


-
文冠果(Xanthoceras sorbifolium Bunge),又名黄角(Yellow horn),落叶灌木或小乔木,是无患子科文冠果属的唯一植物,广泛种植于我国三北地区,抗性强,尤其抗旱,是我国特有的观赏、药用及油用的高收入木本经济树种[1]。文冠果种子富含油脂,种仁含油率达60%以上,不饱和脂肪酸组成高达92%,富含油酸(约30%)、亚油酸(约45%)以及在植物中极为罕见的神经酸(约3%)[2],是生产高品质生物柴油和健康食用油的原料,具有优秀的经济价值。由此可见,研究文冠果种子不饱和脂肪酸的合成机制,对培育优质高产的文冠果品种具有现实意义。截止至今,已有文冠果种子发育转录组的文献分析出一些重要的不饱和脂肪酸合成相关候选关键基因[3-4],探究这些基因在文冠果各组织,尤其不同发育时期种子中的表达模式,尤为关键。
实时荧光定量PCR(qRT-PCR)是大多数科学家常用的检测基因表达丰度的可靠技术,具有高灵敏性、特异性、精确性[5],而一个可靠的内参基因(RGs)是前提。内参基因一般选择在植物中稳定表达的管家基因,常见的管家基因有actin(ACT)、糖醛酸-3-磷酸脱氢酶基因(GADPH)、延长因子1α(EF-1α)、泛素酶基因(UBQ)、泛素结合酶基因(UCE)以及18S 核糖体RNA(18S),然而这些基因的表达在不同物种的不同组织和其不同发育时期中的稳定性有较大差异,需要深入研究[6-7]。
大量关于文冠果基因表达的研究均采用actin作为内参基因[4,8-13],王旭等通过对7个内参基因稳定性研究,发现UBQ和eIF-4α为适用于性别分别相关基因的最优内参基因[14]。然而,近年来神经酸的营养价值和药用价值被逐渐挖掘,其种子油脂合成与代谢相关基因的研究亟待深入,需要进一步研究文冠果种子不同发育时期的最适内参基因需要被进一步研究,以便提高其油脂合成与代谢相关基因表达检测的准确性。基于此,本研究首先以SapBase数据库中文冠果各组织转录组数据(http://www.sapindaceae.com/)中常见内参基因的基因表达量数据作为依据[15],挑选12个在各组织表达量较稳定的候选内参基因;采用qRT-PCR法测试这12个候选内参基因在文冠果种子不同发育时期和不同组织中的表达水平Cq值;利用4种常见的评估内参基因的表达稳定性的数学算法评价这12个候选内参基因在文冠果不同组织和种子发育不同时期中的稳定性,随即利用在线工具RefFinder对这12个候选内参基因进行综合排名选出最佳内参基因。最后,以超长链脂肪酸合成关键酶3-酮酯酰-CoA合酶(KCS)基因验证选择的最佳内参基因的稳定性,本研究旨在选择稳定、可靠的文冠果内参基因。
-
文冠果(Xanthoceras sorbifolium Bunge)成熟实生苗(五年生以上)生长于内蒙古通辽市开鲁县(43°32′ N, 120°30′ E),林间正常管理。文冠果根、茎、叶、花采集于5月10日,种皮采集于6月7日,不同发育时期种子胚乳分别采集于6月18日(谢花后第30 d,30 DAP),6月26日(38 DAP),7月3日(45 DAP),7月11日(53 DAP),7月21日(63 DAP),采集年份均为2022年,5个不同发育时期种子胚乳形态特征见图1。每种文冠果组织采集生物学重复样品3份。

图 1 文冠果种子不同发育时期胚乳形态(30 DAP~63 DAP)
Figure 1. Seed endosperm morphology at different developmental stages (30 DAP~63 DAP)
-
文冠果各植物组织的提取方法参照多糖多酚植物总RNA提取试剂盒(天根,中国)说明书,经过降解酶RQ1 RNase-Free DNase(Promega,美国)降解残留的基因组DNA后,以1 μg RNA为模板,利用GoScriptTM Reverse Transcription System(Promega,美国)试剂盒反转录得到cDNA,稀释10倍后冻于−20 ℃待用
-
参照SapBase数据库(
http://www.sapindaceae.com/ )中文冠果各组织转录组数据(SRA编号:SRP304628),选择在各组织表达量较稳定的12个内参基因作为候选,同时以超长链脂肪酸合成关键酶3-酮酯酰-CoA合酶(KCS)基因作为验证基因,基因编号均来源于SapBase数据库[15]。荧光定量PCR(qRT-PCR)引物由软件Primer Premier 5.0设计,引物信息列于表1。扩增序列均测序检验其正确性。表 1 12个候选内参基因引物和扩增效率
Table 1. 12 candidate reference gene primers and amplification efficiency
基因/基因编号
Gene/Gene number基因名称
Gene name引物序列(5′–3′)正向/反向
Primer sequences (5′-3′)
forward/reverse扩增长度
Ampliction
length/bp扩增效率
Amplification
efficiency/%线性相关系数R2
Correlation
coefficient
R2平均值
Cq Average
Cq valueACT/EVM0004930 肌动蛋白 TACCGAGGCACCATTAAATCCC/
AAGGTCCAAACGAAGAATAGCA226 99.23 0.997 21.51 UBQ/EVM0019311 泛素蛋白 TTCCGTCTTGCTCTCTACACTTCC/
GTTTCTGCTTATTGACATCTTTATCC176 98.75 0.995 24.59 UCE/EVM0006084 泛素结合酶 AATGGGGCTTCAGGAACTCTCTAT/
TATCTTCAGGGCGTTGCTTCACAG256 101.14 0.993 24.33 18r/EVM0012402 18S核糖体RNA CAGGCACTACAGTCCTCACAAAA/
GTCACTTCCACCATCCATATCCA117 99.46 0.998 26.68 GADPH/EVM0014701 糖醛酸-3-磷酸脱氢酶 GCAACCTATGAGGACATCAAAGC/
CCAACGAAATCGGTAGAAACAAC107 98.23 0.992 21.59 EF-1α/EVM0013711 延伸因子 GTGCTATGTCACCCCGATTTCCC/
TTTTACCATTGTCGCAGCCTCCT142 98.88 0.995 24.38 α-TUB/EVM0000325 α微管蛋白 CAATCTCAACCGCCTTGTCTCTC/
AGCTGCTCATGGTATGCCTTCTC183 100.72 0.996 22.94 β-TUB/EVM0013594 β微管蛋白 GGGGCAAAATGAGCACTAAAGAA/
CCAGTCGGAGGGATATCACAGAC127 101.35 0.994 23.67 PP2A/EVM0018196 蛋白质磷酸酯酶 TCTTTGACTTCACTGATCCGACA/
TGGACACCATCTTTACCATTTCTT144 99.46 0.998 27.39 Tip41/EVM0004487 Tip41蛋白 AAGATGAATTGGCTGATAACGGA/
AGCTCTCACGAAGAATGACTGGG183 98.58 0.994 25.53 SAM/EVM0014230 甲硫氨酸腺苷转移酶 TGAGGTGAGAAAGAACAAGACGT/
TGCTGGGTTGAAATGAGAACAGTG132 99.73 0.998 25.34 CYP/EVM0006460 亲环蛋白 GAAAGGTGGAGAACAATGATAGAT/
TTGAAAGCACAACACAGGAGAGT152 101.12 0.993 27.15 KCS/EVM0000575 3-酮酯酰-CoA合酶 TCCAAAGGGAGGATGACAACA/
CTCGGACATGGGAAGGACTAG125 99.32 0.997 25.96 -
qRT-PCR由ABI StepOne7000(美国)实时荧光定量PCR仪完成,反应体系为30.0 μL,包括SYBR qPCR SuperMix 15.0 μL,上游引物(10.0 μmol·L−1)1.0 μL,下游引物(10.0 μmol·L−1)1.0 μL,ROX 0.5 μL,ddH2O 12.5 μL。反应采用三步法,反应程序为:95 °C. 2 min,1个循环;95 °C 10 s,60 °C,20 s,72 °C,20 s,40个循环;72 °C,20 s,1个循环。熔解曲线在65~95 °C间获得以验证引物的特异性。引物的扩增效率和线性相关系数由cDNA按5个稀释倍数(1、10−1、10−2、10−3、10−4)建立标准曲线计算得到。
4种常见的评估内参基因的表达稳定性的数学算法ΔCT[16]、Best-Keeper[17]、geNorm[18]及NormFinder[19]用于评价12个候选内参基因在文冠果种子不同发育时期和不同组织的稳定性。最后利用在线工具RefFinder(
http://150.216.56.64/referencegene )综合排名。 -
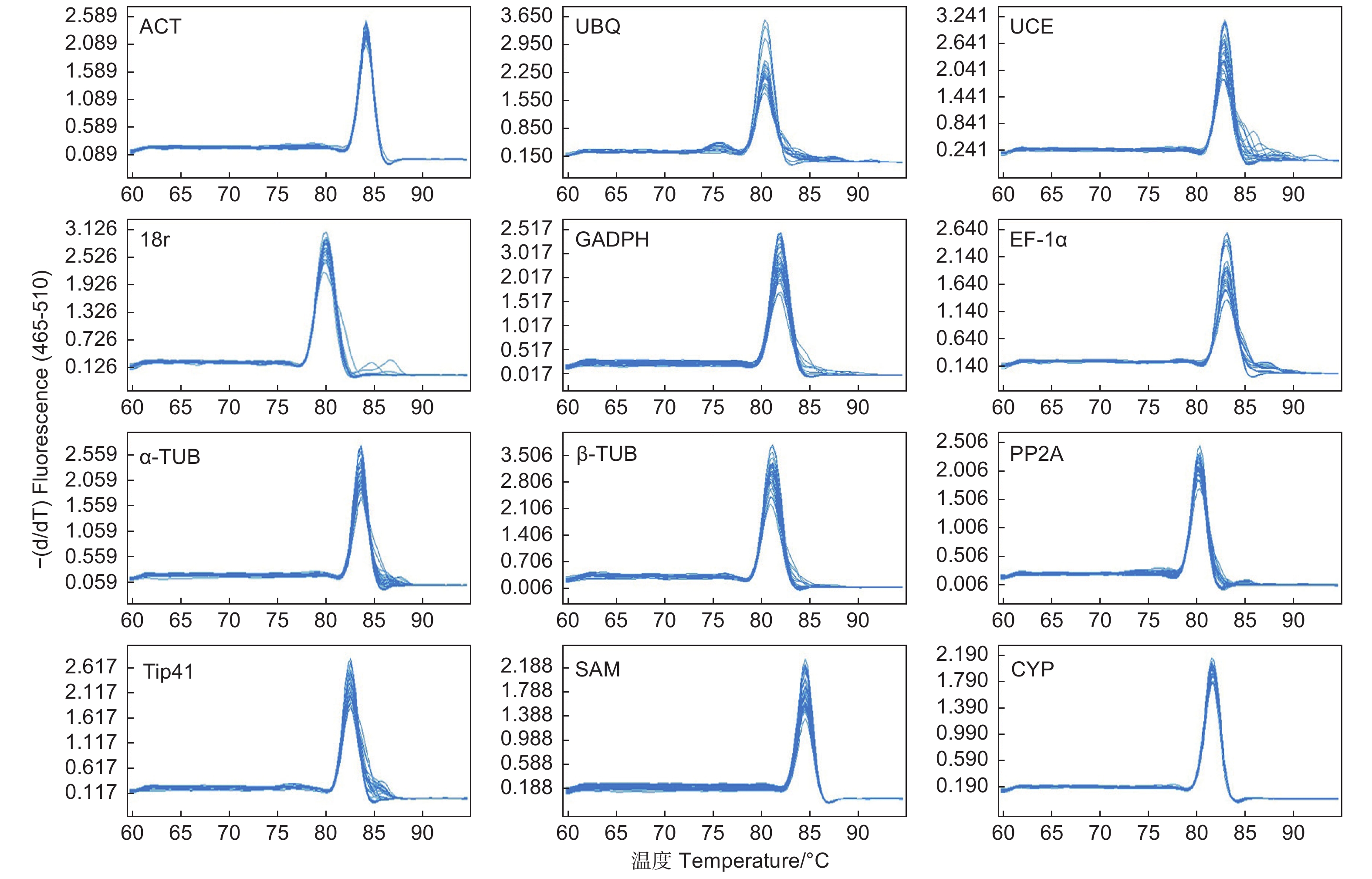
所有样品RNA均利用Agilent 2100 Bioanalyzer生物分析仪测试RNA浓度和完整性,样本RNA浓度分布于128.5~433.9 ng·μL−1,RIN值均大于7.0,琼脂糖电泳检测均显示有明显的28S和18S条带,RNA质量达到后续实验要求。12个候选内参基因的基因名称、SapBase编号、引物序列、扩增长度、扩增效率、相关系数及Cq平均值均列于表1。扩增片段长度在107 bp(GADPH)~256 bp(UCE),扩增效率在98.23%(GADPH)~101.35(β-TUB)。12个候选内参基因熔解曲线均为单一峰(图2),表明无引物二聚体和非特异性扩增发生。
图 2 12个候选内参基因熔解曲线
Figure 2. Melting curves with a single peak for the 12 candidate reference genes
-
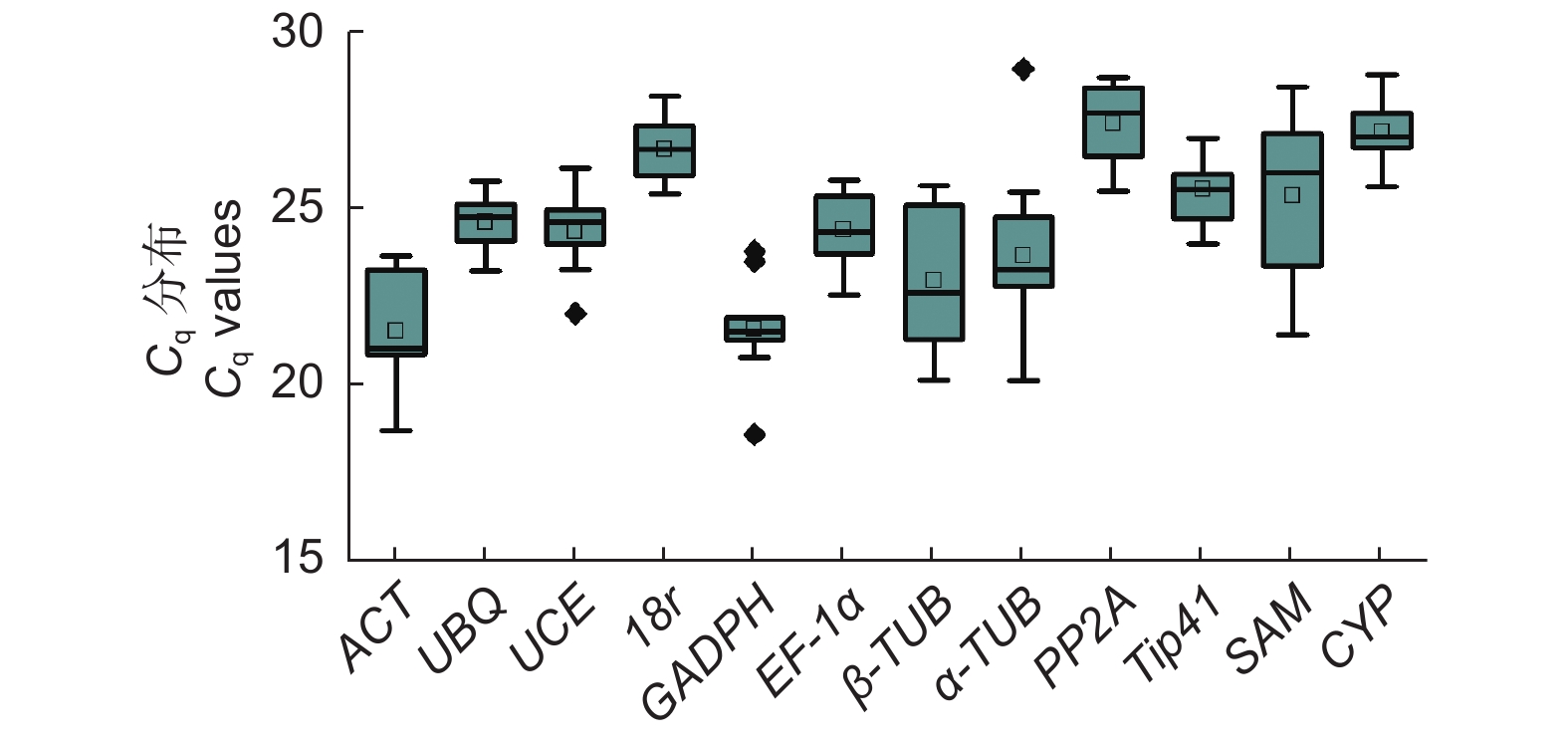
qRT-PCR法得到12个候选内参基因在文冠果不同组织和不同种子发育时期中的表达水平Cq值,Cq值越小,基因表达量越高。这些内参基因Cq值在文冠果各个组织有较大差异。这所有检测的组织中,PP2A平均Cq值最高27.39,分布于25.47~28.69;ACT平均Cq值最低21.51,分布18.69~23.65(表1,图3)。这12个候选内参基因的变异系数分别为7.18%(ACT)、3.01%(UBQ)、4.54%(UCE)、3.24%(18r)、6.64%(GADPH)、4.56%(EF-1α)、8.84%(β-TUB)、10.68%(α-TUB)、4.13%(PP2A)、3.87%(Tip41)、9.60%(SAM)、3.11%(CYP)。

图 3 12个候选内参基因Cq值分布
Figure 3. The quantification cycle (Cq) values of the 12 candidate reference genes
-
常见的内参基因稳定性算法有ΔCT、BestKeeper、NormFinder以及GeNorm。这4种算法中,ΔCT法是根据Cq值的标准差数值来评价,标准差数值越低的内参基因,稳定性越强;BestKeeper算法是根据Cq值的变异系数和标准差数值联合评价,变异系数和标准差数值越低,内参基因的稳定性越强;NormFinder和GeNorm算法均根据平均表达稳定指数计算,表达稳定指数与内参基因的稳定性呈负相关。
上述4种内参基因稳定性算法用于评价12个候选内参基因在文冠果种子不同发育时期和不同组织中的稳定性。如表2所示,根据ΔCT和BestKeeper算法分析,PP2A和Tip41在文冠果种子不同发育时期和不同组织中稳定性均为最优,NormFinder和GeNorm算法分析的最优结果与ΔCT和BestKeeper算法不同,但PP2A和Tip41均在排名前四的行列。在文冠果种子不同发育时期中,所有算法均显示ACT和GADPH为最不稳定的内参基因;在不同组织中,所有算法均显示ACT和β-TUB为最不稳定的内参基因。
表 2 候选内参基因在文冠果种子不同发育时期和不同组织的稳定性评价
Table 2. Stability ranking of candidate reference genes in seeds at different developmental stages and across diverse tissue samples
方法
Method稳定性排名(高-低) Ranking order(high-low) 1 2 3 4 5 6 7 8 9 10 11 12 不同发育时期种子 Seeds at different stages ΔCT Tip41 PP2A UCE EF-1α UBQ 18r β-TUB CYP α-TUB SAM ACT GADPH BestKeeper Tip41 PP2A UBQ EF-1α 18r UCE β-TUB CYP SAM α-TUB ACT GADPH NormFinder β-TUB UCE Tip41 PP2A α-TUB EF-1α UBQ 18r CYP SAM ACT GADPH GeNorm EF-1α/ PP2A Tip41 18r UBQ UCE β-TUB CYP SAM α-TUB ACT GADPH RefFinder PP2A Tip41 EF-1α 18r UCE β-TUB UBQ α-TUB CYP SAM ACT GADPH 不同组织 All tissues ΔCt Tip41 PP2A 18r CYP UBQ EF-1α SAM UCE GADPH α-TUB ACT β-TUB BestKeeper Tip41 PP2A 18r Tip41 UBQ EF-1α UCE SAM GADPH α-TUB ACT β-TUB NormFinder Tip41 SAM PP2A 18r CYP UBQ EF-1α GADPH UCE α-TUB ACT β-TUB GeNorm PP2A/CYP Tip41 18r UBQ EF-1α UCE SAM GADPH α-TUB ACT β-TUB RefFinder PP2A Tip41 CYP 18r UBQ SAM EF-1α UCE GADPH α-TUB ACT β-TUB 在线工具RefFinder根据4种数学算法的排名综合评价这12个候选内参的稳定性。结果如表2所示,在文冠果不同发育时期种子内参基因综合评分排名为PP2A>Tip41> EF-1α>18r>UCE>β-TUB>UBQ>α-TUB>CYP>SAM>ACT>GADPH;在文冠果不同组织内参基因综合评分排名为PP2A>Tip41>CYP>18r>UBQ>SAM>EF-1α>UCE>GADPH>α-TUB>ACT>β-TUB。PP2A和Tip41在文冠果不同发育时期种子和不同组织中的排名均为最高,表明为最合适的内参基因。
-
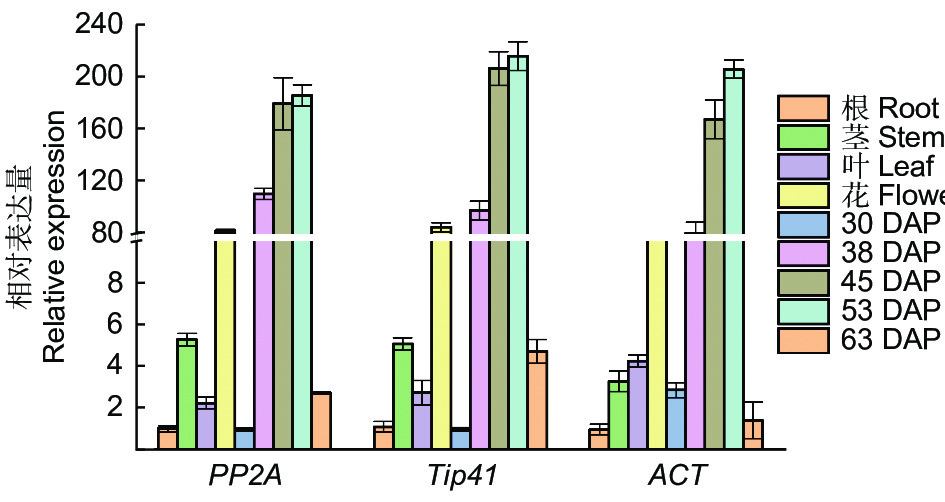
为了评估上述选择的最优内参基因的稳定性,本研究以选择的最优内参基因PP2A和Tip41及文献中最常用的ACT作为后续验证实验的内参基因,利用qRT-PCR法计算种子高表达目标基因KCS在文冠果种子不同发育时期和不同组织中的表达水平。结果如图4所示,以PP2A和Tip41作为内参基因达到大致相同的KCS表达模式(图4A、B)。KCS主要在种子发育期38~53 DAP高表达。在除了种子外的其他组织中,在种皮中几乎不表达,不参与计算,在花中高表达,然而,当采用ACT作为内参基因时,其表达模式有所不同(图4C)。在以PP2A和Tip41作为内参基因时,KCS基因在种子发育30 DAP的表达最低,作为标准值“1”,在茎的表达量高于叶片;而以ACT作为内参基因在根中的表达量最低,且在叶片中的表达量高于茎。在不同发育时期种子中,KCS基因以PP2A和Tip41作为内参基因计算时,种子发育63 DAP的表达量高于30 DAP,而以ACT作为内参基因时相反。上述结果表明,以不同稳定性的内参基因计算得到的组织特异性表达分析结果有所不同。由此可见,选择可靠稳定的内参基因对qRT-PCR实验的准确度非常重要。

图 4 KCS基因在不同内参基因下的组织特异性表达分析
Figure 4. Relative expression of the KCS gene in diverse tissue samples using selected reference genes
-
实时荧光定量(qRT-PCR)技术因其具有高灵敏性、特异性、精确性等特点,是分析基因表达水平和核酸含量的应用最为广泛的技术之一。不同物种之间没有固定的内参基因,例如常用的内参基因ACT、GADPH、UBQ等在不同植物物种的不同组织的不同发育时期可表现出表达差异[20]。由此可见,为特定植物的特定组织发育的qRT-PCR实验选择可靠的内参基因极为必要。
文冠果油是在植物种油中极为罕见的含有神经酸的木本油料来源,大量文献均以内参基因ACT作为文冠果基因表达分析的唯一内参基因[4,8-13],其稳定性是否适宜文冠果种子发育中相关基因的表达分析未有确定。本研究选择了12个常见且表达较为稳定的候选内参基因,利用4种数学算法(ΔCT、BestKeeper、NormFinder、geNorm)评估它们在文冠果种子发育过程中和不同组织中的稳定性,同时采用网络在线工具RefFinder综合这4种算法的结果并排名。
PP2A和Tip41在文冠果不同发育时期种子和不同组织中的排名均为最高,为本研究的最适内参基因,而UBQ和Tip41是油用牡丹(Paeonia ostii T. Hong et J. X. Zhang)种子发育过程中的最适内参基因,PP2A排名中游[6];UBCE2是草果(Amomum tsaoko Crevost et Lemarie)种子发育的最适内参基因[21];UCE、RPS13和RPII是美藤果(Plukenetia volubilis L.)种子发育最适内参基因[22]。以上结果表明不同物种不同发育组织最适内参基因大部分均不相同,值得研究。最为关键的是本研究结论中的最适内参基因与前期大部分研究报道所使用的并不一致,前期大量研究均采用ACT基因作为文冠果基因表达分析的内参基因[4,8-13],而本研究结果表明ACT基因在文冠果种子不同发育时期和不同组织中内参稳定性排名较差,并不适合用于基因表达的研究,同时王旭等研究也表明ACT基因不是研究文冠果性别分化和不同组织的最适内参基因,且排名同样靠后[14]。据此,本研究最重要的意义是更正用于文冠果基因表达分析的内参基因,以期获得更准确的基因表达数据。
-
本研究通过4种数学算法(ΔCT、BestKeeper、NormFinder、geNorm)对文冠果12个候选内参基因稳定性和有效性的评估,发现PP2A和Tip41基因在文冠果种子不同发育时期和不同组织中稳定表达,是文冠果基因表达分析最稳定的内参基因。本研究为文冠果qRT-PCR数据标准化提供了有效依据。
文冠果种子发育基因表达分析内参基因的选择
Selection of Reference Genes for Gene Expression Analysis in Seed Development of Xanthoceras sorbifolium Bunge
-
摘要:
目的 验证ACT基因是否适用于文冠果种子发育的相关基因表达分析,并筛选出在文冠果基因表达分析中最稳定的内参基因。 方法 本研究采用了4种数学算法(ΔCT、BestKeeper、NormFinder、geNorm)评估12个候选内参基因的表达稳定性,并利用在线工具RefFinder综合评价。 结果 在文冠果不同发育时期种子内参基因综合评分排名为PP2A>Tip41> EF-1α>18r>UCE>β-TUB>UBQ>α-TUB>CYP>SAM>ACT>GADPH;在文冠果不同组织内参基因综合评分排名为PP2A>Tip41>CYP>18r>UBQ>SAM>EF-1α>UCE>GADPH>α-TUB>ACT>β-TUB。 结论 PP2A和Tip41基因是文冠果基因表达分析最稳定的内参基因。 Abstract: :Objective : To assess the suitability of the ACT gene for gene expression analysis in seed development of Xanthoceras sorbifolium, and to identify the most stable reference genes for gene expression analysis in X. sorbifolium. Method : In this study, four mathematical algorithms (ΔCT, BestKeeper, NormFinder, geNorm) were carried out for evaluating the expression stability of 12 candidate reference genes based on the web-based tool RefFinder. Result : The RefFinder result showed that the gene PP2A performed the best, followed byTip41, EF-1α,18r, UCE, β-TUB, UBQ, α-TUB, CYP, SAM, ACT, and GADPH at the different seed development stages in X. sorbifolium , and the gene also had the highest score, followed by Tip41, CYP, 18r, UBQ, SAM, EF-1α, UCE, GADPH, α-TUB, ACT, and β-TUB at various tissues. Conclusion : PP2A and Tip41 are the two most stable reference genes for gene expression analysis in X. sorbifolium. -
Key words:
- Xanthoceras sorbifolium
- / reference gene
- / gene expression
- / seed development
-
图 1 文冠果种子不同发育时期胚乳形态(30 DAP~63 DAP)
Figure 1. Seed endosperm morphology at different developmental stages (30 DAP~63 DAP)
图 2 12个候选内参基因熔解曲线
Figure 2. Melting curves with a single peak for the 12 candidate reference genes
图 3 12个候选内参基因Cq值分布
Figure 3. The quantification cycle (Cq) values of the 12 candidate reference genes
图 4 KCS基因在不同内参基因下的组织特异性表达分析
Figure 4. Relative expression of the KCS gene in diverse tissue samples using selected reference genes
表 1 12个候选内参基因引物和扩增效率
Table 1. 12 candidate reference gene primers and amplification efficiency
基因/基因编号
Gene/Gene number基因名称
Gene name引物序列(5′–3′)正向/反向
Primer sequences (5′-3′)
forward/reverse扩增长度
Ampliction
length/bp扩增效率
Amplification
efficiency/%线性相关系数R2
Correlation
coefficient
R2平均值
Cq Average
Cq valueACT/EVM0004930 肌动蛋白 TACCGAGGCACCATTAAATCCC/
AAGGTCCAAACGAAGAATAGCA226 99.23 0.997 21.51 UBQ/EVM0019311 泛素蛋白 TTCCGTCTTGCTCTCTACACTTCC/
GTTTCTGCTTATTGACATCTTTATCC176 98.75 0.995 24.59 UCE/EVM0006084 泛素结合酶 AATGGGGCTTCAGGAACTCTCTAT/
TATCTTCAGGGCGTTGCTTCACAG256 101.14 0.993 24.33 18r/EVM0012402 18S核糖体RNA CAGGCACTACAGTCCTCACAAAA/
GTCACTTCCACCATCCATATCCA117 99.46 0.998 26.68 GADPH/EVM0014701 糖醛酸-3-磷酸脱氢酶 GCAACCTATGAGGACATCAAAGC/
CCAACGAAATCGGTAGAAACAAC107 98.23 0.992 21.59 EF-1α/EVM0013711 延伸因子 GTGCTATGTCACCCCGATTTCCC/
TTTTACCATTGTCGCAGCCTCCT142 98.88 0.995 24.38 α-TUB/EVM0000325 α微管蛋白 CAATCTCAACCGCCTTGTCTCTC/
AGCTGCTCATGGTATGCCTTCTC183 100.72 0.996 22.94 β-TUB/EVM0013594 β微管蛋白 GGGGCAAAATGAGCACTAAAGAA/
CCAGTCGGAGGGATATCACAGAC127 101.35 0.994 23.67 PP2A/EVM0018196 蛋白质磷酸酯酶 TCTTTGACTTCACTGATCCGACA/
TGGACACCATCTTTACCATTTCTT144 99.46 0.998 27.39 Tip41/EVM0004487 Tip41蛋白 AAGATGAATTGGCTGATAACGGA/
AGCTCTCACGAAGAATGACTGGG183 98.58 0.994 25.53 SAM/EVM0014230 甲硫氨酸腺苷转移酶 TGAGGTGAGAAAGAACAAGACGT/
TGCTGGGTTGAAATGAGAACAGTG132 99.73 0.998 25.34 CYP/EVM0006460 亲环蛋白 GAAAGGTGGAGAACAATGATAGAT/
TTGAAAGCACAACACAGGAGAGT152 101.12 0.993 27.15 KCS/EVM0000575 3-酮酯酰-CoA合酶 TCCAAAGGGAGGATGACAACA/
CTCGGACATGGGAAGGACTAG125 99.32 0.997 25.96  下载: 导出CSV
下载: 导出CSV
表 2 候选内参基因在文冠果种子不同发育时期和不同组织的稳定性评价
Table 2. Stability ranking of candidate reference genes in seeds at different developmental stages and across diverse tissue samples
方法
Method稳定性排名(高-低) Ranking order(high-low) 1 2 3 4 5 6 7 8 9 10 11 12 不同发育时期种子 Seeds at different stages ΔCT Tip41 PP2A UCE EF-1α UBQ 18r β-TUB CYP α-TUB SAM ACT GADPH BestKeeper Tip41 PP2A UBQ EF-1α 18r UCE β-TUB CYP SAM α-TUB ACT GADPH NormFinder β-TUB UCE Tip41 PP2A α-TUB EF-1α UBQ 18r CYP SAM ACT GADPH GeNorm EF-1α/ PP2A Tip41 18r UBQ UCE β-TUB CYP SAM α-TUB ACT GADPH RefFinder PP2A Tip41 EF-1α 18r UCE β-TUB UBQ α-TUB CYP SAM ACT GADPH 不同组织 All tissues ΔCt Tip41 PP2A 18r CYP UBQ EF-1α SAM UCE GADPH α-TUB ACT β-TUB BestKeeper Tip41 PP2A 18r Tip41 UBQ EF-1α UCE SAM GADPH α-TUB ACT β-TUB NormFinder Tip41 SAM PP2A 18r CYP UBQ EF-1α GADPH UCE α-TUB ACT β-TUB GeNorm PP2A/CYP Tip41 18r UBQ EF-1α UCE SAM GADPH α-TUB ACT β-TUB RefFinder PP2A Tip41 CYP 18r UBQ SAM EF-1α UCE GADPH α-TUB ACT β-TUB
下载: 导出CSV
-
[1] CHEN X Q, LEI Z L, CAO J, et al. Traditional uses, phytochemistry, pharmacology and current uses of underutilized Xanthoceras sorbifolium bunge: A review[J]. Journal of Ethnopharmacology, 2022, 283: 114747. doi: 10.1016/j.jep.2021.114747 [2] MA Y X, BI Q X, LI G T, et al. Provenance variations in kernel oil content, fatty acid profile and biodiesel properties of Xanthoceras sorbifolium Bunge in northern China[J]. Industrial Crops and Products, 2020, 151: 112487. doi: 10.1016/j.indcrop.2020.112487 [3] LIANG Q, LIU J N, FANG H C, et al. Genomic and transcriptomic analyses provide insights into valuable fatty acid biosynthesis and environmental adaptation of yellowhorn[J]. Frontiers in Plant Science, 2022, 13: 991197. doi: 10.3389/fpls.2022.991197 [4] WANG L, RUAN C J, LIU L Y, et al. Comparative RNA-seq analysis of high- and low-oil yellow horn during embryonic development[J]. International Journal of Molecular Sciences, 2018, 19(10): 3071. doi: 10.3390/ijms19103071 [5] BUSTIN S A, BENES V, NOLAN T, et al. Quantitative real-time RT-PCR – a perspective[J]. Journal of Molecular Endocrinology, 2005, 34(3): 597-601. doi: 10.1677/jme.1.01755 [6] LI C Q, HU L Z, WANG X Q, et al. Selection of reliable reference genes for gene expression analysis in seeds at different developmental stages and across various tissues in Paeonia ostii[J]. Molecular Biology Reports, 2019, 46(6): 6003-6011. doi: 10.1007/s11033-019-05036-7 [7] HU Y, CHEN H, LUO C, et. al. Selection of reference genes for real-time quantitative PCR studies of kumquat in various tissues and under abiotic stress[J]. Scientia Horticulturae, 2014, 174: 207-216. doi: 10.1016/j.scienta.2013.12.003 [8] GUO H H, LI Q Q, WANG T T, et al. XsFAD2 gene encodes the enzyme responsible for the high linoleic acid content in oil accumulated in Xanthoceras sorbifolia seeds[J]. Journal of the Science of Food and Agriculture, 2014, 94(3): 482-488. doi: 10.1002/jsfa.6273 [9] ZHAO N, ZHANG Y, LI Q Q, et al. Identification and expression of a stearoyl-ACP desaturase gene responsible for oleic acid accumulation in Xanthoceras sorbifolia seeds[J]. Plant Physiology and Biochemistry, 2015, 87: 9-16. doi: 10.1016/j.plaphy.2014.12.009 [10] GUO Y X, SONG H F, ZHAO Y Y, et al. Switch from symplasmic to aspoplasmic phloem unloading in Xanthoceras sorbifolia fruit and sucrose influx XsSWEET10 as a key candidate for sugar transport[J]. Plant Science, 2021, 313: 111089. doi: 10.1016/j.plantsci.2021.111089 [11] ZHAO Y, LIU X J, WANG M K, et al. Transcriptome and physiological analyses provide insights into the leaf epicuticular wax accumulation mechanism in yellowhorn[J]. Horticulture Research, 2021, 8(1): 134. doi: 10.1038/s41438-021-00564-5 [12] GUO H H, WANG T T, LI Q Q, et al. Two novel diacylglycerol acyltransferase genes from Xanthoceras sorbifolia are responsible for its seed oil content[J]. Gene, 2013, 527(1): 266-274. doi: 10.1016/j.gene.2013.05.076 [13] WANG L, RUAN C J, BAO A M, et al. Small RNA profiling for identification of microRNAs involved in regulation of seed development and lipid biosynthesis in yellowhorn[J]. BMC Plant Biology, 2021, 21(1): 464. doi: 10.1186/s12870-021-03239-4 [14] 王 旭, 敖 妍, 刘 阳, 等. 文冠果实时荧光定量PCR内参基因的筛选[J]. 分子植物育种, 2020, 18(9):2977-2986. [15] LI J W, CHEN C J, ZENG Z H, et al. SapBase (Sapinaceae Genomic DataBase): a central portal for functional and comparative genomics of Sapindaceae species[J]. BioRxiv, 2022, v1: 517904. [16] SILVER N, BEST S, JIANG J, et al. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR[J]. BMC Molecular Biology, 2006, 7(1): 33. doi: 10.1186/1471-2199-7-33 [17] PFAFFL M W, TICHOPAD A, PRGOMET C, et al. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper – Excel-based tool using pair-wise correlations[J]. Biotechnology Letters, 2004, 26(6): 509-515. doi: 10.1023/B:BILE.0000019559.84305.47 [18] VANDESOMPELE J, De PRETER K, PATTYN F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes[J]. Genome Biology, 2002, 3(7): research0034.1. doi: 10.1186/gb-2002-3-7-research0034 [19] ANDERSEN C L, JENSEN J L, ØRNTOFT T F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets[J]. Cancer Research, 2004, 64(15): 5245-5250. doi: 10.1158/0008-5472.CAN-04-0496 [20] NARANCIO R, JOHN U, MASON J, et al. Selection of optimal reference genes for quantitative RT-PCR transcript abundance analysis in white clover (Trifolium repens L. )[J]. Functional Plant Biology, 2018, 45(7): 737-744. doi: 10.1071/FP17304 [21] 姚李祥, 潘春柳, 余丽莹, 等. 草果种子休眠解除过程中qRT-PCR内参基因筛选[J]. 中国中药杂志, 2021, 46(15):3832-3837. [22] NIU L J, TAO Y B, CHEN M S, et al. Selection of reliable reference genes for gene expression studies of a promising oilseed crop, Plukenetia volubilis, by real-time quantitative PCR[J]. International Journal of Molecular Sciences, 2015, 16(6): 12513-12530. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 664
- HTML全文浏览量: 230
- PDF下载量: 10
- 被引次数: 0
