


-
实时荧光定量PCR(qRT-PCR)是一种常见的检测基因表达水平的技术手段,它可以将常规PCR和荧光检测技术相结合,实时监控PCR扩增的过程[1],具有成本低、灵敏度高、特异性强等特点[2],被广泛应用于新基因挖掘及功能研究[3]。由于qRT-PCR技术的检测结果受到样本、实验条件、RNA质量及纯度、反转录效率等因素的影响[4],因此,想要获取准确度高的试验结果,就必须使用合适的内参基因进行校准[5]。
在大多数基因表达分析研究中,常选用能够维持细胞骨架或参与细胞基本生命过程的管家基因作为内参基因,如肌动蛋白基因(Actin)、α/β微管蛋白基因(TUA/TUB)、3-磷酸甘油醛脱氢酶基因(GAPDH),多聚泛素酶基因(UBQ)以及18S核糖体RNA(18S)等[6]。然而,近年来有研究发现,这类管家基因的表达稳定性也会受到物种和组织差异的影响[7]。如在连翘叶片中UKN1的表达最稳定,但在花和花蕾中最稳定的基因是ACT和SDH[8];在茉莉不同器官(根、茎、叶及花)中筛选出的理想内参也存在差异[9]。此外,同一物种内参基因的选择也受不同胁迫条件的影响,如EXP1和PP2A在高盐胁迫后的北沙参中是最稳定的内参基因,而在MeJA处理后CYP2和α-TUB是最稳定的内参基因[10]。杨树不同发育时期中筛选到的理想内参基因是U6-1、EIF4A和PP2A-2[11],而苏晓娟等发现,锌胁迫下杨树的actin、ubiquitin、EF1α和18S r RNA基因表达最为稳定[12],储文渊等发现,杨树在盐和干旱胁迫下,其新内参基因PtRG1,PtRG3和PtRG5比传统内参基因的表达更加稳定[13]。因此,内参基因并不具有通用性,在开展特定研究材料或实验条件的实时荧光定量分析前,首先应进行该物种特异性内参基因的筛选[14]。
灰楸(Catalpa fargesii Bur.)是紫葳科、梓属落叶乔木,是我国珍贵的用材树种和著名的园林观赏树种,素有“木王”之称。‘麦缘锦楸’是从灰楸实生苗选育出的新品种,其叶片呈现中间绿边缘黄的特征,目前已通过高干嫁接技术广泛应用于园林绿化等方面。课题组前期对‘麦缘锦楸’和灰楸的生理生化指标进行测定,发现不同颜色部位叶片的色素含量及叶绿素荧光参数存在显著差异[15]。想要进一步揭示该生理现象的分子机理,就需要分析‘麦缘锦楸’叶色形成途径中差异基因的表达模式[16],然而,目前尚未见梓树属对不同叶色表型筛选内参基因的相关报道,因此,开展‘麦缘锦楸’叶片内参基因的选择研究,以提高基因表达量的可靠性是十分必要的。本研究借助课题组前期的转录组数据,通过比较‘麦缘锦楸’和灰楸不同组织部位的基因表达量,初步筛选出表达相对稳定的6个基因CfUBC、CfActin11、>CfPP2A、CfMADH、CfGADPH、CfEF-1以及本课题组常用的内参基因CbuActin[17],共7个候选内参基因进行qRT-PCR分析,利用GeNorm、NormFinder和BestKeeper等软件综合分析并筛选出‘麦缘锦楸’不同叶色中表达相对稳定的内参基因;接着以萜类合成相关基因(CfGES)进一步验证上述分析结果的可靠性。本研究将使‘麦缘锦楸’与灰楸叶片基因表达的标准化和定量化更加准确,并为后期开展‘麦缘锦楸’黄绿叶色分区形成的分子研究奠定基础。
-
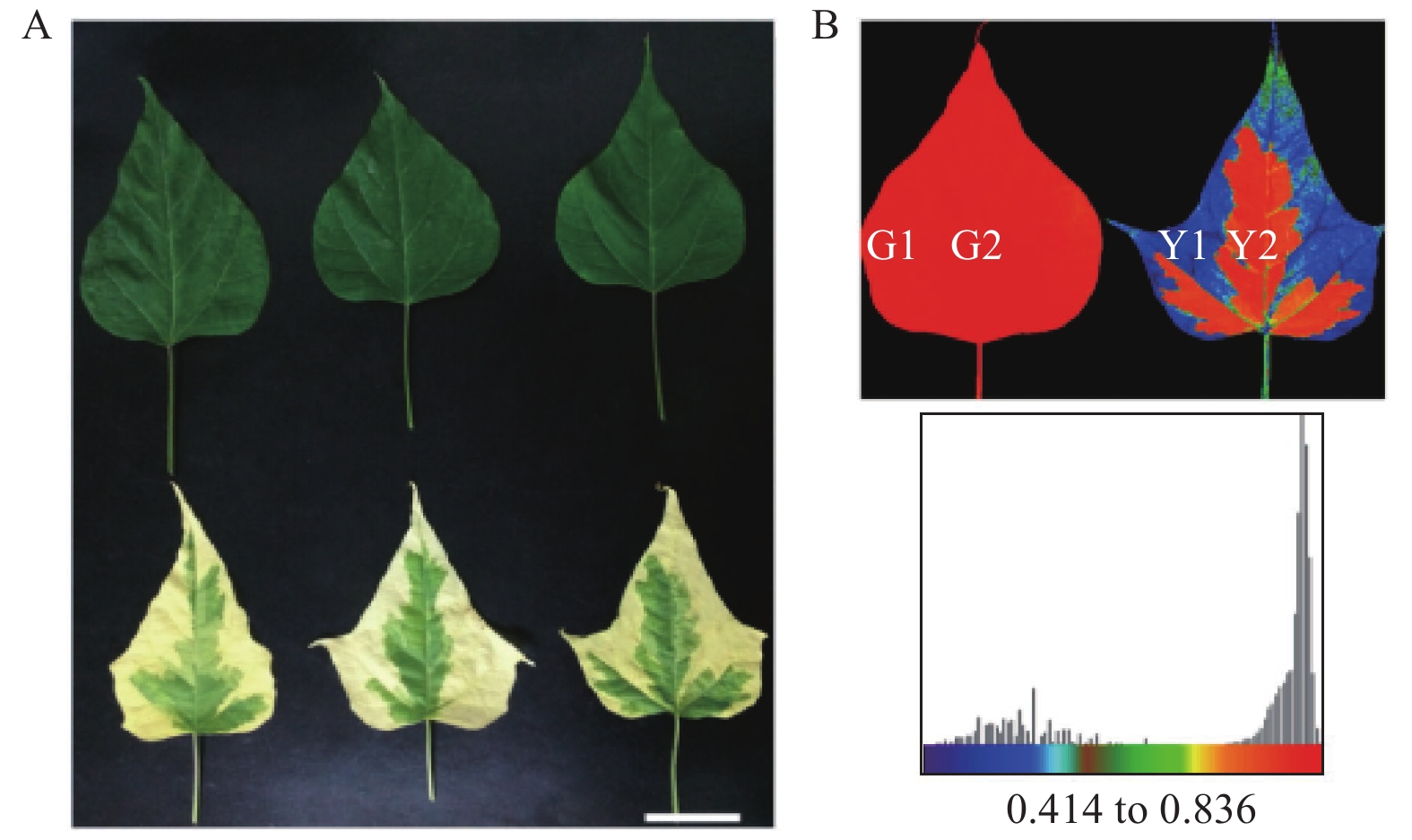
试验材料取自河南省洛阳市扁担赵基地。样品采集后,立即用刀片将‘麦缘锦楸’叶片黄绿部分切分,分别命名为Y1(黄色)、Y2(绿色),灰楸叶片对应部位分别命名为G1、G2(图1)。切好的叶片用锡箔纸包裹后立即置于液氮中速冻,并转至−80℃保存。所有的样品均设置3次生物学重复。
-
RNA提取按照EASY spin植物RNA快速提取试剂盒(北京艾德莱生物科技有限公司)的操作说明进行。通过1%琼脂糖凝胶电泳检测RNA样品的完整性,并利用超微量紫外分光光度计探头(Nanodrop 2000)检测所提取RNA的浓度与纯度。利用PrimeScript™ RT reagent Kit with gDNA Eraser试剂盒(Takara,RR047A),将RNA样品反转录为cDNA,获得的cDNA样品置于−20℃保存备用。
-
以课题组前期未发表的‘麦缘锦楸’和灰楸转录组数据为依据,选取FPKM值大于100且在样品间无显著差异表达的6个基因(CfUBC、CfActin、CfPP2A、CfMADH、CfGADPH、CfEF-1)为候选基因,加上课题组前期常用的CbuActin[17]共7个内参基因。
-
根据引物设计原则,利用在线工具Primer 3 Plus设计内参基因及CfGES基因引物,引物大小为18~27 bp,Tm值在58~61℃,扩增长度为150~250 bp,GC含量为40%~60%,并在NCBI上对引物进行特异性检测。引物由北京擎科生物科技有限公司合成,引物序列见表1。以各候选基因引物做普通PCR扩增,并用1%琼脂糖凝胶电泳检测。
表 1 内参基因及CfGES基因的引物设计
Table 1. Primer design for internal reference genes and CfGES gene
基因名称
Gene引物序列
Primer sequences (5′-3′)退火温度
Tm/℃CfUBC F: CCACTGTTTCGGCCAGTATT
R: GGGTGCCCTCCCATATAAGT60.0
60.0CfActin11 F: TTCCCTGCTTCACTCGATCT
R: TTCCCGGATAACTGAGCATC60.0
60.0CfGADPH F: GGAATCACAGCCGAAGATGT
R: ACCTTGACCATGTCGTCTCC60.1
60.0CfEF-1 F: AGTTTACGCCGCTGTTTTCG
R: CAAGGTCGTCGTCATCATCG60.0
60.0CfPP2A F: GAGCAGTTGCACAGAATGGA
R: CCTGGCGAATTAGAAATGGA60.0
60.0CfMADH F: AGCTTCCATTCTTTGCCTCA
R: TCCGAACAAAAGCAATACCC60.0
59.9CbuActin[17] F: GATGATGCTCCAAGAGCTGT
R: TCCATATCATCCCAGTTGCT60.0
60.0CfGES F: GAGCTACCAAGGCACCTGAG
R: CTCCTTCCACCACCTGGATT59.5
60.0 -
将cDNA模板混合稀释8倍后,按照Takara公司的TB Green Premix Ex Taq (Tli RNaseH Plus) 试剂盒(Takara,RR420A)说明书进行实时荧光定量PCR实验。反应体系为20 μL:TB Green Premix Ex Taq 10 μL,上游引物(10 μmol·L−1)0.8 μL,下游引物(10 μmol·L−1)0.8 μL,DNA模板1 μL,dd H2O 7.4 μL。每个样品设置3个重复,所有操作均在冰上进行。利用LightCycler480实时荧光定量PCR仪对各样品进行扩增,PCR扩增程序为:95℃预变性30 s;定量分析40个循环:95℃ 变性5 s,60℃退火30 s;融解曲线:95℃ 5 s,60℃ 1 min后缓慢上升至95℃;降温:50℃ 30 s。
-
将各样品得到的Ct值按公式Q = E(minCt-sampleCt)(E为扩增效率,默认值为2;minCt为基因在样品中的最小Ct值;sampleCt为基因在样品中的Ct值)进行计算,将得到的Q值导入GeNorm和NormFinder软件中进行稳定性分析。将各样品得到的Ct值输入到BestKeeper软件中,得到候选内参基因的表达稳定性(M值)、稳定值(SV值)、变异系数(CV值)和标准差(SD值),进行表达稳定性分析。使用3款软件分析后,将各样品得到的Ct值输入到在线网站RefFinder(
https://www.heartcure.com.au/for-researchers/ )中进行综合分析,得出最适合‘麦缘锦楸’不同叶色部位稳定表达的内参基因。 -
分别以综合分析排名最靠前的2个基因为内参,对萜类合成基因CfGES的表达情况进行分析,qRT-PCR方法参照1.2.4,结合CfGES基因的转录组数据,对2个内参基因进行表达稳定性验证。
-
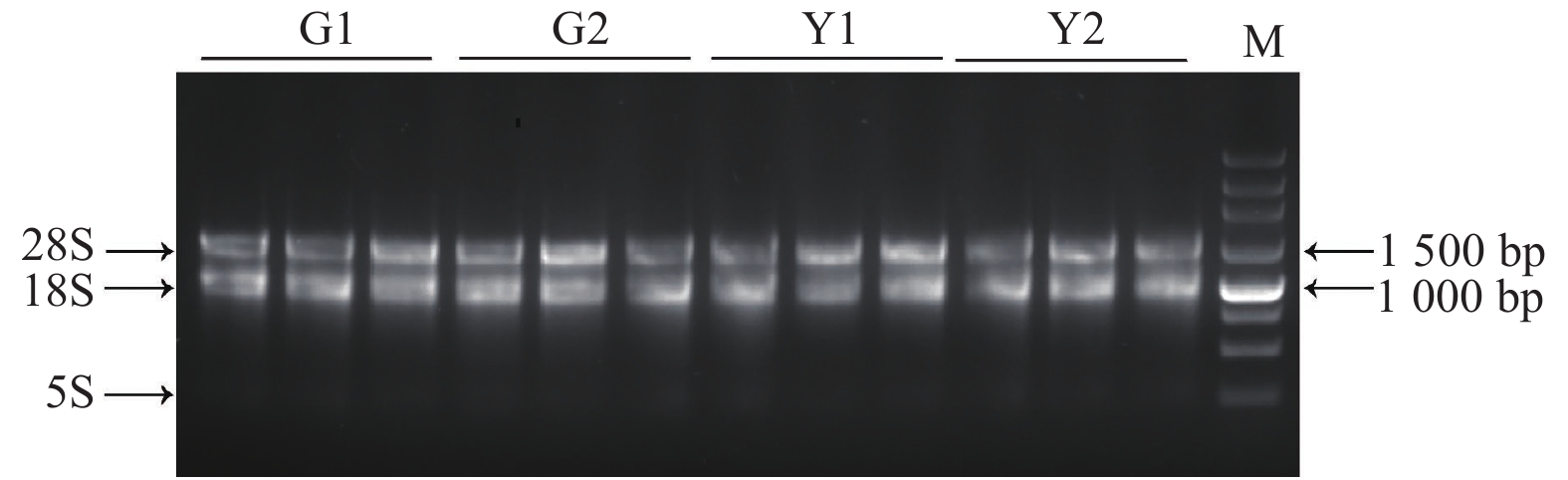
Nanodrop检测到所有样品总RNA的OD260/280及OD260/230值均在1.8~2.2之间,说明所提RNA纯度较好,1%琼脂糖凝胶电泳检测RNA样品完整性(图2),各样品的28S和18S条带明显,说明总RNA完整性较好,均可用于后续试验。
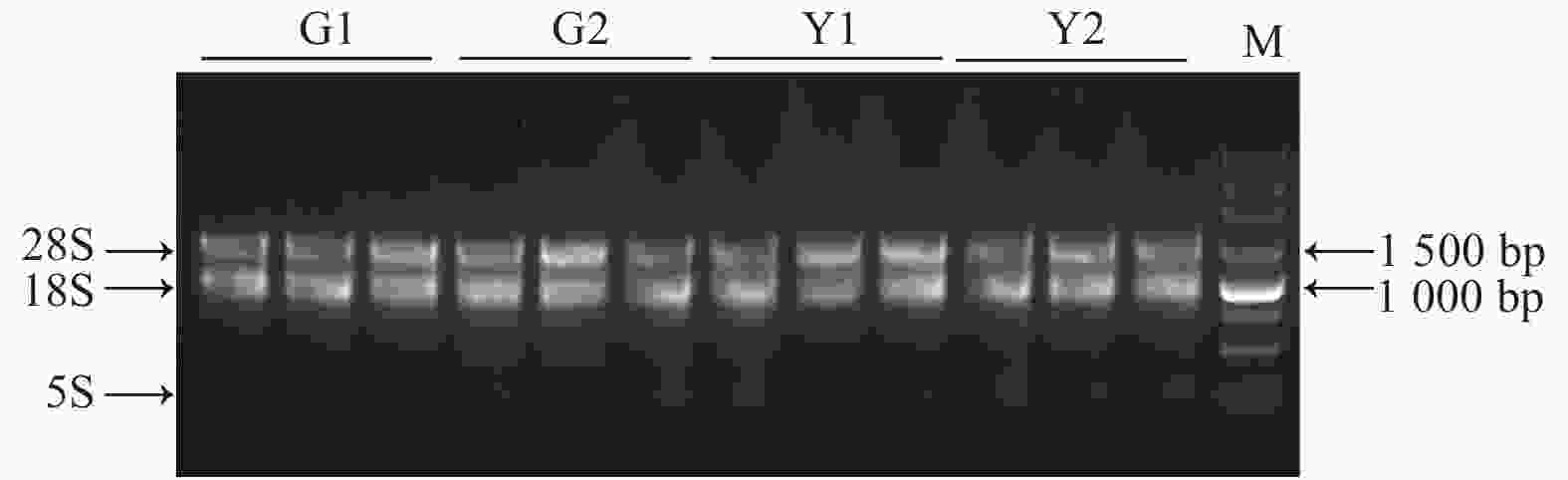
图 2 ‘麦缘锦楸’和灰楸不同叶色部位总RNA电泳图
Figure 2. Total RNA electrophoresis of different leaf color parts of 'Maiyuanjinqiu' and C.fargesii.
-
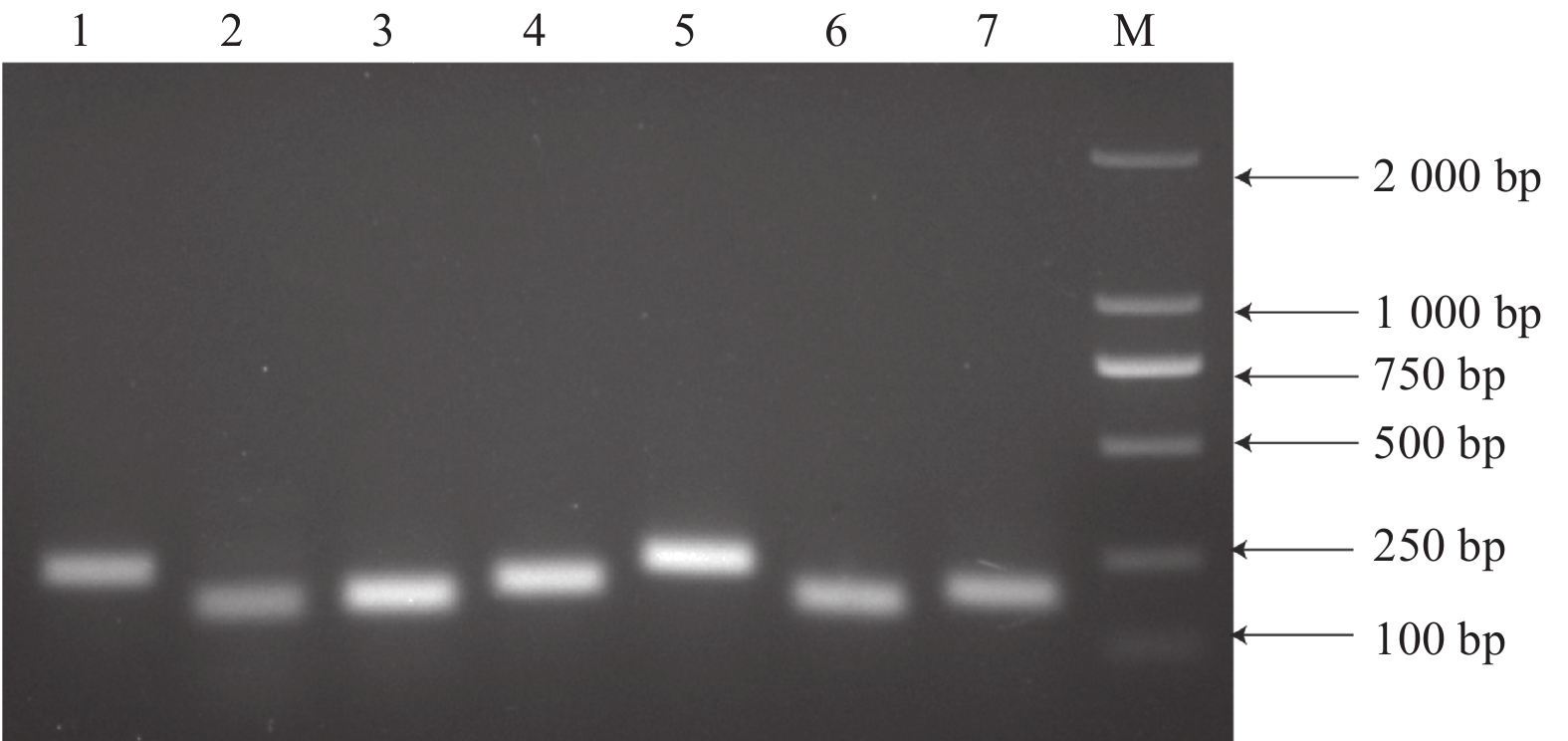
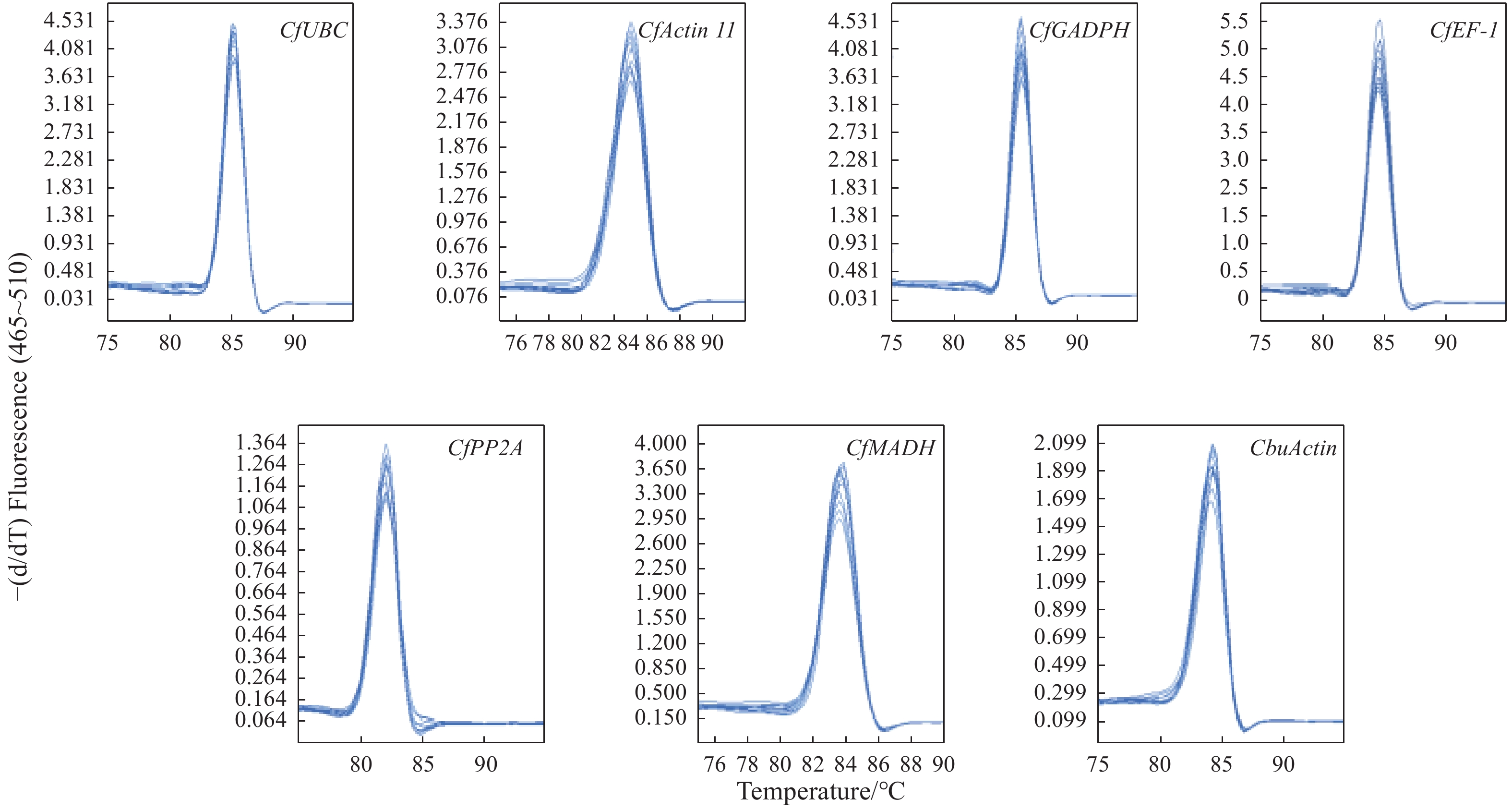
图3显示:所有引物均能扩增出单一且亮的条带,无引物二聚体,条带大小与预期相符,引物特异性完好,可用于后续检测分析。对7个候选内参基因在各个样品的qRT-PCR分析表明:各基因Ct值均在23~35,且溶解曲线都呈现显著单一的峰(图4),表明qRT-PCR所用引物可与模板cDNA特异性结合并扩增靶基因。

图 3 7种内参基因的PCR扩增产物
Figure 3. PCR products of seven reference genes.

图 4 7个内参基因溶解曲线
Figure 4. Real-time PCR melting curves of seven reference genes
-
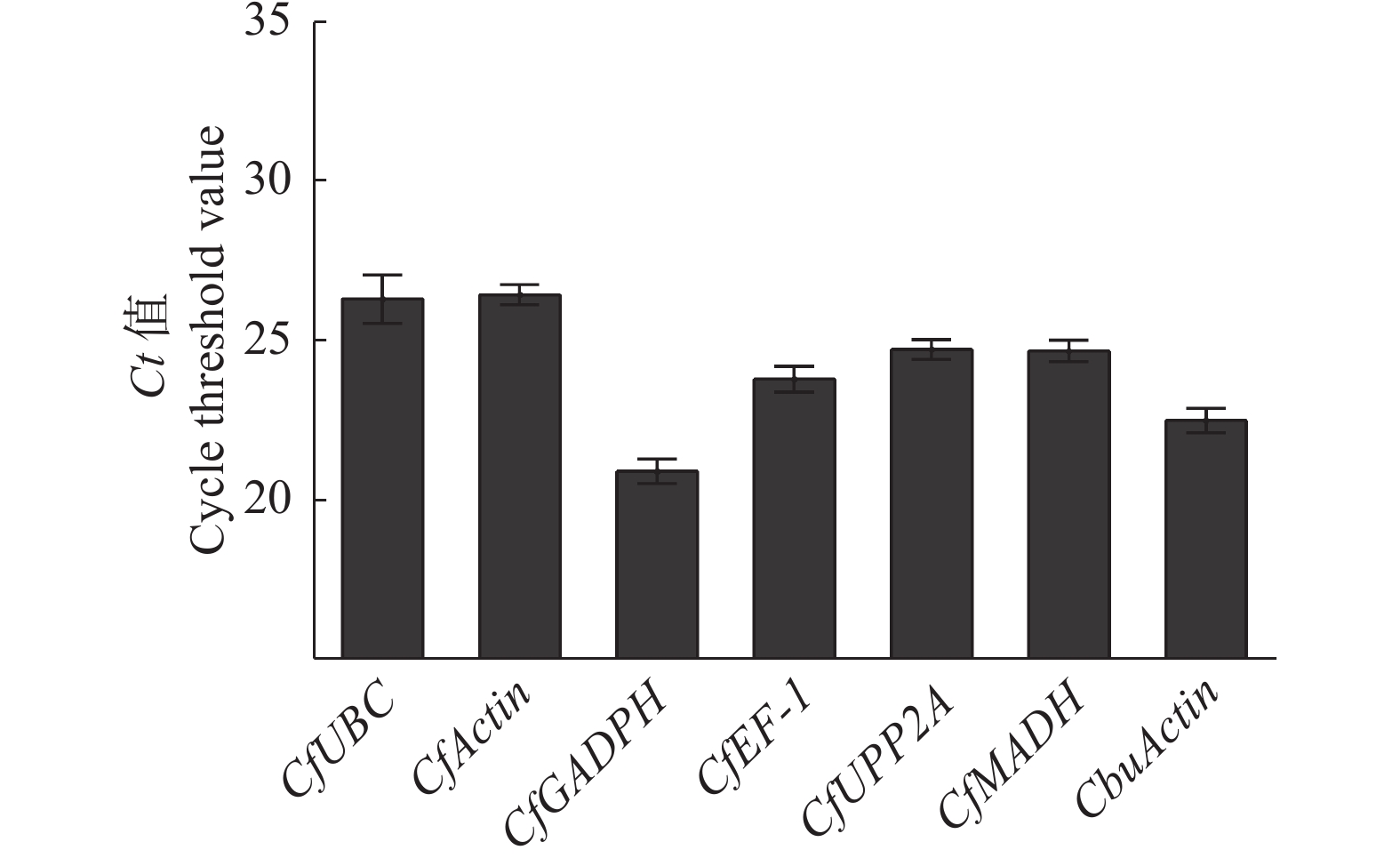
7个候选内参基因中,CfActin的平均Ct值最大,为26.459,说明该基因的表达丰度最低;CfGADPH的平均Ct值最小,为20.91,说明该基因的表达丰度最高。7个内参基因的表达丰度大小排序依次是:CfGADPH > CbuActin > CfEF-1 > CfMADH> CfPP2A > CfUBC > CfActin(图5)。
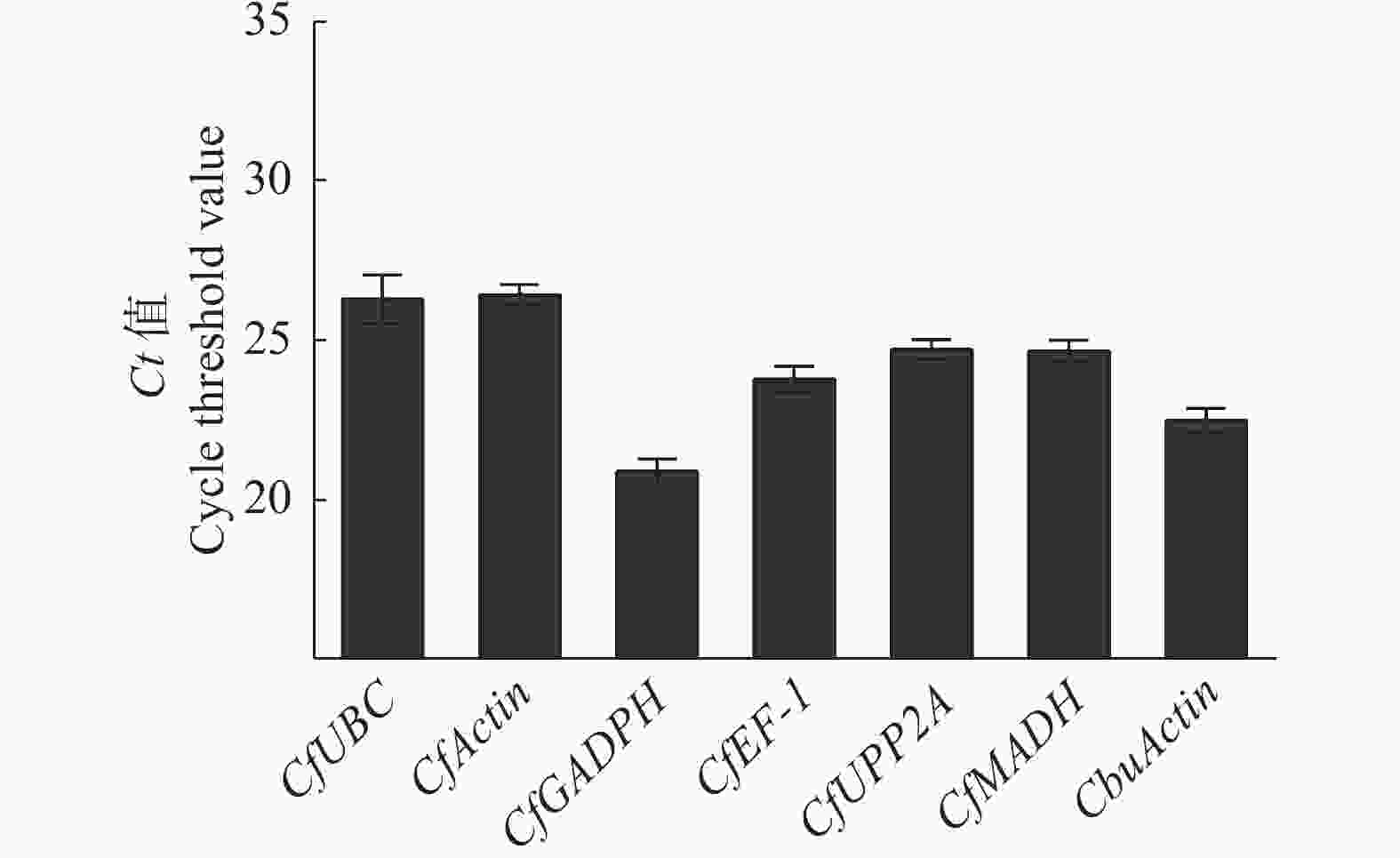
图 5 7个内参基因平均Ct值
Figure 5. Average Ct values of seven reference genes
-
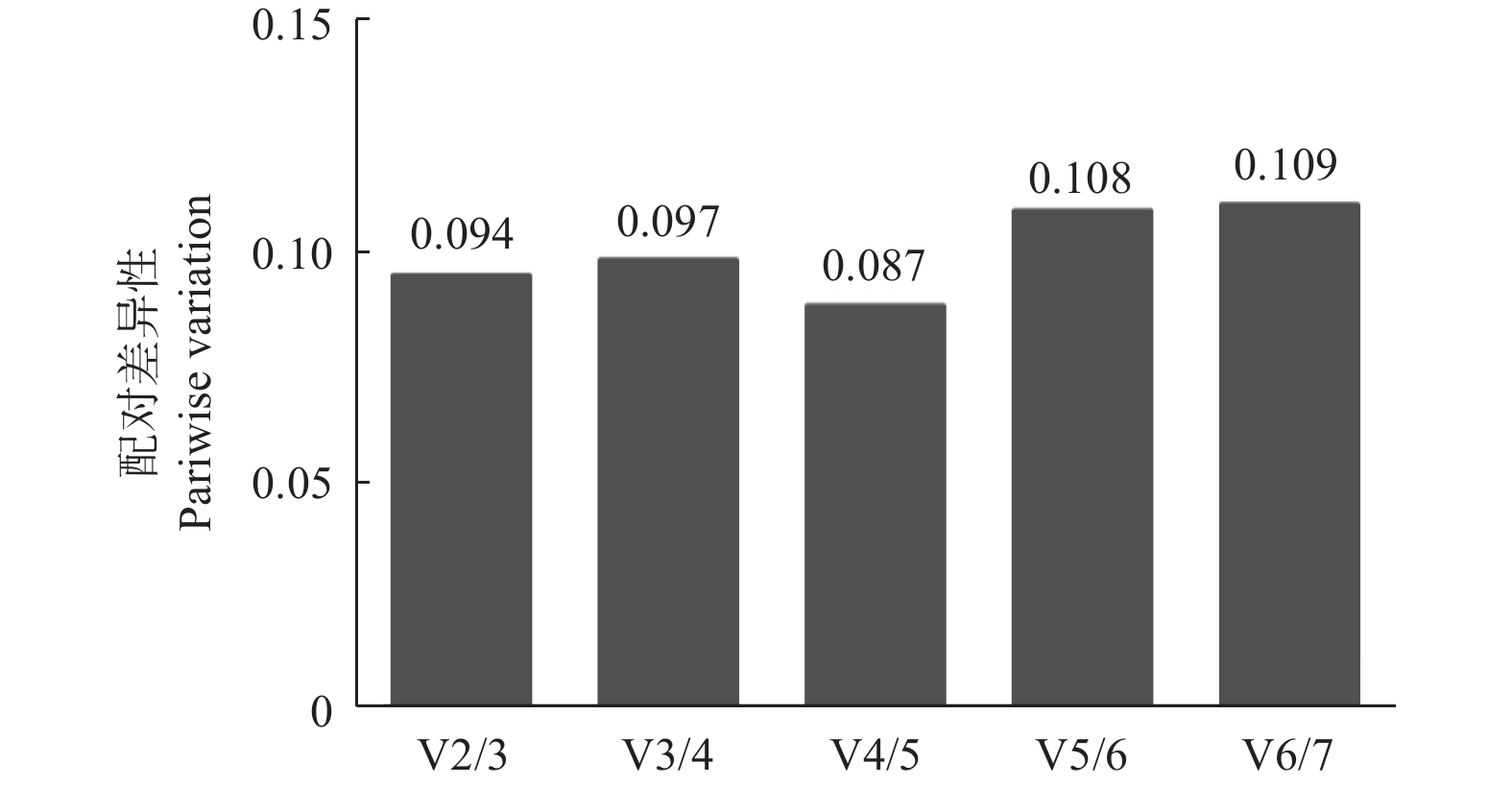
GeNorm是通过比较计算内参基因稳定性的M值,以确定表达最稳定的内参基因。该软件以M=1.5作为临界点,低于该值表明基因的表达相对稳定,且M值越小表示内参基因的表达越稳定[18]。GeNorm分析结果表明:7个候选内参基因的M值均小于1.5(表2),即各候选内参的表达都相对稳定,稳定性排名从高到低依次是:CfMADH > CfEF-1 > CfGADPH > CfPP2A > CfActin11 > CfUBC > CbuActin。为确定内参基因的最佳数量,进一步通过配对变异系数Vn / (n + 1),得出以4个候选基因同时做为内参基因的效果最佳,2个组合使用次之(图6)。
表 2 GeNorm软件分析下内参基因的表达稳定性
Table 2. Analysis of the expression stability of reference genes in leaves by GeNorm
基因名称
Gene name平均表达稳定值
M稳定性排行
Stability rankCfMADH 0.429 1 CfEF-1 0.442 2 CfGADPH 0.467 3 CfPP2A 0.602 4 CfActin11 0.610 5 CfUBC 0.666 6 CbuActin 0.790 7 
图 6 GeNorm软件分析最适合内参基因数目
Figure 6. The number of the most suitable reference genes analyzed by GeNorm software
-
NormFinder的计算原理与GeNorm相似,该软件是基于Excel结合组间方差与组内方差计算SV值,来确定内参基因的稳定性。SV值的大小与基因的稳定性呈负相关。NormFinder分析结果(表3)显示:CfUBC、CbuActin、CfActin11、CfPP2A、CfGADPH、CfEF-1、CfMADH基因在叶片不同颜色部位的表达稳定值分别为0.406、0.520、0.329、0.340、0.068、0.095、0.062,其中,CfMADH、CfEF-1和CfGADPH的SV值最小,说明这3个基因的稳定性较强。
表 3 NormFinder软件分析‘麦缘锦楸’及灰楸不同叶色部位内参基因的表达稳定性
Table 3. Analysis of the expression stability of reference genes by NormFinder software
基因名称
Gene name稳定指数
Stability value稳定性排行
Stability rankCfMADH 0.062 1 CfGADPH 0.068 2 CfEF-1 0.095 3 CfActin11 0.329 4 CfPP2A 0.340 5 CfUBC 0.406 6 CbuActin 0.520 7 -
BestKeeper通过计算变异系数(CV)和标准偏差(SD)来反映内参基因的稳定性,CV和SD的值与基因的稳定性负相关,其中,SD值的默认阈值为1.0,低于该值即认为表达稳定[19]。BestKeeper分析结果(表4)显示:在灰楸及‘麦缘锦楸’叶片中,7个基因的SD值均小于1.0,说明各基因的表达均较为稳定,并且稳定性从高到低排序依次是:CfActin11、CfMADH、CfGADPH、CfEF-1、CbuActin、CfPP2A、CfUBC,其中CfActin11的CV和SD值最小(CV=0.80;SD=0.21),表达最为稳定;CfUBC的CV和SD值最大(CV=2.35;SD=0.62),其表达最不稳定。
表 4 BestKeeper软件分析灰楸及‘麦缘锦楸’不同叶色部位内参基因的表达稳定性
Table 4. BestKeeper software was used to analyze the expression stability of internal reference genes in different leaf color sectors of Maiyuanjinqiu and C.fargesii.
基因名称
Gene name几何平均值
Geometric mean均值
Mean最小值
Min最大值
Max标准差
SD变异系数
CV稳定性排行
Stability rankCfActin11 26.46 26.46 26.08 26.82 0.21 0.80 1 CfMADH 24.69 24.69 24.33 25.01 0.27 1.10 2 CfGADPH 20.91 20.91 20.64 21.49 0.29 1.38 3 CfEF-1 23.81 23.81 23.36 24.22 0.33 1.38 4 CbuActin 22.54 22.54 22.06 22.97 0.37 1.66 5 CfPP2A 28.54 28.55 27.78 29.06 0.46 1.62 6 CfUBC 26.31 26.32 25.48 27.19 0.62 2.35 7 -
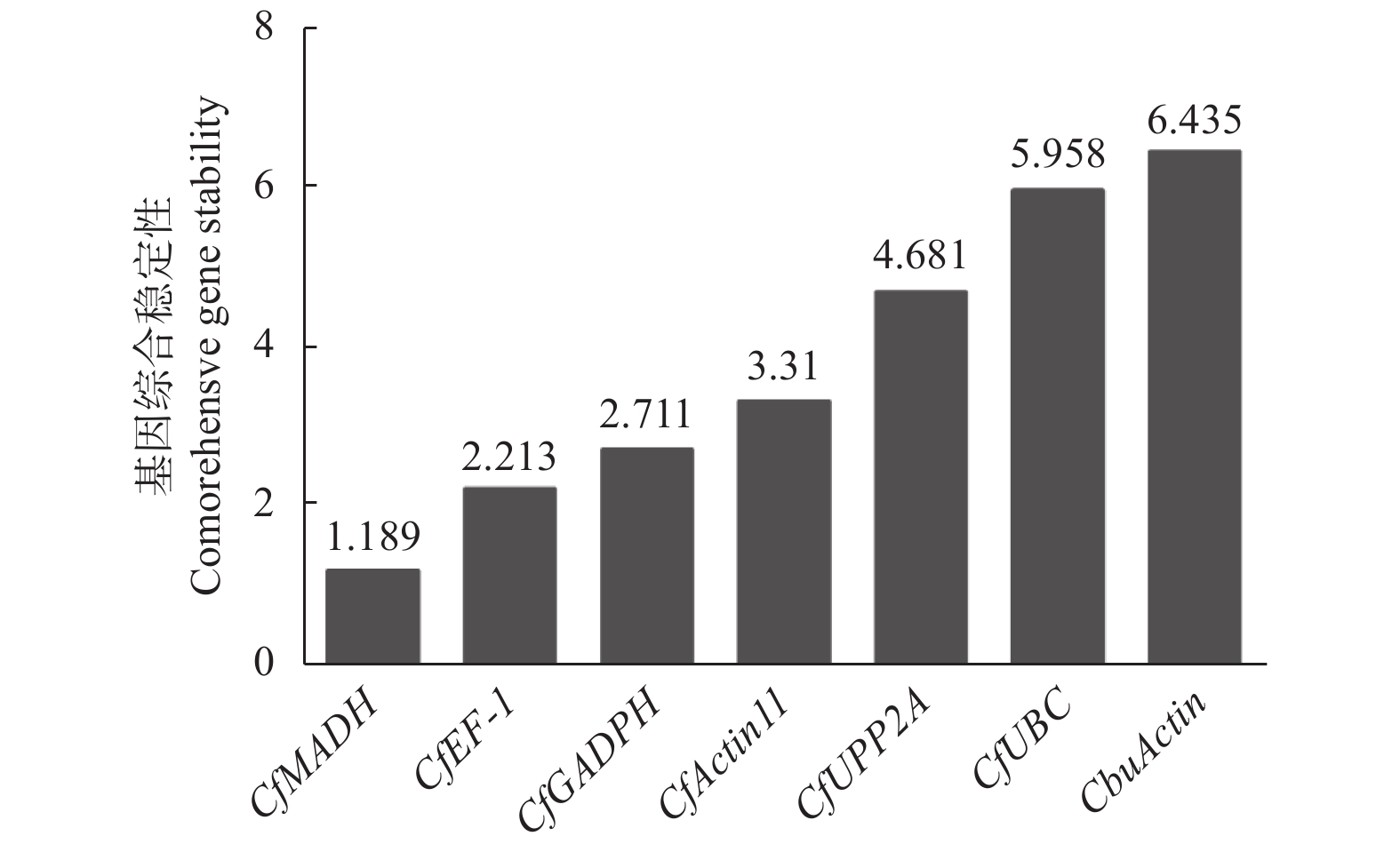
对GeNorm、NormFinder以及BestKeeper 3种软件的数据进行整合分析,综合评估基因的表达稳定性。将各基因在不同样品中的Ct值导入在线网站RefFinder(
https://www.heartcure.com.au/for-researchers/ )中。结果显示,各基因稳定性综合排名由高到低依次为CfMADH > CfEF-1 > CfGADPH > CfActin11 > CfPP2A > CfUBC > CbuActin (图7),其中,CfMADH和CfEF-1的表达稳定性最好,符合qRT-PCR实验内参基因的选择标准。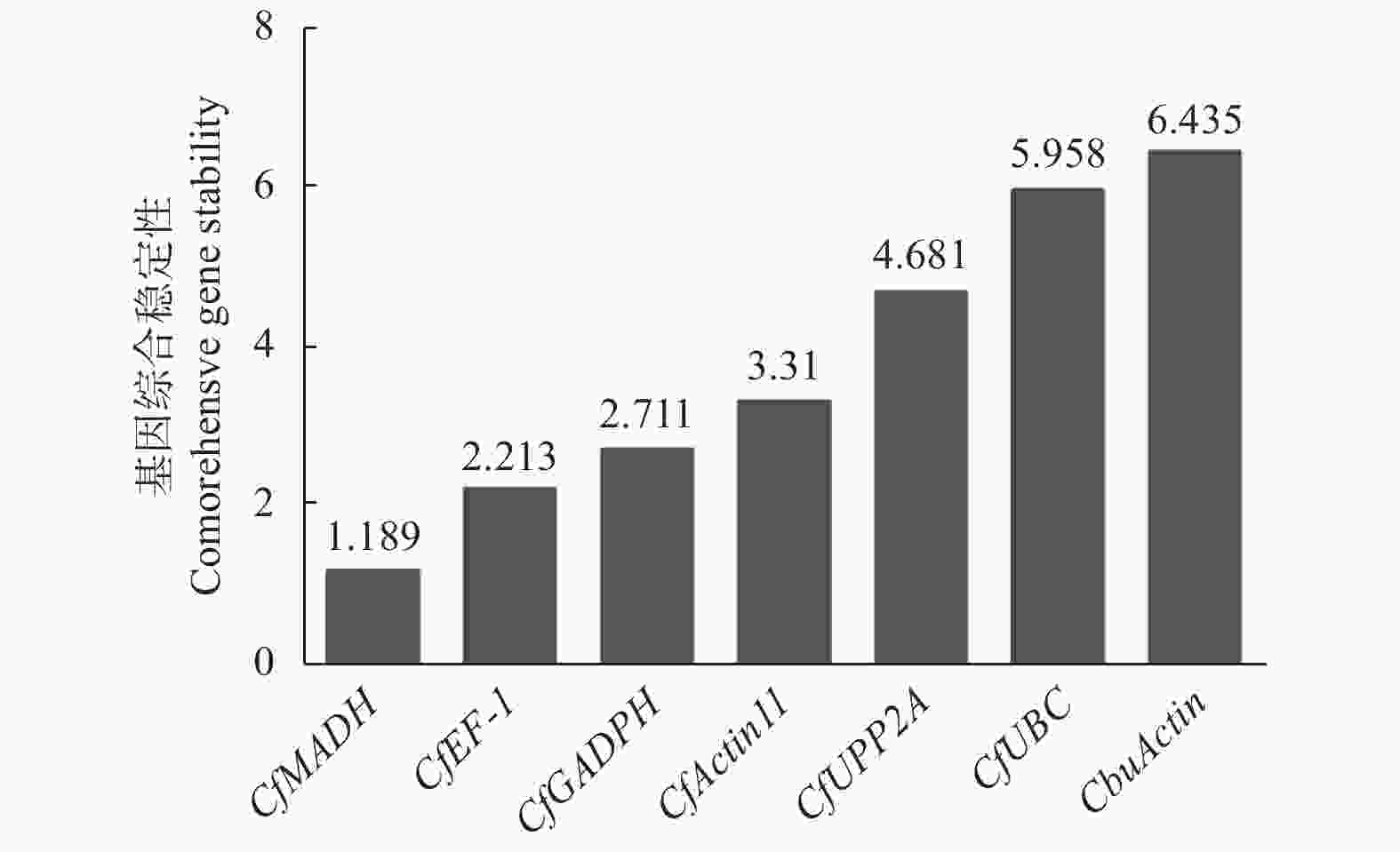
图 7 RefFinder分析7个候选内参基因的稳定性综合排名
Figure 7. Stability ranking of seven genes by RefFinder analysis
-
分别以CfMADH和CfEF-1为内参,分析萜类合成酶基因CfGES在‘麦缘锦楸’和灰楸不同叶色部位(Y1、Y2、G1、G2)的表达量差异,以验证软件预测结果的可靠性。结果表明:单独或组合使用CfMADH及CfEF-1基因为内参时,CfGES基因的表达差异与转录组测序结果趋势一致(图8)。因此,本研究结果表明:单独使用CfMADH、CfEF-1基因或组合使用这2个基因,能够准确校准‘麦缘锦楸’不同叶色部位的荧光定量结果。

图 8 CfGES在‘麦缘锦楸’和灰楸不同叶色部位的基因表达量差异
Figure 8. The expression differences of CfGES among different leaf color sectors of 'Maiyuanjinqiu' and C. fargesii .
-
基因表达分析是分子生物学科中最为重要的研究内容之一,qRT-PCR是当前最常用的检测基因表达情况的技术手段,常用来检测基因在不同样品、组织、生长发育时期及特定实验条件下的表达模式[20]。对基因进行表达情况分析时,必须选择准确的内参基因做标准化分析[21-22]。众多研究中发现,管家基因可作为内参基因来进行表达量分析。然而近年来有研究发现,内参基因并不具有通用性,需要根据特定的物种或实验条件来确定最适宜的内参基因[23-24]。如GADPH在黄山栾树、银杏、肉桂和大叶清化桂中稳定表达[25-27],但在丝瓜和苎麻中却被认为是最不理想的内参基因[28-29]。18SrRNA基因在红豆杉多种处理下都表现出较好的稳定性[30],但在稻瘟病菌侵染的水稻、灵芝及红木不同处理下该基因的表达最不稳定[31-32]。目前尚未发现能在各类实验条件下都稳定表达的内参基因,所以研究者需要根据自己的实验条件找到可以作为标准化的基因进行定量分析。
GeNorm、NormFinder和BestKeeper是基因组研究中筛选稳定内参基因最常见的计算软件,本研究应用3款软件及1个分析网站对7个候选内参基因的稳定性分析发现,这7个内参基因在‘麦缘锦楸’和灰楸叶片中均稳定表达,但稳定性排名略有差异:在GeNorm和NormFinder中均得出CfMADH和CfEF-I是最稳定的内参基因,而在BestKeeper
中CfMADH和CfEF-I的排名却分别居于第2和第4名,排名的差异可能是由于3款软件所设定的统计学算法不同所导致的。GeNorm和NormFinder的原理基本相似,都是将qRT-PCR所得到的Ct值转化为基因相对表达量,再进行最适内参的分析,每个候选基因的稳定性取决于单个样品的最小Ct值;而BestKeeper则直接在内置公式中输入各基因表达的Ct值来进行分析,候选基因的稳定性与每个样品Ct值的离散程度相关,因此,该算法易受极端值影响,无法规避系统误差[33-34]。考虑到每款软件的局限性,笔者利用RefFinder对这3款软件得出的结果进行综合分析,确定了7个候选内参基因在‘麦缘锦楸’叶片中的稳定性排名依次是:CfMADH > CfEF-1 > CfGADPH > CfActin11 > CfPP2A > CfUBC > CbuActin。在GeNorm分析的内参基因变异系数配对值来看,n=4时,Vn / (n+1)的比值最小,是最佳的内参基因组合数量,n=2次之,但综合考虑实验成本及样品用量问题本文认为组合使用2个候选基因作为内参更为合适,此前也有研究认为以2个或2个以上基因为内参更能校准定量分析实验上的系统偏差[35]。 萜类物质(叶绿素、胡萝卜素)在叶色形成过程中有重要的作用。在本课题组转录组数据中发现,萜类合成酶基因CfGES 在‘麦缘锦楸’和灰楸不同叶色部位中差异表达,可以验证候选内参基因的稳定性。CbuActin是课题组前期利用同源克隆法获得的内参基因[17],在‘麦缘锦楸’和灰楸叶片中其M<1.5(M=0.790),符合作为内参基因的标准,但本研究通过软件分析排名最好的内参基因是CfMADH和CfEF-1。笔者分别以这2个最适合的内参基因及其组合来校准CfGES的表达量,结果表明单独或组合使用CfMADH和CfEF-1时,CfGES的表达差异与转录组数据一致,进一步表明了本实验结果的可靠性。本研究所筛选到的最适内参基因CfMADH和CfEF-1均为真核生物中常见的管家基因,在其他观赏性树种(紫薇、花叶唐竹等)的不同叶色叶片中也被报道可作为理想内参基因[36-38]。此外,本研究材料中CfUBC和CfActin11在‘麦缘锦楸’叶片中稳定性不佳,但在其他物种(如景宁木兰、板栗)中却表现出很好的稳定性[39-40],这些结论也体现了内参基因不具有通用性的特点。
-
本研究根据课题组前期研究及转录组数据筛选出7个候选基因(CfUBC、CfActin11、CfPP2A、CfMADH、CfGADPH、CfEF-1及CbuActin),结合qRT-PCR技术及GeNorm、NormFinder和BestKeeper等内参分析软件对各候选基因进行稳定性分析,结果表明CfMADH和CfEF-1是最适合‘麦缘锦楸’和灰楸不同叶色部位的内参基因,萜类合成基因CfGES验证了软件分析结果的可靠性。本研究优化了紫葳科植物‘麦缘锦楸’内参基因的选择,将为‘麦缘锦楸’叶色形成的分子生物学机制提供理论基础,也将为课题组及其它植物研究选择合适内参基因提供参考依据。
‘麦缘锦楸’叶色表型qRT-PCR内参基因筛选及验证
Selection and Validation of Reference Genes for Leaf Color Phenotype in 'Maiyuanjinqiu', a Catalpa fargesii Variety, by qRT-PCR
-
摘要:
目的 筛选不同叶色部位稳定表达的内参基因,为后期开展‘麦缘锦楸’叶色形成的分子机制奠定基础。 方法 以‘麦缘锦楸’不同叶色部位及灰楸对应部位叶片为材料,利用实时荧光定量PCR(qRT-PCR)技术检测了7个候选基因(CfUBC、CfActin11、CfPP2A、CfMADH、CfGADPH、CfEF-1及CbuActin)的相对表达量,利用GeNorm、NormFinder和BestKeeper 等分析软件对内参基因进行了稳定性评价。分析了‘麦缘锦楸’和灰楸不同叶色部位中萜类合成相关基因(CfGES)的表达模式,验证了上述稳定性评价结果的可靠性。 结果 7个候选内参基因在‘麦缘锦楸’和灰楸叶片中均具有作为内参基因的性能,其中,CfMADH和CfEF-1在不同叶色部位的表达量最稳定,CfGADPH和CfActin11次之,CfUBC最差。以萜类合成相关基因CfGES验证内参稳定性发现,单独或组合使用CfEF-1及CfMADH为内参基因时,CfGES的表达差异与转录组数据趋势一致。 结论 单独或组合使用CfMADH和CfEF-1来校准‘麦缘锦楸’不同叶色部位的基因表达量,可显著提高实验结果的可靠性。 Abstract:Objective To select the housekeeping genes stably expressed in leaves in order to support the study of the molecular mechanism of leaf color formation on 'Maiyuanjinqiu', a Catalpa fargesii variety. Method The relative expression of seven candidate genes, including CfUBC, CfActin11, CfPP2A, CfMADH, CfGADPH, CfEF-1 and CbuActin were detected by real-time quantitative PCR (qRT-PCR) technology using leaves of different leaf color sectors of 'Maiyuanjinqiu' and the corresponding sectors of Catalpa fargesii Bur. The stability of the seven candidate reference genes was evaluated by using internal reference analysis software such as GeNorm, NormFinder and BestKeeper. Furthermore, the expression patterns of terpenoid biosynthesis related gene (CfGES) in the leaves of different leaf color parts of 'Maiyuanjinqiu' and the corresponding sectors of C. fargesii were analyzed respectively. The results verified the stability evaluation results mentioned above. Result All the seven candidate reference genes can be used as internal reference genes in the leaves of 'Maiyuanjinqiu' and C. fargesii. The expression levels of CfMADH and CfEF-1 were the most stable in different color tissues, followed by CfGADPH and CfActin11, while the CfUBC was the worst. The stability of internal reference gene was verified by terpenoid biosynthesis related gene (CfGES). It was found that the expressions of CfGES gene were consistent with the trend of transcriptome data, when CfEF-1 and CfMADH genes were used as internal reference for qRT-PCR alone or in combination. Conclusion Using CfMADH and CfEF-1 individually or in combination to calibrate the gene expression levels of different leaf color parts in 'Maiyuanjinqiu' can greatly improve the reliability of experimental results. -
Key words:
- Catalpa fargesii; 'Maiyuanjinqiu'
- / leaf
- / reference gene
- / qRT-PCR
-
图 2 ‘麦缘锦楸’和灰楸不同叶色部位总RNA电泳图
Figure 2. Total RNA electrophoresis of different leaf color parts of 'Maiyuanjinqiu' and C.fargesii.
图 6 GeNorm软件分析最适合内参基因数目
Figure 6. The number of the most suitable reference genes analyzed by GeNorm software
图 7 RefFinder分析7个候选内参基因的稳定性综合排名
Figure 7. Stability ranking of seven genes by RefFinder analysis
图 8 CfGES在‘麦缘锦楸’和灰楸不同叶色部位的基因表达量差异
Figure 8. The expression differences of CfGES among different leaf color sectors of 'Maiyuanjinqiu' and C. fargesii .
表 1 内参基因及CfGES基因的引物设计
Table 1. Primer design for internal reference genes and CfGES gene
基因名称
Gene引物序列
Primer sequences (5′-3′)退火温度
Tm/℃CfUBC F: CCACTGTTTCGGCCAGTATT
R: GGGTGCCCTCCCATATAAGT60.0
60.0CfActin11 F: TTCCCTGCTTCACTCGATCT
R: TTCCCGGATAACTGAGCATC60.0
60.0CfGADPH F: GGAATCACAGCCGAAGATGT
R: ACCTTGACCATGTCGTCTCC60.1
60.0CfEF-1 F: AGTTTACGCCGCTGTTTTCG
R: CAAGGTCGTCGTCATCATCG60.0
60.0CfPP2A F: GAGCAGTTGCACAGAATGGA
R: CCTGGCGAATTAGAAATGGA60.0
60.0CfMADH F: AGCTTCCATTCTTTGCCTCA
R: TCCGAACAAAAGCAATACCC60.0
59.9CbuActin[17] F: GATGATGCTCCAAGAGCTGT
R: TCCATATCATCCCAGTTGCT60.0
60.0CfGES F: GAGCTACCAAGGCACCTGAG
R: CTCCTTCCACCACCTGGATT59.5
60.0 下载: 导出CSV
下载: 导出CSV
表 2 GeNorm软件分析下内参基因的表达稳定性
Table 2. Analysis of the expression stability of reference genes in leaves by GeNorm
基因名称
Gene name平均表达稳定值
M稳定性排行
Stability rankCfMADH 0.429 1 CfEF-1 0.442 2 CfGADPH 0.467 3 CfPP2A 0.602 4 CfActin11 0.610 5 CfUBC 0.666 6 CbuActin 0.790 7
下载: 导出CSV
表 3 NormFinder软件分析‘麦缘锦楸’及灰楸不同叶色部位内参基因的表达稳定性
Table 3. Analysis of the expression stability of reference genes by NormFinder software
基因名称
Gene name稳定指数
Stability value稳定性排行
Stability rankCfMADH 0.062 1 CfGADPH 0.068 2 CfEF-1 0.095 3 CfActin11 0.329 4 CfPP2A 0.340 5 CfUBC 0.406 6 CbuActin 0.520 7
下载: 导出CSV
表 4 BestKeeper软件分析灰楸及‘麦缘锦楸’不同叶色部位内参基因的表达稳定性
Table 4. BestKeeper software was used to analyze the expression stability of internal reference genes in different leaf color sectors of Maiyuanjinqiu and C.fargesii.
基因名称
Gene name几何平均值
Geometric mean均值
Mean最小值
Min最大值
Max标准差
SD变异系数
CV稳定性排行
Stability rankCfActin11 26.46 26.46 26.08 26.82 0.21 0.80 1 CfMADH 24.69 24.69 24.33 25.01 0.27 1.10 2 CfGADPH 20.91 20.91 20.64 21.49 0.29 1.38 3 CfEF-1 23.81 23.81 23.36 24.22 0.33 1.38 4 CbuActin 22.54 22.54 22.06 22.97 0.37 1.66 5 CfPP2A 28.54 28.55 27.78 29.06 0.46 1.62 6 CfUBC 26.31 26.32 25.48 27.19 0.62 2.35 7
下载: 导出CSV
-
[1] Lopez-Nieves S, Yang Y, Timoneda A, et al. Relaxation of tyrosine pathway regulation underlies the evolution of betalain pigmentation in Caryophyllales[J]. New Phytologist, 2018, 217(2): 896-908. doi: 10.1111/nph.14822 [2] Jaiswal P S, Kaur N, Randhawa G S. Identification of reference genes for qRT-PCR gene expression studies during seed development and under abiotic stresses in Cyamopsis tetragonoloba[J]. Crop Science, 2019, 59(1): 252-265. doi: 10.2135/cropsci2018.05.0313 [3] Jia Y, Liu S C, Zhao J, et al. Reference gene selection and validation by qRT-PCR during flower development and in different organs of Primula forbesii[J]. The Journal of Horticultural Science and Biotechnology, 2020, 95(3): 383-394. doi: 10.1080/14620316.2019.1681909 [4] Li C, Xu J, Deng Y, et al. Selection of reference genes for normalization of cranberry (Vaccinium macrocarpon Ait. ) gene expression under different experimental conditions[J]. PLoS ONE, 2019, 14(11): e0224798. doi: 10.1371/journal.pone.0224798 [5] Xiao F, Zheng Y F, Chen J L, et al. Selection and validation of reference genes in all-red Amaranth (Amaranthus tricolor L. ) seedlings under different culture conditions[J]. The Journal of Horticultural Science and Biotechnology, 2021, 96(5): 1-10. [6] Chen G S, Li J T, Liu Y, et al. Selection and validation of reference genes for quantitative RT-PCR analysis in Castanea mollissima[J]. Plant Physiology Journal, 2019, 55(3): 158-166. [7] Yang C L, Yuan X Y, Zhang J, et al. Comprehensive transcriptome analysis of reference genes for fruit development of Euscaphis konishii[J]. PeerJ, 2020, 8: e8474. doi: 10.7717/peerj.8474 [8] Hu X W, Zhang L J, Nan S Z, et al. Selection and validation of reference genes for quantitative real-time PCR in Artemisia sphaerocephala based on transcriptome sequence data[J]. Gene, 2018, 657: 39-49. doi: 10.1016/j.gene.2018.03.004 [9] 齐香玉, 陈双双, 冯 景, 等. 茉莉花实时荧光定量PCR内参基因的筛选与验证[J]. 华北农学报, 2020, 35(6):22-30. doi: 10.7668/hbnxb.20191401 [10] Li L, Li N, Fang H, et al. Selection and validation of reference genes for normalisation of gene expression in Glehnia littoralis[J]. Scientific Reports, 2020, 10(1): 7374. doi: 10.1038/s41598-020-63917-5 [11] Tang F, Chu L, Shu W, et al. Selection and validation of reference genes for quantitative expression analysis of miRNAs and mRNAs in Poplar[J]. Plant Methods, 2019, 15(1): 35. doi: 10.1186/s13007-019-0420-1 [12] 苏晓娟, 樊保国, 袁丽钗, 等. 实时荧光定量 PCR分析中毛果杨内参基因的筛选和验证[J]. 植物学报, 2013, 48(5):507-518. [13] 储文渊, 王玉娇, 朱东悦, 等. 盐和干旱胁迫下杨树新内参基因的筛选[J]. 林业科学, 2017, 53(10):70-79. doi: 10.11707/j.1001-7488.20171008 [14] Li T T, Yuan W G, Qiu S, et al. Selection of reference genes for gene expression analysis in Liriodendron hybrids' somatic embryogenesis and germinative tissues[J]. Scientific Reports, 2021, 11(1): 4957-4957. doi: 10.1038/s41598-021-84518-w [15] Wang N, Zhu T Q, Lu N, et al. Quantitative phosphoproteomic and physiological analyses provide insights into the formation of the variegated leaf in Catalpa fargesii[J]. International Journal of Molecular Sciences, 2019, 20(8): 1895. doi: 10.3390/ijms20081895 [16] Qi C H, Jang H, Zhao X Y, et al. The characterization, authentication, and gene expression pattern of the MdCER Family in Malus domestica[J]. Horticultural Plant Journal, 2019, 5(1): 1-9. doi: 10.1016/j.hpj.2018.11.003 [17] Jing D L, Xia Y, Chen F, et al. Ectopic expression of a Catalpa bungei (Bignoniaceae) PISTILLATA homologue rescues the petal and stamen identities in Arabidopsis pi-1 mutant[J]. Plant Science, 2015, 231: 40-51. doi: 10.1016/j.plantsci.2014.11.004 [18] Vandesompele J, Preter K D, Pattyn F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes[J]. Genome Biology, 2002, 3(7): 00341. [19] Pfaffl M W, Tichopad A, Prgomet C, et al. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-Based tool using pair-wise correlations[J]. Biotechnology Letters, 2004, 26(6): 509-515. doi: 10.1023/B:BILE.0000019559.84305.47 [20] Dong Z, Chen P, Zhang N, et al. Evaluation of reference genes for quantitative real-time PCR analysis of gene expression in Hainan medaka (Oryzias curvinotus)[J]. Gene Reports, 2019, 14: 94-99. doi: 10.1016/j.genrep.2018.11.008 [21] Maldonado-Taipe N, Patirange DSR, Schmöckel SM, et al. Validation of suitable genes for normalization of diurnal gene expression studies in Chenopodium quinoa[J]. PLoS ONE, 2021, 16(3): e0233821. doi: 10.1371/journal.pone.0233821 [22] Cao S H, Hao P P, Shu W S, et al. Phylogenetic and expression analyses of with-no-lysine kinase genes reveal novel gene family diversity in fruit trees[J]. Horticultural Plant Journal, 2019, 5(2): 47-58. doi: 10.1016/j.hpj.2019.01.006 [23] Knopkiewicz M, Wojtaszek P. Validation of reference genes for gene expression analysis using quantitative polymerase chain reaction in pea lines (Pisum sativum) with different lodging susceptibility[J]. Annals of Applied Biology, 2019, 174(1): 86-91. doi: 10.1111/aab.12475 [24] Chen H, Hu B, Zhao L, et al. Differential expression analysis of reference genes in Pineapple (Ananas comosus L. ) during reproductive development and response to abiotic stress, Hormonal Stimuli[J]. Tropical Plant Biology, 2019, 12(2): 67-77. doi: 10.1007/s12042-019-09218-2 [25] 吕运舟, 董筱昀, 黄利斌. 黄山栾树实时荧光定量PCR内参基因的筛选[J]. 分子植物育种印刷版, 2019, 17(2):553-560. [26] 苏西娅, 石元豹, 杨晓明, 等. 银杏实时荧光定量PCR分析中内参基因的选择与验证[J]. 植物生理学报, 2019, 55(6):875-882. [27] 许丹芸, 张辉菊, 刘基柱, 等. 肉桂和大叶清化桂内参基因的筛选和验证[J]. 中国实验方剂学杂志, 2021, 27(4):137-144. [28] Chen M D, Wang B, Li Y P, et al. Reference gene selection for qRT-PCR analyses of luffa (Luffa cylindrica) plants under abiotic stress conditions[J]. Scientific Reports, 2021, 11(1): 3161. doi: 10.1038/s41598-021-81524-w [29] Yu Y T, Zhang G, Chen Y K, et al. Selection of Reference Genes for qPCR Analyses of Gene gene expression in ramie leaves and roots across eleven abiotic/biotic treatments[J]. Scientific Reports, 2019, 9(1): 20004. doi: 10.1038/s41598-019-56640-3 [30] Zhang K K, Fan W, Chen D F, et al. Selection and validation of reference genes for quantitative gene expression normalization in Taxus spp[J]. Scientific Reports, 2020, 10(1): 22205. doi: 10.1038/s41598-020-79213-1 [31] Qu R, Miao Y, Cui Y, et al. Selection of reference genes for the quantitative real-time PCR normalization of gene expression in Isatis indigotica fortune[J]. Physiology and Molecular Biology of Plants, 2019, 20(1): 9. [32] Tajti J, Pál M, Janda T. Validation of Reference genes for studying different abiotic stresses in Oat (Avena sativa L. ) by RT-qPCR[J]. Plants, 2021, 10: 1272. doi: 10.3390/plants10071272 [33] Yang Z, Zhang R, Zhou Z. Identification and validation of reference genes for gene expression analysis in Schima superba[J]. Genes, 2021, 12(5): 732. doi: 10.3390/genes12050732 [34] Linardić M, Braybrook SA. Identification and selection of optimal reference genes for qPCR-based gene expression analysis in Fucus distichus under various abiotic stresses[J]. PLoS ONE, 2021, 16(4): e0233249. doi: 10.1371/journal.pone.0233249 [35] Lv Y, Li Y, Liu X, et al. Identification of Ginger (Zingiber officinale Roscoe) reference genes for gene expression analysis[J]. Frontiers in Genetics, 2020, 11: 586098. doi: 10.3389/fgene.2020.586098 [36] Chen M L, Wang Q, Li Y, et al. Candidate reference genes for quantitative gene expression analysis in Lagerstroemia indica[J]. Molecular Biology Reports, 2021, 48: 1677-1685. doi: 10.1007/s11033-021-06209-z [37] 陈凌艳, 谢德金, 荣俊冬, 等. 花叶唐竹4种叶色表型qRT-PCR内参基因筛选[J]. 分子植物育种, 2019(14):4592-4599. [38] Yu Z C, Zhang P, Lin W, et al. Sequencing of anthocyanin synthesis-related enzyme genes and screening of reference genes in leaves of four dominant subtropical forest tree species[J]. Gene, 2019, 716(C): 144024. [39] 王倩颖, 常鹏杰, 申亚梅, 等. 景宁木兰热胁迫下实时荧光定量PCR内参基因的筛选[J]. 浙江农林大学学报, 2019, 36(5):935-942. [40] 陈国松, 李靖同, 刘 阳, 等. 板栗实时定量PCR内参基因的筛选与验证[J]. 植物生理学报, 2019, 55(3):378-386. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5077
- HTML全文浏览量: 3034
- PDF下载量: 75
- 被引次数: 0
