

 下载:
下载:



-
松枯梢病(又名松梢枯病shoot blight of pine)是世界范围内针叶树种上分布最广、最常见的重要林木枝干病害之一[1]。自从我国自20世纪70年代末报道以来,松枯梢病已蔓延至黑龙江、吉林、辽宁、陕西、江苏、湖北、福建、安徽、江西、广东和广西等10余个省份[2],严重危害松属(Pinus)、冷杉属(Abies)、落叶松属(Larix)、崖柏属(Thuja)、雪松属(Cedrus)、刺柏属(Juniperus)、云杉属(Picea)和黄杉属(Pseudotsuga)约8属60多种针叶树种[3]。该病害病原存在基因型的分化,De Wet通过多基因系谱学和微卫星标记手段将松枯梢病原菌划分为3个不同类型,分别为A型Diplodia scrobiculata,B型Diplodia seriata和C型Sphaeropsis sapinea(同物异名:Diplodia pinea)[4]。在中国引起松枯梢病的病原菌是C型松球壳孢菌(Sphaeropsis sapinea)[5],该病原菌可危害大树和幼树,大树多为侧枝发病,小树上顶梢发病,导致顶芽枯死、枯梢、根腐,严重时会出现流脂现象[6],对松树人工林造成了严重危害。目前,对松枯梢病主要采取化学防治与营林措施相结合的防治技术[7]。但长期使用化学农药不仅会导致环境污染、农药残留、威胁人类健康,而且容易诱导病原菌产生抗药性[8],甚至会导致病害的再次流行。而营林措施见效慢,投入成本较大。为有效防控松枯梢病,必须寻求一种生态友好的防治策略,木霉菌由于特有的拮抗和促生作用,现已成为一类应用较多的生防真菌[9],它通过竞争、寄生或产生抗菌素等次生代谢产物,从而对病原菌起到抑制作用[10],并能促进植物生长[11],提高寄主抵抗性。常见的生防木霉有哈茨木霉(Trichoderma harzianum)、绿色木霉(T. viride)、棘孢木霉(T. asperellum)等。研究表明,哈茨木霉ES323可有效抑制番茄灰霉病菌灰葡萄孢菌菌丝生长,引起菌丝腔质液泡化并导致菌体裂解[12]。非洲哈茨木霉菌(Trichoderma afroharzianum)株NAIMCC-F-01938发酵液可使葡萄白粉病原菌分生孢子扭曲变形,可作为安全杀菌剂在田间使用,可使葡萄白粉病发病率降低43%[13]。接种棘孢木霉可使洋葱鳞茎内酚类化合物含量提高97.6%,提高了洋葱对叶枯病的抵御能力,对洋葱叶枯病病原菌Stemphylium vesicarium具有显著的防控效果[14]。内生木霉菌株V76-12可抑制油棕叶斑病原菌Curvularia oryzae菌丝生长,同时提高了油棕幼苗苯丙氨酸解氨酶PAL、过氧化物酶POD和多酚氧化酶PPO的活性[15],提高了寄主植物的抗性。在前期的研究中,本实验室以松枯梢病原松球壳孢菌(Sphaeropsis sapinea)为目标菌株,从土壤真菌中分离筛选到1株对松球壳孢菌具有良好抑菌效果的森吉木霉菌株M75。为明确森吉木霉M75对松球壳孢菌的抑制作用机理,本研究从森吉木霉M75对病原菌代谢系统酶活性和电导率、丙二醛含量等方面,探究生防菌森吉木霉M75发酵粗提液对松球壳孢菌的抑菌机制,为松枯梢病的生物防治提供理论依据。
-
松球壳孢菌(Sphaeropsis sapinea)由中国林业科学研究院森林保护重点实验室菌种保藏中心提供。森吉木霉M75由本实验室分离筛选获得,现保藏于中国林业微生物菌种保藏管理中心,保藏编号:CFCC54490。
-
将保藏的M75菌株接种于PDA培养基上,28℃活化5 d,用打孔器选取直径5 mm菌饼,接入装有250 mL PDB培养液的三角瓶中,180 r·min−1、28 ℃振荡培养4 d,10000 r·min−14℃离心10 min,取上清经0.45 μm无菌微孔滤膜过滤,得到发酵粗提液,于4 ℃冰箱中保存备用。
-
将保藏的松球壳孢菌菌株接种于PDA培养基上,28 ℃活化5 d,用打孔器选取直径5 mm菌饼,接入装有250 mLPDB培养液的三角瓶中,160 r·min−1、28 ℃振荡培养3~4 d,将产生的直径约为2 cm的病原菌菌丝团用无菌水洗净后置于20 mL含20%发酵粗提液的无菌水溶液中,静置2、4、6、8、10、12、24、48、72、96 h后测定松球壳孢菌各项代谢酶活性和生理指标。以不添加发酵粗提液作为空白对照,每个时间段处理重复3次,指标测定时技术重复3次。
-
测定的病原菌代谢系统酶包括:保护酶系统中的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、多酚氧化酶(PPO);糖酵解途径中的己糖激酶(HK)、丙酮酸激酶(PK)和乳酸脱氢酶(LDH);三羧酸循环中的琥珀酸脱氢酶(SDH)和苹果酸脱氢酶(MDH);辅酶Ⅰ含量和ATP酶。酶活性测定试剂盒由南京建成生物工程研究所提供。
生理指标包括电导率、丙二醛(MDA)含量2个指标。电导率测定使用电导仪测定[16],丙二醛含量测定采用硫代巴比妥酸法[17]。
-
采用Excel 2010软件对实验数据进行统计和分析,绘图使用Origin 2018软件。
-
如图1所示,经森吉木霉M75粗提液处理的前8 h内,与对照组相比,松球壳孢菌代谢系统SOD酶活性显著上升,并在处理8 h时达到峰值,此时酶活性为72.717 U·(g·min)−1。说明病原菌在粗提液的处理下,前期SOD酶活性增加以达到保护菌体的作用。处理8 h后,SOD酶活性大幅降低,在处理96 h时酶活性最低,为1.510 U·(g·min)−1,说明随着处理时间增加,SOD酶活性不断降低。对照组在前8 h内SOD酶活性也呈上升趋势,8 h时酶活性达37.167 U·(g·min)−1,显著低于同时段处理组酶活性。但8 h后,对照组SOD酶活性呈平稳变化趋势,无大幅下降,48 h时达峰值62.180U·(g·min)−1,在处理96 h时酶活性为52.985 U·(g·min)−1。

图 1 森吉木霉M75粗提液对松球壳孢菌保护系统酶活性的影响
Figure 1. Effects of the crude extract of Trichoderma songyi M75 on pathogen protective enzyme activities
经森吉木霉M75粗提液处理的前12 h内,CAT酶活性快速上升并在12 h时达到峰值,0.067 U·(g·s)−1,说明发酵粗提液导致松球壳孢菌代谢系统中过氧化氢含量增加,但处理12 h后酶活性大幅下降,并逐渐趋于零,在处理96 h时为0.004 U·(g·s)−1,是由于SOD酶活性逐渐降低,及膜脂过氧化严重,导致CAT酶活性不断降低。而对照组在48 h内CAT酶活性都处于上升状态,在48 h时达到峰值0.064 U·(g·s)−1,在之后随时间增加酶活性缓慢下降,96 h时为0.054 U·(g·s)−1,且在24 h后对照组CAT酶活性始终高于处理组。
在经森吉木霉M75粗提液处理的前12 h内,与对照组相比,松球壳孢菌代谢系统POD酶活性显著上升,在12 h时达到峰值,为5.733 U·(g·s)−1,在12 h后,处理组酶活性大幅下降,并在48 h后趋于平稳。对照组POD酶活性一直呈上升趋势。PPO酶活性与POD酶活性变化趋势基本一致,处理组在10 h时达到酶活性峰值,11.909 U·(g·s)−1,此后酶活性大幅快速下降,并在处理96 h时达到最低值0.046 U·(g·s)−1。
-
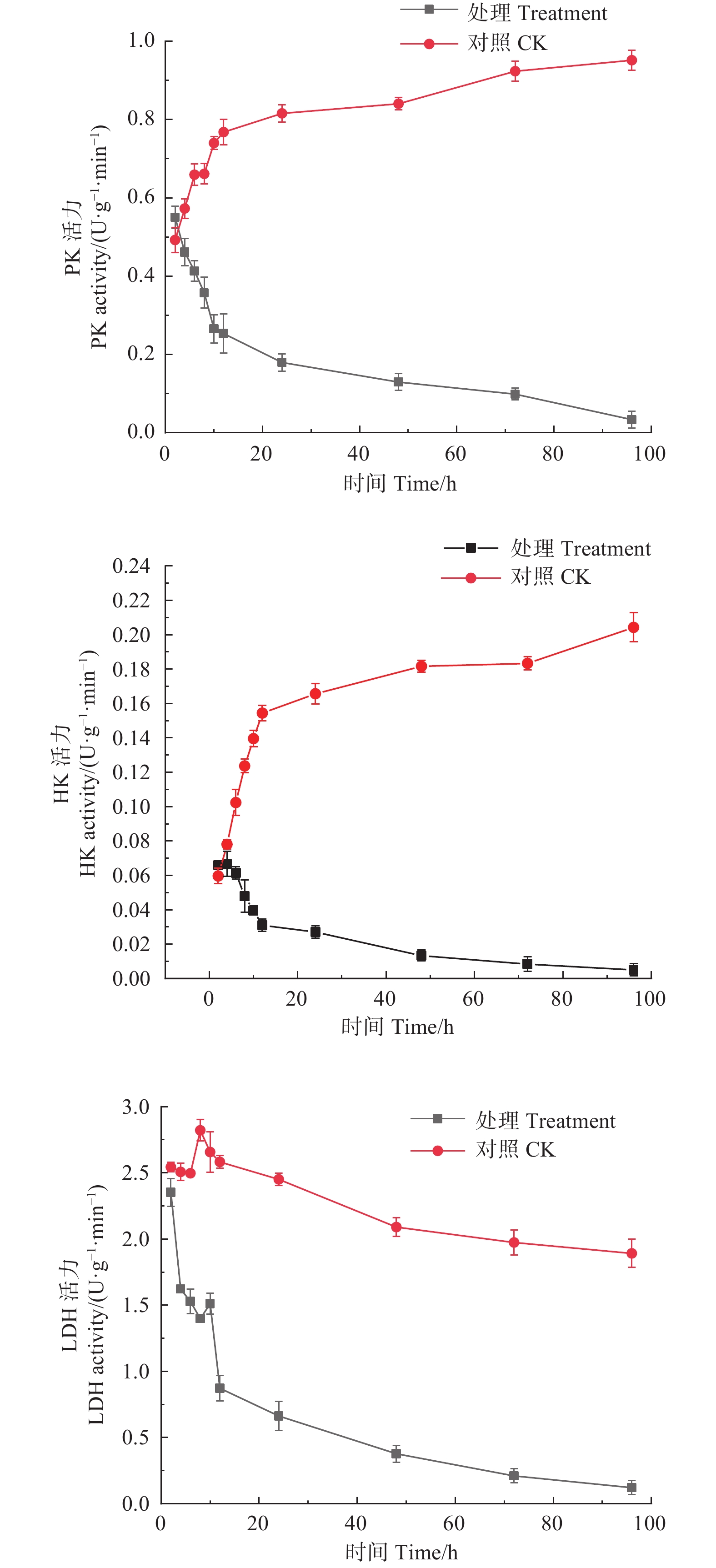
如图2所示,经M75发酵粗提液处理后,松枯梢病原菌松球壳孢菌菌丝团的PK、HK、LDH均随时间增加呈下降趋势,PK酶活性从初始2 h时的0.550 U·(g·min)−1,至96 h时下降至0.033 U·(g·min)−1。HK酶活性从初始2 h时的0.066 U·(g·min)−1,至96 h时下降至0.005 U·(g·min)−1。LDH酶活性由初始2 h的2.351 U·(g·min)−1,至96 h时下降至0.122 U·(g·min)−1。而对照组的PK、HK活力都随时间增加呈持续上升趋势,PK酶活性由初始2 h的0.492 U·(g·min)−1,至96 h时上升至0.952 U·(g·min)−1;HK酶活性由初始2 h的0.060 U·(g·min)−1,至96 h时上升至0.204 U·(g·min)−1;对照组的LDH酶活性整体趋于平稳,且酶活性始终高于处理组,说明M75发酵粗提液对糖酵解途径中的PK、HK及LDH的合成产生了抑制作用,严重干扰了其正常代谢。
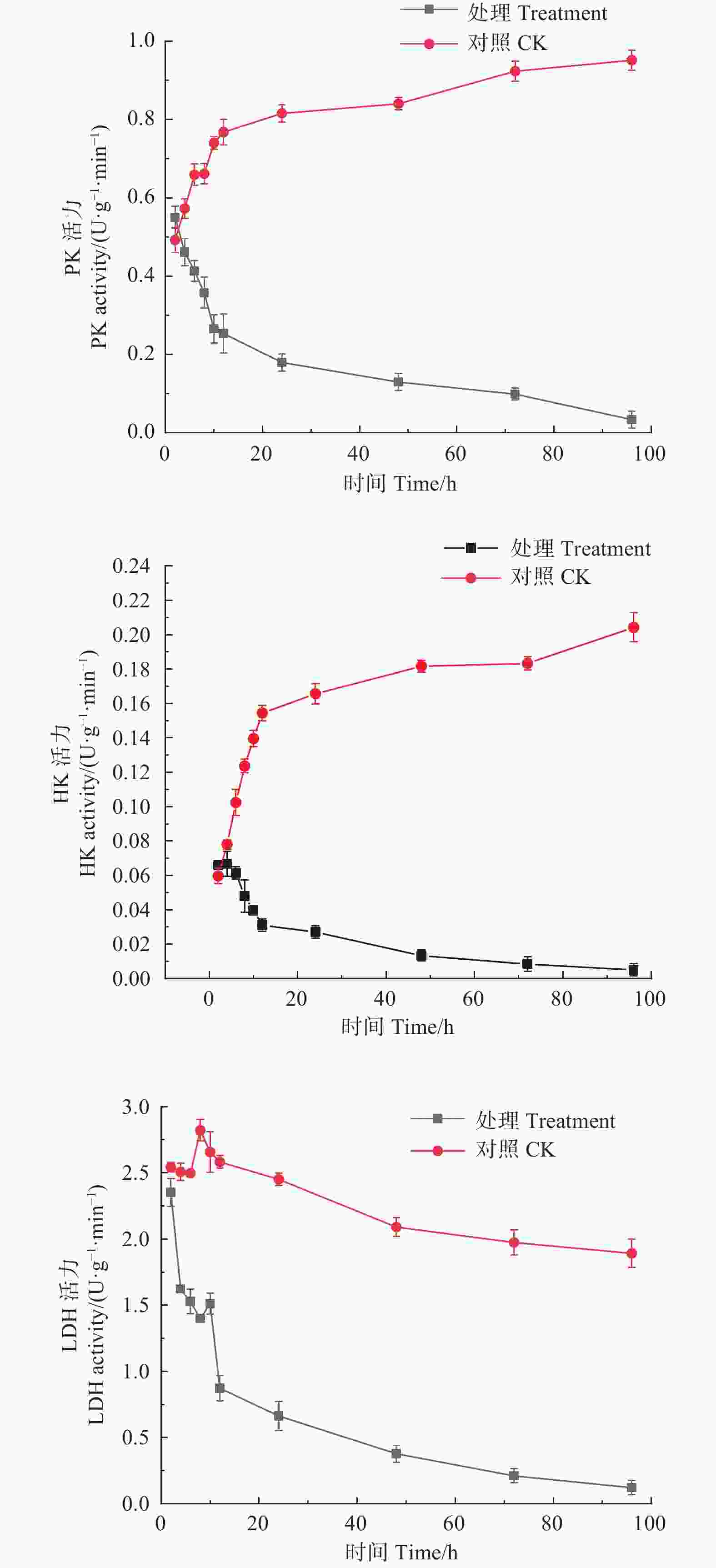
图 2 森吉木霉M75粗提液对松球壳孢菌糖酵解途径中关键酶活性的影响
Figure 2. Effects of the crude extract of Trichoderma songyi M75 on pathogen key enzyme activities in sugar metabolic pathways
-
琥珀酸脱氢酶SDH是细胞能量代谢的重要酶类,其活性变化可直接反应细胞能量代谢状况[18]。如图3所示,经生防菌M75粗提液处理后,松球壳孢菌菌丝团的MDH与SDH酶活性在处理后的12 h内急速下降,MDH酶活性由初始2 h时的15.550 U·(g·min)−1下降至24 h时的1.275 U·(g·min)−1,之后下降趋势变缓,但仍随时间呈逐渐下降趋势,在96 h时为0.196 U·(g·min)−1。SDH酶活性在处理12 h内由初始2 h的30.327 U·(g·min)−1急速下降至9.997 U·(g·min)−1,之后下降趋势减缓,处理96 h时为0.948 U·(g·min)−1。而对照组MDH与LDH酶活性则随时间呈逐渐上升趋势。这说明粗提液抑制了病原菌代谢系统中MDH和SDH的正常合成,导致三羧酸循环停滞。
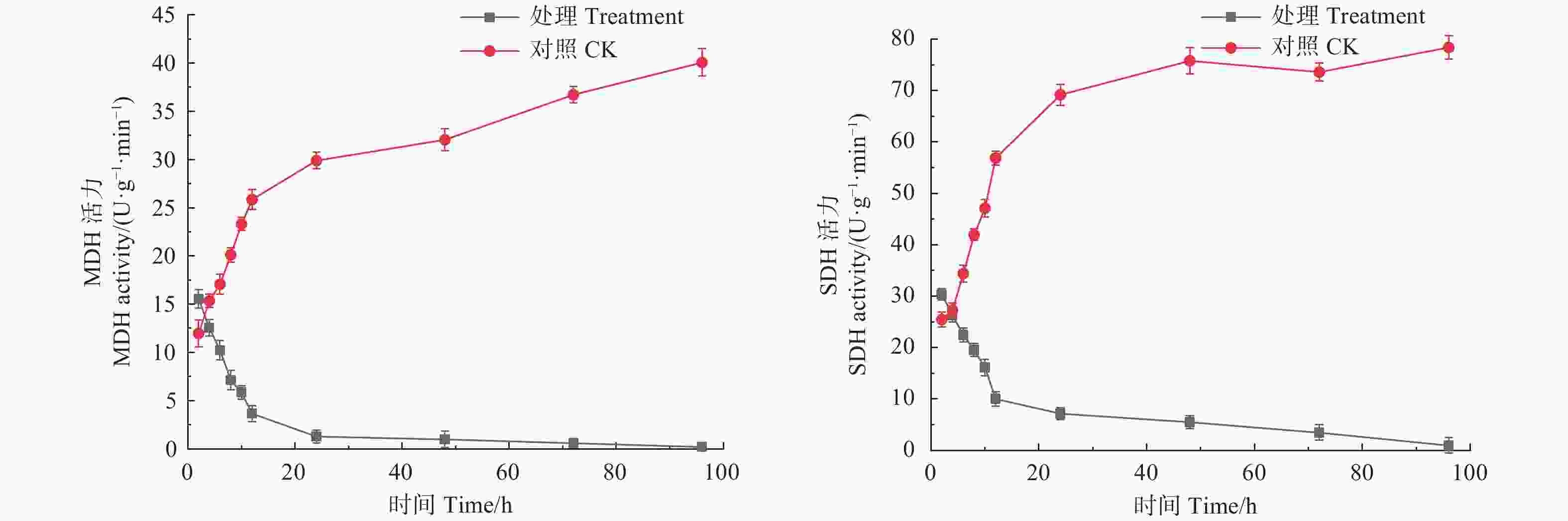
图 3 森吉木霉M75粗提液对球壳孢菌TCA循环中关键酶活性的影响
Figure 3. Effects of the crude extract of Trichoderma songyi M75 on pathogens key enzyme activities in TCA cycle
-
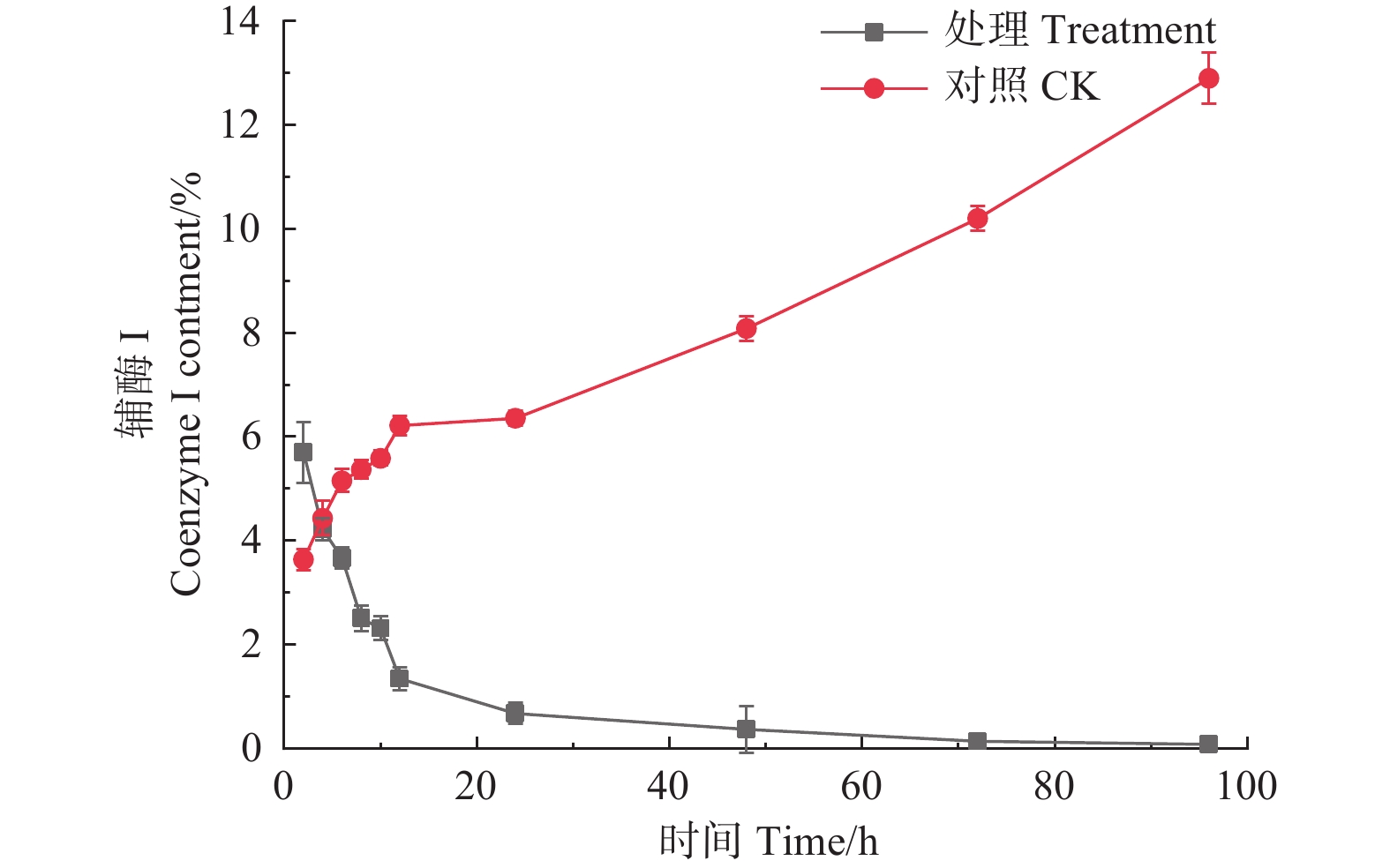
如图4所示,经森吉木霉菌M75发酵粗提液处理后的12 h内,松枯梢病原菌松球壳孢菌菌丝团的辅酶I含量快速下降,由2 h的5.700%下降至1.340%,在之后随处理时间的增加不断趋于零。对照组辅酶I含量则不断增加。
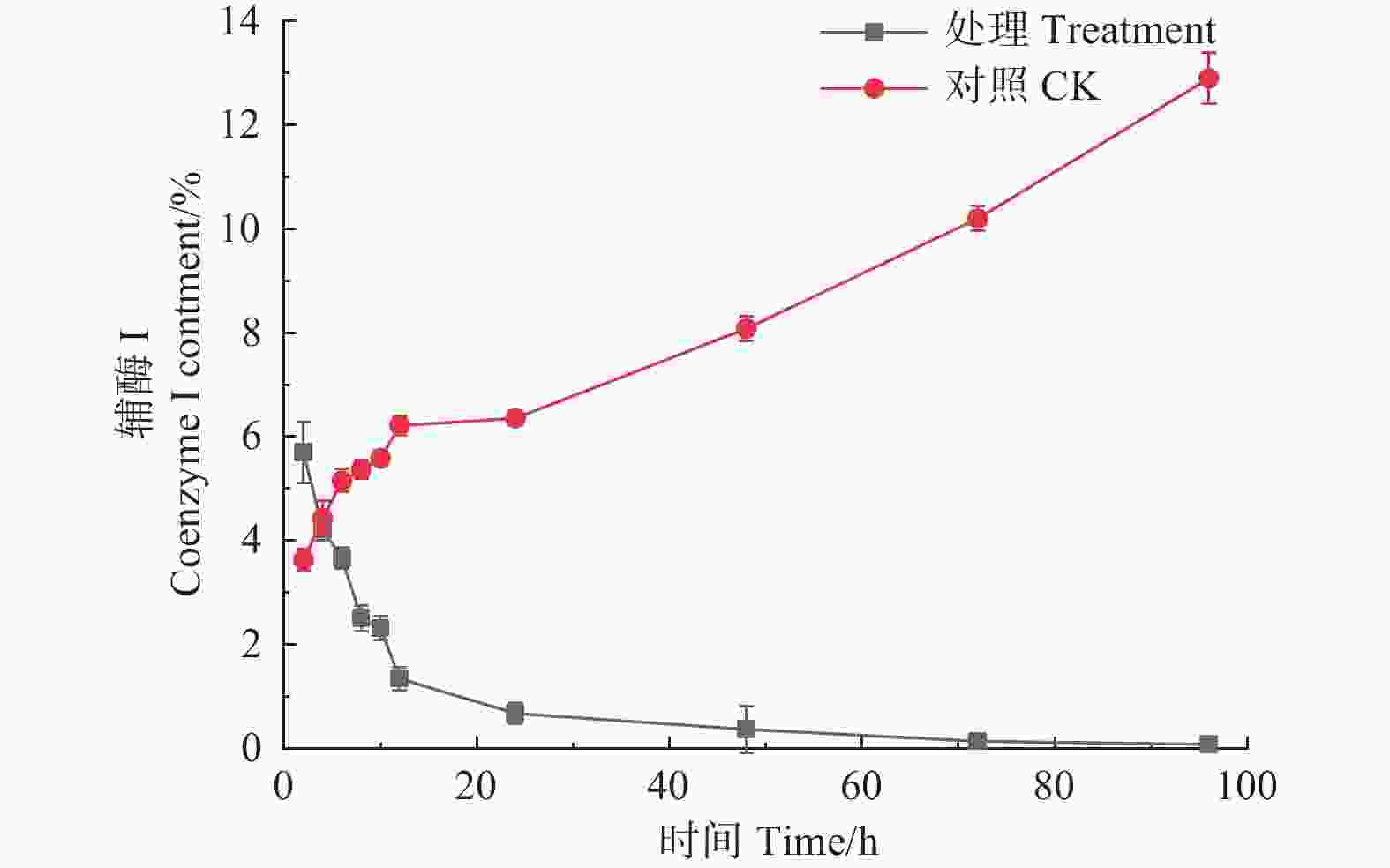
图 4 森吉木霉M75粗提液松球壳孢菌辅酶I含量的影响
Figure 4. Effects of the crude extract of Trichoderma songyi M75 on the content of pathogens enzyme I
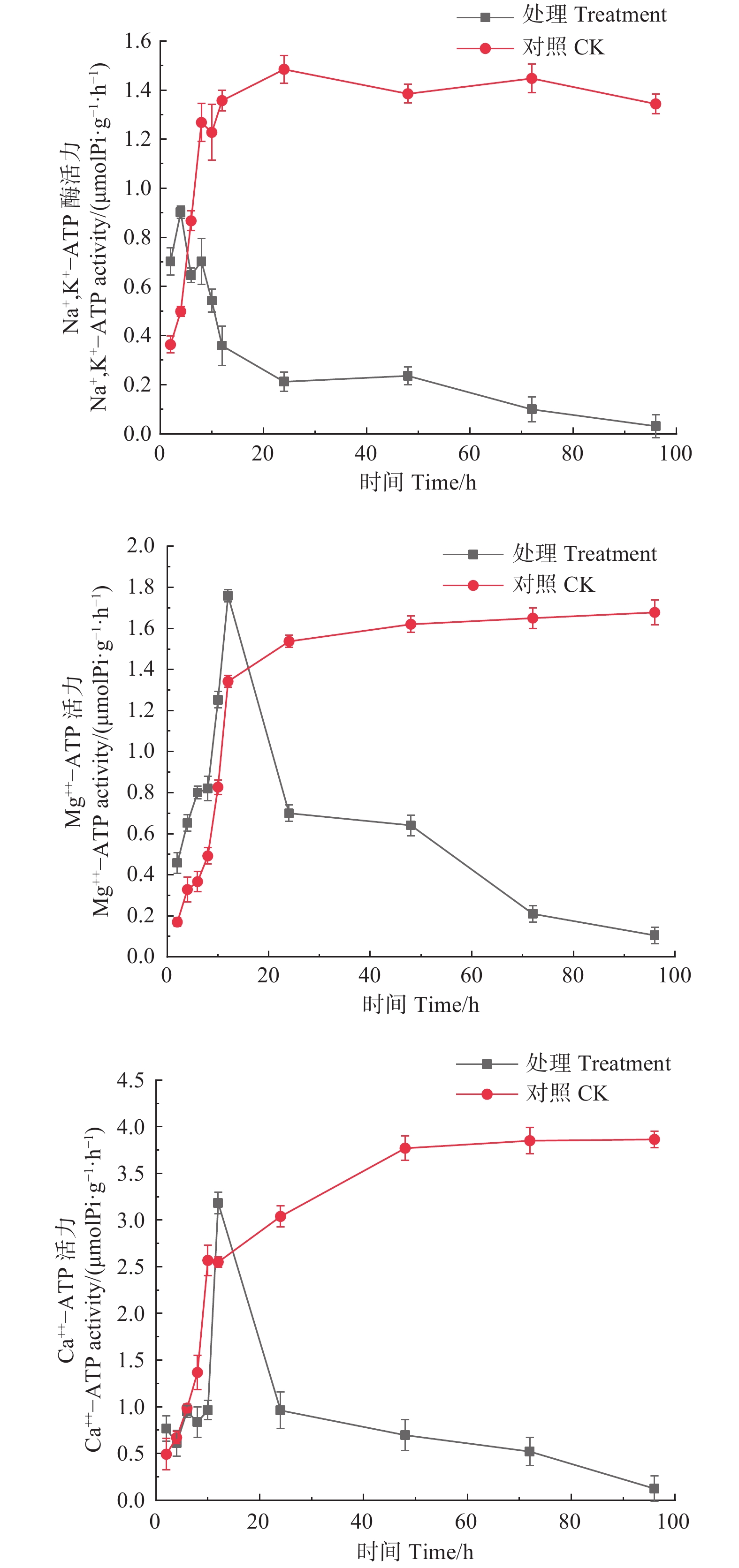
如图5所示,经森吉木霉菌M75发酵粗提液处理的Na+,K+-ATP、Mg2+-ATP、Ca2+-ATP酶活性总体上呈先上升后急速下降的趋势。处理4 h时,Na+,K+-ATP酶活性达到峰值0.902 μmol Pi·(g·h)−1,处理12 h时,Mg2+-ATP、Ca2+-ATP酶活性达到峰值,分别为1.759和3.184 μmol Pi·(g·h)−1,此后急剧下降并逐渐趋于零。对照组酶活性在24 h内处于快速上升趋势,后上升趋缓,但酶活性在10或12 h后高于处理组。
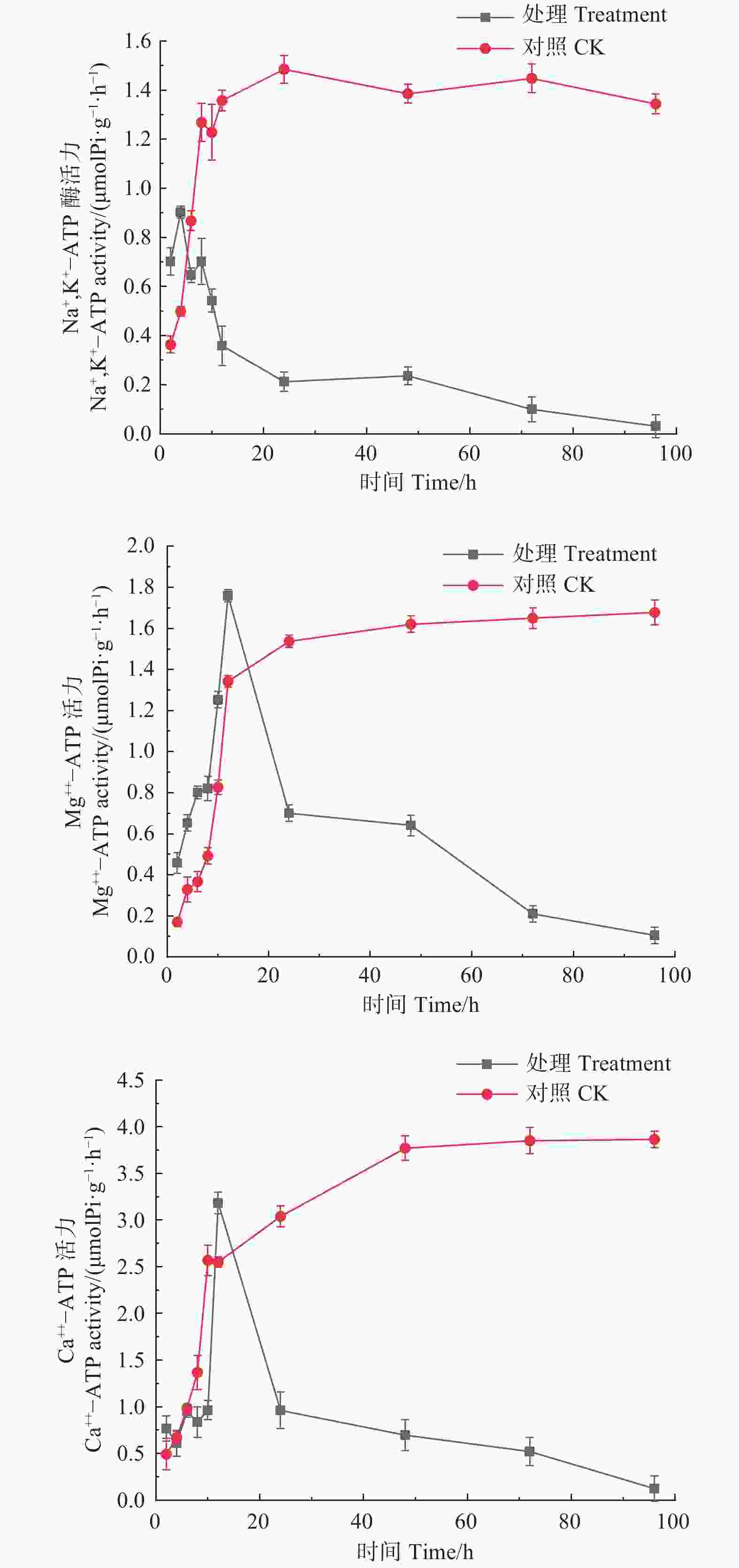
图 5 森吉木霉M75粗提液对松球壳孢菌ATP酶活的影响
Figure 5. Effects of the crude extract of Trichoderma songyi M75 on ATP enzyme activities
-
如图6所示,松球壳孢菌经森吉木菌M75发酵粗提液处理后,电导率随处理时间呈明显增大趋势,并且在各处理时段都远高于对照组,说明经处理的病原菌体电解质泄露,导致电导率值增加。
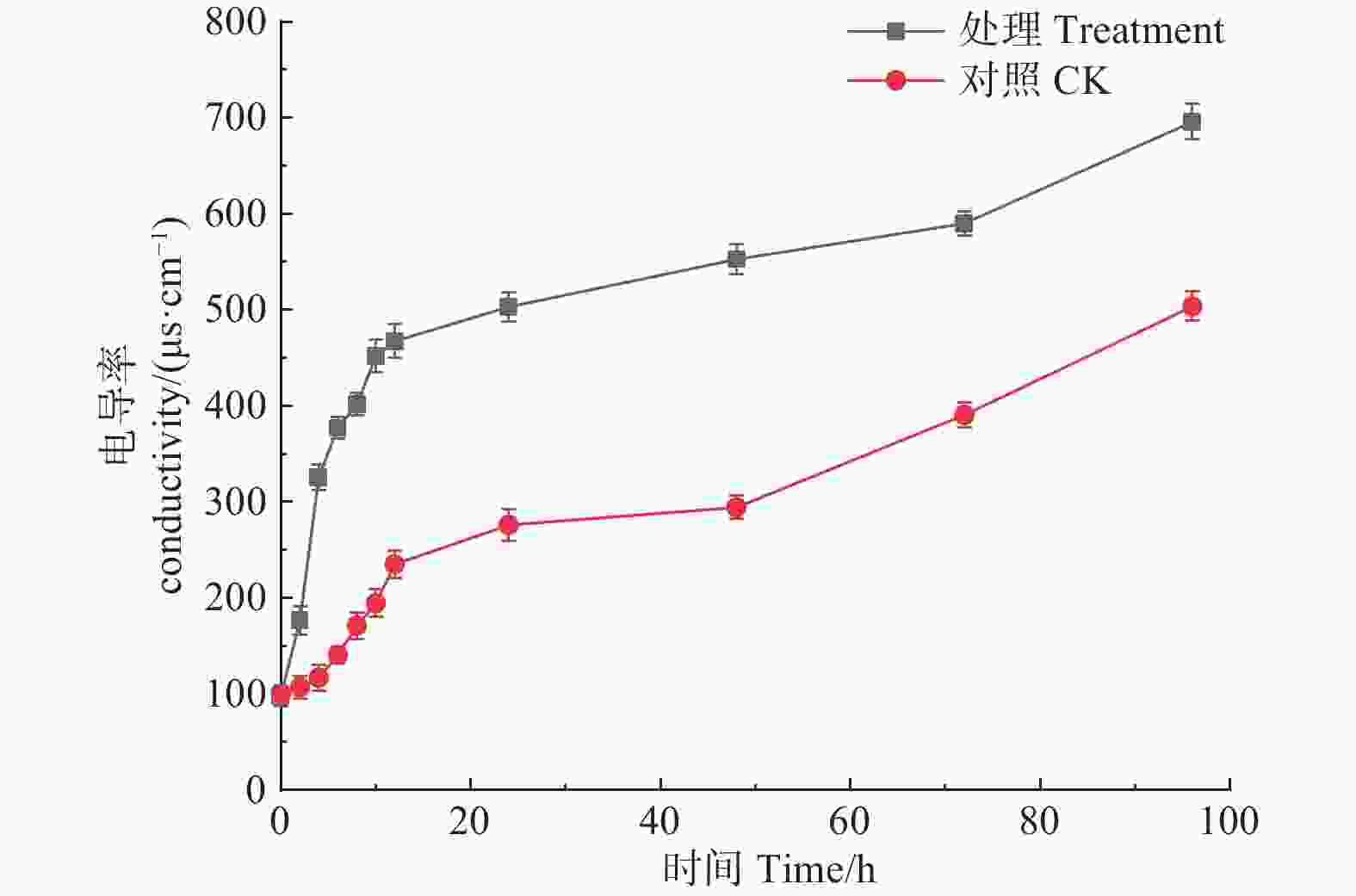
图 6 森吉木霉M75粗提液对球壳孢菌电导率的影响
Figure 6. Effects of the crude extract of Trichoderma songyi M75 on pathogens conductivity
-
如图7所示,经森吉木霉M75粗提液处理后,松球壳孢菌MDA含量在24 h内呈急速增加趋势,在24 h时达到峰值,1.063 nmol·g−1,而后呈下降趋势。对照组MDA含量整体随时间增加呈平稳上升趋势,且对照组MDA含量始终低于处理组。
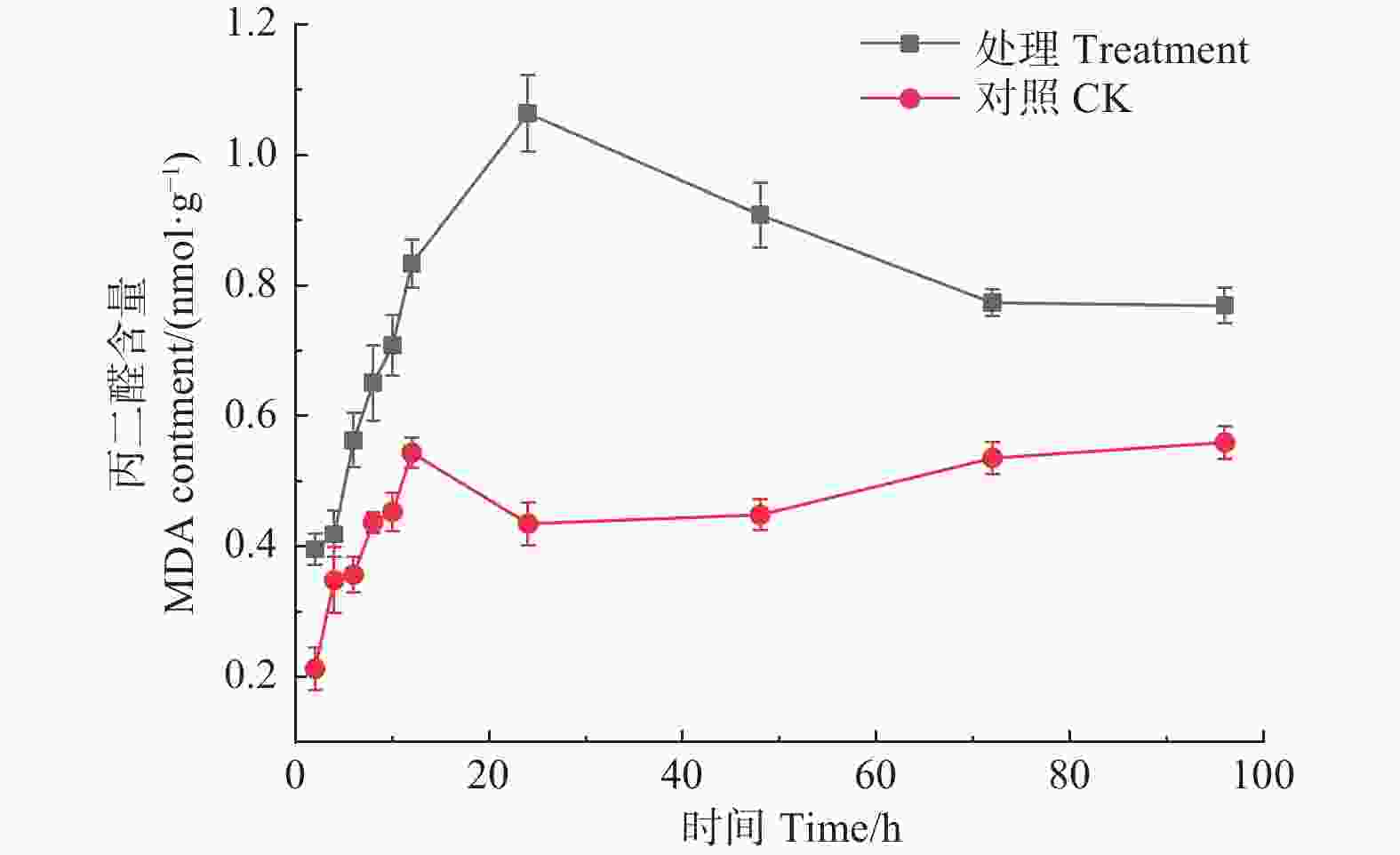
图 7 森吉木霉M75粗提液对松球壳孢菌丙二醛含量的影响
Figure 7. Effects of the crude extract of Trichoderma songyi M75 on pathogens MDA contents
-
木霉菌作为应用最广泛的一类生防真菌,对18个属的29种植物病原真菌表现出显著的抑菌活性[19],生防菌株能够抑制病原菌的生长,必然是破坏了菌体正常的生理代谢途径[20]。森吉木霉M75发酵粗提液对松枯梢病原菌松球壳孢菌的抑制作用主要体现在,发酵粗提液有效抑菌成分可使松球壳孢菌代谢保护酶系统中的SOD、CAT、POD、PPO酶活性先上升后下降,并在处理10 h至12 h后显著低于正常水平,说明随着处理时间增加,氧自由基不断增多,细胞膜脂过氧化加重,导致保护系统酶活性不断降低,影响了病原菌保护酶系统的正常代谢。同时导致糖酵解途径中的HK、PK和LDH酶活性持续下降,影响了松球壳孢菌正常的糖酵解途径[21]。作为三羧酸循环中的关键酶,SDH和MDH代表TCA循环活化状态[22],其酶活数值变化能直接反应细胞能量代谢的情况[23],SDH与MDH活性降低,代表菌体的TCA循环受阻,正常的细胞代谢无法进行。ATP是生命体维持生命活动的能量库,其含量能在一定程度上反应出自身的生理状态[24],辅酶I含量可用于评价糖酵解和TCA循环的强弱,ATP和辅酶I的变化曲线反映出菌体在受到发酵粗提液胁迫时,细胞代谢路径受阻,糖酵解和TCA循环合成减慢,细胞能量供给不足[25]。MDA是细胞膜脂过氧分解的主要产物,MDA的含量可直接反映细胞膜过氧化程度[26],当生物体受到胁迫时,会引起细胞质膜过氧化,使得MDA含量大幅上升[27],本研究中松球壳孢菌MDA含量在处理24 h内急剧上升,且处理组MDA含量始终高于对照组,说明发酵液导致病原菌菌体过氧化程度加剧[28]。细胞内物质成分渗出是菌体膜脂受损的标志,能直接反映菌体细胞膜通透性的变化[29],本研究中松球壳孢菌的电导率随处理时间增加,电导率值不断上升,说明发酵粗提液破坏了菌体细胞膜的完整性,使菌体细胞膜透性改变,导致电解质渗漏[30],抑制了松球壳孢菌的正常生长。本研究中所用为此前分离得到的森吉木霉M75,这是森吉木霉首次应用于生物防治,目前还未见有关森吉木霉菌抑菌机理的报道,本研究对丰富生防木霉菌的资源库具有重要意义。
-
通过对松枯梢病病原菌松球壳孢菌代谢系统中保护酶系统、糖代谢、三羧酸循环、能量代谢中各关键酶和电导率、MDA含量等指标的测定,说明森吉木霉M75发酵粗提液对松枯梢病原松球壳孢菌有较好的抑制作用,可使其代谢系统相关酶活性降低,并导致电解质外渗,电导率升高,MDA大量累积,阻碍松球壳孢菌正常的生理代谢途径,使得病原菌无法进行正常的生理生化反应,从而抑制病原菌的生长。
森吉木霉M75发酵粗提液对松球壳孢菌的抑菌机制研究
Study on the Inhibition Mechanism of Trichoderma songyi Against Sphaeropsis sapinea
-
摘要:
目的 研究森吉木霉M75发酵粗提液对松球壳孢菌Sphaeropsis sapinea代谢系统及生理生化的影响,探究其抑菌机制。 方法 采用分光光度计法,分别测定在不同处理时间段内森吉木霉M75发酵粗提液对松球壳孢菌代谢系统酶活性,以及对电导率、丙二醛含量的影响。 结果 森吉木霉M75发酵粗提液可显著影响松球壳孢菌代谢系统中各关键酶活性。其中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、多酚氧化酶(PPO)在处理8~12 h内达到峰值,为75.25 U·(g·min)−1、0.066 U·(g·s)−1、5.8 U·(g·s)−1和12.33 U·(g·s)−1,分别是初始时的1.96、6.09、16.37和7.86倍;丙酮酸激酶(PK)、己糖激酶(HK)和乳酸脱氢酶(LDH)在处理12~24 h内显著下降,由初始时的0.55、0.066、2.35 U·(g·min)−1下降至0.18、0.03、0.66 U·(g·min)−1,后下降趋缓;琥珀酸脱氢酶(SDH)、苹果酸脱氢酶(MDH)和辅酶Ⅰ含量在处理12 h内呈快速下降趋势,由初始时的30.33 U·(g·min)−1、15.55 U·(g·min)−1、5.70 nmol·(g·min)−1分别下降至10.00 U·(g·min)−1、3.65 U·(g·min)−1、1.34 nmol·(g·min)−1,处理12 h后呈缓慢下降趋势,而对照组则不断升高;Na+,K+-ATP、Mg++-ATP和Ca++-ATP酶活性在处理8 h或12 h时达到峰值,分别为0.09、1.76和3.18 μmolPi·(g·h)−1;此后迅速下降,处理24 h后下降趋缓并逐渐趋于0;松球壳孢菌细胞膜电导率在处理12 h内显著升高,且处理组在各时间段的电导率值始终高于对照组;丙二醛含量在处理24 h内显著升高并达到峰值1.06 nmol·g−1,处理24 h至96 h内呈缓慢下降趋势,但在各处理时间段仍显著高于对照组。 结论 森吉木霉M75发酵粗提液通过破坏松球壳孢菌的防御系统,阻碍了病原菌正常的生理代谢路径,从而抑制病原菌的正常生长。 Abstract:Objecive To study the effects of crude fermentation extract of Trichoderma songyi M75 on the metabolism system, physiological and biochemical mechanisms against the pathogen Sphaeropsis sapinea of pine shoot blight. Method Spectrophotometer was used to determine the effect of the crude fermentation extract of T. songyi M75 on the metabolic system enzyme activity of S. sapinea, as well as the electrical conductivity and the content of malondialdehyde in different treatment periods. Result The crude fermentation extract of T. songyi M75 significantly reduced the activity of the key enzymes in the metabolic system of S. sapinea. The treatment of SOD, CAT, PPO and POD reached their peaks within 8 to 12 hours, which were 75.25 U·(g·min)−1, 0.066 U·(g·s) −1, 5.8 U·(g·s) −1 and 12.33 U·(g·s) −1, respectively, and were 1.96, 6.09, 16.37 and 7.86 times of the initial. PK, HK and LDH decreased significantly within 12 to 24 hours of treatment, from 0.55, 0.066, 2.35 U·(g·min)−1 at the initial time to 0.18, 0.03, 0.66 U·(g·min)−1 respectively, and then the downward trend gradually slowed down. The contents of SDH, MDH and coenzyme Ⅰ decreased rapidly from 30.33 U·(g·min)−1, 15.55 U·(g·min)−1 and 5.70 nmol·(g·min)−1 to 10.00 U·(g·min)−1, 3.65 U·(g·min)−1 and 1.34 nmol·(g·min)−1 respectively, and then the decline was slow and the control group was in the continuous increasing trend. The activities of Na+, K+-ATP, Mg++-ATP和Ca++-ATP peaked at 0.09, 1.76 and 3.18μmolPi·(g·h) −1 at the 8th or the 12th hours, after that, it decreased rapidly, and then decreased slowly and gradually to 0 after 24 hours of treatment. The conductivity increased significantly within 12 hours, and the value of the treatment group was always higher than that of the control. MDA increased significantly in 24 hours and reached the peak value of 1.06 nmol·g −1, and decline slowly from the 24th to the 96th hours, however, it was still significantly higher than that of the control in each treatment period. Conclusion The crude fermentation extract of T. songyi M75 can inhibit the normal growth of the S. sapinea by destroying the defense system and normal metabolic pathway of the fungus. -
Key words:
- Trichoderma songyi
- / extract
- / Sphaeropsis sapinea
- / antibacterial mechanism
-
图 1 森吉木霉M75粗提液对松球壳孢菌保护系统酶活性的影响
Figure 1. Effects of the crude extract of Trichoderma songyi M75 on pathogen protective enzyme activities
图 2 森吉木霉M75粗提液对松球壳孢菌糖酵解途径中关键酶活性的影响
Figure 2. Effects of the crude extract of Trichoderma songyi M75 on pathogen key enzyme activities in sugar metabolic pathways
图 3 森吉木霉M75粗提液对球壳孢菌TCA循环中关键酶活性的影响
Figure 3. Effects of the crude extract of Trichoderma songyi M75 on pathogens key enzyme activities in TCA cycle
图 4 森吉木霉M75粗提液松球壳孢菌辅酶I含量的影响
Figure 4. Effects of the crude extract of Trichoderma songyi M75 on the content of pathogens enzyme I
图 5 森吉木霉M75粗提液对松球壳孢菌ATP酶活的影响
Figure 5. Effects of the crude extract of Trichoderma songyi M75 on ATP enzyme activities
图 6 森吉木霉M75粗提液对球壳孢菌电导率的影响
Figure 6. Effects of the crude extract of Trichoderma songyi M75 on pathogens conductivity
-
[1] 叶建仁, 贺 伟. 林木病理学[M]. 北京: 中国林业出版社, 2011: 224-227. [2] 唐 旭. 松枯梢病拮抗细菌的筛选及其抗病机制初探[D]. 南京: 南京林业大学, 2017. [3] 满晓国. 松树枯梢病的发生原因及防治技术[J]. 农村实用科技信息, 2014(1):30. [4] Juanita de Wet, Treena Burgess, Bernard Slippers, et al. Multiple gene genealogies and microsatellite markers reflect relationships between morphotypes of Sphaeropsis sapinea and distinguish a new species of Diplodia[J]. Mycological Research, 2003, 107(5): 557-566. doi: 10.1017/S0953756203007706 [5] 刘满光, 张 娣, 路文雅, 等. 松枯梢病病原菌的研究进展[J]. 中国农学通报, 2018, 34(27):97-101. doi: 10.11924/j.issn.1000-6850.casb17110042 [6] 李文辉. 松蚜虫和松树枯梢病防治技术[J]. 农业与技术, 2015, 35(18):138. [7] 卢颖超. 樟子松枯梢病的发生及其预防[J]. 吉林农业, 2017(11):82. [8] 黄华毅. 杨树炭疽病菌内生拮抗细菌抑菌活性研究[D]. 北京: 北京林业大学, 2017. [9] Hermosa R, Viterbo A, Chet I, et al. Plant-beneficial effects of Trichoderma and of its genes[J]. Microbiology, 2012, 158(1): 17-25. doi: 10.1099/mic.0.052274-0 [10] Shoresh M, Yedidia I, Chet I. Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203[J]. Phytopathology, 2005, 95(1): 76-84. doi: 10.1094/PHYTO-95-0076 [11] Harman G E. Overview of mechanisms and uses of Trichoderma spp[J]. Phytopathology, 2006, 96(2): 190-194. doi: 10.1094/PHYTO-96-0190 [12] Cheng C H, Yang C A, Peng K C. Antagonism of Trichoderma harzianum ETS 323 on Botrytis cinerea mycelium in culture conditions[J]. Phytopathology, 2012, 102(11): 1054-1063. doi: 10.1094/PHYTO-11-11-0315 [13] Sawant I S, Wadkar P N, Ghule S B, et al. Enhanced biological control of powdery mildew in vineyards by integrating a strain of Trichoderma afroharzianum with sulphur[J]. Biological Control, 2017, 114: 133-143. doi: 10.1016/j.biocontrol.2017.08.011 [14] Diego H Z, Elibeth F P, Aida A R et al. Trichoderma asperellum, a potential biological control agent of Stemphylium vesicarium, on onion (Allium cepa L.)[J]. Biological Control., 2020, 140: 104105. doi: 10.1016/j.biocontrol.2019.104105 [15] Sunpapao A, Chairin T, Ito S I. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings[J]. Biological Control, 2018, 123:36-42. [16] 黄运红, 任雨涵, 邹 龙, 等. 炭样小单孢菌产抗生素对水稻细菌性条斑病菌的抗菌作用及其机理[J]. 微生物学通报, 2019, 46(5):85-94. [17] 李 凤, 刘会涛, 田 莎, 等. 复配芽孢杆菌对撑绿杂交竹枯萎病的生物防治效果研究[J]. 北京林业大学学报, 2018, 40(12):76-84. [18] 李丽萍, 谢响明, 宋洪英, 等. 紫茎泽兰提取物对番茄青枯菌的抑菌作用及其机理[J]. 生物技术通报, 2010, 7:146-152. [19] 郑柯斌, 林 海, 周 沙, 等. 海洋生境棘孢木霉TCS007菌株的鉴定及抑菌活性[J]. 农药学学报, 2020, 22(5):801-807. [20] 杨 蕾, 周国英, 梁 军. 杨树溃疡病生防菌株的抑菌机理研究[J]. 植物保护, 2016, 42(2):47-54. doi: 10.3969/j.issn.0529-1542.2016.02.008 [21] 李冲伟, 杨立宾, 邓 勋, 等. 木霉菌株对金黄壳囊孢菌的抑菌效应及机制[J]. 林业科学, 2012, 48(9):88-94. doi: 10.11707/j.1001-7488.20120914 [22] 郭聪聪, 沈永宝, 史锋厚. 白皮松种子萌发过程中呼吸代谢和内源激素对温度变化的响应[J]. 中南林业科技大学学报, 2021, 41(3):25-36. [23] 廖石榴, 尹 维, 廖晓兰, 等. 苋菜乙酸乙酯提取物对柑橘溃疡病菌的抑制及机理研究[J]. 湖南农业大学学报: 自然科学版, 2017, 43(5):544-550. [24] 王艳娜, 王贵禧, 梁丽松, 等. 低温及轮纹病菌胁迫对鸭梨果肉ATP含量及H+-ATPase, Ca2+-ATPase活性的影响[J]. 林业科学, 2008, 44(12):72-76. doi: 10.3321/j.issn:1001-7488.2008.12.013 [25] 王艳玲, 尚敏敏, 张紊玮, 等. 假单胞菌YL11对扩展青霉的抑制作用及其机理初探[J]. 微生物学通报, 2019, 46(5):114-124. [26] 韩庆典, 胡晓君, 黄坤艳, 等. 小麦白粉病菌对小麦幼苗MDA含量及防御酶活性的影响[J]. 分子植物育种, 2016, 14(10):2803-2811. [27] 肖生鸿, 邓倩仪, 黄洁婷, 等. 汞胁迫对桉树3个保护酶活性及基因表达的影响[J]. 西北林学院学报, 2021, 36(2):91-96. doi: 10.3969/j.issn.1001-7461.2021.02.13 [28] 杨立宾, 宋瑞清, 李冲伟. 哈茨木霉发酵液乙酸乙酯提取物对致病疫霉生理指标的影响[J]. 北京林业大学学报, 2013, 35(2):92-96. [29] Kong M, Xi G C, Xing K, et al. Antimicrobial properties of chitosan and mode of action: a state of the art review[J]. International Journal of Food Microbiology, 2010, 144(1): 51-63. doi: 10.1016/j.ijfoodmicro.2010.09.012 [30] 蓝蔚青, 杨 歆, 王 蒙, 等. 壳聚糖对腐生葡萄球菌的作用机制[J]. 广东海洋大学学报, 2021, 41(1):63-71. doi: 10.3969/j.issn.1673-9159.2021.01.009 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3359
- HTML全文浏览量: 2080
- PDF下载量: 45
- 被引次数: 0
