



-
作为一种固着生物,植物的生长发育时刻受到各种外界环境条件的影响。面对各种逆境胁迫,植物已经进化出包括形态、细胞、分子及生理生化水平在内的复杂调控机制予以应答[1-3]。作为一种调节蛋白,转录因子(Transcription factors,TFs)通过调控目的基因的表达在植物生长发育及逆境胁迫响应中发挥着重要作用[4-5]。
HD-Zip (homeodomain leucine zipper)蛋白是植物特有的转录因子,根据结构与功能差异可以进一步将其分为4个亚家族(HD-Zip I~IV)[6]。植物HD-Zip I蛋白仅包含C-端的同源异型结构域(Homeodomain, HD)和紧密相连的亮氨酸拉链结构域(leucine zipper,LZ),它们广泛参与植物的生长发育以及非生物胁迫应答等过程[7]。研究人员已经对拟南芥(Arabidopsis thaliana L.)、水稻(Oryza sativa L.)、番茄(Solanum lycopersicum Mill.)、向日葵(Helianthus annuus L.)等植物中的HD-Zip I蛋白进行了家族成员鉴定及相关基因的功能研究[8-11]。拟南芥ATHB12和ATHB7都受到ABA和干旱胁迫的诱导表达,且ATHB12的异源表达可增强转基因酵母对NaCl的耐受能力[12-13]。水稻Oshox22基因影响内源ABA的合成,并通过依赖ABA的信号转导途径调控水稻的干旱和盐胁迫耐受能力[14]。向日葵Hahb-4基因受干旱和外源ABA的强烈诱导表达,过表达该基因的拟南芥植株对缺水胁迫表现出更强的耐受能力[4]。向日葵HaHB1基因的异源表达可提高拟南芥对低温的耐受能力,并上调转基因株系内抗冻相关基因的表达[15]。番茄SlHB2基因的下调表达则可诱导胁迫相关基因的表达并提高植株对干旱和盐胁迫的耐受能力[7]。玉米HD-Zip I亚家族成员Zmhdz10的表达受外源ABA和盐胁迫的强烈诱导,该基因的过表达可明显提高水稻和拟南芥对干旱和盐胁迫耐受能力[16]。综上可见, 植物HD-ZIP I亚家族蛋白在激素响应及应对非生物胁迫过程中发挥着重要的调控作用。
作为一种重要的经济作物,桑树(Morus notabilis L.)在我国已有几千年的栽种历史。桑叶可养蚕,桑树皮可作造纸原料,桑果既可鲜食也可以加工成果酒、果汁和果酱,桑树的根、茎、叶和桑果还具备一定的药用价值。此外,桑树对干旱、盐碱及水淹等逆境胁迫具有较强的耐受能力,是木本植物抗逆境研究的优良材料。本研究从基因组水平对桑树HD-ZIP I亚家族基因进行了鉴定和分析,对家族蛋白的理化性质、进化关系及基因结构等进行了分析。利用 RNA-seq数据研究了HD-ZIP I亚家族基因的组织表达模式,通过qPCR检测了该家族基因在激素及非生物胁迫处理后的表达情况。为进一步阐明桑树HD-ZIP I亚家族基因的生物学功能和抗逆基因的挖掘奠定了基础。
-
从拟南芥基因组数据库(
https://www.arabidopsis.org/tools/index.jsp )获取拟南芥HD-Zip I蛋白序列[17],并以此作为查询序列在NCBI数据库(https://www.ncbi.nlm.nih.gov/ )及桑树基因组数据库(https://morus.swu.edu.cn/morusdb/ )进行Blast同源搜索,下载桑树HD-Zip I亚家族的基因及氨基酸序列。所有序列均通过InterPro(http://www.ebi.ac.uk/interpro/ )、NCBI CDD (https://www.ncbi.nlm. nih.gov/Structure/cdd/wrpsb.cgi )及SMART (http://smart.embl-heidelberg.de )数据库进行保守结构域分析。利用MEME Version 5.4.1(http://meme-suite.org/tools/meme )对蛋白保守基序进行预测,基序数量设为10,其余参数采用默认值。桑树HD-Zip I蛋白的多序列分析由ClustalW及DNAMAN 9软件完成,系统进化分析采用邻接法(Neighbor-joining method, NJ)由MEGA 7.0完成,bootstrap值设置为1000 [18]。蛋白分子量、等电点等均由在线软件(https://web.expasy.org/protparam/ )计算获得[19]。蛋白亚细胞定位预测分析在WoLFPsort(https://wolfpsort.hgc.jp/ )网站中进行。基因的内含子与外显子分布由Tbtool (windows-x64_1_068)软件绘制,并利用Tbtool软件从桑树基因组数据中获取HD-Zip I基因起始密码子(ATG)上游2 000 bp的启动子序列,在plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )数据库对启动子区域的顺式作用元件进行预测分析,并用Tbtool软件进行可视化[20]。 -
利用桑树基因组数据库中根、枝条、冬芽、雄花和叶的RNA-seq数据,分析桑树HD-Zip I基因的组织表达模式。相关基因的RPKM值经log2转换后,通过Tbtool软件绘制热图。
-
桑树品种育711扦插苗于25 ℃温室条件下(光照16 h,黑暗8 h)生长45 d后,选取长势一致的单株进行处理。将100 µmol·L−1 脱落酸( ABA)溶液均匀喷施于幼苗叶面进行激素处理,用200 mmol·L−1的NaCl对树苗进行盐处理,拔出幼苗并洗掉根上的泥土置于室温进行脱水处理,淹水至树苗地上部分5 cm处作淹水处理,以正常浇水树苗为对照(CK)。对以上各处理的材料于处理后0、3、6、12、24 h摘取相同发育时期的叶片,利用液氮冷冻后置于–80 ℃冰箱备用,每个时间点设置3次重复,每次重复取3株树苗叶片。
-
RNAiso plus(TaKaRa)用于材料总RNA提取,M-MLV反转录酶(Promega)试剂盒用于合成cDNA。利用Primer Premier 5软件设计特异性定量分析引物,以ACTIN基因作为内标基因(表1)。定量PCR体系(10 μL)中包含qPCR Mix(2 × GoTaq®, 5.0 μL)、引物混合物(10 μmol·L−1, 0.5 μL)、模板cDNA(1.0 μL)、Rnase-free H2O(3.5 μL)。在eppendorf荧光定量仪上进行定量分析(95 ℃预变性2 min,95 ℃变性15 s,60 ℃退火延伸30 s,40次循环),3次技术重复,采用2−∆∆CT公式计算基因相对表达量,使用Origin 9.0绘制柱形图。
表 1 桑树HD-Zip I基因qRT-PCR分析引物
Table 1. Primers used in qRT-PCR analysis of HD-Zip I genes in mulberry
基因名称
Gene Name引物序列(5′–3′)
Primer sequence产物大小/bp
Product lengthMnHD-Zip 1 F:TTGACGGAGAAACTTCAGGCTAA R:TGAAAGTAGGAGTCACCGCTGTC 209 MnHD-Zip 2 F:TCAGAGGGTGAGGTCTCCAAAG R:GATTGGTCGGCTTCAAACACA 161 MnHD-Zip 3 F:CAGGCAGGTTGCTATTTGGTTT R:ATTTTCTCGCCCTCCTTCGT 208 MnHD-Zip 4 F:TGAAAAATAGAGAGCCAACGGAGT R:TGGAAAGAGAGGTCTGCTTGTGAT 176 MnHD-Zip 5 F:GTGTTTGAGCCAGAGCAGTCG R:TCGCACACGGGTCATCATAAC 127 MnHD-Zip 6 F:GCCCTAATGTCCGTCTGCC R:CCGCTTCCTCCACACATCC 121 MnHD-Zip 7 F:GGCTTGAAGTTGAAGGATACGG R:GACATCCCATTTAGCCCAAGTG 142 MnHD-Zip 8 F:TGGCTTCCCAGTTCGAGTCA R:CCAATGTCGTTTTCCAATCCC 166 MnHD-Zip 9 F:AGAAATGCCGTCTTGAATCCG R:ATCCTTCAGACACCACCCCAA 183 MnHD-Zip 10 F:CCTCGTCAGATTGCTGTTTGG R:GTAACCGCCCGTTGACACTT 198 MnHD-Zip 11 F:GCTAACAAAGGACCAAGTCAGGC R:GGCATTTTCAAGCCTCACCTG 205 MnHD-Zip 12 F:TAGGAAAAGACGCTGAAGTTGTAGA R:TCAAGACAAGAACTCCCACCACT 140 MnHD-Zip 13 F:TGAACAAGGCTCAACAGATGC R:ATCATCCACGGCTGACACTG 101 MnHD-Zip 14 F:ATCCTTCCCAACCACCTTCC R:TTCCAGCGAGCCCTCTTGT 136 ACTIN F:CCAAGTCATCACAATCGGAGC R:TACCAGGGAACATAGTTGAACCAC 191 -
经保守结构域分析并排除冗余序列后,共获得14个桑树HD-Zip I成员,分别命名为MnHD-Zip 1~MnHD-Zip 14(表2)。从表2可以看出:桑树HD-Zip I蛋白的氨基酸数及其分子量变化较大,分别介于173~351 aa和19 927.40~39 913.22;等电点pI则介于4.54~8.88,弱酸性蛋白偏多。平均亲水性值则位于−1.040~−0.657之间。蛋白亚细胞定位结果表明,桑树HD-Zip I蛋白主要定位于细胞核。HD-Zip I亚家族蛋白的不同特点及成员间的差异可能预示着该亚家族转录因子在桑树生长发育过程中发挥着不同的调控作用。
表 2 桑树HD-Zip I家族基因及其编码蛋白信息
Table 2. The detailed information of HD-Zip I members of mulberry
基因名称
Gene Name登录号
Gene ID蛋白长度/aa
Protein length分子量/kDa
Molecular weight等电点
PI亲水性值
Hydropathicity亚细胞定位
Subcellular localizationMnHD-Zip 1 L484_008420 281 31634.93 4.70 −0.817 nucl:14 MnHD-Zip 2 L484_010105 325 36528.92 4.68 −0.839 nucl:14 MnHD -Zip 3 L484_027159 326 36766.68 6.54 −0.982 nucl:13.5, cyto_nucl: 7.5 MnH D-Zip 4 L484_021122 328 37377.30 6.42 −1.020 nucl: 13.5, cyto_nucl: 7.5 MnHD-Zip 5 L484_021079 292 33207.53 4.54 −0.922 nucl: 13, pero: 1 MnHD-Zip 6 L484_019806 342 39104.85 5.03 −0.997 nucl: 13.5, cyto_nucl: 7.5 MnHD-Zip 7 L484_012750 351 39913.22 4.94 −0.945 nucl: 13, pero: 1 MnHD-Zip 8 L484_001654 219 25604.55 6.66 −1.029 nucl: 14 MnHD-Zip 9 L484_009043 209 24036.19 6.93 −0.820 nucl: 14 MnHD-Zip 10 L484_002276 205 23523.59 8.53 −0.888 nucl: 14 MnHD-Zip 11 L484_012158 197 23221.98 6.66 −0.990 nucl: 14 MnHD-Zip 12 L484_015344 192 21891.73 8.88 −0.718 nucl: 14 MnHD-Zip 13 L484_010529 173 19927.40 7.89 −0.657 nucl: 14 MnHD-Zip 14 L484_023454 182 20630.97 6.78 −1.040 nucl: 14 注:nucl:细胞核;cyto:细胞质;pero:过氧物酶体。
Notes: nucl: nucleus; cyto: cytosol; pero: peroxisome. -
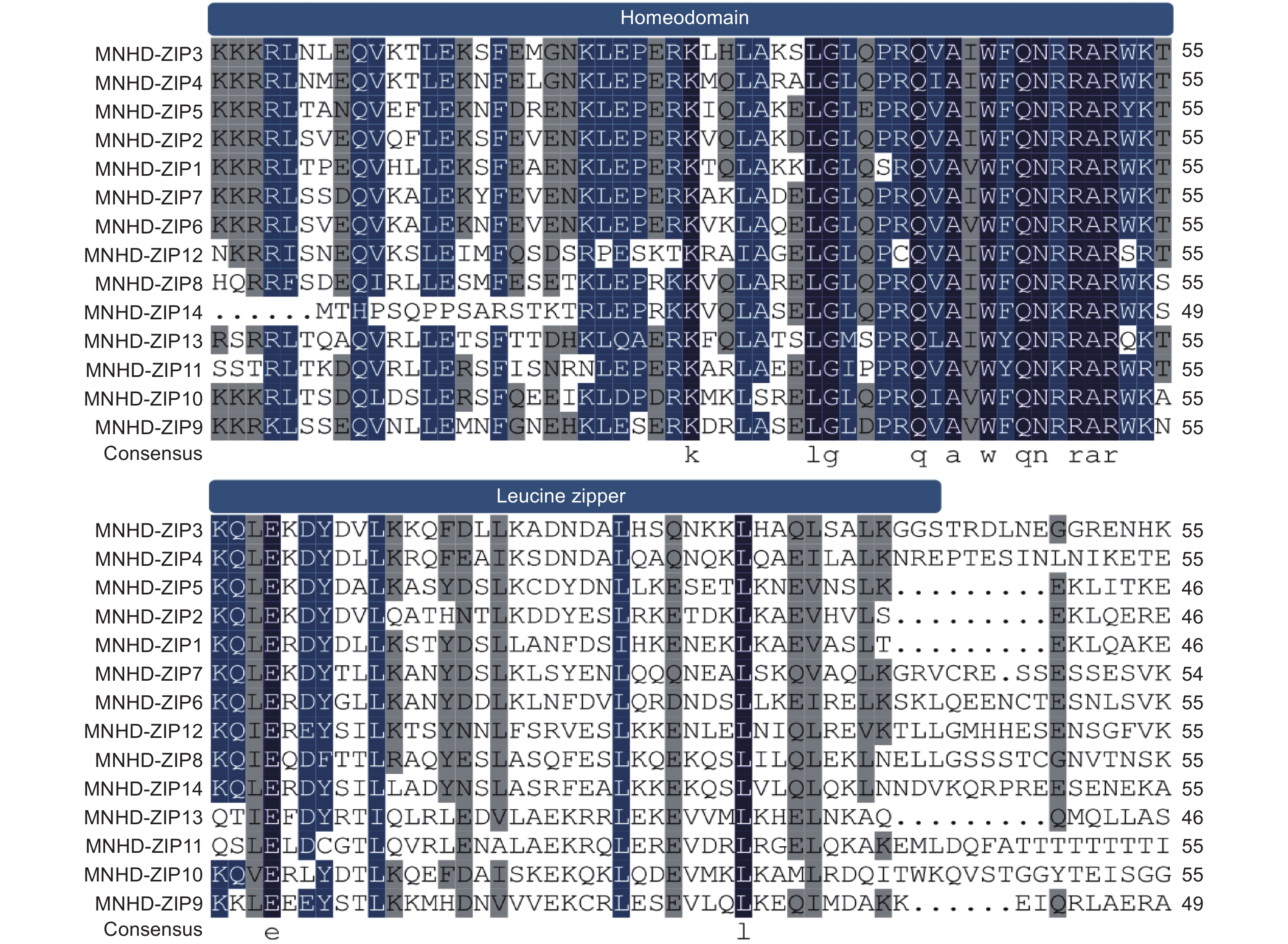
蛋白多序列比对结果发现,桑树HD-Zip I亚家族成员均含有HD-Zip家族转录因子的保守结构域HD(Homeomain)和LZ(leucine zipper)。其中,HD结构域的序列高度保守,LZ结构域的氨基酸残基则相对多变(图1)。

图 1 桑树 HD-Zip I 蛋白 Homeodomain 及 leucine zipper 结构域多序列比对
Figure 1. Multiple sequence alignments of Homeodomain and leucine zipper domains
为分析桑树HD-Zip I亚家族成员的进化关系,利用MEGA 7.0软件构建了拟南芥和桑树该家族蛋白的进化树。参考拟南芥该家族成员的进化关系,将桑树HD-Zip I亚家族成员分为α、β、γ、δ、ε、φ 6个分枝,其中,MnHD-Zip 3和MnHD-Zip 4属于α分枝,MnHD-Zip 5、MnHD-Zip 2、MnHD-Zip 1、MnHD-Zip 7和MnHD-Zip 6属于β分枝,MnHD-Zip 8、MnHD-Zip 12和MnHD-Zip 14属于γ分枝,MnHD-Zip 11和MnHD-Zip 13属于φ分枝,MnHD-Zip 10和MnHD-Zip 9分别属于ε和δ分枝(图2)。

图 2 桑树与拟南芥 HD-Zip I 家族的系统进化分析
Figure 2. Phylogenetic analysis of HD-Zip I family in mulberry and Arabidopsis
-
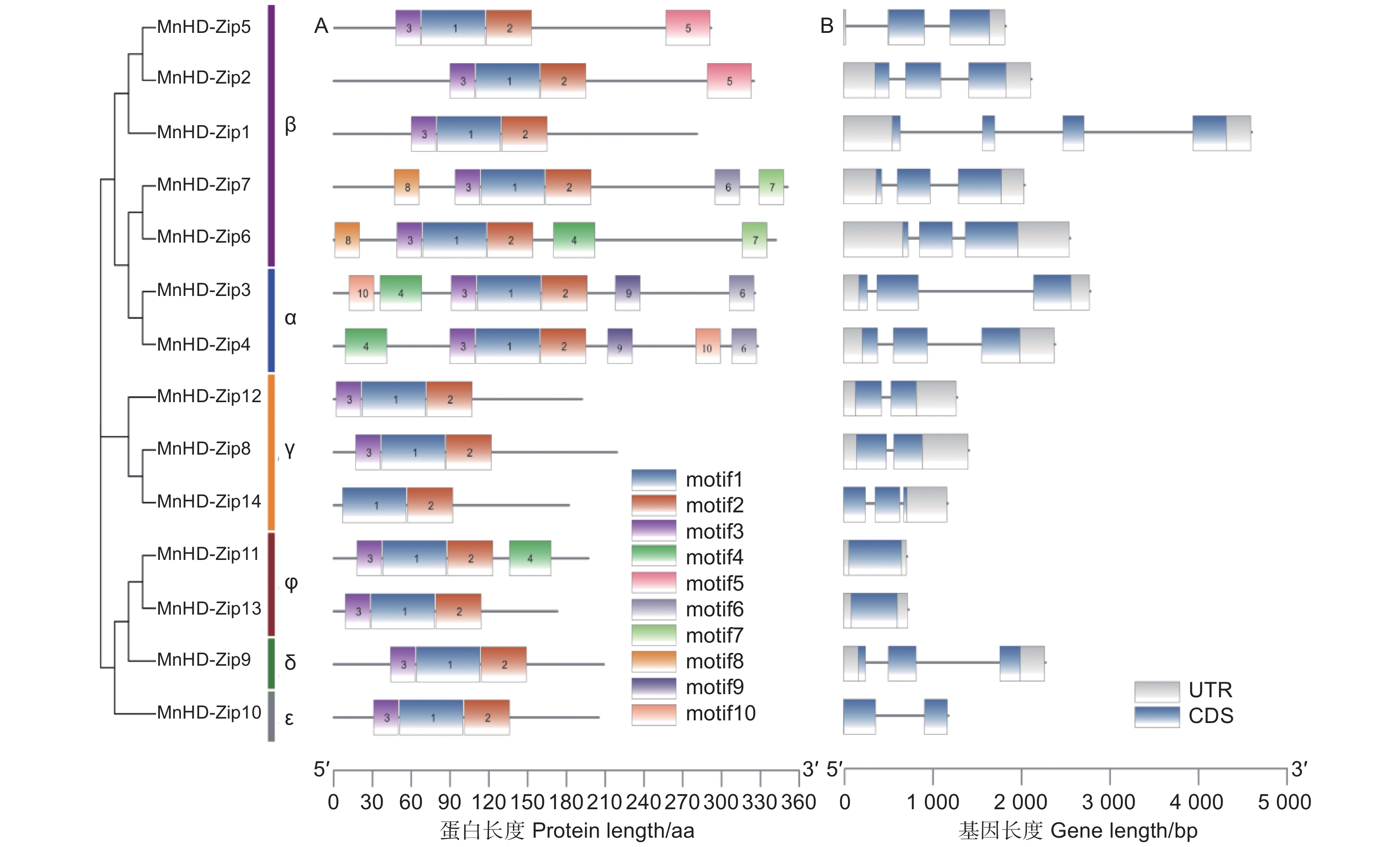
蛋白motif分析结果表明:桑树HD-Zip I亚家族蛋白检测出的10个基序中,motif 1和motif 2为该家族蛋白共有基序,进一步序列分析发现,motif 1主要参与构成桑树MnHD-Zip I蛋白的Homeomain结构域,motif 2则是leucine zipper结构域的主要组成部分。桑树HD-Zip I家族内各成员所含有的motif种类及数量存在较大差异,如MnHD-Zip 3和MnHD-Zip 4共检测出7个基序,MnHD-Zip 14仅发现2个基序;同一进化分枝内各转录因子所含有的基序种类及数量则表现出较高的相似性,如α分枝的2个成员均含有7个相同的motif,φ分枝成员均含有motif 1、motif 2和motif 3(图3A)。

图 3 桑树 HD-Zip I 家族蛋白保守基序(A)及基因结构(B)分析
Figure 3. Analysis of conserved motifs(A)and exon–intron organization(B)of MnHD-Zip I genes
通过比较MnHD-Zip I基因的mRNA和基因组序列,获得该家族基因的外显子/内含子结构(图3B)。从图3B可以看出:桑树HD-Zip I家族成员中,同一进化分枝成员具有相似的外显子/内含子数量及分布规律,其中,β分枝的MnHD-Zip 1基因独具4个外显子,该分枝的其余成员则均只含有3个外显子;α和δ分枝的所有成员都含有3个外显子;γ分枝成员中,MnHD-Zip 14基因有3个外显子,其余两个成员同ε分枝成员都只含有2个外显子;φ分枝成员的外显子数量最少,均只含有1个外显子。
-
为了进一步明确桑树HD-Zip I家族基因启动子区域的相关顺式作用元件,探索该家族基因对激素及逆境的响应机制,利用PlantCARE对该家族基因的启动子进行了分析,结果(表3)表明:桑树14个HD-Zip I基因的启动子区检测出参与激素及非生物胁迫响应的顺式作用元件共计11种。其中,激素响应元件6种,即ABRE(ABA响应元件)、P-box(赤霉素响应元件)、TCA-element(水杨酸响应元件)、CGTCA-motif(茉莉酸响应元件)、TGACG-motif(茉莉酸响应元件)和AuxRR-core(生长素响应元件);非生物胁迫响应元件5种,即TC-rich repeats(防御和胁迫响应元件)、LTR(低温胁迫响应元件)、ARE(厌氧诱导元件)、MBS(干旱诱导的MYB结合位点)及WUN-motif(损伤反应元件)。
表 3 桑树HD-Zip I家族基因启动子区顺式作用元件分析
Table 3. Analysis of cis acting elements in HD-Zip I gene’s promoter region
分枝
Clades基因名
Gene name激素响应元件
Phytohormone response element非生物胁迫响应元件
Abiotic stress response elementABRE P-box TCA-element CGTCA-motif TGACG-motif AuxRR-core TC-rich repeats LTR ARE MBS WUN-motif β MnHD-Zip 5 2 0 0 0 1 0 2 0 4 1 1 MnHD-Zip 2 1 0 2 1 1 1 0 1 2 0 0 MnHD-Zip 1 3 0 1 4 4 0 0 0 1 2 0 MnHD-Zip 7 1 0 1 0 0 0 3 0 5 1 0 MnHD-Zip 6 0 0 0 2 2 0 0 0 4 0 1 α MnHD-Zip 3 1 1 1 3 3 0 0 0 2 0 0 MnHD-Zip 4 3 0 0 1 1 0 0 1 1 2 0 γ MnHD-Zip 12 8 0 0 1 1 0 0 0 2 0 1 MnHD-Zip 8 2 1 3 0 0 0 0 1 2 0 2 MnHD-Zip 14 1 0 0 3 3 0 0 2 0 0 2 φ MnHD-Zip 11 6 0 0 0 0 0 0 0 5 0 1 MnHD-Zip 13 2 1 4 2 2 0 1 2 1 0 0 δ MnHD-Zip 9 4 2 2 4 4 0 0 0 2 0 0 ε MnHD-Zip 10 2 0 2 2 2 0 0 0 0 1 0 桑树HD-Zip I家族基因启动子区顺式作用元件的数量和种类存在明显差异,但不同分枝成员的元件分布则具有一定的相似性,如β分枝成员启动子均不含有P-box元件,γ和φ分枝成员的启动子区都没有MBS元件。从数量上看,MnHD-Zip 9基因启动子区拥有的元件数最多(18个),MnHD-Zip 2、MnHD-Zip 4、MnHD-Zip 6和MnHD-Zip 10的启动子所含元件最少(9个)。从顺式作用元件的种类看,ABRE元件数量最多,除MnHD-Zip 6外的13个MnHD-Zip I基因启动子区共含有36个ABRE;ARE元件次之,在12个MnHD-Zip I基因启动子区中共有31个;AuxRR-core元件最少,仅在MnHD-Zip 2基因启动子区中出现1次。以上结果表明,MnHD-Zip I基因启动子区的顺式作用元件在数量和种类方面存在明显差异,预示该家族基因可能在激素及非生物胁迫响应中具有不同的生物学功能。
-
为了解桑树HD-Zip I家族基因的组织表达模式,对桑树基因组数据库中根、枝条、冬芽、雄花和叶片的转录组数据进行分析,结果(图4)表明:从组织特异性方面看,大部分桑树HD-Zip I家族基因在雄花和枝条中的表达量相对较高,根和叶片中大部分基因的表达量则相对较低;从进化分枝方面看,同一分枝基因具有相似的组织特异性表达。β分枝的5个成员中,MnHD-Zip 2和MnHD-Zip 6在根、枝条、冬芽、雄花和叶片中均呈现高水平表达,MnHD-Zip 5在除叶片外的器官/组织中也有较高水平的表达,预示该分枝内的基因在桑树的不同组织/器官生长发育调控中发挥着重要作用。α分枝中2个成员的组织/器官特异性表达呈现高度的一致性,均在雄花和枝条中高水平表达;γ分枝中,MnHD-Zip 8和MnHD-Zip 14在雄花和枝条中的转录水平也都较高,说明α和γ分枝的4个基因在桑树雄花发育及枝条生长方面具有潜在调控功能。此外,φ分枝的MnHD-Zip 13基因分别在雄花、根、枝条和δ分枝的MnHD-Zip 9基因分别在雄花、冬芽中也有较高水平的表达,暗示它们也在桑树相应组织/器官的生长发育中发挥着重要调控作用。
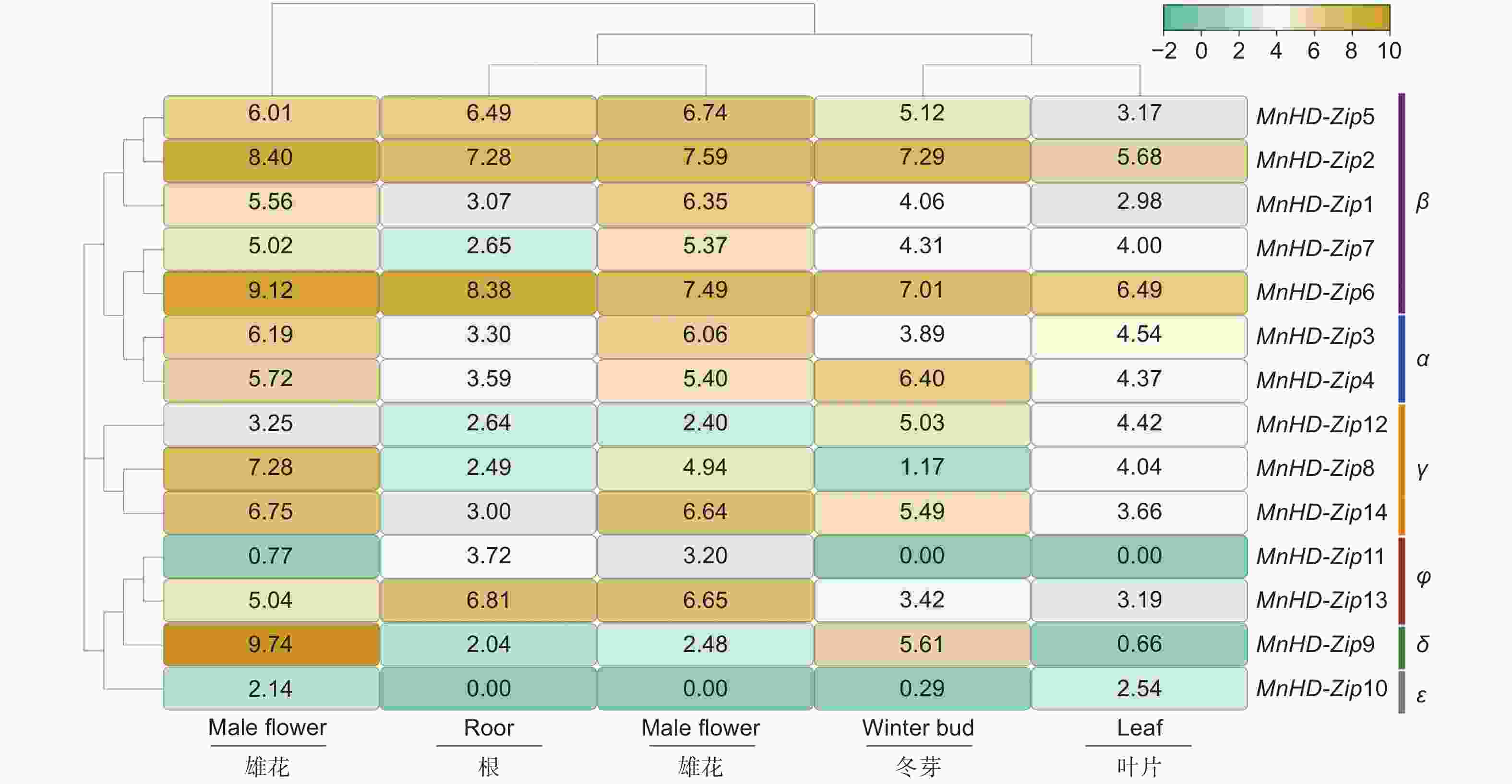
图 4 桑树 HD-Zip I 家族基因的器官特异性表达
Figure 4. Organ specific expression analysis of MnHD-Zip I genes
-
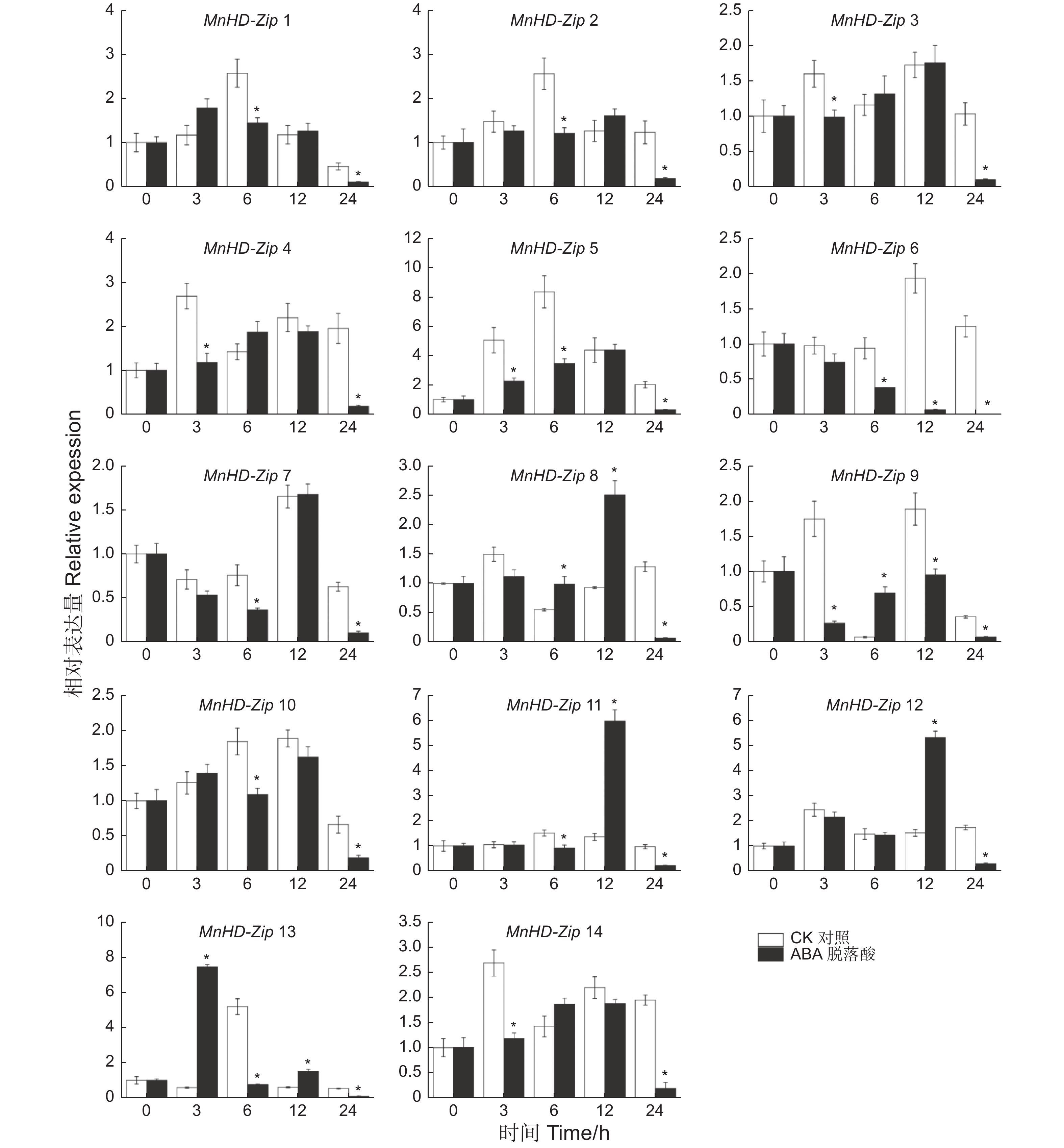
为了分析桑树HD-Zip I家族基因对激素的响应情况,对桑树苗进行了ABA处理,以正常浇水桑树苗为对照,利用qRT-PCR技术分别检测了处理0、3、6、12、24 h后叶片中MnHD-Zip I基因的转录水平,结果(图5)表明:多数桑树HD-Zip I基因并未受到ABA的上调诱导,甚至很多基因的转录水平被ABA所抑制。β分枝中,MnHD-Zip 1、MnHD-Zip 2与MnHD-Zip 5的表达模式相似,MnHD-Zip 6和MnHD-Zip 7则呈现另一种相似的表达趋势;但ABA处理后,β分枝中所有基因的表达水平都受到不同程度的抑制。α分枝的MnHD-Zip 3和MnHD-Zip 4基因也具有相似的表达模式,且其转录都在3、24 h时间点被ABA显著抑制。γ分枝中,MnHD-Zip 8和MnHD-Zip 12的转录水平在处理12 h后均达到对照的2倍以上,但到24 h后,它们的表达水平又显著低于对照;MnHD-Zip 14基因的表达则在3、24 h时间点受到ABA的显著抑制。φ分枝中,MnHD-Zip 11基因的表达水平在12 h被显著上调,随后快速下降并显著低于对照;MnHD-Zip 13则在处理3 h时便受到ABA的快速上调诱导,随后开始降低直至显著低于对照。MnHD-Zip 9基因仅在处理 6 h时被显著上调诱导,3、12、24 h则均被显著抑制。MnHD-Zip 10基因的表达在6、24 h也受到ABA的抑制。
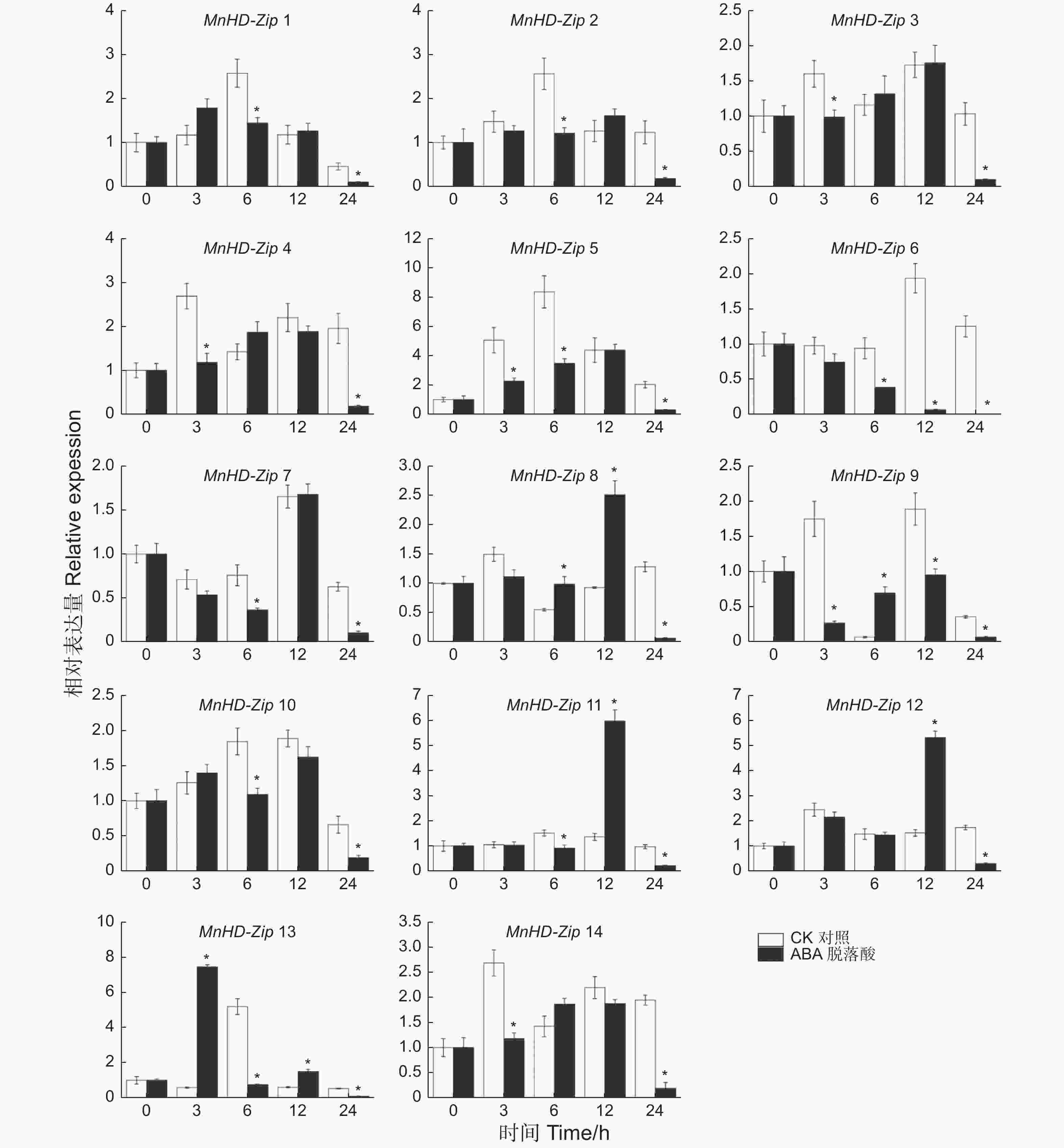
图 5 桑树 HD-Zip I 家族基因激素处理后的差异表达分析
Figure 5. Temporal differential expression of MnHD-Zip I genes under ABA
-
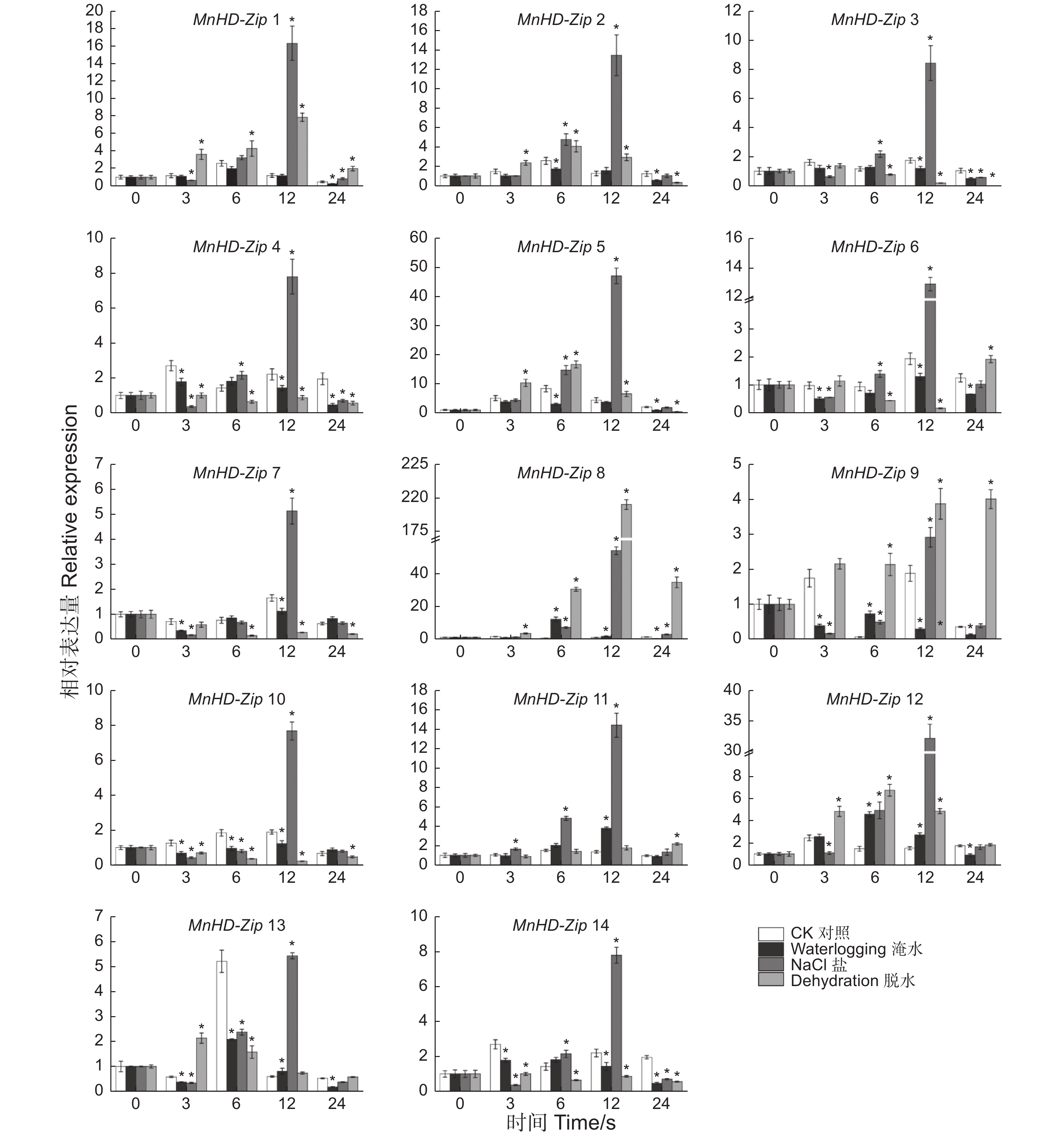
为分析非生物胁迫对桑树HD-Zip I家族基因表达的影响,对桑树苗进行了淹水、盐及脱水处理,以正常浇水桑树苗为对照,利用qRT-PCR技术分别检测了处理0、3、6、12、24 h后叶片中MnHD-Zip I基因的转录水平,结果(图6)表明:桑树HD-Zip I亚家族基因均受到3种非生物胁迫不同程度的诱导表达,且多数基因的表达水平在处理后12 h上升至峰值。β分枝的5个成员中,MnHD-Zip 1基因的转录水平在脱水胁迫处理后3~24 h的几个时间点均被显著诱导上调;在盐处理后12 h,该基因的表达被诱导上调约14倍;MnHD-Zip 5和MnHD-Zip 2基因转录水平的变化趋势与MnHD-Zip 1较相似,且这2个基因在盐处理后12 h的表达水平均有10倍以上的上调;MnHD-Zip 6和MnHD-Zip 7表达水平的变化呈另一种相似的趋势,且都在盐处理后12 h出现2倍以上的上调表达。α分枝的MnHD-Zip 3和MnHD-Zip 4基因的表达水平都只在盐处理后12 h有2倍以上的上调;处理后24 h,2个基因的表达水平又同时下调至CK以下。γ分枝中,MnHD-Zip 8基因在盐和脱水胁迫处理后6~24 h,其表达水平都出现2倍以上的上调诱导,在淹水胁迫3 h时也显著上调诱导;MnHD-Zip 12基因在淹水、盐和脱水处理后6、12 h也被显著诱导上调表达;MnHD-Zip 14在盐处理后12 h显著上调,在处理后3、24 h该基因的表达同时受到3种非生物胁迫不同程度的下调诱导。φ分枝中,MnHD-Zip 11在盐胁迫下全程上调诱导表达,淹水处理后12 h该基因的表达量也显著高于对照;MnHD-Zip 13基因在盐处理后12 h被显著诱导上调。MnHD-Zip 9基因在脱水胁迫处理后3~24 h表达水平均显著诱导上调。MnHD-Zip 10仅在盐处理后12 h受到显著的上调诱导,在非生物胁迫处理后3、6 h,其表达量显著低于CK。
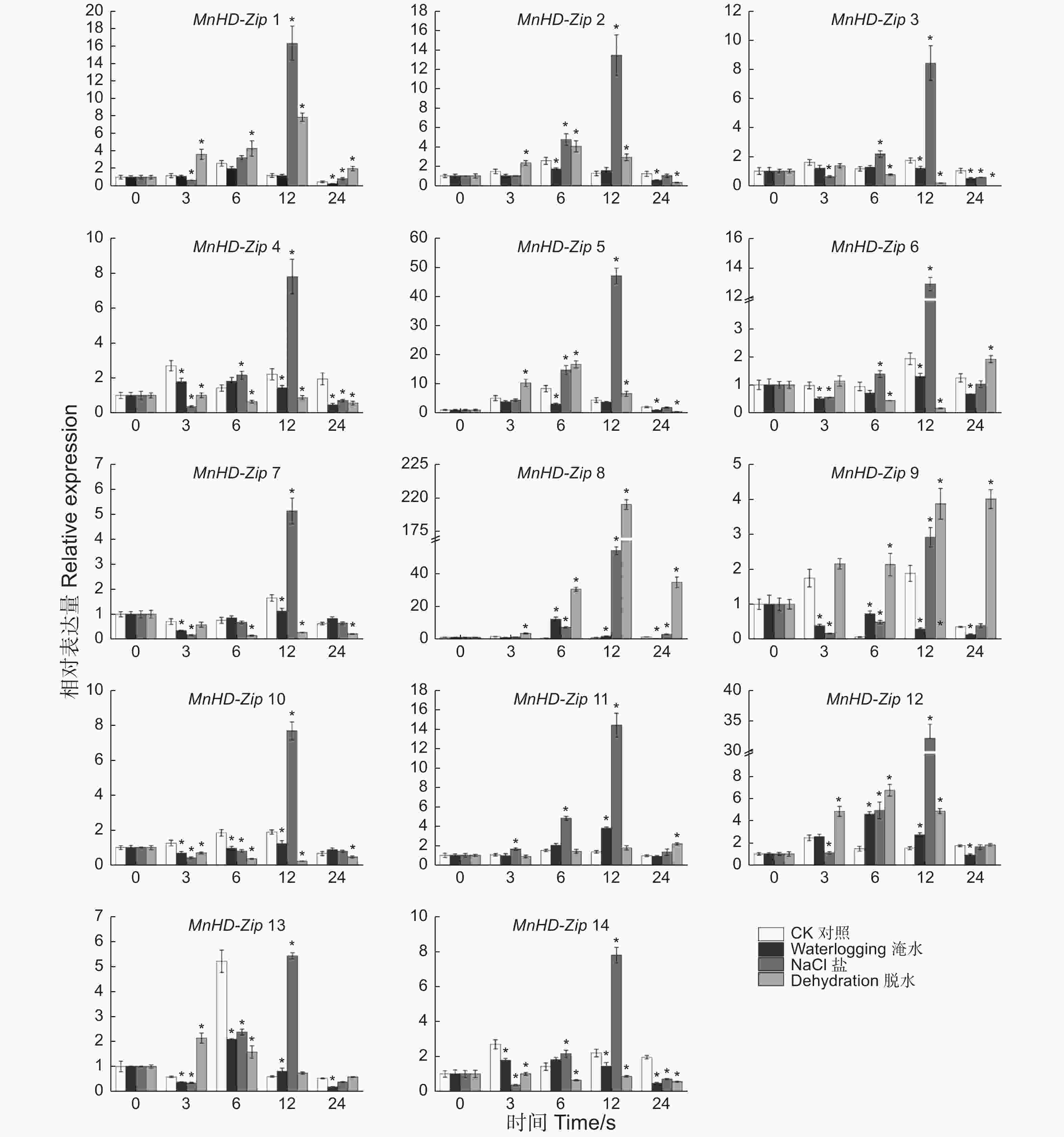
图 6 桑树 HD-Zip I 家族基因胁迫处理后的差异表达分析
Figure 6. Temporal differential expression of MnHD-Zip I genes under abioticstresses
-
自桑树基因组测序工作完成以来[21- 22],PHR、bHLH、Trihelix、bZIP、MYB、ERF等家族的转录因子已进行过相关的分析与研究[23-25],但桑树HD-Zip I亚家族成员的系统鉴定及其对淹水等非生物胁迫及激素响应的研究还未见报道。本研究通过系统进化分析、基因结构分析、顺式作用元件预测、基因的组织/器官特异性表达分析、ABA和淹水等非生物胁迫诱导后的转录水平检测等,对桑树HD-Zip I亚家族基因进行了鉴定和研究。通过全基因组分析,本研究共鉴定到14个桑树HD-Zip I亚家族基因,与拟南芥、水稻和玉米(Zea mays L.)中同一亚家族基因的数量相似,但明显少于苹果(Malus pumila Mill.)和杨树(Populus L.)[8-9, 26-28]。
根据氨基酸序列的相似性,桑树HD-Zip I亚家族蛋白可进一步被分为6个分枝,这一研究结果同拟南芥、水稻、玉米的研究结果相似[8-9, 26]。从成员分布看,桑树HD-Zip I亚家族中,β分枝有5个成员,γ分枝有3个成员,α和φ分枝各有2个成员,δ和ε分枝各1有个成员。可见,桑树该亚家族成员相对集中在β分枝,而δ和ε分枝成员数偏少,这与拟南芥中各分枝成员相对均衡的分布规律不尽一致[8]。
桑树HD-Zip I亚家族蛋白含有多个基序,其中,家族成员共有基序motif 1和motif 2分别参与构成该家族蛋白的Homeomain结构域和leucine zipper结构域,说明motif 1、motif 2为桑树HD-Zip I亚家族蛋白的特征性基序。此外,同一分枝的成员在所含有的基序种类及数量方面具有高度的相似性。家族基因结构分析也得出类似的结论,即桑树HD-Zip I亚家族中属于同一分枝的基因具有高度相似的外显子/内含子数量及分布规律,这一分析结果进一步印证了桑树HD-Zip I亚家族蛋白进化树分枝关系的可靠性。
基因启动子区域中的不同顺式作用元件通常暗示该基因具备对相应激素或者胁迫诱导的响应能力[29]。研究发现,桑树HD-Zip I亚家族基因的启动子区存在多种参与激素及非生物胁迫响应的顺式作用元件,如ABRE(ABA响应元件)、MBS(干旱诱导的MYB结合位点)及WUN-motif(损伤反应元件)等,这些元件的大量存在表明桑树HD-Zip I基因在激素及非生物胁迫响应及相关的生长调控方面发挥着潜在的重要作用。后续的基因表达分析发现,桑树HD-Zip I亚家族基因的表达都受到激素和非生物胁迫不同程度的诱导,这一结果很好地证实了上述顺式作用元件分析所得出的结论。
研究表明,HD-Zip I亚家族基因的表达具有组织和器官特异性,通常受到包括干旱、极端温度及渗透胁迫等外界环境因素的影响[6]。作为一类植物转录因子,HD-Zip I蛋白广泛参与了植物的生长发育以及非生物胁迫应答等生命过程,包括花序发育及开花时间调节、侧根发育调控、抗逆境胁迫调节等[15-16, 30-32]。本研究对14个桑树HD-Zip I亚家族基因在根、枝条、冬芽、雄花和叶片中的转录水平进行了分析,结果发现,MnHD-Zip I基因在所检测的5个组织中均有不同程度的表达。从组织特异性上看,该家族基因主要在雄花及枝条中有相对较高的表达量,大部分基因在根和叶片中的表达量则相对偏低,这与柑橘(Citrus sinensis cv. Valencia)HD-Zip I基因主要在叶片和花中高表达的结果不太一致[33]。但桑树HD-Zip I基因是否具有与水稻HD-Zip I家族基因相同的花发育调控功能还需后续实验进一步验证[34]。从进化树分枝看,处于同一进化分枝内的基因具有相似的组织/器官表达特异性,其中,β分枝的成员在多数组织/器官中都呈现高水平表达,暗示该分枝成员在桑树生长发育中具有潜在重要功能;同时,这一结果与黄瓜(Cucumis sativus L.)的研究结果较吻合[35]。
激素及非生物胁迫处理后的表达模式分析结果表明,ABA处理对除MnHD-Zip 8、MnHD-Zip 9、MnHD-Zip 11-13外的桑树HD-Zip I亚家族基因的表达没有显著上调诱导作用,不少成员甚至出现下调诱导,如β分枝及α分枝的所有基因均在处理后24 h受到明显下调诱导,这一结果与拟南芥、黄瓜、水稻、向日葵和玉米中该家族基因的研究结果不同[4, 8, 16, 35-38]。研究发现,与MnHD-Zip 8同属γ分枝的荔枝LcHB2基因受到乙烯处理的上调诱导,且该基因在果实离区高水平表达。LcHB2蛋白可与LcCEL2 和 LcCEL8基因启动子区的顺式元件结合并激活这2个基因的表达,从而促进果实脱落[39]。棉花γ分枝蛋白GhHB12则可通过抑制下游GhFT、GhFUL和GhSOC1基因的表达起到调控棉花开花时间的作用[40]。MnHD-Zip 8基因也受到ABA的诱导表达,且其在桑树花器官中也呈现高水平表达,推测MnHD-Zip 8基因可能也在桑树花器官发育中发挥相应的重要功能。植物HD-Zip I亚家族β分枝的基因中,拟南芥ATHB1和ATHB16分别在叶片细胞的发育及株高、花序发育方面具有重要调控功能[41-42],番茄LeHB-1基因则参与调控花发育及果实成熟调控[10],说明β分枝成员在植物中发挥着多方面的发育调控功能。本研究结果表明,桑树中β分枝成员受到盐和脱水胁迫的显著诱导表达,且在各种器官中也有较广泛的高表达,因此,推测这些基因在桑树生长发育及逆境胁迫中均有潜在的调控作用。γ分枝中,Oshox22基因的沉默转基因株系具有较高的干旱和盐胁迫耐受能力[14],向日葵Hahb-4基因过表达株系也对缺水胁迫表现出更强的耐受能力[4, 11, 43],说明该分支成员在植物非生物胁迫调控中发挥着重要作用。本研究中,桑树γ分枝基因MnHD-Zip 8和MnHD-Zip 12受到盐、淹水及脱水胁迫的显著诱导,由此推测,它们在桑树应对上述逆境胁迫中具有潜在重要功能。总体看,桑树HD-Zip I亚家族基因受到淹水、盐和脱水处理的诱导表达,其中,盐和淹水处理的诱导作用最明显。盐和脱水处理后,大部分MnHD-Zip I基因的表达水平随时间的推移呈逐步升高后再降低的趋势,且多数基因在处理后12 h时达到峰值,出现这一现象的原因可能与基因表达的节律性存在一定联系[40]。
-
本研究从桑树基因组数据库中鉴定出14个HD-Zip I亚家族基因,这些基因可进一步被划分为6个进化分枝。基因结构及蛋白保守基序分析结果表明,同一进化分枝内的桑树HD-Zip I成员具有结构方面的相似性。RNA-seq数据分析发现,MnHD-Zip 2和MnHD-Zip 6基因在根、枝条、冬芽、雄花和叶片中均呈现高水平表达,MnHD-Zip5、MnHD-Zip1和MnHD-Zip 7也在多数器官中具有较高的表达水平。激素处理后的基因表达分析发现,除MnHD-Zip 8、MnHD-Zip 9、MnHD-Zip 11、MnHD-Zip 12和MnHD-Zip 13外,其余MnHD-Zip I基因的表达水平均受到ABA不同程度的抑制。非生物胁迫处理后的基因表达分析发现,桑树HD-Zip I亚家族基因均受到3种非生物胁迫不同程度的诱导表达,其中,γ分枝基因MnHD-Zip 8受到盐和脱水胁迫诱导后大幅度上调表达,MnHD-Zip 12也受到盐胁迫的强烈上调诱导。综合以上结果推测,β分枝中的所有基因在桑树生长发育及逆境胁迫中均有潜在的重要调控作用,γ分枝成员MnHD-Zip 8和MnHD-Zip 12则在桑树逆境胁迫响应中具有潜在重要功能。
桑树HD-Zip I亚家族基因的鉴定及表达分析
Identification and Expression Analysis of the HD-Zip I Subfamily Genes in Mulberry
-
摘要:
目的 研究桑树中HD-Zip I亚家族成员的进化及结构等特点,明确该家族基因的器官特异性表达模式,阐明ABA及淹水、盐和脱水处理对桑树HD-Zip I基因表达水平的影响。 方法 通过桑树基因组数据库鉴定桑树HD-Zip I基因,并进行生物信息学分析;利用桑树与拟南芥HD-Zip I蛋白的序列比对结果,构建系统进化树;利用桑树RNA-seq数据分析桑树HD-Zip I基因的组织/器官特异性表达谱;利用qRT-PCR分析桑树HD-Zip I基因在激素及非生物胁迫处理下的表达模式。 结果 共鉴定出14个桑树HD-Zip I基因,根据进化关系可进一步将其分为6个分枝,各分枝内的成员具有相似的基因结构及保守基序。RNA-seq数据分析发现,MnHD-Zip 2和MnHD-Zip 6在根、枝条、冬芽、雄花和叶片中均呈现高水平表达。激素处理后的基因表达分析发现,MnHD-Zip 8、MnHD-Zip 9、MnHD-Zip 11、MnHD-Zip 12和MnHD-Zip 13受到ABA不同程度的上调诱导,桑树其余HD-Zip I基因的表达水平均受到ABA不同程度的抑制。非生物胁迫处理表达分析发现,桑树HD-Zip I亚家族基因均受到3种非生物胁迫不同程度的诱导表达。 结论 桑树β分枝成员受到盐和脱水胁迫的显著诱导表达,且在各种器官中也有较广泛的高表达,因此,推测这些基因在桑树生长发育及逆境胁迫中均发挥着重要的调控作用。此外,MnHD-Zip 8和MnHD-Zip 12受到了盐、淹水及脱水胁迫的显著诱导,由此推测它们在桑树逆境胁迫响应中具有潜在重要功能。 -
关键词:
- 桑树
- / HD-Zip I转录因子
- / 鉴定
- / 基因表达分析
Abstract:Objective The aims of this study were to investigate the gene structure and phylogenetic of the mulberry (Morus alba L.) HD-Zip I subfamily members, clarify the tissue-specific expression of these genes in different organs, and reveal the expression level of the family genes under ABA and abiotic stress. Method Identification of putative HD-Zip I subfamily genes from MorusDB and bioinformatics analysis were performed. The evolutionary tree was constructed based on the multiple sequence alignment of the Arabidopsis and mulberry. RNA-seq data was used to analyze the tissue-specific expression of mulberry HD-Zip I genes. The transcription level of the family genes under hormone and abiotic stress was detected by qRT-PCR. Result A total of 14 HD-Zip I genes were identified in the mulberry genome. These members could be divided into six classes by phylogenetic tree. Members in the same class were similar in the gene structure and protein motifs. The expression level of both MnHD-Zip 2 and MnHD-Zip 6 were high in root, branch, winter bud, male flower, and leaf according to the RNA-seq data. All the MnHD-Zip I gene except MnHD-Zip 8, MnHD-Zip 9, MnHD-Zip 11, MnHD-Zip 12, and MnHD-Zip 13 were inhibited by ABA. The result of qRT-PCR showed that all the mulberry HD-Zip I genes could be induced by the abiotic stresses. Conclusion The β clades genes that were highly expressed in most of the organs were observably induced by the NaCl and dehydration. These results indicates that genes from the β clades of MnHD-Zip I play important roles in the development and stress response. In addition, MnHD-Zip 8 and MnHD-Zip 12 were dramatically up regulated by waterlogging, NaCl and dehydration, which indicates that these genes have essential roles in stress regulation of mulberry. -
Key words:
- Mulberry
- / HD-Zip I transcription factor
- / identification
- / gene expression analysis
-
图 1 桑树 HD-Zip I 蛋白 Homeodomain 及 leucine zipper 结构域多序列比对
Figure 1. Multiple sequence alignments of Homeodomain and leucine zipper domains
图 2 桑树与拟南芥 HD-Zip I 家族的系统进化分析
Figure 2. Phylogenetic analysis of HD-Zip I family in mulberry and Arabidopsis
图 3 桑树 HD-Zip I 家族蛋白保守基序(A)及基因结构(B)分析
Figure 3. Analysis of conserved motifs(A)and exon–intron organization(B)of MnHD-Zip I genes
图 4 桑树 HD-Zip I 家族基因的器官特异性表达
Figure 4. Organ specific expression analysis of MnHD-Zip I genes
图 5 桑树 HD-Zip I 家族基因激素处理后的差异表达分析
Figure 5. Temporal differential expression of MnHD-Zip I genes under ABA
图 6 桑树 HD-Zip I 家族基因胁迫处理后的差异表达分析
Figure 6. Temporal differential expression of MnHD-Zip I genes under abioticstresses
表 1 桑树HD-Zip I基因qRT-PCR分析引物
Table 1. Primers used in qRT-PCR analysis of HD-Zip I genes in mulberry
基因名称
Gene Name引物序列(5′–3′)
Primer sequence产物大小/bp
Product lengthMnHD-Zip 1 F:TTGACGGAGAAACTTCAGGCTAA R:TGAAAGTAGGAGTCACCGCTGTC 209 MnHD-Zip 2 F:TCAGAGGGTGAGGTCTCCAAAG R:GATTGGTCGGCTTCAAACACA 161 MnHD-Zip 3 F:CAGGCAGGTTGCTATTTGGTTT R:ATTTTCTCGCCCTCCTTCGT 208 MnHD-Zip 4 F:TGAAAAATAGAGAGCCAACGGAGT R:TGGAAAGAGAGGTCTGCTTGTGAT 176 MnHD-Zip 5 F:GTGTTTGAGCCAGAGCAGTCG R:TCGCACACGGGTCATCATAAC 127 MnHD-Zip 6 F:GCCCTAATGTCCGTCTGCC R:CCGCTTCCTCCACACATCC 121 MnHD-Zip 7 F:GGCTTGAAGTTGAAGGATACGG R:GACATCCCATTTAGCCCAAGTG 142 MnHD-Zip 8 F:TGGCTTCCCAGTTCGAGTCA R:CCAATGTCGTTTTCCAATCCC 166 MnHD-Zip 9 F:AGAAATGCCGTCTTGAATCCG R:ATCCTTCAGACACCACCCCAA 183 MnHD-Zip 10 F:CCTCGTCAGATTGCTGTTTGG R:GTAACCGCCCGTTGACACTT 198 MnHD-Zip 11 F:GCTAACAAAGGACCAAGTCAGGC R:GGCATTTTCAAGCCTCACCTG 205 MnHD-Zip 12 F:TAGGAAAAGACGCTGAAGTTGTAGA R:TCAAGACAAGAACTCCCACCACT 140 MnHD-Zip 13 F:TGAACAAGGCTCAACAGATGC R:ATCATCCACGGCTGACACTG 101 MnHD-Zip 14 F:ATCCTTCCCAACCACCTTCC R:TTCCAGCGAGCCCTCTTGT 136 ACTIN F:CCAAGTCATCACAATCGGAGC R:TACCAGGGAACATAGTTGAACCAC 191  下载: 导出CSV
下载: 导出CSV
表 2 桑树HD-Zip I家族基因及其编码蛋白信息
Table 2. The detailed information of HD-Zip I members of mulberry
基因名称
Gene Name登录号
Gene ID蛋白长度/aa
Protein length分子量/kDa
Molecular weight等电点
PI亲水性值
Hydropathicity亚细胞定位
Subcellular localizationMnHD-Zip 1 L484_008420 281 31634.93 4.70 −0.817 nucl:14 MnHD-Zip 2 L484_010105 325 36528.92 4.68 −0.839 nucl:14 MnHD -Zip 3 L484_027159 326 36766.68 6.54 −0.982 nucl:13.5, cyto_nucl: 7.5 MnH D-Zip 4 L484_021122 328 37377.30 6.42 −1.020 nucl: 13.5, cyto_nucl: 7.5 MnHD-Zip 5 L484_021079 292 33207.53 4.54 −0.922 nucl: 13, pero: 1 MnHD-Zip 6 L484_019806 342 39104.85 5.03 −0.997 nucl: 13.5, cyto_nucl: 7.5 MnHD-Zip 7 L484_012750 351 39913.22 4.94 −0.945 nucl: 13, pero: 1 MnHD-Zip 8 L484_001654 219 25604.55 6.66 −1.029 nucl: 14 MnHD-Zip 9 L484_009043 209 24036.19 6.93 −0.820 nucl: 14 MnHD-Zip 10 L484_002276 205 23523.59 8.53 −0.888 nucl: 14 MnHD-Zip 11 L484_012158 197 23221.98 6.66 −0.990 nucl: 14 MnHD-Zip 12 L484_015344 192 21891.73 8.88 −0.718 nucl: 14 MnHD-Zip 13 L484_010529 173 19927.40 7.89 −0.657 nucl: 14 MnHD-Zip 14 L484_023454 182 20630.97 6.78 −1.040 nucl: 14 注:nucl:细胞核;cyto:细胞质;pero:过氧物酶体。
Notes: nucl: nucleus; cyto: cytosol; pero: peroxisome.
下载: 导出CSV
表 3 桑树HD-Zip I家族基因启动子区顺式作用元件分析
Table 3. Analysis of cis acting elements in HD-Zip I gene’s promoter region
分枝
Clades基因名
Gene name激素响应元件
Phytohormone response element非生物胁迫响应元件
Abiotic stress response elementABRE P-box TCA-element CGTCA-motif TGACG-motif AuxRR-core TC-rich repeats LTR ARE MBS WUN-motif β MnHD-Zip 5 2 0 0 0 1 0 2 0 4 1 1 MnHD-Zip 2 1 0 2 1 1 1 0 1 2 0 0 MnHD-Zip 1 3 0 1 4 4 0 0 0 1 2 0 MnHD-Zip 7 1 0 1 0 0 0 3 0 5 1 0 MnHD-Zip 6 0 0 0 2 2 0 0 0 4 0 1 α MnHD-Zip 3 1 1 1 3 3 0 0 0 2 0 0 MnHD-Zip 4 3 0 0 1 1 0 0 1 1 2 0 γ MnHD-Zip 12 8 0 0 1 1 0 0 0 2 0 1 MnHD-Zip 8 2 1 3 0 0 0 0 1 2 0 2 MnHD-Zip 14 1 0 0 3 3 0 0 2 0 0 2 φ MnHD-Zip 11 6 0 0 0 0 0 0 0 5 0 1 MnHD-Zip 13 2 1 4 2 2 0 1 2 1 0 0 δ MnHD-Zip 9 4 2 2 4 4 0 0 0 2 0 0 ε MnHD-Zip 10 2 0 2 2 2 0 0 0 0 1 0
下载: 导出CSV
-
[1] Acosta-Motos J R, Ortuño M F, Bernal-Vicente A et al. Plant responses to salt stress: adaptive mechanisms[J]. Agronomy, 2017, 7(1): 18. doi: 10.3390/agronomy7010018 [2] Yamaguchi-Shinozaki K, Shinozaki K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses[J]. Annu. Rev Plant Biol, 2006, 57(1): 781-803. doi: 10.1146/annurev.arplant.57.032905.105444 [3] Ohama N, Sato H, Shinozaki Ket al. Transcriptional regulatory network of plant heat stress response[J]. Trends in plant science, 2017, 22(1): 53-65. doi: 10.1016/j.tplants.2016.08.015 [4] Dezar C A, Gago G M, González D H, et al. Hahb-4, a sunflower homeobox-leucine zipper gene, is a developmental regulator and confers drought tolerance to Arabidopsis thaliana plants[J]. Transgenic research, 2005, 14(4): 429-440. doi: 10.1007/s11248-005-5076-0 [5] Riechmann J, Heard J, Martin, G, et al. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes[J]. Science, 2000, 290(5499): 2105-2110. doi: 10.1126/science.290.5499.2105 [6] Ariel F D, Manavella P A, Dezar C A, et al. The true story of the HD-Zip family[J]. Trends in Plant ence, 2007, 12(9): 419-426. doi: 10.1016/j.tplants.2007.08.003 [7] Hu J, Chen G, Yin W, et al. Silencing of SlHB2 improves drought, salt stress tolerance, and induces stress-related gene expression in tomato[J]. Journal of Plant Growth Regulation, 2017, 36(3): 578-589. doi: 10.1007/s00344-017-9664-z [8] Henriksson E, Olsson A S, Johannesson H, et al. Homeodomain leucine zipper class I genes in Arabidopsis. Expression patterns and phylogenetic relationships[J]. Plant physiology, 2005, 139(1): 509-518. doi: 10.1104/pp.105.063461 [9] Agalou A, Purwantomo S, Övernäs E, et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members[J]. Plant molecular biology, 2008, 66(1-2): 87-103. doi: 10.1007/s11103-007-9255-7 [10] Lin Z, Hong Y, Yin M, et al. A tomato HD‐Zip homeobox protein, LeHB‐1, plays an important role in floral organogenesis and ripening[J]. The Plant Journal, 2008, 55(2): 301-310. doi: 10.1111/j.1365-313X.2008.03505.x [11] Manavella P A, Arce A L, Dezar C A, et al. Cross‐talk between ethylene and drought signalling pathways is mediated by the sunflower Hahb‐4 transcription factor[J]. The Plant Journal, 2006, 48(1): 125-137. doi: 10.1111/j.1365-313X.2006.02865.x [12] Olsson A, Engström P, Söderman E. The homeobox genes ATHB12 and ATHB7 encode potential regulators of growth in response to water deficit in Arabidopsis[J]. Plant molecular biology, 2004, 55(5): 663-677. doi: 10.1007/s11103-004-1581-4 [13] Shin D, Koo Y D, Lee J, et al. Athb-12, a homeobox-leucine zipper domain protein from Arabidopsis thaliana, increases salt tolerance in yeast by regulating sodium exclusion[J]. Biochemical and biophysical research communications, 2004, 323(2): 534-540. doi: 10.1016/j.bbrc.2004.08.127 [14] Zhang S, Haider I, Kohlen W, et al. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice[J]. Plant molecular biology, 2012, 80(6): 571-585. doi: 10.1007/s11103-012-9967-1 [15] Cabello J V, Arce A L, Chan R L. The homologous HD‐Zip I transcription factors HaHB1 and AtHB13 confer cold tolerance via the induction of pathogenesis‐related and glucanase proteins[J]. The Plant Journal, 2012, 69(1): 141-153. doi: 10.1111/j.1365-313X.2011.04778.x [16] Zhao Y, Ma Q, Jin X, et al. A novel maize homeodomain–leucine zipper (HD-Zip) I gene, Zmhdz10, positively regulates drought and salt tolerance in both rice and Arabidopsis[J]. Plant and Cell Physiology, 2014, 55(6): 1142-1156. doi: 10.1093/pcp/pcu054 [17] Khanna R, Kronmiller B, Maszle D R, et al. The Arabidopsis B-box zinc finger family[J]. The Plant Cell, 2009, 21(11): 3416-3420. doi: 10.1105/tpc.109.069088 [18] Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular biology and evolution, 2016, 33(7): 1870-1874. doi: 10.1093/molbev/msw054 [19] Gasteiger E, Gattiker A, Hoogland C, et al. ExPASy: the proteomics server for in-depth protein knowledge and analysis[J]. Nucleic acids research, 2003, 31(13): 3784-3788. doi: 10.1093/nar/gkg563 [20] Chen C, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular plant, 2020, 13(8): 1194-1202. doi: 10.1016/j.molp.2020.06.009 [21] He N, Zhang C, Qi X, et al. Draft genome sequence of the mulberry tree Morus notabilis[J]. Nature communications, 2013, 4(1): 1-9. [22] Jiao F, Luo R, Dai X, et al. Chromosome-level reference genome and population genomic analysis provide insights into the evolution and improvement of domesticated mulberry (Morus alba)[J]. Molecular Plant, 2020, 13(7): 1001-1012. doi: 10.1016/j.molp.2020.05.005 [23] 韩利红, 刘 潮, 刘学林, 等. 桑树PHR家族基因的鉴定及生物信息学分析[J]. 分子植物育种, 2020, 18(18):5991-5999. [24] 惠 甜, 沈兵琪, 王连春, 等. 桑树bHLH转录因子家族全基因组鉴定与分析[J]. 分子植物育种, 2019, 17(17):5624-5637. [25] 周 宏, 李荣芳, 陈丹丹, 等. 桑树Trihelix转录因子家族研究[J]. 基因组学与应用生物学, 2018, 37(2):874-880. [26] Zhao Y, Zhou Y, Jiang H, et al. Systematic analysis of sequences and expression patterns of drought-responsive members of the HD-Zip gene family in maize[J]. PloS ONE, 2011, 6(12): e28488. doi: 10.1371/journal.pone.0028488 [27] Mukherjee K, Brocchieri L, Bürglin T R. A comprehensive classification and evolutionary analysis of plant homeobox genes[J]. Molecular biology and evolution, 2009, 26(12): 2775-2794. doi: 10.1093/molbev/msp201 [28] Li R, Ge H, Dai Y, et al. Genomewide analysis of homeobox gene family in apple (Malus domestica Borkh. ) and their response to abiotic stress[J]. Journal of genetics, 2019, 98(1): 1-11. doi: 10.1007/s12041-018-1053-2 [29] Bhalothia P, Sangwan C, Alok A, et al. PP2C-like promoter and its deletion variants are induced by ABA but not by MeJA and SA in Arabidopsis thaliana[J]. Frontiers in plant science, 2016, 7: 547. [30] Son O, Hur Y-S, Kim Y K, et al. ATHB12, an ABA-inducible homeodomain-leucine zipper (HD-Zip) protein of Arabidopsis, negatively regulates the growth of the inflorescence stem by decreasing the expression of a gibberellin 20-oxidase gene[J]. Plant and Cell Physiology, 2010, 51(9): 1537-1547. doi: 10.1093/pcp/pcq108 [31] Re D A, Dezar C A, Chan R L, et al. Nicotiana attenuata NaHD20 plays a role in leaf ABA accumulation during water stress, benzylacetone emission from flowers, and the timing of bolting and flower transitions[J]. Journal of Experimental Botany, 2011, 62(1): 155-166. doi: 10.1093/jxb/erq252 [32] Ariel F, Diet A, Verdenaud M, et al. Environmental regulation of lateral root emergence in Medicago truncatula requires the HD-Zip I transcription factor HB1[J]. The Plant Cell, 2010, 22(7): 2171-2183. doi: 10.1105/tpc.110.074823 [33] Ge X X, Liu Z, Wu X M, et al. Genome-wide identification, classification and analysis of HD-ZIP gene family in citrus, and its potential roles in somatic embryogenesis regulation[J]. Gene, 2015, 574(1): 61-68. doi: 10.1016/j.gene.2015.07.079 [34] Shao J, Haider I, Xiong L, et al. Functional analysis of the HD-Zip transcription factor genes Oshox12 and Oshox14 in rice[J]. PLoS ONE, 2018, 13(7): e0199248. doi: 10.1371/journal.pone.0199248 [35] Liu W, Fu R, Li Q, et al. Genome-wide identification and expression profile of homeodomain-leucine zipper Class I gene family in Cucumis sativus[J]. Gene, 2013, 531(2): 279-287. doi: 10.1016/j.gene.2013.08.089 [36] Hjellström, M. Drought stress signal transduction by the HD-ZIP transcription factors ATHB6 and ATHB7[D]. Sweden: Universitatis Upsaliensis, 2002: 1-50. [37] Huang X, Duan M, Liao J, et al. OsSLI1, a homeodomain containing transcription activator, involves abscisic acid related stress response in rice (Oryza sativa L. )[J]. The Scientific World Journal, 2014, 2014: 1-9. [38] Skinner D J, Gasser C S. Expression-based discovery of candidate ovule development regulators through transcriptional profiling of ovule mutants[J]. BMC plant biology, 2009, 9(1): 1-20. doi: 10.1186/1471-2229-9-1 [39] Li C, Zhao M, Ma X, et al. The HD-Zip transcription factor LcHB2 regulates litchi fruit abscission through the activation of two cellulase genes[J]. Journal of experimental botany, 2019, 70(19): 5189-5203. doi: 10.1093/jxb/erz276 [40] He X, Wang T, Zheng X, et al. The cotton HD-Zip transcription factor GhHB12 regulates flowering time and plant architecture via the GhmiR157-GhSPL pathway[J]. Communications biology, 2018, 1(1): 1-8. doi: 10.1038/s42003-017-0002-6 [41] Aoyama T, Dong C H, Wu Y, et al. Ectopic expression of theArabidopsis transcriptional activator Athb-1 alters leaf cell fate in tobacco[J]. The Plant Cell, 1995, 7(11): 1773-1785. [42] Wang Y. The role of the homeobox gene ATHB16 in development regulation in Arabidopsis thaliana[J]. Acta Universitatis Upsaliensis, 2001. [43] Manavella P A, Dezar C A, Bonaventure G, et al. HAHB4, a sunflower HD-Zip protein, integrates signals from the jasmonic acid and ethylene pathways during wounding and biotic stress responses[J]. The Plant Journal, 2008, 56(3): 376-388. doi: 10.1111/j.1365-313X.2008.03604.x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5435
- HTML全文浏览量: 2679
- PDF下载量: 73
- 被引次数: 0
