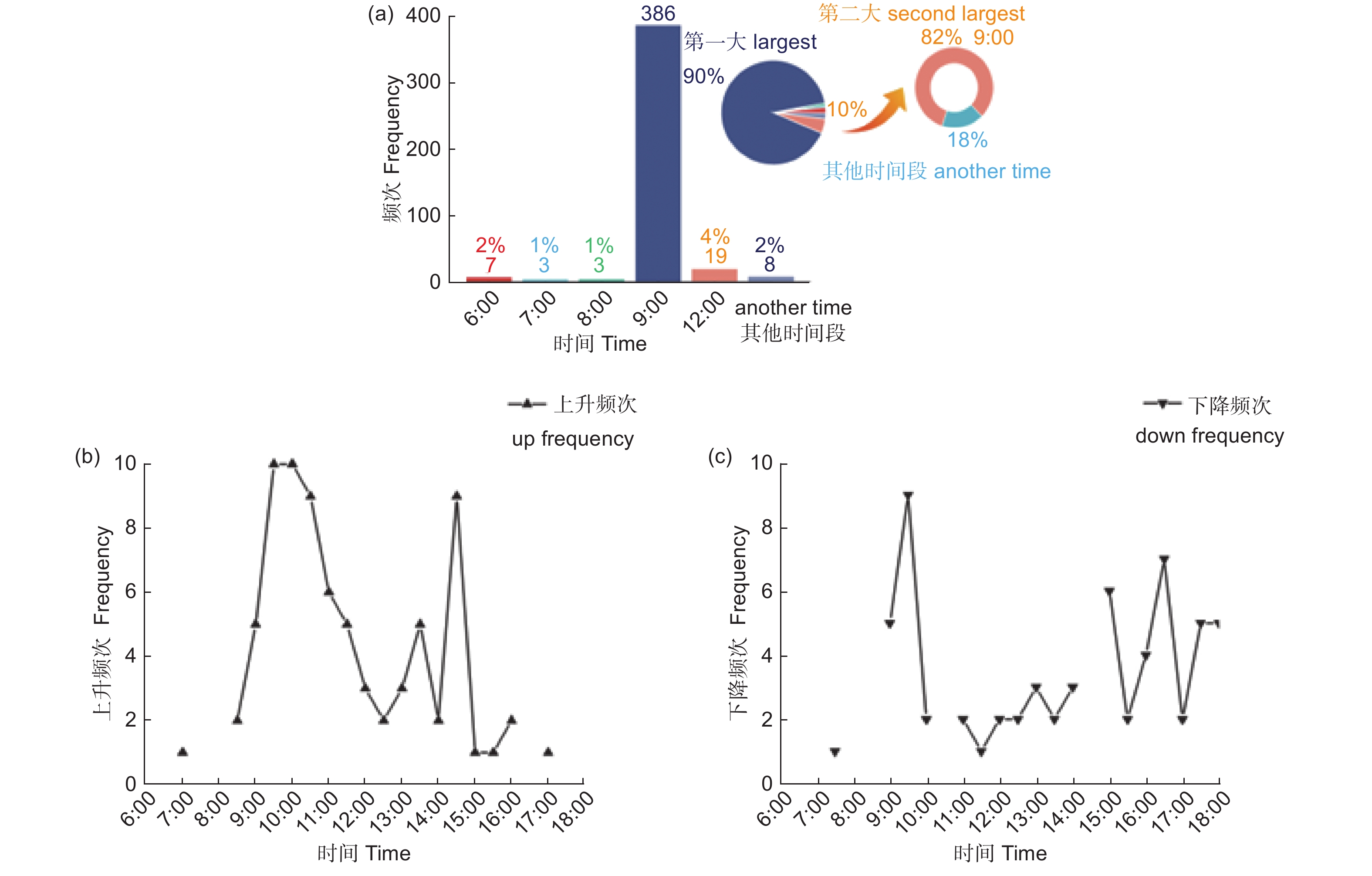
对试验区2020年8月至2021年9月的太阳辐射、气温、湿度和饱和水汽压差进行1 h为尺度监测,分析饱和水汽压差增幅时,发现了典型的“9点现象”,如图4a所示。饱和水汽压差90%增幅最大出现在9:00 am,其余10%的第二大82%也出现在9:00 am,所以饱和水汽压差最大增幅几乎出现在每天9:00 am,本文将这一现象称为“9点现象”。由于研究地点经纬度的原因,太阳辐射、气温和湿度等增幅出现典型的自然现象,太阳辐射增幅最大的时间出现在6:00 am,气温增幅最大出现在9:00 am,湿度增幅最大出现在 5:00 pm。液流上升增幅超过100%的77个同步数据多数发生在9:00至15:00,而且9:00至10:00和14:00至15:00两个高发期,如图4b所示。液流下降增幅超过50%的63个同步数据多数发生在9:00至18:00,而且9:00至10:00和15:00至18:00两个高发期,如图4c所示。

图 4 饱和水汽压差和同步数据各时间段分布
Figure 4. Distribution of saturated water vapour differential pressure and synchronous data by time period
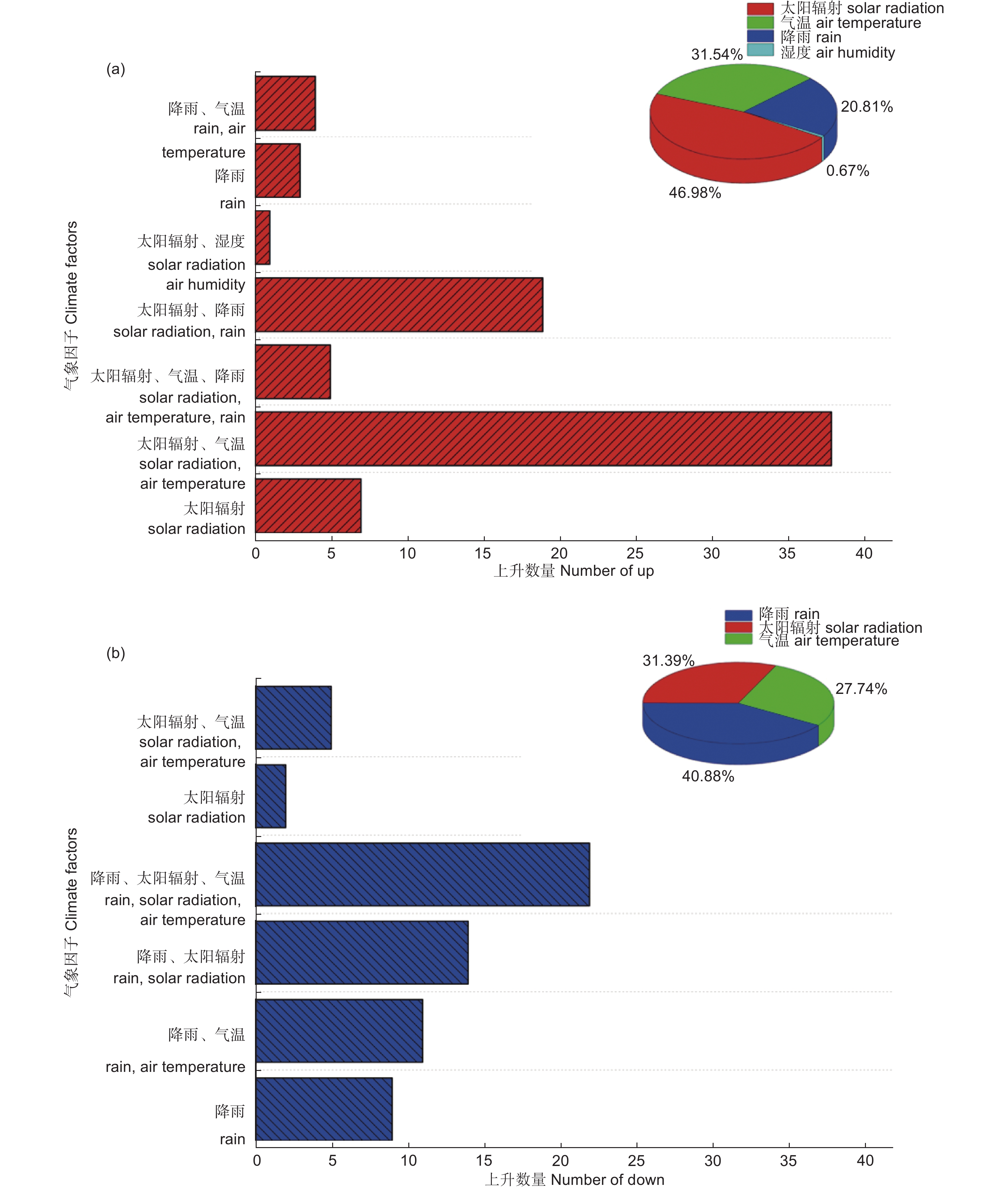
在气象数据中找到增幅超过100%的77个和减幅超过50%的63个同步数据发生时间对应的时间点,每个点对太阳辐射、气温、饱和水汽压差、湿度、风速、大气压、降雨等气象因子进行前后各2 h的分析,如果在分析时间内某个或多个气象因子发生了突变,将它们作为影响原因进行统计。如图5a所示,太阳辐射、气温和降雨是产生上升同步数据的原因,太阳辐射的突然增加是液流增幅超过100%的直接原因,77条数据中有70条的首要原因是太阳辐射的突然增加。但是影响树干液流快速上升的气象因子不是单一的,太阳辐射单一因子影响上升同步数据的只有7条,43条是由于太阳辐射的突然增加而导致气温的快速升高,19条是由于短时间内降雨后太阳辐射突然增加。如图5b所示,降雨、太阳辐射和气温是产生下降同步数据的原因,降雨是液流降幅超过50%的直接原因,63条数据中有56条的首要原因是降雨,7条是由于太阳辐射的快速减少。但是影响树干液流快速下降的气象因子也不是单一的,降雨单一因子影响下降同步数据的只有9条,36条是由于降雨而太阳辐射快速变化,11条是由于降雨而导致气温快速变化。以上分析可知,太阳辐射的突然增加是产生液流上升同步数据的原因,而降雨则是产生液流下降同步数据的原因,但是影响液流同步上升或下降的气象因子不是单一的,多数情况下是由于太阳辐射、降雨等主要因子发生变化,导致其它一个或多个因子变化,从而影响树干液流的变化。
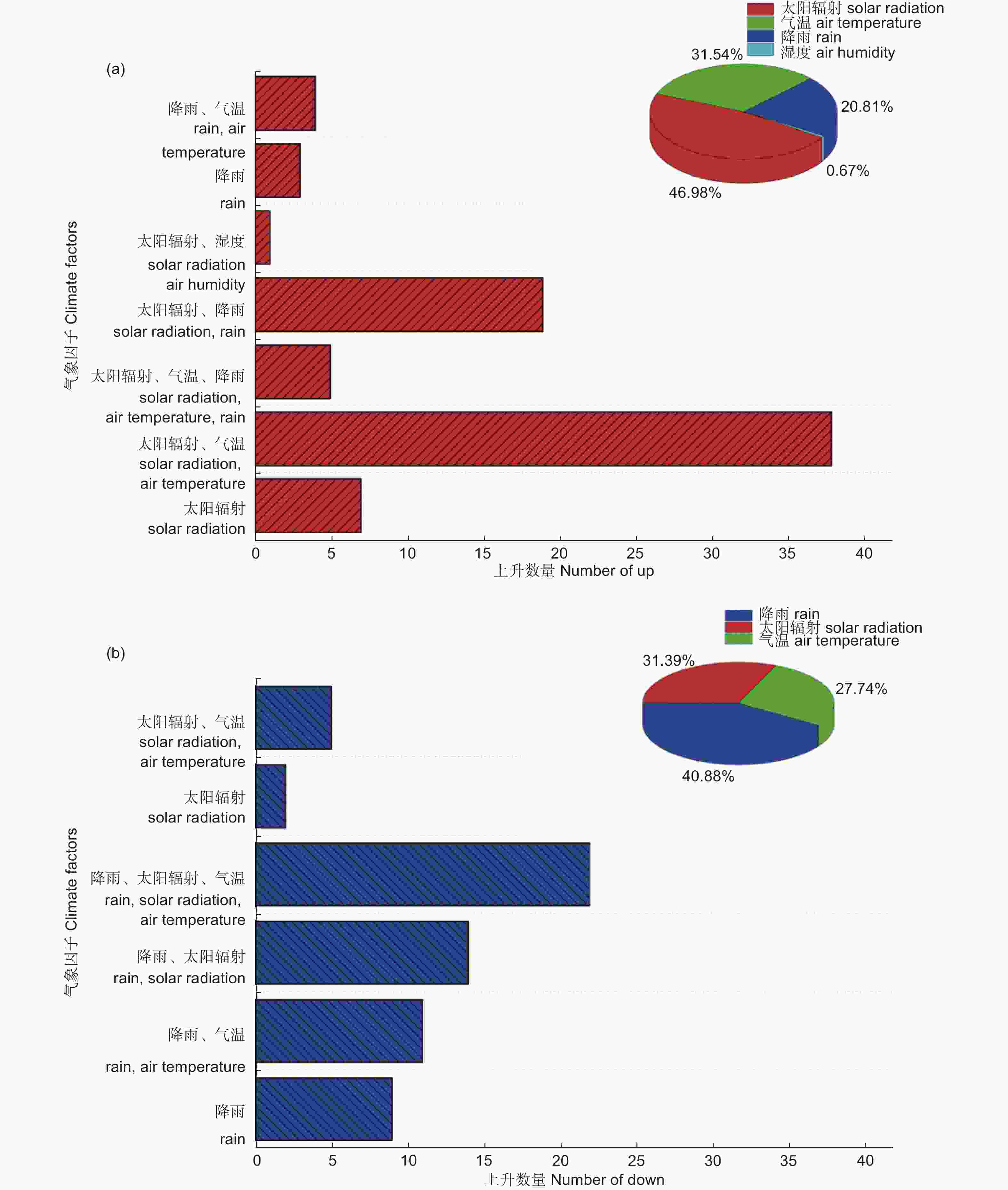
图 5 影响同步树干液流的气象原因分析
Figure 5. Analysis of the meteorological causes affecting synchronous trunk sap flow
-
3种林分类型优势树种正常液流速均呈“昼高夜低”日变化规律,随着每天日出和气温上升,液流上午启动,中午达到峰值后开始下降(单峰曲线),而部分树种液流又返回另一峰值(双峰曲线),每天下午六七点钟几乎为零,很少有其他时刻出现超过峰值的异常数据。相同树种各月树干液流速率的变化趋势非常相似,不同树种各月液流速峰值大小不同,有的树种不同径级树干液流速差异较大,而有的差异却较小。在典型晴天条件下,不同时期不同树种液流启动和停止时间也不相同,但同一树种的变化规律却非常相似,所有树种液流启动并到达峰值的时间从3月春季开始逐渐提前,至夏季最早后,到秋季、冬季再逐渐推迟,相同树种在相同时期液流速率大小呈晴天>阴天>雨天的规律。树干液流与树种、树形、环境因子和气象因子密切相关,对各月份气象因子和不同树种的树干液流速率进行了Pearson相关性分析,如表3所示。结果表明研究区各树种生长季树干液流与太阳辐射、气温、饱和水汽压差、湿度极显著相关,与风速和大气压在多数月份呈显著或极显著相关,与降雨量相关性不强。树干液流速率与太阳辐射、气温、饱和水汽压差和风速呈正相关,而与湿度和大气压呈负相关。同一树种不同月份由于环境原因与各气象因子的相关强弱不同[30],例如喜树3月、7月、8月、9月树干液流与太阳辐射的相关系数分别为0.69、0.75、0.84和0.77;不同树种同一月份由于树种原因与气象因子的相关强弱也不同,例如云南樟、西南红山茶、猴樟、桤木、喜树7月树干液流与饱和水汽压差的相关系数分别为0.66、0.60、0.25、0.44和0.71。影响树干液流的气象因子主要是太阳辐射、饱和水汽压差、气温和湿度,但是与气象因子的相关性大小由不同树种决定。这与王小菲[5, 31]、于占辉[6]、张璇[9]、涂洁和万艳芳[32]研究结论一致。
表 3 各树种生长季树干液流速率与各气象因子的Pearson相关系数分析
Table 3. Pearson correlation coefficient between SAP flow rate of tree species and meteorological factors in growing season
树种
Varieties of trees月份
Month太阳辐射
Rn气温
Ta饱和水汽压差
VPD湿度
RH风速
SP大气压
P降雨量
Pn云南樟
Cinnamomum glanduliferum5月 0.70** 0.68** 0.74** −0.71** 0.48** −0.2** 0.08* 6月 0.51** 0.57** 0.59** −0.54** 0.26** −0.19* −0.02 7月 0.63** 0.63** 0.66** −0.64** 0.25** −0.06 −0.03 西南红山茶
Camellia pitardii5月 0.68** 0.62** 0.64** −0.59** 0.43** −0.15* 0.14** 6月 0.50** 0.45** 0.49** −0.45** 0.19* −0.027 −0.07 7月 0.59** 0.57** 0.60** −0.57** 0.21** −0.03 −0.01 猴樟
Cinnamomum bodinieri5月 0.33** 0.29** 0.28** −0.26** 0.11 −0.03 0.08* 6月 0.20** 0.19** 0.15** −0.15** 0.12* −0.09 0.02 7月 0.28** 0.25** 0.25** −0.21** 0.06 −0.26** 0.17** 三桠苦
Evodia lepta5月 0.23** 0.17** 0.22** −0.23** 0.14** 0.04 −0.01 6月 0.34** 0.46** 0.45** −0.41** 0.18** −0.14* −0.01 7月 0.30** 0.33** 0.33** −0.33** 0.27** 0.10 0.28** 桤木
Alnus cremastogyne3月 0.63** 0.66** 0.66** −0.55** 0.60** −0.54** −0.02 7月 0.45** 0.46** 0.44** −0.41** 0.12* 0.44** 0.20** 8月 0.58** 0.67** 0.72** −0.68** 0.02 −0.32** 0.02 9月 0.52** 0.63** 0.62** −0.56** 0.23** −0.45** 0.06 喜树
Camptotheca acuminata3月 0.69** 0.62** 0.58** −0.46** 0.61** −0.34** 0.08 7月 0.75** 0.73** 0.71** −0.69** 0.17* −0.33** 0.11* 8月 0.84** 0.79** 0.80** −0.79** 0.12* −0.18* 0.01 9月 0.77** 0.74** 0.69** −0.64** 0.27** 0.28** 0.05 注:**表示在0.01水平上极显著相关,*表示在0.05水平上显著相关
Notes: ** shows correlation is significant at the 0.01 level, * shows correlation is significant at the 0.05 level通常,由于气象因子相对稳定,在非峰值时间很少出现超过峰值的异常数据,但某个或多个气象因子发生瞬态性的大变化,树干液流瞬态响应。根据上文的分析,产生7个同步异常的主要原因是太阳辐射发生了瞬时的大变化,在11:00至13:30接近中午最热期间太阳辐射的快速异常是产生同步异常数据的直接原因,所以太阳辐射是影响同步异常树干液流的直接驱动力。太阳辐射又直接或者间接影响了气温和湿度[13],气温和湿度又决定了饱和水汽压差,所以影响树干液流的太阳辐射、饱和水汽压差、气温和湿度四个气象因子中,太阳辐射是主要原因,饱和水汽压差、气温和湿度是次要原因。
-
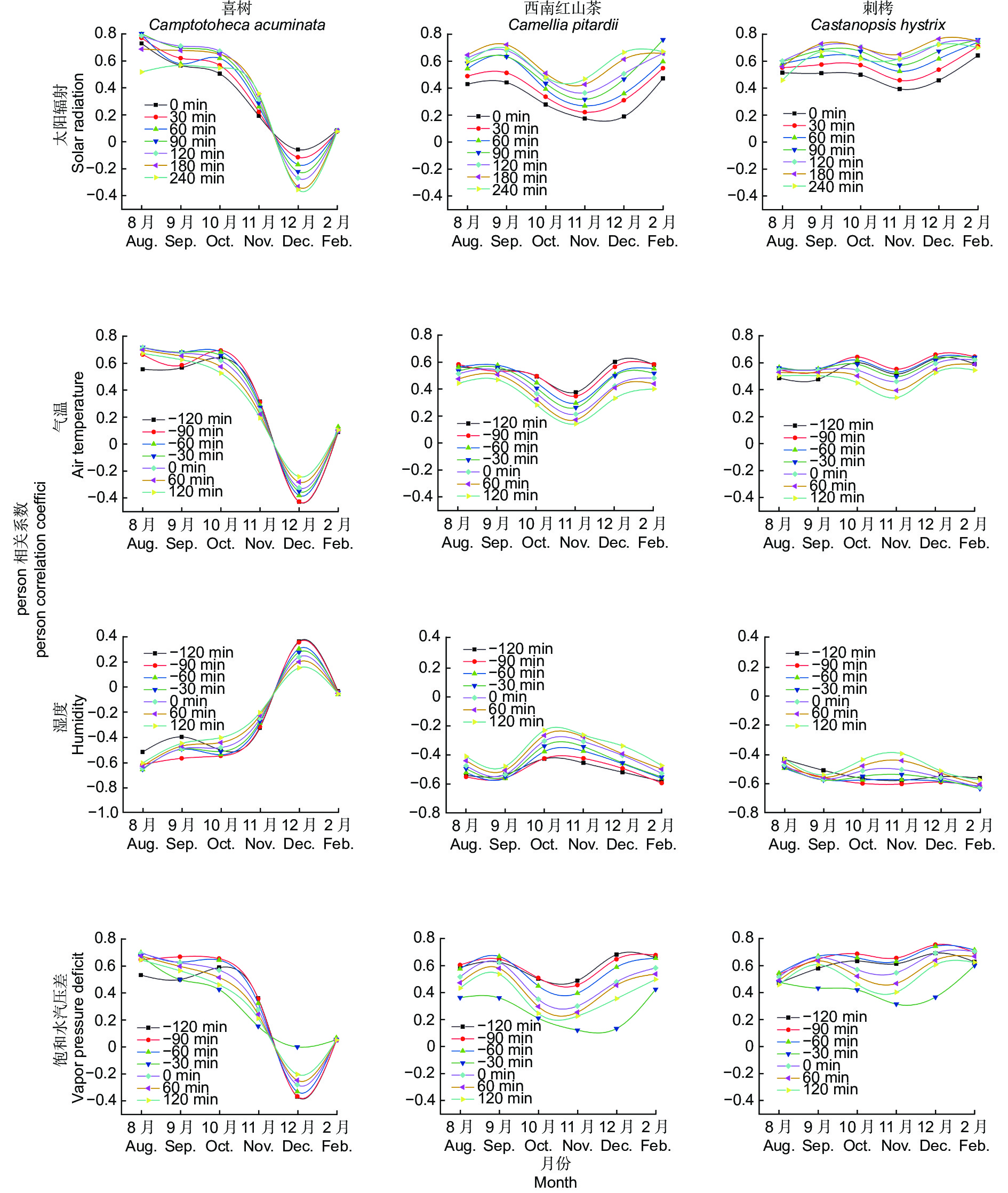
树干液流与气象因子具有一定的时滞效应,本文以喜树、西南红山茶、刺栲为例,采用错位相关法分析各月树干液流与气象因子的Pearson相关系数,分析2020年8月至2021年2月3种树各月树干液流和气象因子的时滞效应。如图6所示,原生常绿阔叶林不同树种的树干液流滞后太阳辐射90~240 min,提前气温60~120 min,提前湿度90~120 min,提前于饱和水汽压差0~120 min。这与其他学者[24, 33-36]的研究结果基本一致,树干液流滞后于太阳辐射,提前于气温、饱和水汽压差、湿度和风速,但是滞后和提前时间不同,可能是地域、时间、物种和环境等原因造成时滞时间不同[21, 33, 37-40]。党宏忠等[20]研究得出新疆杨在不同年份同一时期的气象因子与树干液流的时滞时间都不一样,马长明等[8]研究得出不同健康状况的北京杨的时滞时间也不一样。本文由同步异常数据分析可知,所有监测树种的树干液流滞后太阳辐射0~30 min,提前于饱和水汽压差、气温和湿度0~60 min。这一结果与马长明[8]、武鹏飞[18]、涂洁[41]等研究结果非常接近。用同步异常数据分析得出的太阳辐射、饱和水汽压差、气温和湿度滞后或提前时间远小于正常数据。一般情况下,因为不同树种内部结构和生理特性不一样、地域和时间不一样,气象因子无法100%满足的所有树种树干液流发生同步异常,所以会出现时滞时间远大于异常数据。但是本文的7个同步异常数据是满足了所有外部条件,已经突破了树种的限制,纯粹反映了气象因子与乔木树木生理生态特性的关系和时滞时间。马长明等[8]研究得出健康和亚健康北京杨单株液流速率滞后于太阳辐射30 min,提前于空气温度、湿度、饱和水汽压差60 min;而不健康单株液流速率则滞后于太阳辐射60 min,与空气温度、湿度、饱和水汽压差同步。他们的结论也从另外一个角度证明了本文的结果,如果满足所有条件,树木的树干液流将滞后太阳辐射0~30 min,提前于饱和水汽压差、气温和湿度0~60 min,如果不能100%满足条件,滞后时间或提前时间将更大。

图 6 喜树、西南红山茶和刺栲3种树种树干液流与气象因子时滞效应分析
Figure 6. Analysis of SAP flow and time delay effects of meteorological factors in three species of native evergreen broad-leaved forest and secondary deciduous broad-leaved forest
-
本文通过对哈尼梯田水源区3种林分中10种乔木树种树干液流与气象因子的关系分析中发现,如果考虑树种因素,影响树干液流的气象因子主要是太阳辐射、饱和水汽压差、气温和湿度,与太阳辐射、气温、饱和水汽压差呈正相关,而与湿度呈负相关,相关性大小由不同树种决定。如果消除树种因素限制,太阳辐射、气温和降雨是影响乔木生理生态树干液流的主要气象因子,其中太阳辐射是影响乔木树干液流的主要原因。在正常气候天气下,原生常绿阔叶林不同树种的树干液流滞后太阳辐射90~240 min,提前气温60~120 min,提前湿度90~120 min,提前于饱和水汽压差0~120 min;在满足发生同步异常数据的气候条件下,树干液流滞后太阳辐射0~30 min,提前于饱和水汽压差、气温和湿度0~60 min,滞后或提前时间远小于正常气候条件下各树种的时滞时间。本研究为乔木树干液流与气象因子的关系和时滞时间提供了科学依据,对天气突变等原因影响乔木蒸腾有重要意义。
从同步数据分析乔木树干液流与气象因子的关系
Relationship between Tree Trunk Sap Flow and Meteorological Factors from Synchronous Data
-
摘要:
目的 树干液流是研究植物蒸腾耗水的重要生理指标,树种和气象因子均是其关键影响因素。本文旨在消除树种因素限制,探讨树干液流与气象因子之间的关系。 方法 以哈尼梯田水源区3种林分中10种乔木树种为研究对象,采用 Granier热扩散探针技术测定其树干液流速率,将其同步数据与气象因子进行关联分析。 结果 从次生落叶阔叶林和原生常绿阔叶林的7 779条乔木树干液流速率数据中发现7条异常数据,且这些异常数据在各个树种中都表现出同步一致性;进一步关联分析表明,太阳辐射的突然增加是产生乔木树干液流数据同步上升的主要原因,而降雨则是产生树干液流数据同步下降的主要原因,两者的变化可导致其他气象因子发生变化,从而影响树干液流速率;树干液流与气象因子存在一定的时滞效应,在发生同步异常数据的气候条件下,树干液流滞后太阳辐射0~30 min,提前于饱和水汽压差、气温和湿度0~60 min,滞后或提前时间远小于正常气候条件下各树种的时滞时间。 结论 乔木树干液流受到太阳辐射和降雨等气象因子的综合调节,其中太阳辐射的影响最为显著。研究结果可为从生理生态角度探究乔木的蒸腾作用与气象因子的关系提供科学依据。 Abstract:Objective Trunk sap flow is an important physiological indicator for the study of plant transpiration water consumption, and both tree species and meteorological factors are key influencing factors. This study aims to remove the limitation of tree species factor and explore the relationship between tree trunk sap flow and meteorological factors. Method Based on ten tree species in 3 forest stands in the water source area of Hani Terraces, their trunk sap flow rates were measured by using thermal dissipation probe (TDP) technology, and the relationship between synchronized data and meteorological factors were analyzed. Result Seven abnormal synchrony data were found from 7 779 tree trunk sap flow rate data in secondary deciduous broadleaf and primary evergreen broadleaf forests, and these abnormal synchrony data showed synchronous consistency across tree species; Further correlation analysis showed that the sudden increase in solar radiation was the main cause of the synchronous increase in sap flow rate, while rainfall was the main cause of the synchronous decrease in sap flow rate, and changes in both could lead to changes in other meteorological factors, which could affect the trunk sap flow rate; There was a certain time lag effect between tree trunk sap flow and meteorological factors: under abnormal climate conditions, sap flow lagged solar radiation by 0~30 min and was ahead of saturated water vapor pressure difference, with air temperature and humidity of 0~60 min, and the lag or advance time was much smaller than the time of each tree species under normal climatic conditions. Conclusion Tree trunk sap flow is regulated by a combination of meteorological factors such as solar radiation or rainfall, with solar radiation having the most pronounced effect. The paper may provide a scientific basis for studying the relationship between transpiration and meteorological factors in trees at the physiological-ecological level. -
Key words:
- Trunk sap flow
- / meteorological factor
- / relationship analysis
- / abnormal synchrony
- / time-lag effect
-
图 1 研究区位置及Granier热扩散测量树干液流速率的原理[28]与现场测定
Figure 1. Schematic diagram of study area location and Granier thermal diffusion measurement of SAP flow rate
图 2 次生落叶阔叶林和原生常绿阔叶林出现的7个同步异常数据
Figure 2. Seven synchronous anomalies in secondary deciduous broad-leaved forest and primary evergreen broad-leaved forest
图 3 同步异常树干液流与气象因子关系分析
Figure 3. Relationship analysis diagram of synchronous abnormal SAP flow and meteorological factors
图 4 饱和水汽压差和同步数据各时间段分布
Figure 4. Distribution of saturated water vapour differential pressure and synchronous data by time period
图 5 影响同步树干液流的气象原因分析
Figure 5. Analysis of the meteorological causes affecting synchronous trunk sap flow
图 6 喜树、西南红山茶和刺栲3种树种树干液流与气象因子时滞效应分析
Figure 6. Analysis of SAP flow and time delay effects of meteorological factors in three species of native evergreen broad-leaved forest and secondary deciduous broad-leaved forest
表 1 树干液流速率监测样木基本参数
Table 1. Basic parameters of sample wood were monitored by SAP flow rate
试验点
Study sites林分类型
Forest type样树编号
Sample tree number胸径
DBH/cm树高
HT/m冠幅(东西,南北)
RTC/(m*m)试验点1
Study plot1次生常绿阔叶林
Secondary evergreen broad-leaved forest液1-尖齿木荷1 Schima khasiana 10.7 11.5 2.1,1.6 液1-尖齿木荷2 Schima khasiana 10.8 13.5 1.8,1.5 液1-元江锥1 Castanopsis orthacantha 11.3 14.5 2.0,1.4 液1-元江锥2 Castanopsis orthacantha 11.2 15.0 2.4,1.6 液1-喜树1 Camptotheca acuminata 12.5 13.0 1.8,1.6 液1-喜树2 Camptotheca acuminata 11.5 12.2 1.6,1.5 试验点2
Study plot2次生落叶阔叶林
Secondary deciduous broad-leaved forest液2-桤木1 Alnus cremastogyne 33.5 23.0 6.3,4.2 液2-桤木2 Alnus cremastogyne 25.2 20.3 4.2,3.2 液2-桤木3 Alnus cremastogyne 24.8 20.2 4.2,1.0 液2-喜树3 Camptotheca acuminata 15.3 15.6 2.1,2.0 液2-喜树4 Camptotheca acuminata 28.4 22.1 4.5,4.7 试验点3
Study plot3原生常绿阔叶林
Native evergreen broad-leaved forest液3-云南樟1 Cinnamomum glanduliferum 22.0 17.2 4.2,3.1 液3-云南樟2 Cinnamomum glanduliferum 24.0 18.1 4.5,3.4 液3-西南红山茶1 Camellia pitardii 18.7 14.5 4.2,4.1 液3-西南红山茶2 Camellia pitardii 20.0 15.2 5.7,4.6 液3-西南红山茶3 Camellia pitardii 31.0 18.3 6.9,5.1 液3-刺栲1 Castanopsis hystrix 48.0 25.0 6.2,5.3 液3-刺栲2 Castanopsis hystrix 12.5 13.2 2.3,1.5 液3-猴樟1 Cinnamomum bodinieri 27.8 22.3 5.3,4.2 液3-猴樟2 Cinnamomum bodinieri 23.0 19.2 4.2,3.6 液3-三桠苦 Evodia lepta 31.0 25.3 7.3,6.5 注:DBH为胸径,HT为树高,RTC为冠幅
Notes: DBH, diameter at breast height; HT, height of tree; RTC, range of tree-crown 下载: 导出CSV
下载: 导出CSV
表 2 样树树干液流变化幅度最大值和频次
Table 2. Table of frequency and maximum values of trunk sap flow variation of sample trees
样树编号
Sample tree number变化幅度频次 Frequency of change amplitude 最大值/%
MaximumR<−50% −50%≤R<0 0%≤R<50% 50%≤R<100% 100%≤R 液2-桤木2 Alnus cremastogyne 109 822 862 132 224 1 568 液2-喜树4 Camptotheca acuminata 100 890 817 120 222 2 355 液2-喜树3 Camptotheca acuminata 194 740 865 132 218 9 632 液3-云南樟1 Cinnamomum glanduliferum 163 692 723 128 194 2 750 液3-西南红山茶2 Camellia pitardii 167 717 856 45 115 6 520 液3-猴樟2 Cinnamomum bodinieri 164 701 819 87 129 55 127 注:R表示每日6:00至18:00时间内每30 min的变化幅度
Note: R represents the range of change every 30 minutes between 6:00 and 18:00 per day
下载: 导出CSV
表 3 各树种生长季树干液流速率与各气象因子的Pearson相关系数分析
Table 3. Pearson correlation coefficient between SAP flow rate of tree species and meteorological factors in growing season
树种
Varieties of trees月份
Month太阳辐射
Rn气温
Ta饱和水汽压差
VPD湿度
RH风速
SP大气压
P降雨量
Pn云南樟
Cinnamomum glanduliferum5月 0.70** 0.68** 0.74** −0.71** 0.48** −0.2** 0.08* 6月 0.51** 0.57** 0.59** −0.54** 0.26** −0.19* −0.02 7月 0.63** 0.63** 0.66** −0.64** 0.25** −0.06 −0.03 西南红山茶
Camellia pitardii5月 0.68** 0.62** 0.64** −0.59** 0.43** −0.15* 0.14** 6月 0.50** 0.45** 0.49** −0.45** 0.19* −0.027 −0.07 7月 0.59** 0.57** 0.60** −0.57** 0.21** −0.03 −0.01 猴樟
Cinnamomum bodinieri5月 0.33** 0.29** 0.28** −0.26** 0.11 −0.03 0.08* 6月 0.20** 0.19** 0.15** −0.15** 0.12* −0.09 0.02 7月 0.28** 0.25** 0.25** −0.21** 0.06 −0.26** 0.17** 三桠苦
Evodia lepta5月 0.23** 0.17** 0.22** −0.23** 0.14** 0.04 −0.01 6月 0.34** 0.46** 0.45** −0.41** 0.18** −0.14* −0.01 7月 0.30** 0.33** 0.33** −0.33** 0.27** 0.10 0.28** 桤木
Alnus cremastogyne3月 0.63** 0.66** 0.66** −0.55** 0.60** −0.54** −0.02 7月 0.45** 0.46** 0.44** −0.41** 0.12* 0.44** 0.20** 8月 0.58** 0.67** 0.72** −0.68** 0.02 −0.32** 0.02 9月 0.52** 0.63** 0.62** −0.56** 0.23** −0.45** 0.06 喜树
Camptotheca acuminata3月 0.69** 0.62** 0.58** −0.46** 0.61** −0.34** 0.08 7月 0.75** 0.73** 0.71** −0.69** 0.17* −0.33** 0.11* 8月 0.84** 0.79** 0.80** −0.79** 0.12* −0.18* 0.01 9月 0.77** 0.74** 0.69** −0.64** 0.27** 0.28** 0.05 注:**表示在0.01水平上极显著相关,*表示在0.05水平上显著相关
Notes: ** shows correlation is significant at the 0.01 level, * shows correlation is significant at the 0.05 level
下载: 导出CSV
-
[1] 王 媛, 魏江生, 周 梅, 等. 大兴安岭南段白桦树干液流对土壤水分的响应[J]. 水土保持研究, 2020, 27(4):128-133. [2] NADEZHDINA N. Sap flow index as an indicator of plant water status[J]. Tree Physiology, 1999, 19(13): 885-891. doi: 10.1093/treephys/19.13.885 [3] 陈孟涤, 覃鑫浩, 毛炎新, 等. 半干旱区华北落叶松林冠层蒸腾特征及其影响因子[J]. 西北林学院学报, 2022, 37(4):10-17. [4] 张 俊, 李晓飞, 李建贵, 等. 干旱荒漠区银白杨树干液流动态[J]. 生态学报, 2013, 33(18):5655-5660. [5] 王小菲, 孙永玉, 李 昆, 等. 干热河谷大叶相思树干液流季节动态及其与气象因子的关系[J]. 林业科学研究, 2013, 26(2):145-150. [6] 于占辉, 陈云明, 杜 盛. 黄土高原半干旱区侧柏(Platycladus orientalis)树干液流动态[J]. 生态学报, 2009, 29(7):3970-3976. doi: 10.3321/j.issn:1000-0933.2009.07.061 [7] 于占辉, 陈云明, 杜 盛. 黄土高原半干旱区人工林刺槐展叶期树干液流动态分析[J]. 林业科学, 2009, 45(4):53-59. doi: 10.3321/j.issn:1001-7488.2009.04.009 [8] 马长明, 马玉洁, 程月明. 冀西北坝上干旱区北京杨树干液流特征及影响因素分析[J]. 水土保持学报, 2017, 31(6):338-344. [9] 张 璇, 张会兰, 王玉杰, 等. 缙云山典型树种树干液流日际变化特征及与气象因子关系[J]. 北京林业大学学报, 2016, 38(3):11-20. [10] 蒋文伟, 杨广远, 赵明水, 等. 天目山柳杉树干液流的昼夜及季节变化[J]. 南京林业大学学报(自然科学版), 2012, 36(5):77-80. [11] MCDOWELL N G, WHITE S, POCKMAN W T. Transpiration and stomatal conductance across a steep climate gradient in the southern Rocky Mountains[J]. Ecohydrology, 2008, 1(3): 193-204. doi: 10.1002/eco.20 [12] FAN C, QIU Y P, LI Z Q, et al. Relationships between trunk sap flow rate of litchi trees and meteorological parameters[J]. Acta Ecologica Sinica, 2014, 34(9): 2401-2410. [13] 买尔当·克依木, 玉米提·哈力克, 古丽卡玛尔·迪力木拉提, 等. 阿克苏市3种绿化乔木树干液流日变化特征[J]. 森林与环境学报, 2016, 36(4):473-479. [14] 孙 旭, 杨文慧, 焦 磊, 等. 不同时间尺度北京蟒山油松树干液流对环境因子的响应[J]. 生态学报, 2022, 42(10):4113-4123. [15] 方伟伟, 吕 楠, 傅伯杰. 植物夜间液流的发生、生理意义及影响因素研究进展[J]. 生态学报, 2018, 38(21):7521-7529. [16] 高照全, 邹养军, 王小伟. 植物水分运转影响因子的研究进展[J]. 干旱地区农业研究, 2004, 22(2):200-204. doi: 10.3321/j.issn:1000-7601.2004.02.042 [17] 马长明, 刘广营, 张艳华, 等. 核桃树干液流特征研究[J]. 西北林学院学报, 2010, 25(2):25-29. [18] 武鹏飞, 刘云强, 李冬梅, 等. 环境因子对沙地人工杨树林树干液流的驱动影响[J]. 中国农业气象, 2021, 42(5):402-411. [19] 彭 苓, 韩 磊, 韩永贵, 等. 宁夏河东沙区丝棉木树干液流昼夜变化及其受气象因子的影响[J]. 东北林业大学学报, 2022, 50(8):28-32 + 48. [20] 党宏忠, 杨文斌, 李 卫, 等. 新疆杨树干液流的径向变化及时滞特征[J]. 生态学报, 2015, 35(15):5110-5120. [21] 王 瑛, 刘美君, 杜 盛. 树干液流时滞特征及影响因素研究进展[J]. 应用与环境生物学报, 2023, 29(2):507-514. [22] TU J, WEI X H, HUANG B B. Improvement of sap flow estimation by including phenological index and time-lag effect in back-propagation neural network models[J]. Agricultural and Forest Meteorology, 2019(2): 276-287. [23] ZHAO C Y, SI J H, FENG Q, et al. Trunk sap flow research: progress and prospect[J]. Journal of Northwest Forestry University, 2015, 30(5): 98-105. [24] 田 原, 张秋良, 刘 璇, 等. 兴安落叶松树干液流与太阳辐射的时滞效应[J]. 东北林业大学学报, 2018, 46(5):23-26. [25] 张 荣, 毕华兴, 焦振寰, 等. 生长季刺槐树干液流昼夜变化特征及其对气象因子的响应[J]. 浙江农林大学学报, 2022, 39(6):1238-1246. [26] 姚 敏, 崔保山. 哈尼梯田湿地生态系统的垂直特征[J]. 生态学报, 2006, 26(7):2115-2124. [27] 宋维峰, 吴锦奎. 哈尼梯田-历史现状、生态环境、持续发展 [M]. 北京: 科学出版社, 2016. [28] GRANIER A. Evaluation of transportation in a Douglas-firstand by means of sap flow measurements[J]. Tree Physiology, 1987, 3: 309-320. doi: 10.1093/treephys/3.4.309 [29] CAMPBELL G S, NORMAN J M. An Introduction to Environmental Biophysics [M]. New York: Springer, 1998. [30] FORD C R, GORANSON E C, MITCHELL J R, et al. Diurnal and seasonal variability in the radial distribution of sap flow: Predicting total trunk flow in Pinus taedatres[J]. Tree physiology, 2004, 24(9): 951-960. doi: 10.1093/treephys/24.9.951 [31] 王小菲, 孙永玉, 李 昆, 等. 山合欢树干液流的季节变化[J]. 生态学杂志, 2013, 32(3):597-603. [32] 万艳芳, 于澎涛, 刘贤德, 等. 祁连山青海云杉树干液流密度的优势度差异[J]. 生态学报, 2017, 37(9):3106-3114. [33] 赵春彦, 司建华, 冯 起, 等. 树干液流研究进展与展望[J]. 西北林学院学报, 2015, 30(5):98-105. [34] 张 雷, 孙鹏森, 刘世荣. 树干液流对环境变化响应研究进展[J]. 生态学报, 2009, 29(10):5600-5610. doi: 10.3321/j.issn:1000-0933.2009.10.051 [35] ZHANG H D, WEI W, CHEN L D, et al. Analysis of sap flow characteristics of the Chinese pine in typical Loess Plateau region of China[J]. Environmental Science, 2015, 36(1): 349-356. [36] ZHANG R, XU X, LIU M, et al. Hysteresis in sap flow and its controlling mechanisms for a deciduous broad-leaved tree species in a humid karst region[J]. Sci China Earth Sci, 2019, 62(11): 1744-1755. doi: 10.1007/s11430-018-9294-5 [37] 郭孟霞, 毕华兴, 刘 鑫, 等. 树木蒸腾耗水研究进展[J]. 中国水土保持科学, 2006, 4(4):114-120. [38] WULLSCHLEGER S D, MEINZER F C, VERTESSY R A. A critique of whole-plant water use studies in trees[J]. Tree Physiology, 1998, 18: 499-512. doi: 10.1093/treephys/18.8-9.499 [39] JONARD F, ANDRé F, PONETTE Q, et al. Sap flux density and stomatal conductance of European beech and common oak trees in pure and mixed stands during the summer drought of 2003[J]. Journal of Hydrology, 2011, 409(1-2): 371-381. doi: 10.1016/j.jhydrol.2011.08.032 [40] WANG W J, SUN W, QIU L, et al. Relations between trunk sap flow density of Larix gmelinii and environmental factors under different temporal scale[J]. Scientia Silvae Sinicae, 2012, 48(1): 77-85. [41] 涂 洁, 王永文. 退化红壤区针阔混交林3种优势树种树干液流特征比较[J]. 河南农业科学, 2016, 45(12):120-126. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 762
- HTML全文浏览量: 382
- PDF下载量: 26
- 被引次数: 0




