





-
自工业革命以来,大气中CO2等温室气体浓度的不断增加引起气候变化和全球增温,从而使人们开始关注陆地生态系统的碳循环过程[1]。森林生态系统是陆地生态系统的主体,其占比陆地生态系统的31%[2-3],其作为陆地生态系统的重要碳汇[4],对实现碳中和起到极为关键的作用。因此,对于明晰森林生态系统的碳交换量的变化显得尤为重要。
大气湍流是生态系统和大气进行气体和能量交换的主要媒介,涡协方差相关法的诞生就是为了记录这一过程的物质和能量的量[5],是目前测定大气与陆地生态系统之间气体和能量交换的最直接的方法[6-7]。在实际应用过程中,涡度相关技术的监测受到多种环境因素的影响,如生态系统类型、观测高度、下垫面地形、大气因素等都会影响其监测结果的准确性[5,8]。需要通过对数据处理后的能量闭合和源区分析对其的适用性和准确性进行验证。许多学者都利用涡度相关技术,对复杂的森林生态系统与大气之间的CO2交换进行过探究[9-10],同时针对不同类型的生态系统和站点进行比较分析。目前大部分的研究表明,森林生态系统为碳汇,只有一小部分的研究得出的结论为碳源且包括幼龄林在内[11-12]。尽管森林生态系统的碳汇作用得到了极大多数学者的广泛认可,但由于环境因子的影响,使得全球森林生态系统与大气间的CO2交换量在不断变化,所以,对于森林生态系统与大气之间的碳交换监测依旧重要[13]。
重庆缙云山地处三峡库区的尾端,是重要的生态屏障,其中针阔叶混交林是该地区的典型林分之一,也是我国西南山地森林生态系统的典型代表。王倩、王杰帅等都曾利用二次坐标旋转对其碳通量进行分析[10, 14]。本研究以2019.11—2020.10缙云山针阔叶混交林通量监测数据为基础,坐标旋转处理上采用平面拟合法,通过湍流分析,能量闭合和源区分析,来对涡度相关对其的适用性进行探究,同时对该地区碳通量的时间变化进行分析。以期为预测未来碳通量变化下的森林动态和经营提供科学依据。
-
研究区所在地为重庆市北碚区缙云山国家级自然保护区国家定位观测研究站内(106°17'~106°24' E、29°41'~29°52' N),该保护区的整体面积约7 600 hm2,海拔200~952 m,地处三峡库区的尾端,是三峡库区的重要生态屏障。缙云山气候为典型的亚热带季风性湿润气候,降雨充沛,年平均降水1 611.8 mm,主要集中在生长季(4—10月);年平均蒸发量 777.1 mm;该地区受局部小气候影响潮湿多雾,平均全年有雾日达89.8 d。研究区为针阔叶混交林森林生态系统,主要的优势树种为四川山矾(Symplocos setchuensis Brand)、广东山胡椒(Lindera kwangtungensis Allen)、杨桐(Adinandra bockiana Pritzel ex Diels)、马尾松(Pinus massoniana Lamb)和杉木(Cunninghamia lanceolata Hook)。
-
重庆缙云山通量塔位于观测站的针阔叶混交林内,塔高35 m,配备的观测系统包括两个部分,即开路式涡度相关监测系统和微气象观测系统,具体的仪器布设详见表1。
表 1 通量塔仪器布设
Table 1. Instrument layout of flux tower
观测系统 仪器名称 型号 生产商 安装高度/m 频率/Hz 开路式涡度相关监测系统 开路式气体分析仪 Li-7500 LiCor Inc.,USA 35 10 三维超声风速仪 CAST-3 Campbell,USA 35 10 数据采集器 CR3000 35 10 微气象观测系统 空气温湿度传感器 HMP60 Vaisala, FIN 25
3510 翻斗式雨量筒 TR-525M Texas Electronics, USA 25
3510 光合有效辐射传感器 LI-190SB LiCor Inc.,USA 25
3510
净辐射仪NR01 Hukseflux,NED
3510 风速风向仪 03002 RM Young,CN 35 10 涡度相关法可以直接测定植物群落与大气之间的气体交换通量,其原理是通过测定大气中湍流运动产生的风速脉动和物理量脉动的协方差来测定物质或能量的通量。
大气与植物群落之间的CO2湍流通量的公式为:
$ Fc=\rho \overline{{c}{{{'}}}{\omega }{{{'}}}} $
(1) Fc(mg·m−2·s−1)为CO2的湍流通量,
$ \rho $ c′为CO2密度脉动,ω′为垂直风速的脉动量。 -
原始数据处理主要在EddyPro-7.0.6中进行,具体方法如下:
(1)采用Vicker提出的处理办法对原始数据进行质量检测,并将野点及异常值进行剔除。
(2)采用Mauder和Foken[15]提出的0~2级分级法作为本研究数据的质量分级标准。
(3)坐标校正,考虑到研究区位于山区,下垫面不平稳,且生态系统结构较为复杂,所以采用平面拟合法[16]对其进行校正。
(4)通量校正,主要包括:超声虚温修正、WPL校正、频率响应校正、夜间通量校正。
(5)数据插补,采用由Max Planck Institute for Biogeochemistry开发的REddyProcWeb在线数据处理工具对不连续的通量数据进行插补。
后期的数据处理和制图分别在Excel、MATLAB、Tovi和Origin中进行。
-
根据能量守恒定律,输入生态系统的能量将会以某种形式输出,因此能量的闭合与否是评判通量数据是否可用的衡量标准之一。森林生态系统中的能量闭合(EBR)计算公式为:
$ Rn-G=H + LE + Fs + Q $
(2) Rn为太阳净辐射,G为土壤热通量,H为显热通量,LE为潜热通量,Fs为冠层储热,Q为其他能量输出项。
由于冠层储热和其他能量输出项所占比例很少,且部分数据在测量中存在较大难度,所以大量的学者在研究能量闭合时都将其忽略不计,本研究中只利用(Rn-G)和(H+LE)来进行能量闭合的计算。
-
质量评价是0和1的为高质量通量数据,图中显示为蓝色;质量评价是2的数据为中低质量数据,图中显示为黄色(图1)。图中2019.12.06—2019.12.19数据缺失是由于通量塔断电导致。质量评价为0和1的高质量数据占比为77%,质量评价为2的中低质量数据占比16%,缺失数据为7%。结果表明,77%的数据趋于理想的稳态条件,并且很好的符合湍流观测的发展特点。
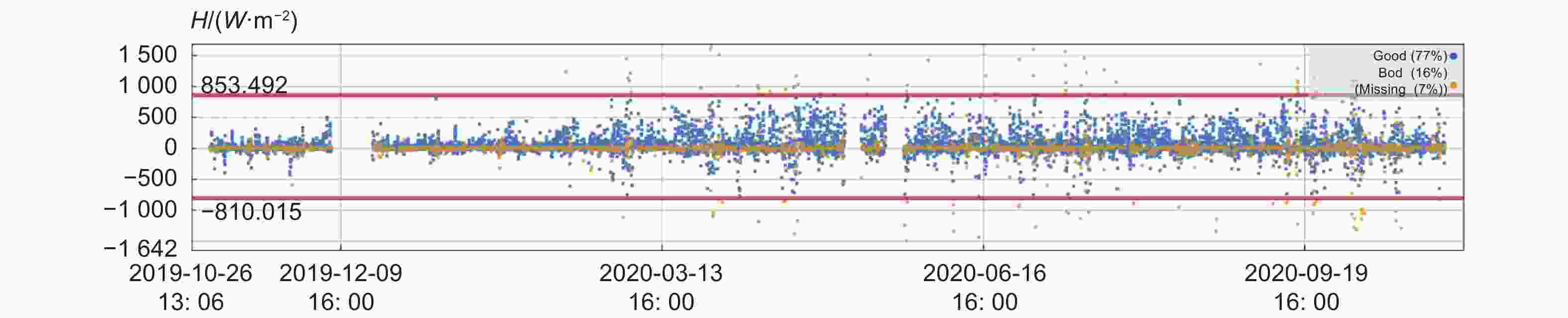
图 1 显热通量质量等级分级
Figure 1. Quality classification of sensible heat flux
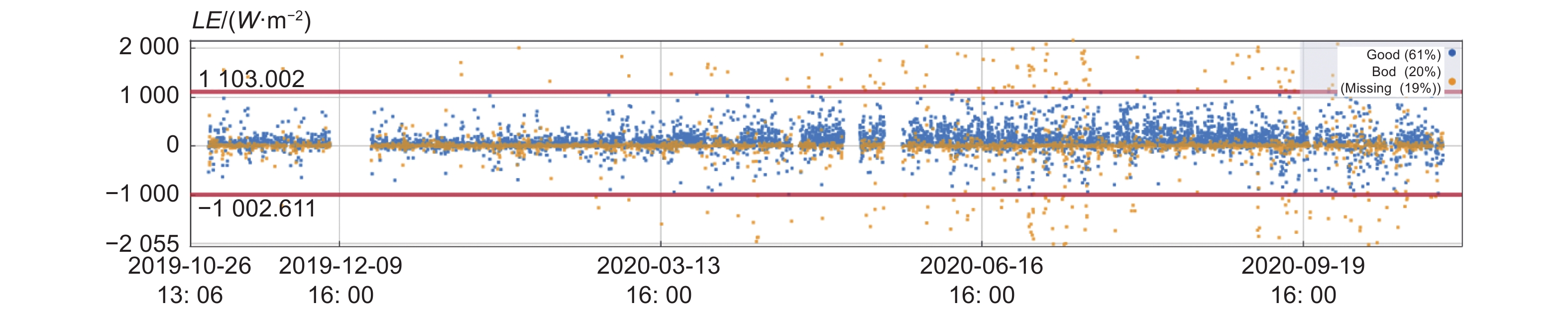
由图2可见,质量评价为0和1的高质量数据占比为61%,质量评价为2的中低质量数据占比20%,缺失数据为19%。结果表明,61%的数据趋于理想的稳态条件。并且很好的符合湍流观测的发展特点。
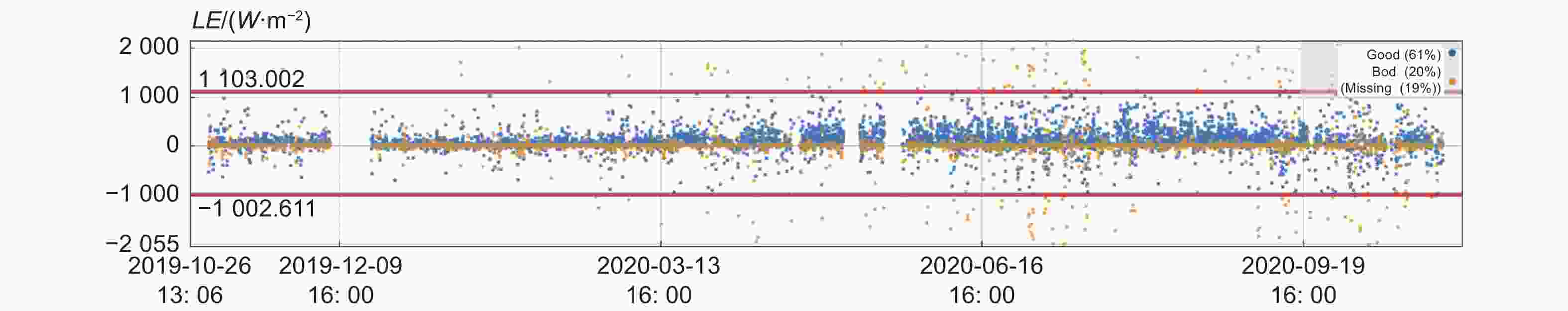
图 2 潜热通量质量等级分级
Figure 2. Quality classification of latent heat flux
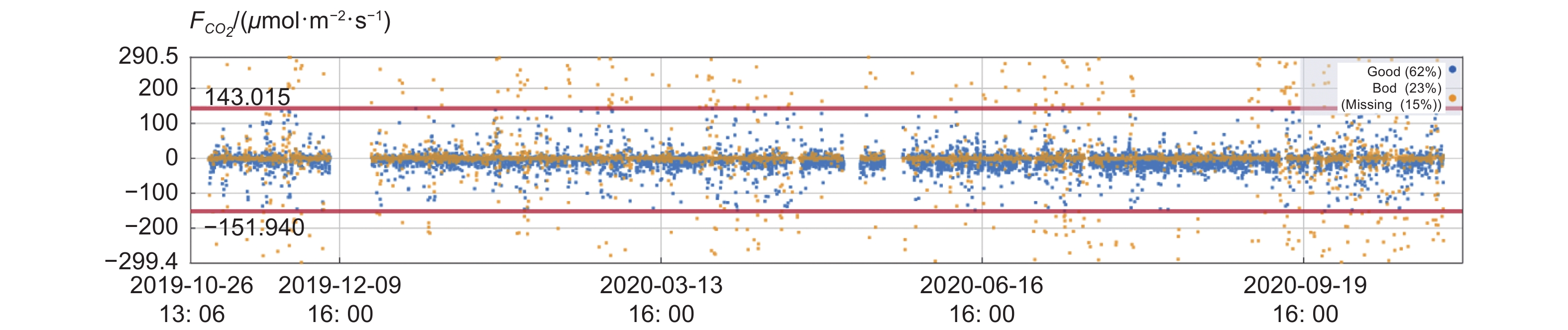
碳通量的数据质量容易受到降水的影响,样地所在地全年多雨(4—10月为雨季),且非雨季时,由于受山地小气候影响,该山地长时期水雾笼罩。从图3可以看出,CO2湍流的高质量数据占比为62%,中低质量数据为23%,缺失数据为15%。符合湍流观测的发展特点。
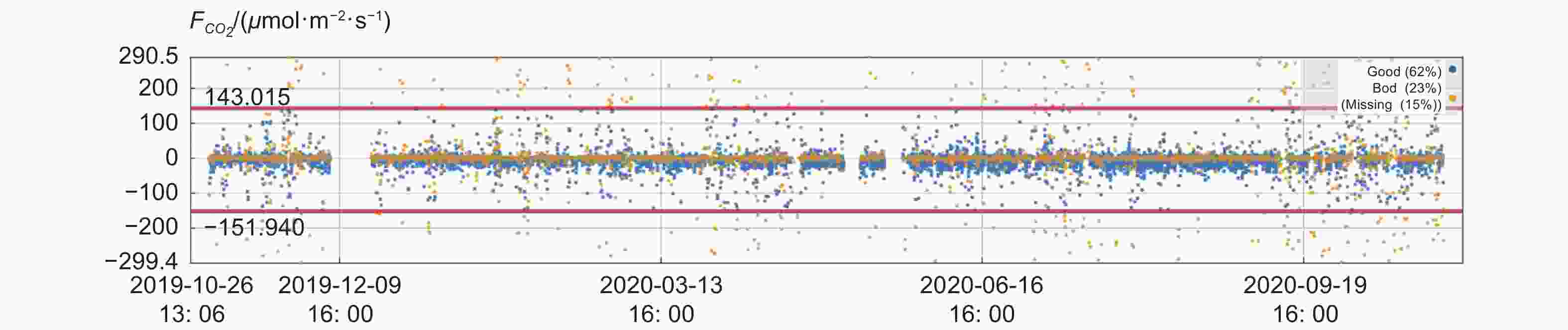
图 3 碳通量质量等级分级
Figure 3. Quality classification of carbon flux
-
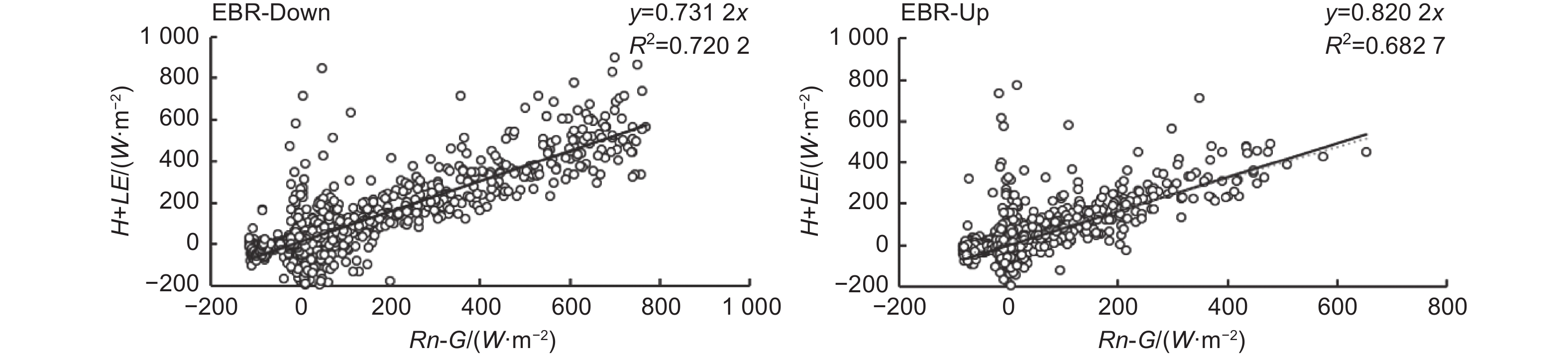
分别对25 m和35 m处的通量观测系统所观测到的生长季通量数据进行了能量闭合分析(图4)。该观测系统下层能量闭合率为0.73,上层能量闭合率为0.82,均在国际认可的能量闭合范围之内(0.7~0.9),表明能量闭合情况良好。
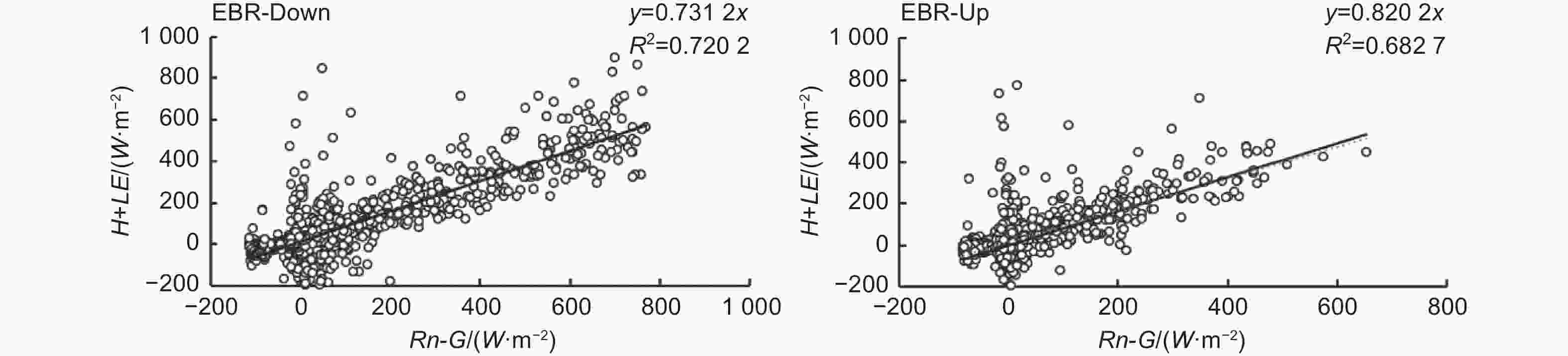
图 4 能量闭合分析
Figure 4. Diagram of energy closure analysis
-
本研究中的通量足迹如图5所示。左图为全年白天通量足迹模拟,右图为全年夜间通量足迹模拟。有色区域即为通量贡献区,颜色越亮代表通量观测的数据来源越密集,闭合圈线代表不同累积通量贡献率的范围。因为通量塔观测的通量数据一般来源于上风向,因此贡献区形状为椭圆,长轴在东北和西南方向,同时可以看出高亮区域大部分集中在东北侧,即全年大部分月份主要通量观测区域来源于东北方。全年风速风向变化如风玫瑰图(图6)所示,主风向来自东北方向,这与通量的高贡献区相吻合,说明通量数据来源可信且质量良好。
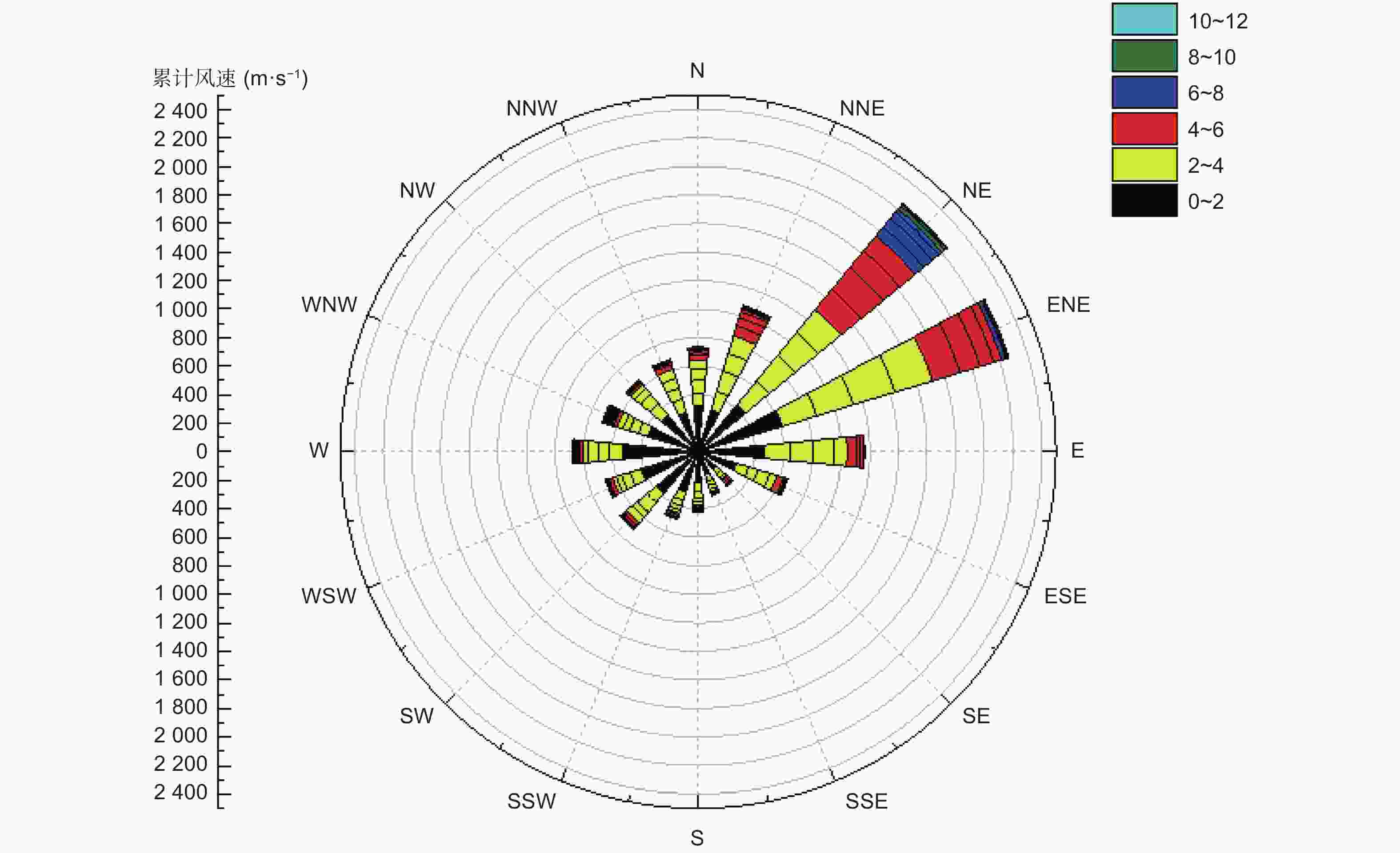
图 5 全年通量足迹(左为白天,右为夜间)
Figure 5. Annual flux footprint (day on the left and night on the right)

图 6 风速玫瑰图
Figure 6. Wind speed rose
通量的峰值年贡献区的年平均距离为82.25 m,10%、30%、50%、70%和90%累计通量贡献区年平均距离分别为28.23、70.31、107.22、150.38 m和225.29 m(表2)。由表2可以看出9月份为贡献区距离最大月,峰值距离为91.12 m,90%累计贡献距离为249.59 m,最小月份为4月,峰值距离为83.43 m,90%累计贡献距离为228.55 m。贡献区距离在正常范围内,且不同月份间相差不大。
表 2 通量贡献区距离
Table 2. Distance of flux contribution area
月份 x_peak/m x_10%/m x_30%/m x_50%/m x_70%/m x_90%/m 4 83.43 28.64 71.32 108.77 152.55 228.55 5 89.26 30.64 76.30 116.36 163.2 244.49 6 80.69 27.70 68.97 105.19 147.53 221.02 7 84.39 28.97 72.14 110.01 154.29 231.16 8 88.32 30.32 75.50 115.55 162.4 244.16 9 91.12 31.28 77.89 118.78 166.6 249.59 10 84.90 29.14 72.57 110.68 155.23 232.56 -
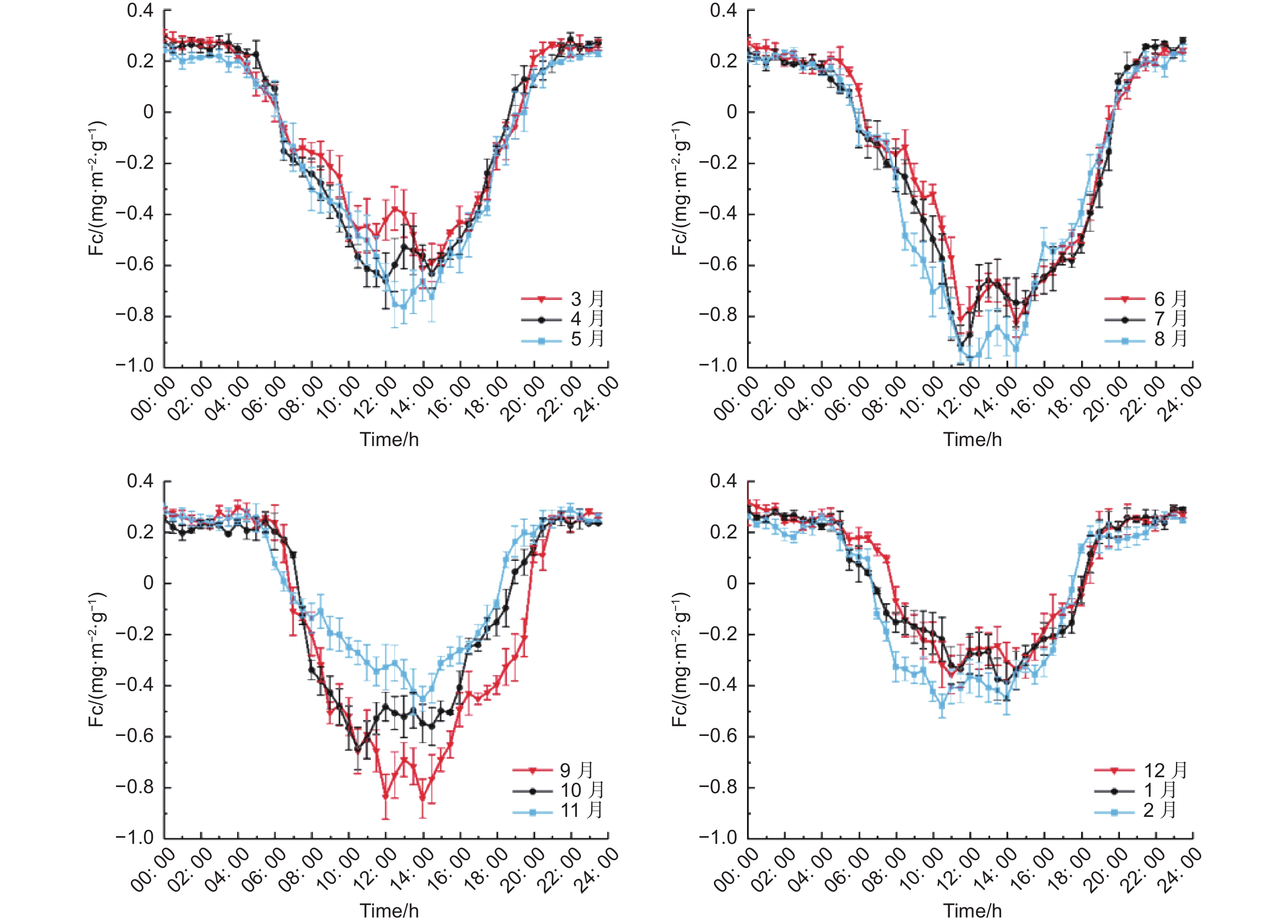
利用MATLAB对全年半小时通量数据进行平均,得到各月份同一时刻下半小时的平均值,得到全年各月同一时刻的Fc的日变化,由图7可以看出,各月份的Fc日变化大体相同,均成U字形,Fc为负值时,说明生态系统在吸收CO2,当Fc为正值时,说明生态系统在释放CO2,也就是说在全年各个月份当中,缙云山针阔叶混交林生态系统在白天都为碳汇,在夜间则为碳源。主要原因是受到辐射的影响,从而使生态系统在光合与呼吸作用之间相互切换。其中碳汇的波动范围为:−0.97~−0.01 mg·m−2·s−1,最小值出现在8月的12:00,最大值出现在5月的19:00。碳源多集中在夜间,其波动区间是:0.04~0.32 mg·m−2·s−1,最大值出现在12月的0:00点,最小值出现在1月份的6:30。
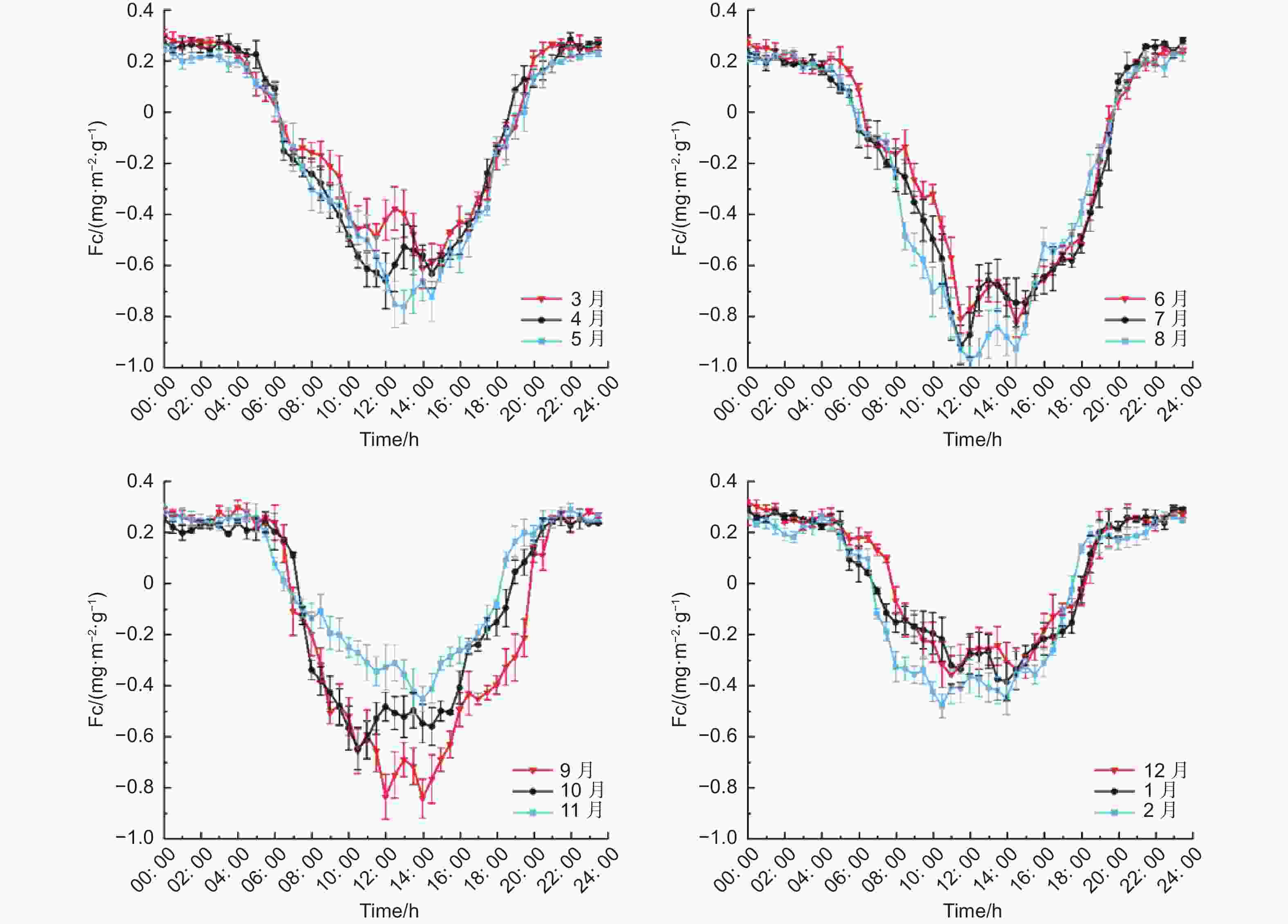
图 7 各月份碳通量日变化
Figure 7. Daily variation of carbon flux in different months
研究期间各月份碳汇峰值分布范围为11:30~15:00,6月最早达到碳汇峰值,1月最晚达到峰值;碳源峰值主要分布范围为 20:00—次日 3:00。除了11月,其他各月均出现两个碳汇峰值,即在中午段Fc出现增大的现象。碳汇能力最大的是8月份,日均Fc为−0.24 mg·m−2·s−1,最小的是12月份,FC为0.03 mg·m−2·s−1。每个月份碳汇碳源的转化时刻也不尽相同,最早由碳源转为碳汇的是7月和8月均为6:00,最晚为12月时间是7:30;最早由碳汇转变为碳源的是2月(18:00),最晚为5、6、7、8和9月(19:30)。其中,日固碳时间最长的月份为7月,日固碳时间为13.5 h,最短为12月,日固碳9.5 h。
-
按照月份的不同,划分为四季,分别是春季(3—5月)、夏季(6—8月)、秋季(9—11月)、冬季(12—次年2月),分别求同一时刻下的Fc的平均值。得到碳通量各季节的日变化(图8)。各季节的Fc大致呈现U字形的日变化,在季节上也会出现午间的双峰变化。夏季峰值最早出现,发生时间为11:00,春季最不明显。不同季节的碳汇峰值排列如下:夏季(−0.83 mg·m−2·s−1) > 春季(−0.65 mg·m2·s−1) > 秋季(−0.62 mg·m−2·s−1) > 冬季(−0.38 mg·m−2·s−1);日碳汇时长夏季(13.5 h) > 春季(12 h) = 秋季(12 h) > 冬季(10.5 h),可见缙云山针阔叶混交林生态系统冬季的碳汇能力要小于其他季节。
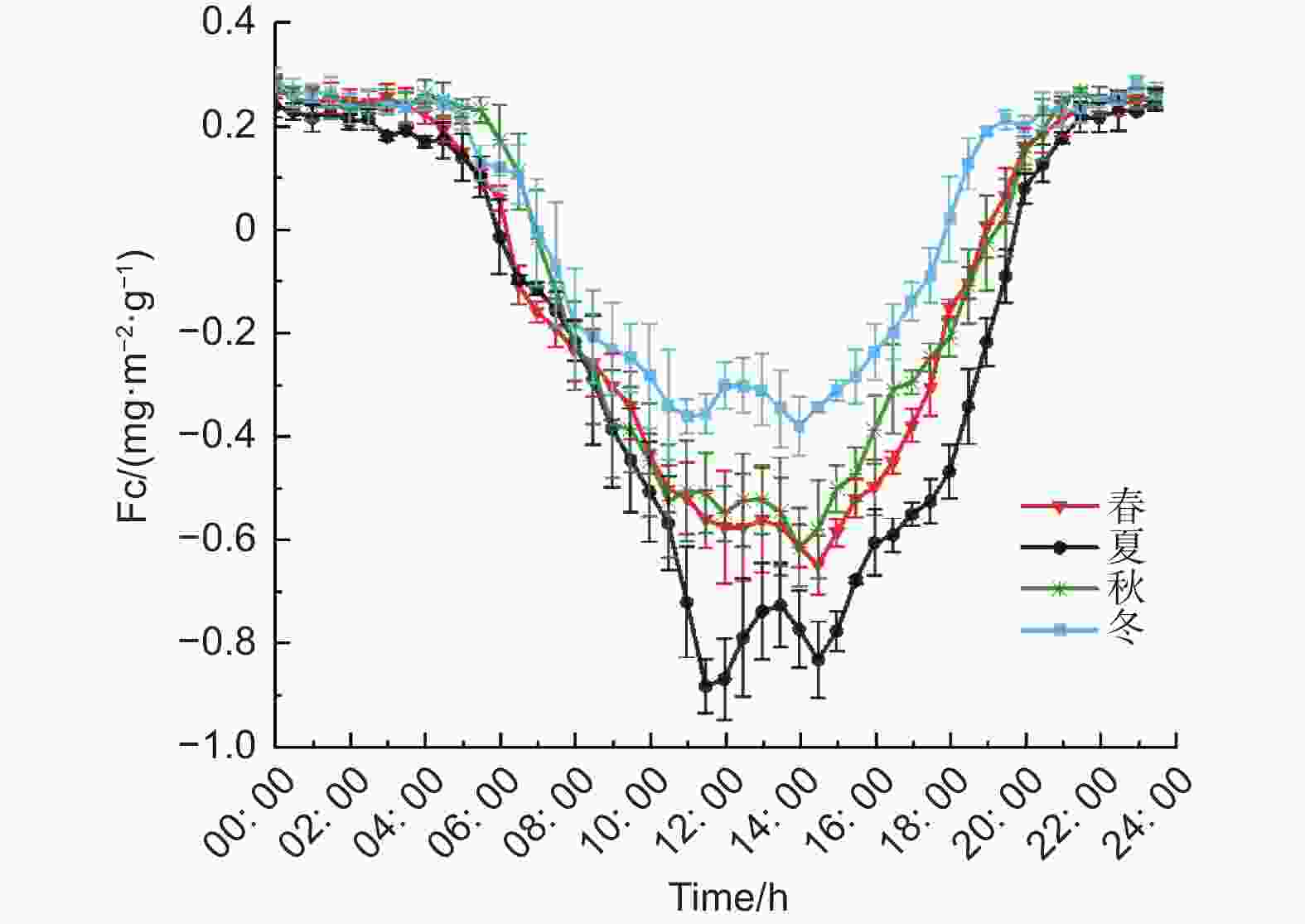
图 8 各季节碳通量的日变化
Figure 8. Seasonal variation of carbon flux
-
图9为净生态系统交换量(NEE)的全年月累积变化。全年的月平均NEE为−73.95 g·m−2,年累积NEE值为-887.40 g·m−2,可知缙云山针阔叶混交林生态系统为碳汇。不同月份NEE的累积值也不尽相同。最高值出现在12月份,为22.21 g·m−2·mon−1;最低值出现在8月份,为−172.27 g·m−2·mon−1。全年所有月份中,只有12月和1月为正值,其他季节均为负值,说明该生态系统冬季处于碳源过程,而其它季节处于碳汇过程。碳汇月份之间也存在差异,最大固碳月份为8月(−172.27 g·m−2·mon−1);最小固碳月份为11月(−3.71 g·m−2·mon−1)。碳汇月份从2月开始,碳汇能力逐渐增强,到8月份到达峰值;从9月开始逐渐减弱,一直到11月降为最低。从植物的生长季角度来看,生长季(4月—10月)与非生长季之间的固碳能力也存在着较大差异。生长季的Fc累积变化量为:−826.2 g·m−2,非生长季则只有:−61.2 g·m−2。这是因为在生长季节,植物需要消耗大量的养分,所以,需要进行强烈的光合作用来提供,从而需要吸收大量的CO2。
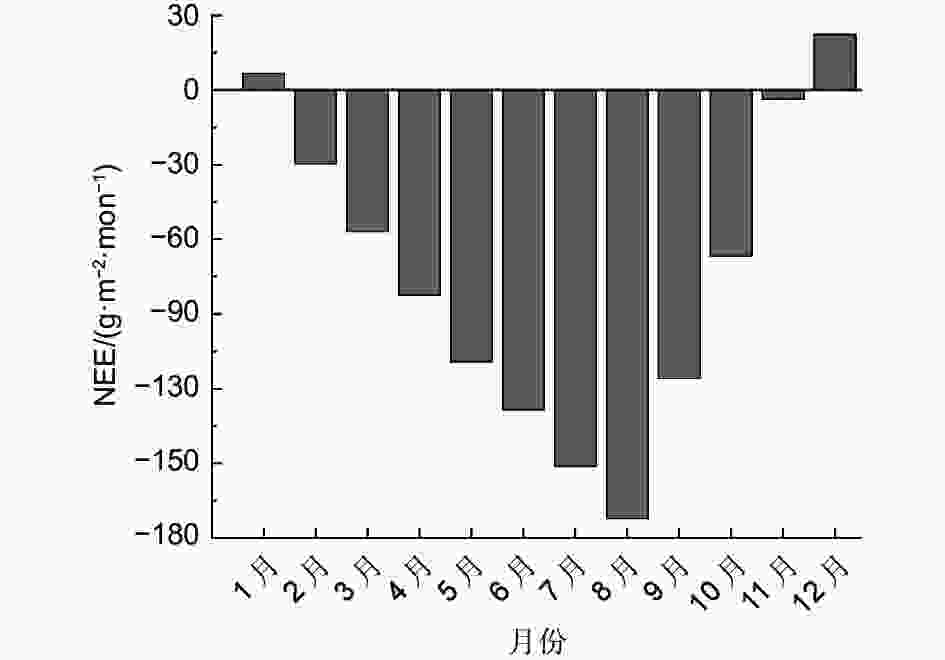
图 9 碳通量月累积变化
Figure 9. monthly cumulative variation of carbon flux
-
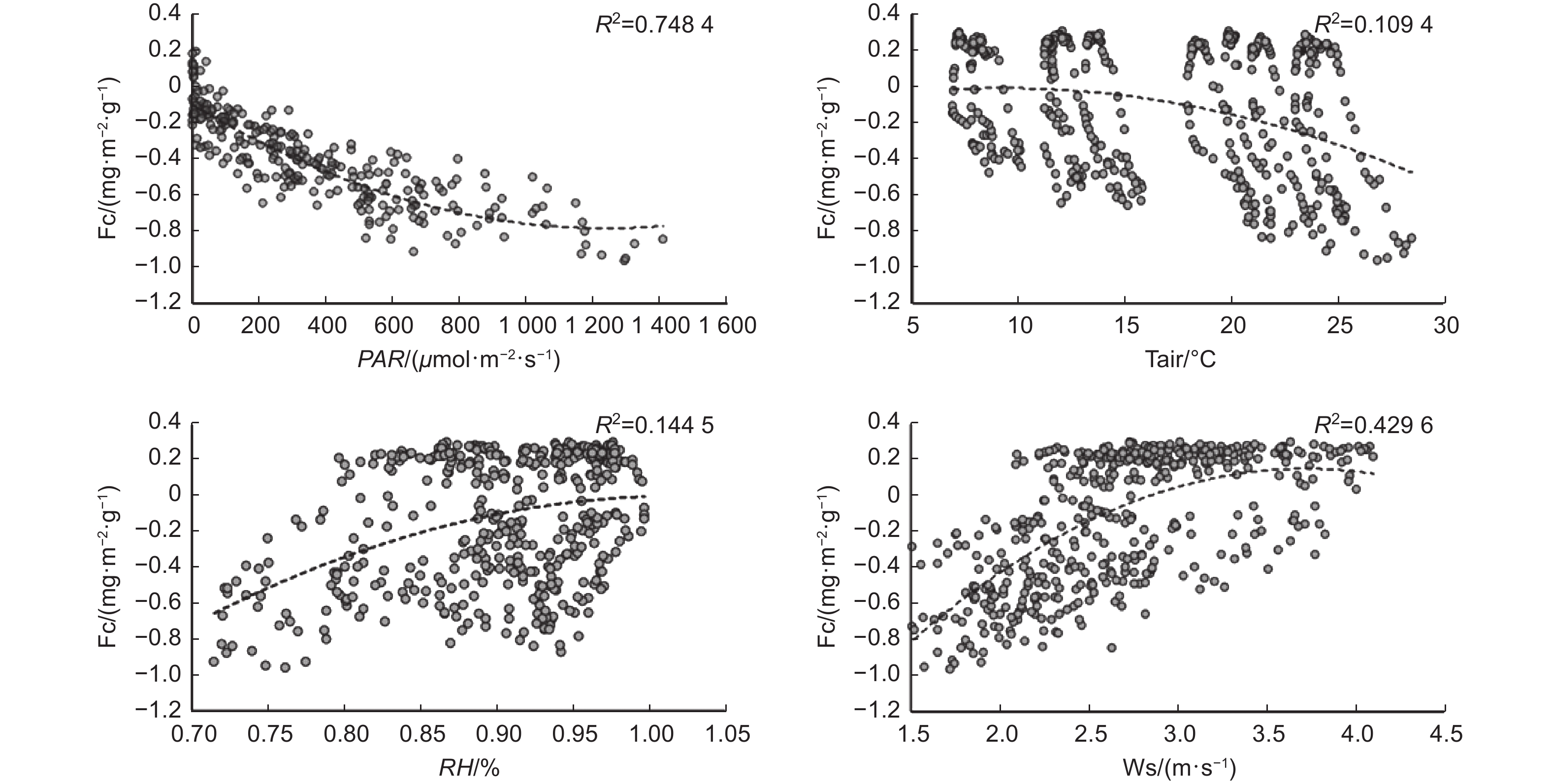
通过F检验以及碳通量与主要环境因子的拟合关系表明,光合有效辐射和风速是影响碳通量变化的主要因子,R2分别为0.75和0.43(图10)。光辐射作为光合作用的必要条件,其对植物光合至关重要。随着光辐射强度的增加,碳通量的值逐渐减小,植物吸收CO2的能力逐渐增强。风速作为影响涡度相关测量的主要气象因子,在本研究中,对碳通量的影响也很大,风速越小碳通量越平稳。空气相对湿度主要是通过影响植物的气孔导度从而间接影响碳通量的变化,空气湿度过高时植物的气孔会有不同程度的闭合。空气温度对碳通量变化的影响不显著。
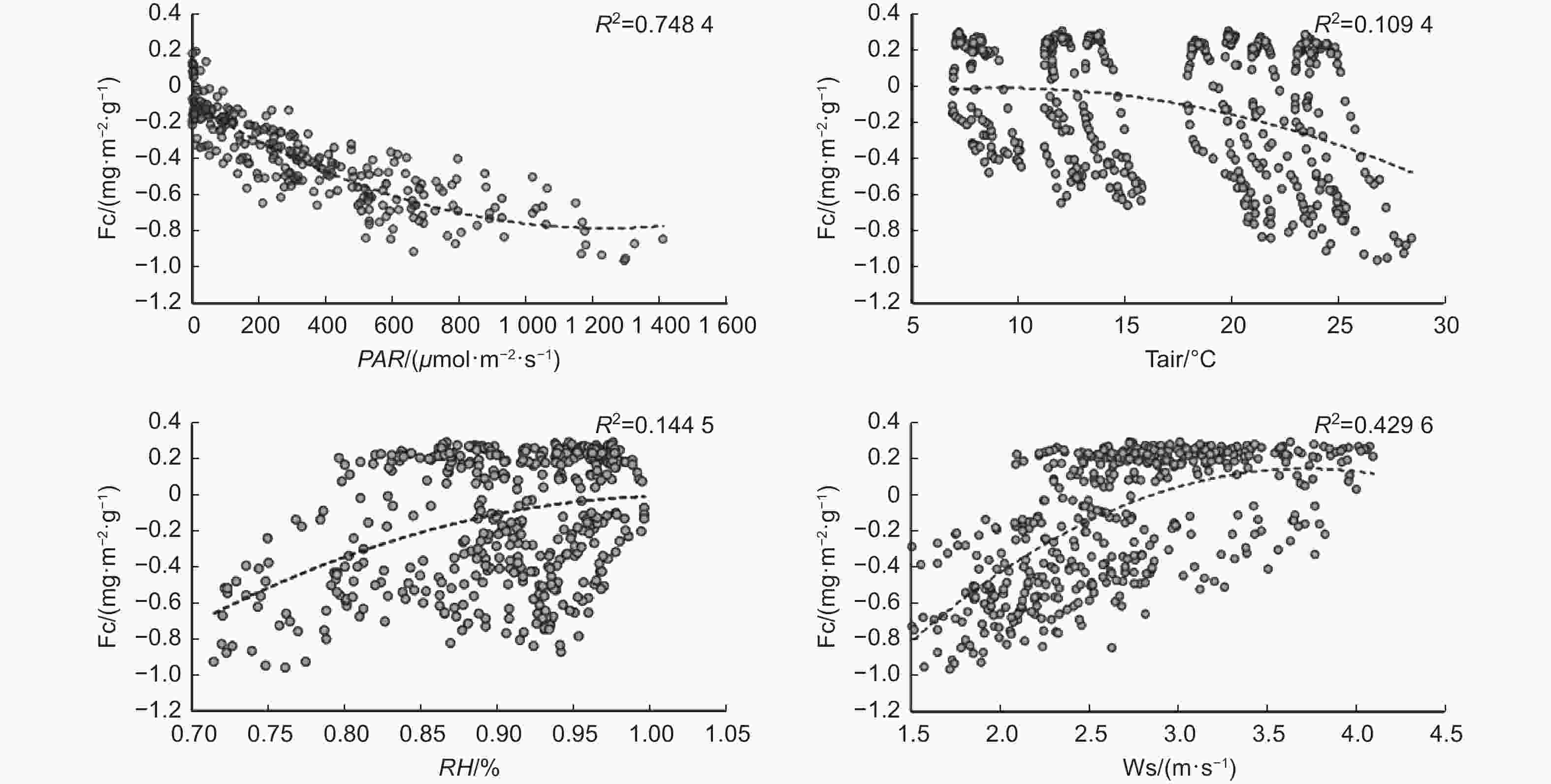
图 10 碳通量与主要环境因子的关系
Figure 10. The relationship between carbon flux and major environmental factors
-
评判涡度相关在一个地区的适应性,主要包括:湍流质量的评价、能量闭合分子以及通量足迹模拟。
本研究中H、LE、Fc的高质量湍流数据占比分别为77%、61%、62%。均超过60%。仅有少量的数据需要进行剔除插补。湍流的平稳状态,表明了针阔叶混交林生态系统近地面湍流发展平稳。王杰帅[14]在之前的研究中利用二次坐标旋转所得到的湍流质量评价分别为:77.5%、65.7%、68.5%,另外比较国内已有研究,吴志祥[17]研究了海南橡胶林生态系统的湍流数据质量评价分别为:63.4%、52.6%和59.1%与本研究结果相似。说明本研究中湍流趋于理想的稳态条件并且很好的符合湍流观测的发展特点。
关于能量闭合,张璇[18]、王杰帅[14]等均在缙云山针阔叶混交林生态系统进行过研究,能量闭合率分别为:0.84、0.78。在国内外相关的森林生态系统研究中,吴亚楠[19]关于小浪底山地人工生态系统的通量研究中,能量闭合率为0.77;李桐[20]在对九寨沟针阔叶混交林生态系统的通量研究中得出能量闭合率为0.71;Y. Kosugi等[21]在研究中,能量闭合率为0.74。均与本研究得到类似结果,说明缙云山针阔叶混交林生态系统能量闭合良好,涡度相关法在该研究区具有良好的适用性。
最后是通量足迹模拟,涡度相关法在测量时,需要生态系统下垫面具有足够的代表性[22],由于观测方法受到大气层和下垫面等环境因素的限制,通量足迹的函数模拟的贡献峰值区主要来自上风向[8, 23]。本研究所得到通量足迹模拟的峰值贡献区方向与研究期内的主风向一致,都是东北方向,说明通量数据信号来源正确。本研究中通量足迹的90%累计贡献区年平均距离为225.29 m,相比于其他平缓的林地[24]和农田[25-26]生态系统来说偏小,这是因为山地丘陵地当中的森林生态系统的下垫面通常是非均一的,且生态系统类型较为复杂[18],从而导致下垫面附近的大气在摩擦作用下产生环流,另外不同的观测高度也会使得通量贡献区距离不同,通常情况下观测高度越高,通量贡献区的距离就越大[27],这也是本研究中通量贡献区偏小的原因。
-
本研究期内碳通量的日变化中碳通量的波动范围是−0.97~0.38 mg·m−2·s−1,最小值出现在8月的12:00,最大值出现在5月的23:00,且波动范围在植物的生长季更为广泛,这是由于植物在生长季需要吸收大量的CO2来进行光合作用,从而合成有机物供应自身的生长,同时夜间呼吸也更为旺盛。2016年在对浙江天目山常绿阔叶落叶混交林生态系统的检测中,所得结果为:−1.00~0.30 mg·m−2·s−1[28];2019年对广东鼎湖山的针阔叶混交林的监测结果中碳通量的变化范围是−1.18~0.37 mg·m−2·s−1[29],与本研究结果相似。说明气候条件和生态系统类型相似的情况下,碳通量的变化也相近。而陈晓峰[30]在对安吉毛竹林的监测中,所得结果为−0.52~0.20 mg·m−2·s−1,碳汇能力弱于本研究结果。说明不同类型的森林生态系统,碳汇能力之间存在着较大差异。
本研究中碳通量成双峰变化,这主要是由于植物的“光合午休”造成的。因为中午光照强烈,致使气温升高、空气干燥,植物叶片为减少蒸腾,防止水分蒸发,被迫关闭气孔来减少水分散失,同时降低与光合作用有关的酶活性,这种现象被称为“光合午休”[31]。引起植物光合午休的主要原因是光辐射过高,过高的光辐射会引起叶绿体的回避反应,从而使植物的光合作用受到抑制[32],此外在高温的胁迫下会使植物的气孔关闭[33],从而导致其光合能力下降。徐勇峰[34]和牛晓栋[35]在对洪泽湖湿地生态系统和天目山常绿阔叶混交林的研究中也得到了相近的结果,说明植物在相应的环境胁迫下会出现光合午休的现象。光合午休的现象在夏季最为明显,这是因为夏季太阳高度角大,太阳辐射强度大,能够更快地使植物叶片因高温干旱而关闭气孔。春季之所以不明显,一方面是由于春季太阳辐射强度较小,另一方面是因为冬季的林地蒸散发作用不强烈,从而使土壤含水量充足以便于植物在春季吸收,用于蒸腾和生长。另外,由于本研究中所观测的生态系统为针阔叶混交林生态系统,植物种类并非单一,由于不同植物的光饱和点不同,其吸收CO2所达到峰值的时间也不近相同[36-37]。
-
(1)研究期内,湍流因子中的H、LE和Fc的湍流数据质量评价较好,高质量数据占比分别77%、61%和62%,趋于理想的稳态条件;在生长季内,上、下层能量闭合率良好分别为:0.82和0.73,均在国际山地森林生态系统能量闭合认可范围内(0.7~0.9)。
(2)研究期内,通量足迹模拟高贡献区域与研究期内主风向(东北)一致,表明通量监测的数据来源可信且质量良好。
(3)研究期内,该生态系统处于碳汇过程,年固碳量为887.40 g·m−2,月均日变化和各季节日变化大多为双峰变化。光合有效辐射和风速是影响碳通量变化的主要因子,R2分别为0.75和0.43。空气温度对碳通量变化的影响不显著。
缙云山针阔叶混交林涡相关适用性及碳通量变化特征
Eddy Covariance Applicability and Carbon Flux Variation Characteristics of Coniferous and Broad-leaved Mixed Forests in Jinyun Mountain
-
摘要:
目的 探讨涡度相关法在缙云山针阔叶混交林的适用性,并明晰针阔叶混交林生态系统的碳源/碳汇情况,从而为该地区的森林经营和林业可持续发展提供科学依据。 方法 利用涡度协方差观测方法,获得了缙云山针阔叶混交林2019.11—2020.10的碳通量监测数据。通过湍流数据质量评价、能量闭合及通量足迹,分析对涡度相关应用于该下垫面的适用性进行验证。在时间序列上对碳通量的变化进行分析,并对研究区内碳源或碳汇情况进行评估。 结果 研究期内,显热通量(H)、潜热通量(LE)和碳通量(Fc)的湍流数据质量评价较好,高质量数据占比分别为77%、61%和62%;在生长季(4—10月),上层能量闭合率为0.82,下层能量闭合率为0.73;研究期内通量高贡献区域所处方向与全年主风向(东北)一致;在研究期内,该生态系统年固碳量为887.40 g·m−2,月均日变化和季节均日变化大多为双峰型,生长季的Fc累积变化量为:−826.2 g·m−2,非生长季则只有:−61.2 g·m−2;影响碳通量的主要环境因子是光合有效辐射和风速,R2分别是0.75和0.43。 结论 涡度相关法在缙云山针阔叶混交林生态系统的监测过程中具有较好的适用性,且通量监测的数据来源可信,质量评价良好。研究期内,该生态系统处于碳汇过程,生长季的碳汇能力显著高于非生长季。光合有效辐射和风速是影响碳通量变化的主要环境因子。 Abstract:Objective To provide helpful insights into forest management and sustainable forestry development, the applicability of eddy covariance method in the coniferous and broad-leaved mixed forest in Jinyun Mountain was explored and the carbon source or sink of the coniferous and broad-leaved mixed forest ecosystem was clarified. Method Used the eddy covariance observation method(EC) to obtain the carbon flux monitoring data of the coniferous and broad-leaved mixed forest in Jinyun Mountain from 2019.11 to 2020.10. Through turbulence data quality evaluation, energy closure and flux footprint analysis, the applicability of the eddy covariance to the underlying surface was verified. The changes of carbon flux in the ecosystem were analyzed, and the carbon source or sink in the study area were evaluated. Results During the study period, the turbulence data quality check of sensible heat flux(H), latent heat flux(LE) and carbon flux(Fc) was good, and the proportions of high-quality data were 77%, 61% and 62% respectively. During the growing season(April-October), the upper-layer energy closure rate was 0.82, and the lower-layer energy closure rate was 0.73. The direction of the high flux contribution area was consistent with the main wind direction (northeast)The. During the study period, the annual carbon sequestration of the ecosystem was 887.40 g·m−2. The monthly average daily changes and seasonal average daily changes were mostly bimodal, and the cumulative change of Fc was −826.2 g·m−2 in the growing season and −61.2 g·m−2 in the non-growing season. The main environmental factors affecting carbon flux were photosunthetically active radiation and wind speed, and R2 was 0.75 and 0.43, respectively. Conclusion The EC had good applicability in the monitoring process of the Jinyun Mountain coniferous and broad-leaved mixed forest ecosystem, and flux monitoring data are creadible, and flux quality check is evaluated well. During the study period, the ecosystem is in the process of carbon sink,and the carbon sink capacity in the growing season is significantly higher than that in the non-growing season. Photosynthetically active radiation and wind speed are the main environmental factors affecting carbon flux changes. -
图 5 全年通量足迹(左为白天,右为夜间)
Figure 5. Annual flux footprint (day on the left and night on the right)
图 10 碳通量与主要环境因子的关系
Figure 10. The relationship between carbon flux and major environmental factors
表 1 通量塔仪器布设
Table 1. Instrument layout of flux tower
观测系统 仪器名称 型号 生产商 安装高度/m 频率/Hz 开路式涡度相关监测系统 开路式气体分析仪 Li-7500 LiCor Inc.,USA 35 10 三维超声风速仪 CAST-3 Campbell,USA 35 10 数据采集器 CR3000 35 10 微气象观测系统 空气温湿度传感器 HMP60 Vaisala, FIN 25
3510 翻斗式雨量筒 TR-525M Texas Electronics, USA 25
3510 光合有效辐射传感器 LI-190SB LiCor Inc.,USA 25
3510
净辐射仪NR01 Hukseflux,NED
3510 风速风向仪 03002 RM Young,CN 35 10  下载: 导出CSV
下载: 导出CSV
表 2 通量贡献区距离
Table 2. Distance of flux contribution area
月份 x_peak/m x_10%/m x_30%/m x_50%/m x_70%/m x_90%/m 4 83.43 28.64 71.32 108.77 152.55 228.55 5 89.26 30.64 76.30 116.36 163.2 244.49 6 80.69 27.70 68.97 105.19 147.53 221.02 7 84.39 28.97 72.14 110.01 154.29 231.16 8 88.32 30.32 75.50 115.55 162.4 244.16 9 91.12 31.28 77.89 118.78 166.6 249.59 10 84.90 29.14 72.57 110.68 155.23 232.56
下载: 导出CSV
-
[1] Christopher B. Field. Plant physiology of the "missing" carbon sink[J]. Plant Physiology, 2001, 125(1): 25-28. doi: 10.1104/pp.125.1.25 [2] Rodney J, Keenan, Gregory A, et al. Dynamics of global forest area: results from the FAO global forest resources assessment[J]. Forest Ecology and Management, 2015, 352(1): 9-20. [3] 康满春. 北方典型杨树人工林能量分配与碳水通量模拟[D]. 北京:北京林业大学, 2016. [4] Xiao Jun-lan, Zeng Feng, He Qiu-lan, et al. Responses of forest carbon cycle to drought and elevated CO2[J]. Atmosphere, 2021, 12(2): 212. doi: 10.3390/atmos12020212 [5] 龚 元, 纪小芳, 花雨婷, 等. 基于涡动相关技术的森林生态系统二氧化碳通量研究进展[J]. 浙江农林大学学报, 2020, 37(3):593-604. doi: 10.11833/j.issn.2095-0756.20190412 [6] Wilson K,Goldstein A,Falge E, et al. Energy balance closure at FLUXNET sites[J]. Agricultural and Forest Meteorology, 2002, 113(1): 223-243. [7] 于贵瑞, 张雷明, 孙晓敏, 等. 亚洲区域陆地生态系统碳通量观测研究进展[J]. 中国科学:D辑 地球科学, 2004, 34(S2):15-29. [8] Kim J H, Hwang T,Schaaf C L,et al. Seasonal variation of source contributions to eddy-covariance CO2 measurements in a mixed hardwood-conifer forest[J]. Agricultural and Forest Meteorology, 2018, 253(1): 71-83. [9] Rodda S R, Thumaty K C, Jha C S, et al. Seasonal variations of carbon dioxide, water vapor and energy fluxes in tropical indian mangroves” [J]. Forests ,2016,7 (35):1–18. [10] 王 倩, 王云琦, 马 超, 等. 缙云山针阔混交林碳通量变化特征及影响因子研究[J]. 长江流域资源与环境, 2019, 28(3):565-576. [11] Nave L E, Swanston C W, Mishra U, et al. Afforestation effects on soil carbon storage in the United States: a synthesis[J]. Soil Science Society of America Journal, 2013, 77(3): 1035-1047. doi: 10.2136/sssaj2012.0236 [12] Makkonen M,Hhttunen S,Primmer E, et al. Policy coherence in climate change mitigation: An ecosystem service approach to forests as carbon sinks and bioenergy sources[J]. Forest Policy and Economics, 2015, 50(1): 153-162. [13] Niu S, Fu Z, Luo Y, et al. Interannual variability of ecosystem carbon exchange: From observation to prediction[J]. Global Ecology and Biogeography, 2017, 26(11): 1225-1237. doi: 10.1111/geb.12633 [14] 王杰帅. 基于涡度法的西南丘陵区森林碳通量观测研究[D]. 北京:北京林业大学, 2020. [15] Foken T, Göckede M, Mauder M, et al. Handbook of micrometeorology: post-field data quality control[M]. Dordrecht: Springer Netherlands: 2004.181-208. [16] Wilczak J M, Oncley S P, Stage S A. Sonic anemometer tilt correction algorithms[J]. Boundary-Layer Meteorology, 2001, 99(1): 126-150. [17] 吴志祥. 海南岛橡胶林生态系统碳平衡研究[D]. 海口:海南大学, 2013. [18] 张璇. 重庆缙云山针阔混交林水分传输特征及对环境因子的响应机制[D]. 北京: 北京林业大学,2019. [19] 吴雅楠. 黄河小浪底山地人工混交林生态系统水碳交换及环境响应[D]. 北京:北京林业大学, 2018. [20] 李 桐, 鄢春华, 王 蓓, 等. 九寨沟针阔混交林能量平衡特征[J]. 生态学报, 2018, 38(22):8098-8106. [21] Y. Kosugi, M. Katsuyama. Evapotranspiration over a Japanese cypress forest. II. Comparison of the eddy covariance and water budget methods[J]. Journal of Hydrology, 2006, 334(3): 56. [22] Aubinet M. Estimates of the annual net carbon and water exchange of forests: The EUROFLUX methodology[J]. Advances in Ecological Research, 2000, 30: 114-175. [23] Kljun N, Calanca P, Rotach M W, et al. A simple two-dimensional parameterisation for Flux Footprint Prediction (FFP)[J]. Geoscientific Model Development, 2015, 8(11): 3695-3713. doi: 10.5194/gmd-8-3695-2015 [24] 刘 伟, 韦志刚, 郑志远, 等. 珠海凤凰山林地下垫面观测通量的贡献区分析[J]. 热带地理, 2016, 36(5):840-849. [25] 朱明佳, 赵谦益, 刘绍民, 等. 农田下垫面观测通量的变化特征及其气候学足迹分析[J]. 地球科学进展, 2013, 28(12):1313-1325. [26] 吴东星, 李国栋, 张 茜. 华北平原冬小麦农田生态系统通量贡献区[J]. 应用生态学报, 2017, 28(11):3663-3674. [27] 郭智娟, 龚 元, 张凯迪, 等. 复杂下垫面下不透水层的CO2通量足迹分析——以上海市奉贤大学城为例[J]. 环境科学学报, 2018, 38(2):772-779. [28] 方成圆. 天目山常绿落叶阔叶混交林能量和CO2通量的研究[D]. 杭州: 浙江农林大学, 2016. [29] 黄 俊, 麦博儒, 邓雪娇, 等. 鼎湖山针阔叶混交林CO2浓度及通量变化特征[J]. 广西植物, 2019, 39(2):261-274. [30] 陈晓峰. 浙江安吉毛竹林生态系统碳通量及响应机制研究[D]. 杭州: 浙江农林大学, 2016. [31] 王 倩. 缙云山针阔混交林碳通量变化特征及影响因子研究[D]. 北京: 北京林业大学, 2018. [32] Maai E, Nishimura K, Takisawa R, et al. Light stress-induced chloroplast movement and midday depression of photosynthesis in sorghum leaves[J]. Plant Production Science, 2019, 23(2): 1-10. [33] 高冠龙, 冯 起, 刘贤德. 自然条件下胡杨叶片光合作用的气孔、非气孔限制[J]. 干旱区资源与环境, 2020, 34(11):182-188. [34] 徐勇峰, 季 淮, 韩建刚, 等. 洪泽湖湿地杨树林生长季碳通量变化特征及其影响因子[J]. 生态学杂志, 2018, 37(2):322-331. [35] 牛晓栋, 江 洪, 张金梦, 等. 浙江天目山老龄森林生态系统CO2通量特征[J]. 应用生态学报, 2016, 27(1):1-8. [36] 吴 勇, 张海燕, 卢腾飞, 等. 基于Landsat 8 OLI的滇西北高山亚高山针叶林地上生物量遥感估测模型构建及光饱和点确定[J]. 云南大学学报:自然科学版, 2021, 43(4):818-830. [37] 陈晓萍, 吕 敏, 王满堂, 等. 江西阳际峰73种阔叶植物叶片光截获和水分排除策略对树冠位置的响应研究[J]. 中国科学:生命科学, 2021, 51(1):91-101. -

 点击查看大图
点击查看大图

图(10) / 表(2)
计量
- 文章访问数: 3574
- HTML全文浏览量: 2016
- PDF下载量: 60
- 被引次数: 0



