

-
硒是人体所必需的微量营养元素,随着食物链进入人体,以两种形式影响人体的健康。其一,参与蛋白的生物合成。在人体中,几种氨基酸含有硒元素,如硒代半胱氨酸和硒代甲硫氨酸[1]。含有这样氨基酸的蛋白被称为硒蛋白。在人的基因组中,有25个基因编码硒蛋白[1],如谷胱甘肽过氧化物酶(GPX)中的GPX1、GPX2、GPX3、GPX4和GPX6,氧还原蛋白还原酶(TXNRD)中的TXNRD1、TXRND2和TXRND3,碘化甲腺氨酸脱碘酶(DIO)中的DIO1、DIO2和DIO3[2]。其二,硒元素以配位体的形式参与蛋白的活动。例如,硒作为配位体参与调节硒结合蛋白SBP1的活动[3]。这些蛋白在人体中具有多种重要的生理活动,如上述的GPXs可以作为抗氧化酶起作用,TXNRDs则在氧化还原信号转导过程中起作用,DIOs参与甲状腺激素的生物合成[1]。人体对硒的摄入量将影响上述蛋白的生物合成及其活动。如果日常饮食中硒摄入量过多,则引起硒中毒。然而,日常饮食中硒摄入量偏少,则引起人体各种疾病。据统计,高达80%的人群经历过饮食性硒缺乏[1]。硒摄入量不足引起的疾病中,最著名的例子就是克山病和卡斯钦-贝克病。克山病的发生与人体GPX1活动低下有关[4],硒元素的补充可以阻止克山病的发生[5]。在卡斯钦-贝克病发生过程中,硒则与抗氧化和甲状腺功能的维持有关[6]。此外,硒的摄入量不足,还会引起其它疾病,如心脏病[7-8]、癌症[9-10]、腮腺疾病[11-12]等。因此,硒的足量摄入可以消除这些疾病,还可以用于治疗南美洲锥虫病[13-14]。世界上许多国家,针对本国人群的身体状况以及当地的水土中硒的含量,制定了各自的硒摄入标准。
在日常生活中,人们可以通过饮食强化的方式来解决硒摄入不足的问题。鉴于硒的毒性,这种强化方式以生物强化为宜[15]。植物的硒生物强化可以通过多种方式进行,如通过对植物有益的微生物的作用[16-17]和叶面施用亚硒酸钠、硒酸钠、硒酸钾[18]。目前,人们已通过硒生物强化的方式来栽培一些农作物、水果、蔬菜以及食用与药用菌类,如水稻[19]、大豆[20]、小麦[21-22]、苹果[23]、猴头菇[24]、灵芝[25]、羽衣甘蓝[26]、薄叶二行芥、羊莴苣以及生菜和菠菜[27]。坚果是人们日常食用的休闲果品,也可以用于烘焙食品的生产,目前关于坚果类产品的硒生物强化研究报道较少,仅见巴西坚果[28]。
近些年来,香榧(Torreya grandis Fort. ex Lindl.)果实作为休闲食品在我国越来越受到人们的青睐,对新品种选育及其产量研究尤受重视[29-31],但对其果实的硒生物强化研究很少。作者曾在施用硒肥条件下使用植物根内生真菌印度梨形孢与香榧共生以促进其硒的累积[32]。本研究对香榧幼苗叶片施加叶面肥亚硒酸钠,然后对其代谢组进行了分析,以期了解硒叶面肥对香榧叶片代谢的影响,探讨硒叶面肥的吸收转化以及有益于人体健康的次生代谢物的累积。
-
60株香榧实生幼苗从四川省雅安市业之欣种植专业合作社购买,苗龄2年。将每株香榧幼苗定植于1个塑料花盆(高30 cm,直径25 cm)中。每盆装入等量的园艺培养基质(山东莘县一亩田农业科技有限公司生产)。将花盆连带香榧幼苗移入人工智能温室。根据培养基质的湿度进行水分的日常管理。栽培1年后,将60株幼苗随机分成2组,每组30株,对其中一组的叶片喷洒亚硒酸钠(浓度为100 μg·L−1),另一组作为对照,喷洒去离子水,均以叶面全部喷湿为度。15 d后再重复1次。第二次喷施叶面肥后15 d时采样。本研究只取幼苗中上部枝条上叶龄为1 年的叶片为非靶向代谢组分析的实验材料。采样时,因考虑到每株所采的1 年叶片数量不足,因而随机采取2棵幼苗上的1 年叶片合并为1个样品。处理组和对照组各6个样品,共计12个样品。取样后迅速以无菌水进行4次清洗,然后以液氮进行处理,保存于−80 ℃冰箱以待分析。
-
首先取100 mg液氮研磨的组织样本,置于EP管中,加入500 μL的80%甲醇水溶液;然后进行涡旋震荡,冰浴静置5 min,离心20 min (15 000 g,4 ℃)。取一定量的上清加质谱级水稀释至甲醇含量为53%,离心20 min (15 000 g,4 ℃),收集上清,进样LC-MS 进行分析[33]。所有样品均采用正负离子化模式分析。
色谱仪采用Thermo Fisher (型号:Vanquish UHPLC,德国)。液相色谱分析的条件如下:采用Hypersil Gold column (C18)色谱柱(100 mm × 2.1 mm,1.9 μm),其柱温:40 ℃;流速:0.2 mL·min−1。正离子化模式下,流动相A:0.1%甲酸;流动相B:甲醇。负离子化模式下,流动相A:5 mmol·L−1醋酸铵,pH 9.0;流动相B:甲醇。液相色谱梯度洗脱程序如表1所示:
表 1 色谱梯度洗脱程序
Table 1. Process of chromatographic gradient elution
时间 Time/min A/% B/% 0.0 98 2 1.5 98 2 3.0 15 85 10.0 0 100 10.1 98 2 11.0 98 2 12.0 98 2 质谱仪采用Fermo Fisher (型号:Q ExactiveTM HF,德国)。质谱条件如下:扫描范围选择m/z 100~1 500;ESI源的设置如下:喷雾电压:3.5 kV;鞘气流速:35 psi;辅助气流速: 10 L·min−1;离子传输管温度:320 ℃;离子导入射频电平:60;辅助气加热器温度:350 ℃;极性:正负离子化模式;MS/MS二级扫描为数据依赖性扫描。氮气(纯度 ≥ 99.999%)用作碰撞气以便产生碎片离子,其能量水平设置为正常碰撞能量的30%。
-
首先将质谱检测得到的原始文件(.raw)导入Compound Discoverer 3.1(以下简称CD3.1) 搜库软件中进行处理,对每个代谢物进行保留时间、质荷比(m/z)等参数的简单筛选,然后设置保留时间偏差0.2 min和质量偏差5 mg·kg−1对不同样品进行峰对齐,使鉴定更准确,随后设置质量偏差5 mg·kg−1 、信号强度偏差30%、信噪比3、最小信号强度、加合离子等信息进行峰提取,同时对峰面积进行定量,再整合目标离子,然后通过分子离子峰和碎片离子进行分子式的预测,并与mzCloud (
https://www.mzcloud.org/ )、mzVault和Masslist 数据库进行比对,用blank样本去除背景离子,将原始定量结果依据公式:样本原始定量值/(样本代谢物定量值总和/QC1样本代谢物定量值总和),进行标准化处理,得到相对峰面积;并将质量控制(QC)样本中相对峰面积的变异系数(CV)大于30%的化合物删除,最后得到代谢物的鉴定和相对定量结果。数据处理部分基于Linux操作系统(CentOS版本6.6)以及R语言软件包和Python软件包进行,其中代谢物定性和定量分析采用Python 软件(Python-3.5.0);代谢物分类、QC样本控制、总样品PCA分析均采用R语言(R-3.4.3)。 -
使用KEGG数据库(
https://www.genome.jp/kegg/pathway.html )、HMDB数据库(https://hmdb.ca/metabolites )和LIPIDMaps数据库(http://www.lipidmaps.org/ )对鉴定到的代谢物进行注释。多元统计分析部分,使用代谢组学数据处理软件metaX[6]对数据进行转换后进行主成分分析(PCA)和偏最小二乘法判别分析(PLS-DA),进而得到每个代谢物的VIP值。单变量分析部分,基于t检验来计算各代谢物在两组间统计学显著性(P值),并计算代谢物在两组间的差异倍数(fold change,FC)。差异代谢物筛选的默认标准为变量投影重要度(Variable Importance in the Projection,VIP) > 1,P值< 0.05 且FC ≥ 2 或FC ≤ 0.5。火山图用R软件包ggplot2绘制,可以综合代谢物的VIP值、log2(FoldChange)和−log10(p值)3个参数,来筛选感兴趣的代谢物。聚类热图,用R软件包Pheatmap 进行绘制,使用z-score 对代谢物数据进行归一化。差异代谢物之间的相关性分析(Pearson相关系数)使用R语言中的cor()进行,统计显著性通过R语言中cor.mtest()实现,P值<0.05 为在统计学上显著,并用R语言中的corrplot软件包绘制相关性图。KEGG途径气泡图用R语言中的软件包ggplot2进行绘制。使用KEGG数据库来研究代谢物的功能和代谢途径,当x/n > y/N时,认为该代谢途径富集(其中,x为与该通路相关的差异代谢物的数目;y为与该通路相关的背景(所有)代谢物的数目; n为KEGG注释的差异代谢物数目; N为KEGG注释的背景(所有)代谢物的数目);当代谢途径的P值< 0.05时,认为该代谢途径是显著富集的。代谢物差异分析和KEGG富集分析均采用Python-3.5.0和R-4.0.3进行。 -
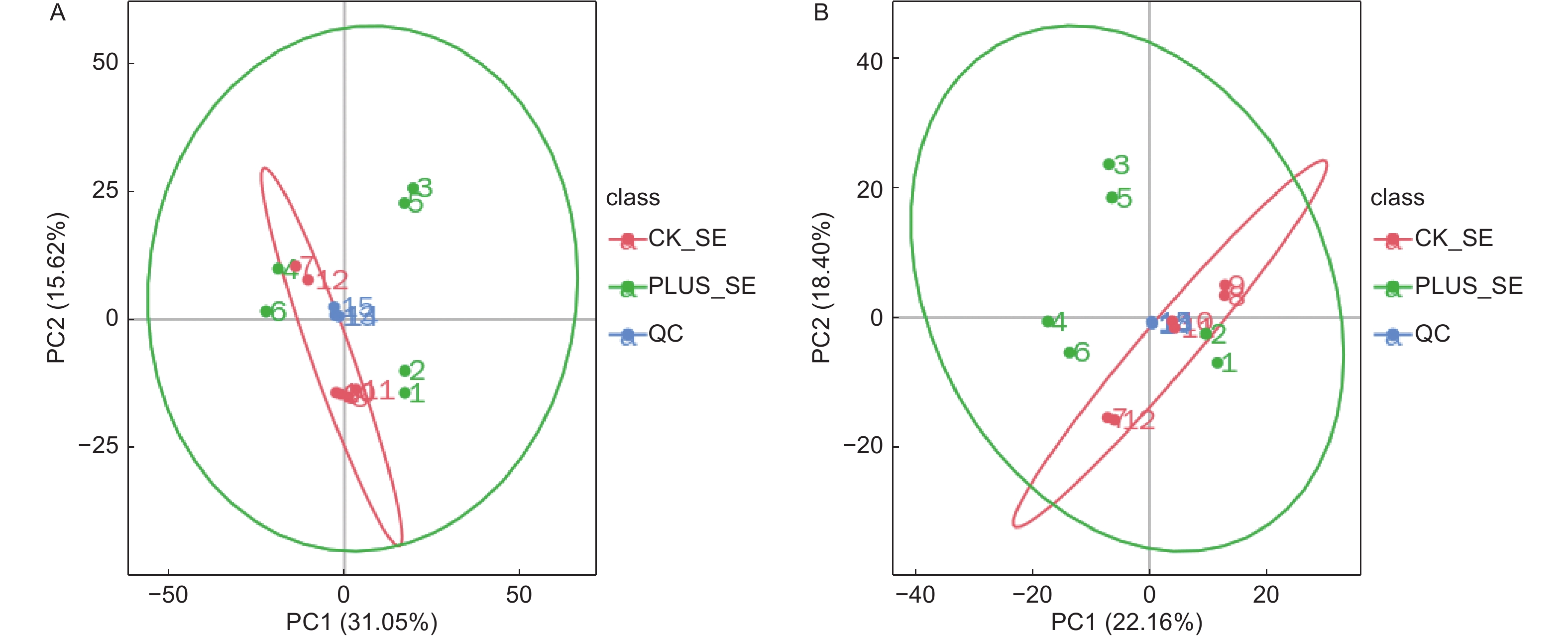
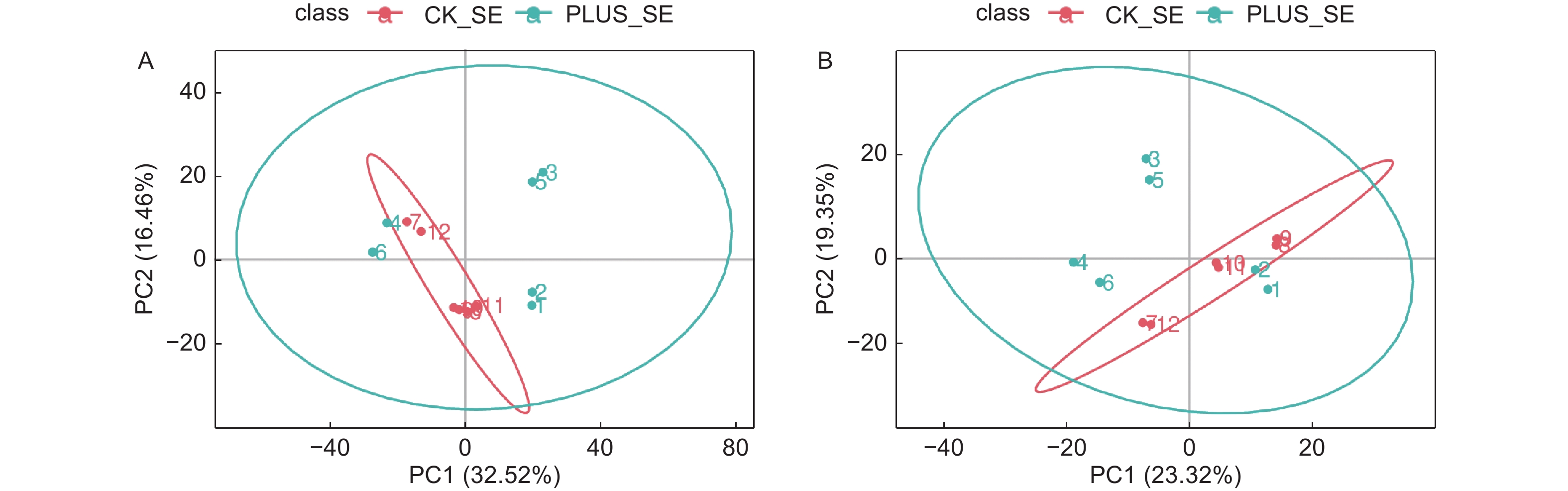
对从6个硒处理样本和6个对照样本获得的数据,采用PCA方法进行归类分析,从总体上反映各组样本之间的总体代谢差异和组内样本之间的变异大小。从图1可以看出,在正负离子化模式下,6个硒处理样本和6个对照样本都处于95%的置信区间范围内,且QC样本非常集中,表明实验数据质量高,可以满足后续的分析。在差异代谢物的总体分布趋势上,无论是在第一主成分方向上还是在第二主成分方向上,正负离子化模式下,硒处理和对照样本的代谢物差异都不大,表明硒处理只引起香榧叶片内少数代谢物发生差异变化(图2)。
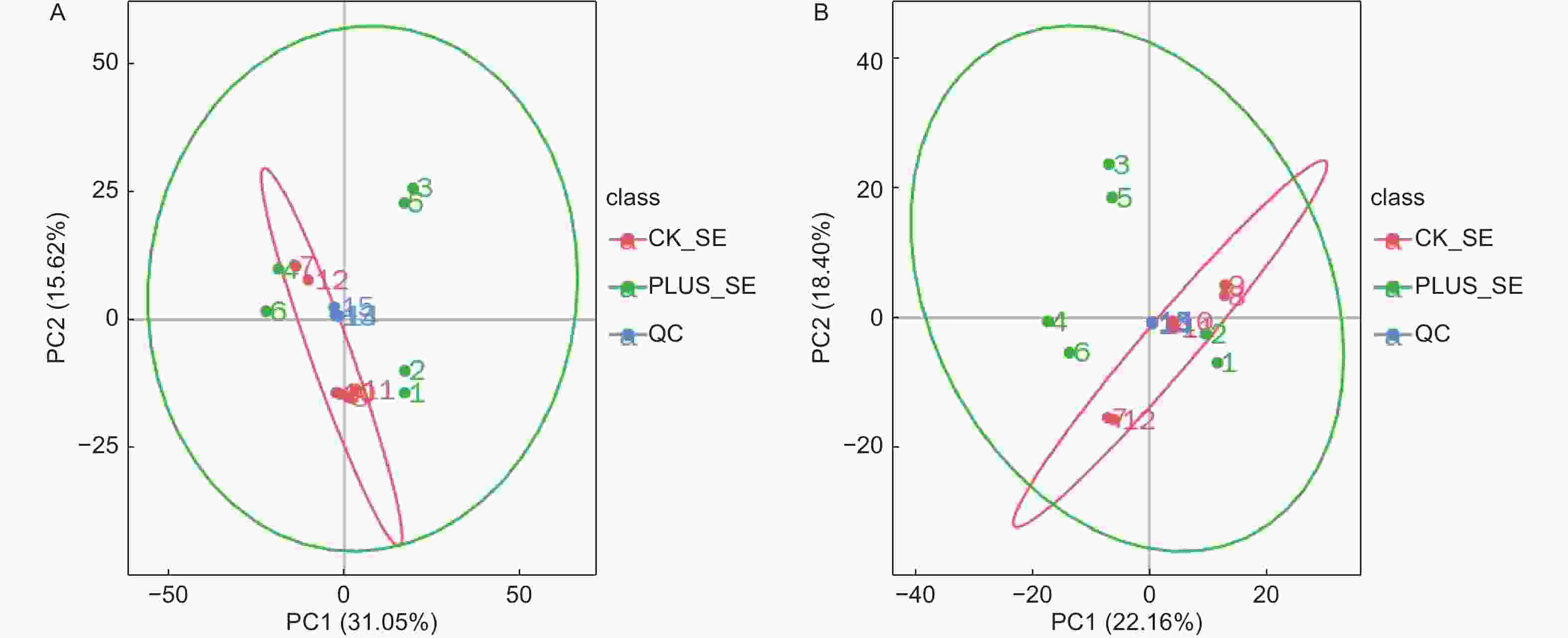
图 1 总样本PCA分析。A-正离子模式;B-负离子模式。 CK_SE为对照,PLUS_SE为硒处理。QC为质控
Figure 1. PCA analysis of total samples. A-Positive ionization mode; B- negative ionization mode. CK_SE stands for control, PLUS_SE for selenium treatment, QC for quality control
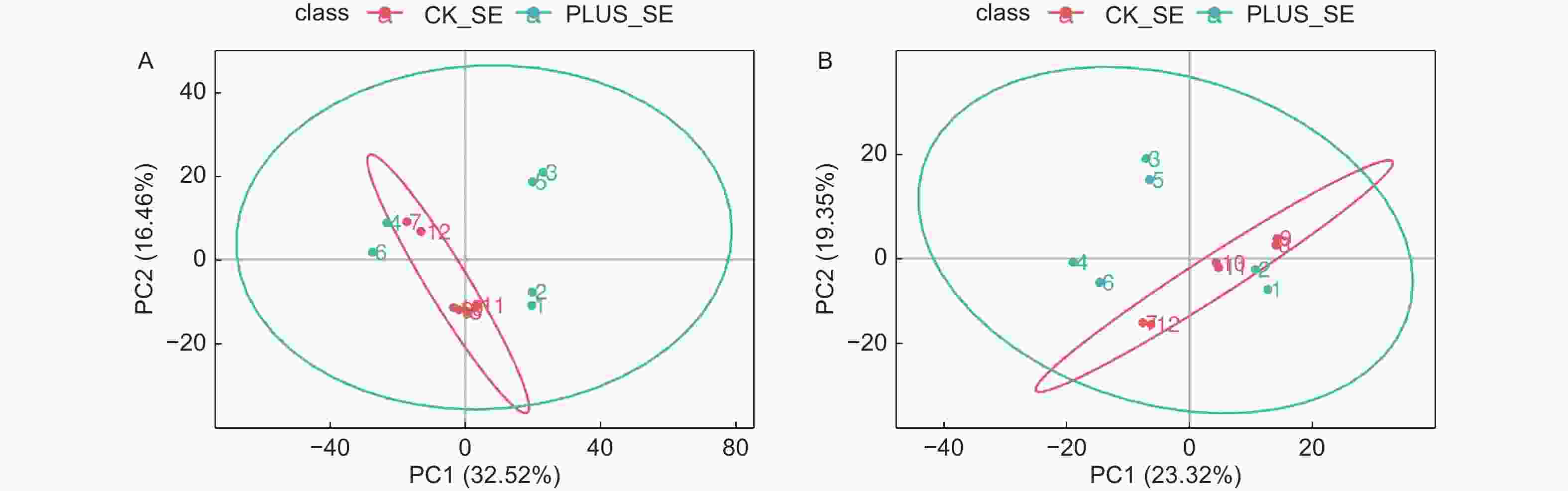
图 2 差异代谢物筛选的PCA分析。A-正离子模式;B-负离子模式。 CK_SE为对照,PLUS_SE为硒处理。QC为质控
Figure 2. PCA used for selection of differential metabolites. A-positive ionization mode; B- negative ionization mode. CK_SE stands for control, PLUS_SE for selenium treatment, QC for quality control
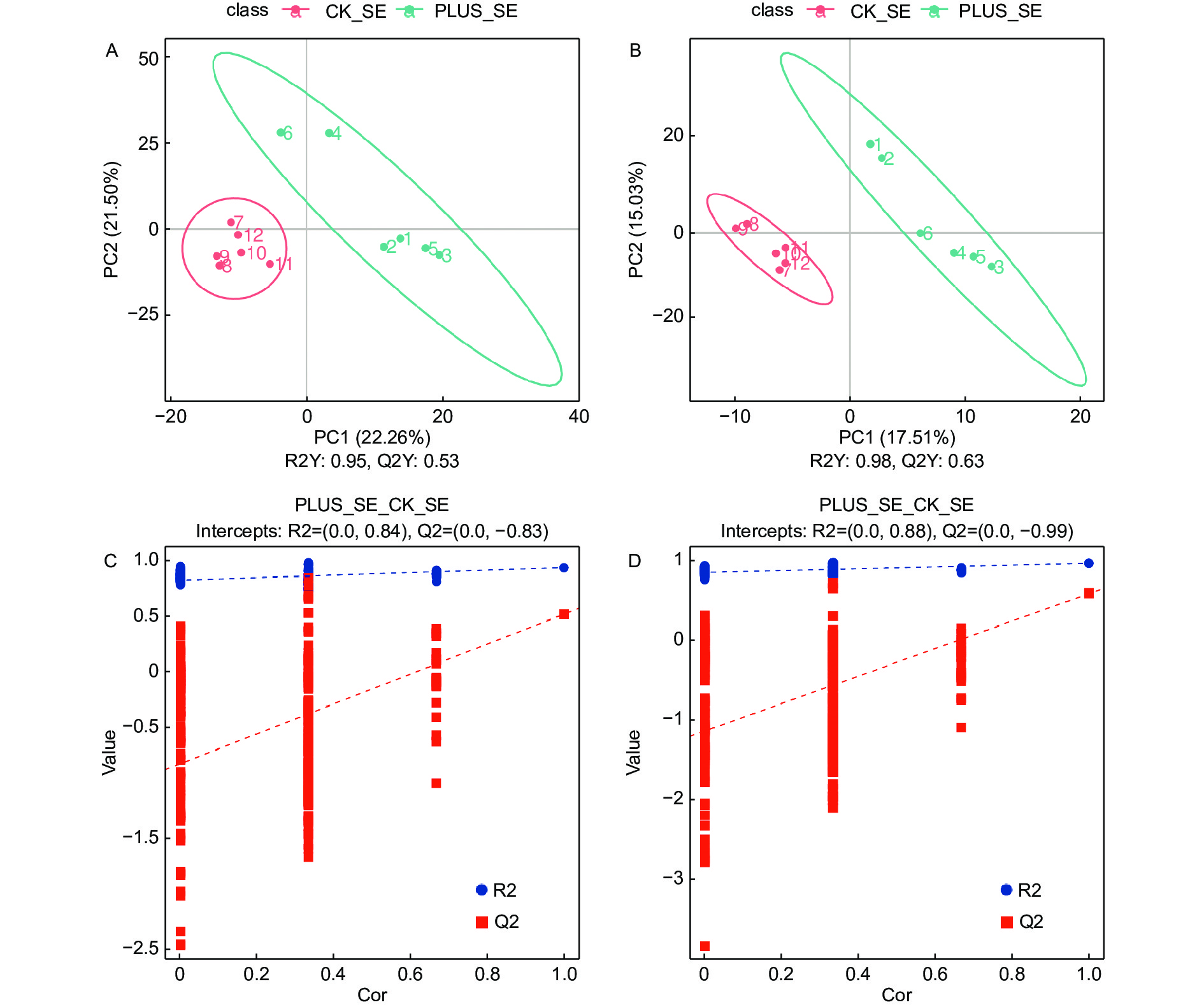
为了确定硒处理和对照之间的代谢物差异,采用有监督的PLS-DA模型放大硒处理和对照之间的差异,并建立代谢物表达量与延平类别之间的关系模型。交叉验证结果显示,在正离子化模式下,R2Y(模型对分类变量Y的可解释性)和Q2Y(模型的可预测性)分别为0.95和0.53(图3A);在负离子化模式下,R2Y和Q2Y分别为0.98和0.63(图3B)。在正负离子化模式下,其R2Y和Q2Y均大于0.5,且R2Y均大于Q2Y(图3A, B)。这些结果表明拟合模型的准确度和预测能力良好。从PLS-DA得分图可以看出,在正负离子化模式下,硒处理样本和对照具有非常显著的差别,且全部处于95%的置信区间内(图3A, B)。通过置换检验对模型有效性进行检验,其结果表明原模型具有良好的稳定性,不存在过拟合现象,因为在正负离子化模式下,R2均大于Q2,且Q2回归线在Y轴上的截距小于0(图3C, D)。
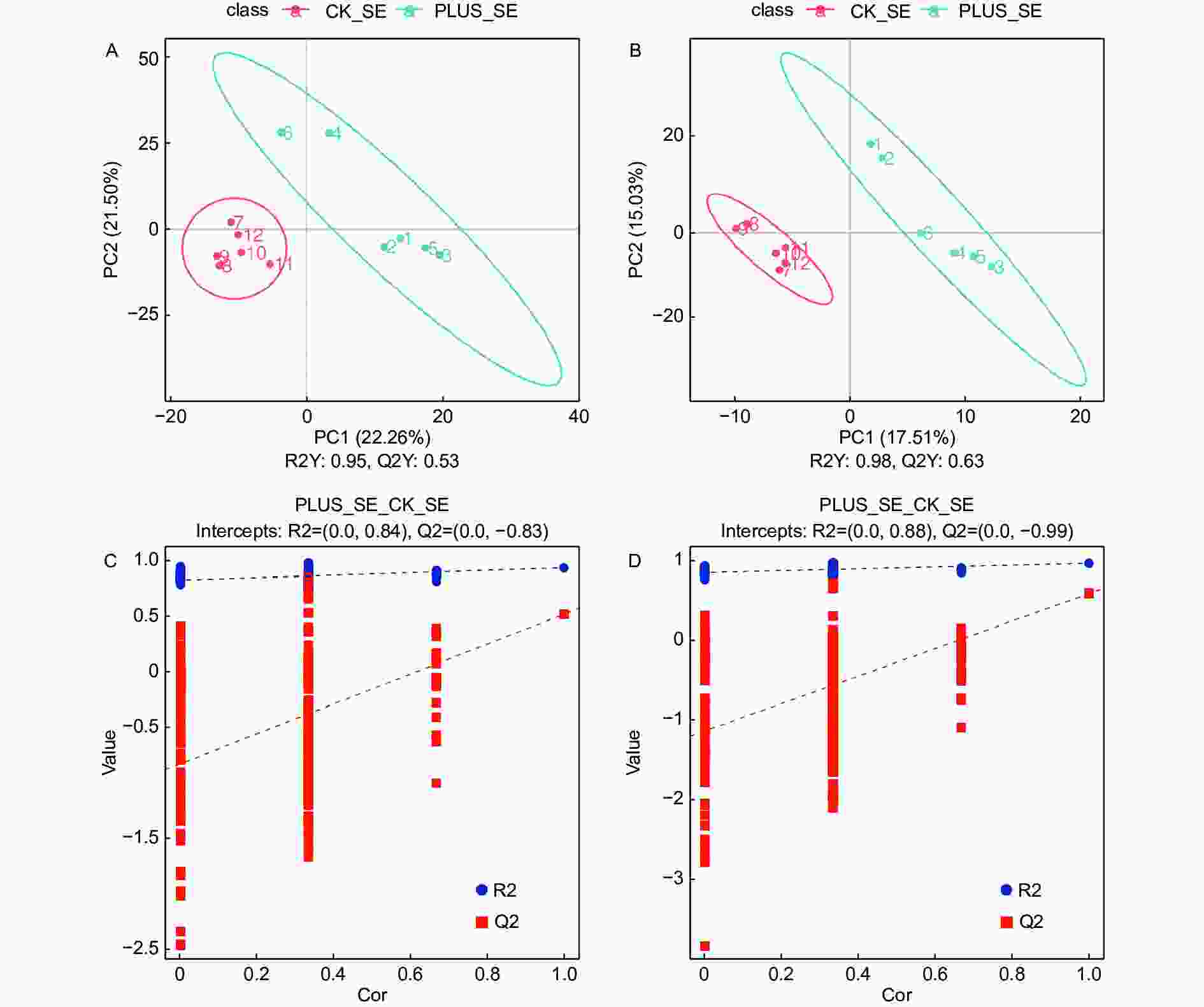
图 3 代谢物筛选的PLS-DA分析
Figure 3. PLS-DA used for selection of differential metabolites
-
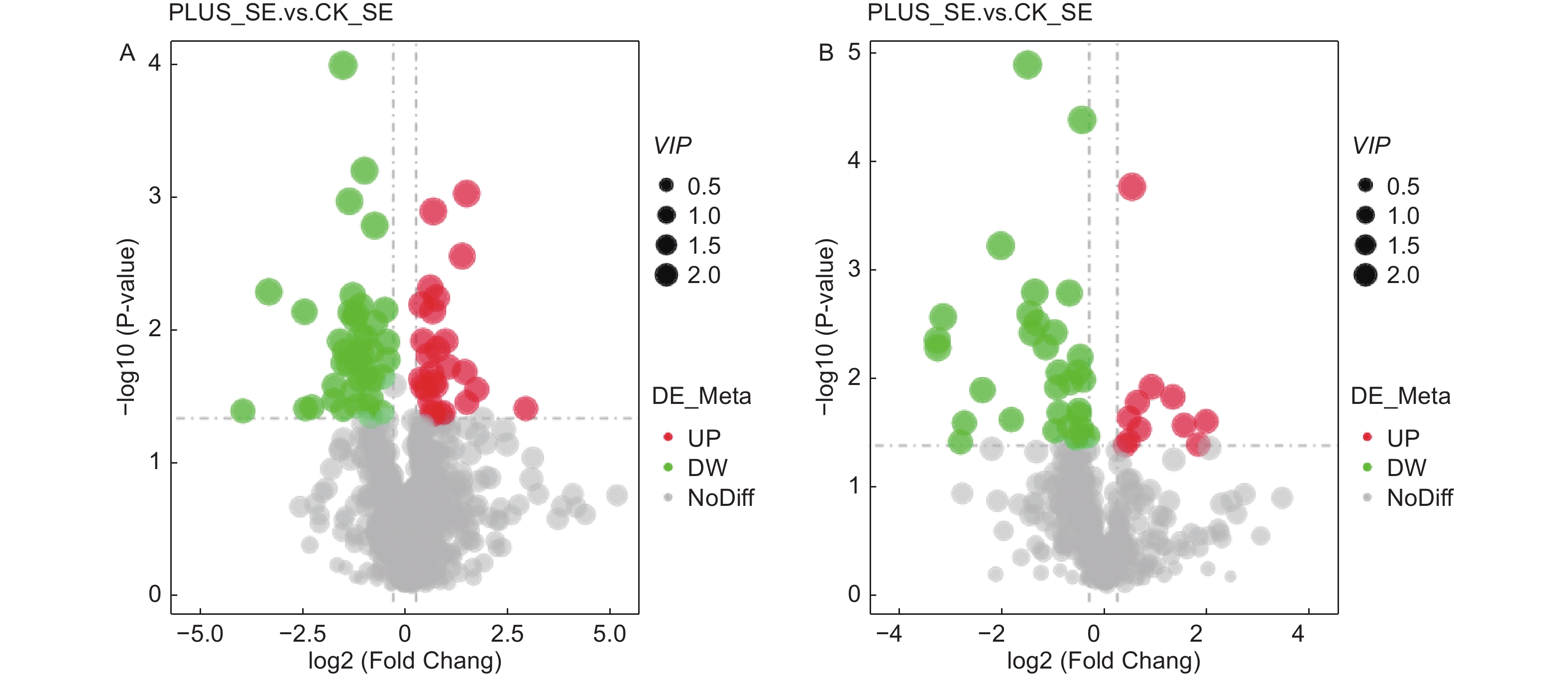
在正离子化模式下,硒处理与对照之间存在909种差异代谢物,其中69种差异显著(27种显著上调,42种显著下调);在负离子化模式下,硒处理与对照之间存在433种差异代谢物,其中43种差异显著(11种显著上调,32种显著下调)(表2)。其火山图显示,在正负离子化模式下,无论是显著上调的代谢物还是显著下调的代谢物,其VIP大致一致(图4)。在正离子化模式下,显著上调的代谢物前3种为N'-[6-(tert-butyl)thieno[3,2-d]pyrimidin-4-yl]-4-methylbenzohydrazide、4-dihydroxy-1,4-dimethyl-7-(propan-2-ylidene)-decahydroazulen-6-one和butanedioicacid,其上调倍数分别为7.009、3.179和2.718;在负离子化模式下,显著上调的代谢物前3种为polyphyllin V、C-pentosyl-apigenin O-p-coumaroylhexoside和plantagoside,其上调倍数分别为3.761、3.399和2.827(表3)。
表 2 差异代谢物筛选结果
Table 2. Selective results of differential metabolites
样品比对
Compared samples代谢物总数
Num. of total ident.差异显著的代谢物总数
Num. of total Sig.显著上调代谢物总数
Num. of Sig.up显著下调代谢物总数
Num. of Sig.down正离子化模式
+ Se vs. CK909 69 27 42 负离子化模式
+ Se vs. CK433 43 11 32 注:差异代谢物筛选时,设定阈值为VIP > 1.0,FC > 1.2 Note: Threshold values were set as VIP > 1.0 and FC > 1.2 when differential metabolites were selected. 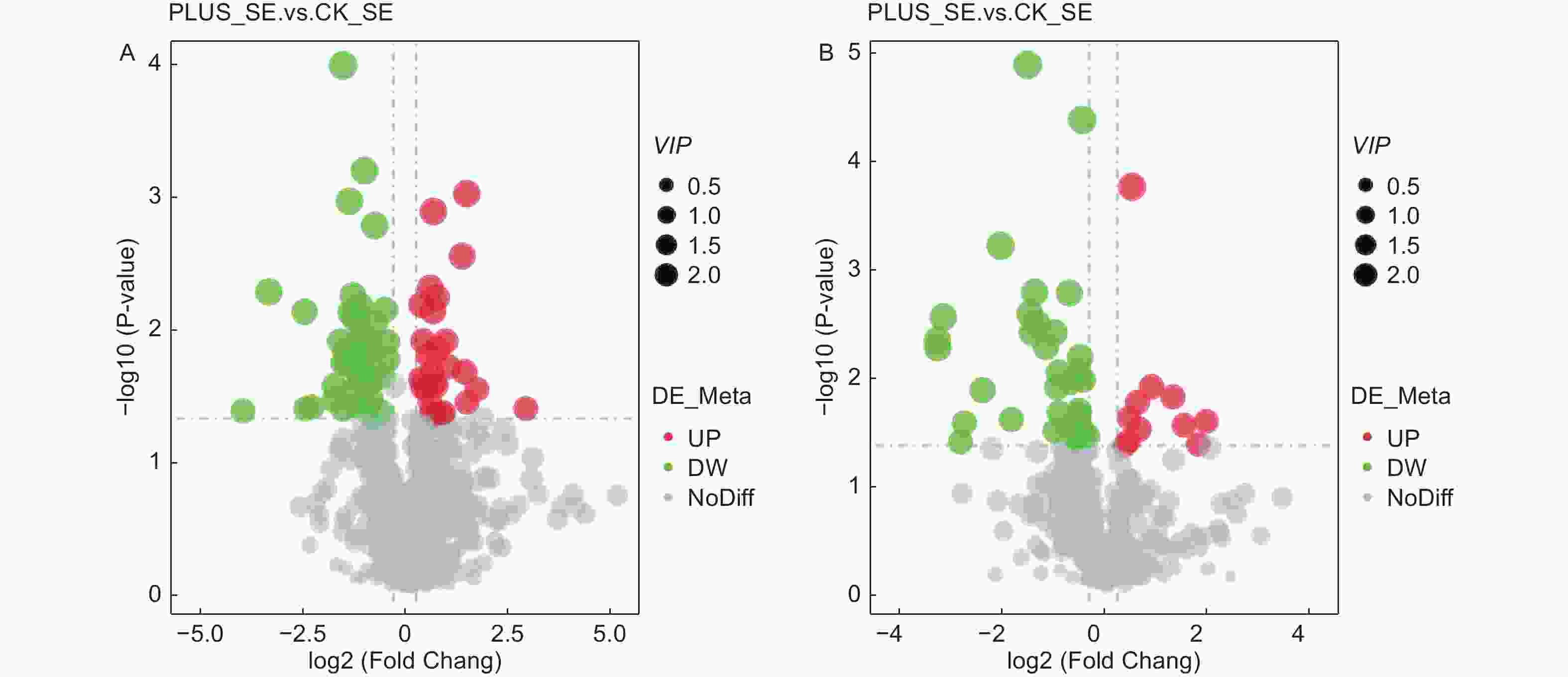
图 4 差异代谢物火山图。 A-正离子化模式;B-负离子化模式
Figure 4. Volcano maps of differential metabolites. A-positive ionization mode; B-negative ionization mode.
表 3 硒肥叶面处理后正负离子化模式下上调最多10种差异代谢物
Table 3. Top 10 differential metabolites significantly up-regulated under positive and negative ionization modes in leaves of Torreya grandis seedlings after selenium foliar application
差异代谢物名称
Names of metabolites分子式
Formula分子量
Molecular weight滞留时间
RT/min质荷比
m/z倍数FC log2FC P value 正离子化模式下 N'-[6-(tert-butyl)thieno[3,2-d]pyrimidin-4-yl]-
4-methylbenzohydrazideC18H20N4OS 362.12 4.44 363.13 7.009 2.809 0.041 8 1,4-dihydroxy-1,4-dimethyl-7-(propan-2-ylidene)-
decahydroazulen-6-oneC15H24O3 234.16 7.041 235.17 3.179 1.669 0.029 7 butanedioic acid C28H36O10 532.23 6.167 533.24 2.718 1.442 0.037 6 HQH C17H24N8O5 442.18 7.086 443.18 2.712 1.439 0.000 9 (2,6-dimethylmorpholino)
(1-methyl-5-nitro-1H-pyrazol-4-yl)methanoneC11H16N4O4 290.10 1.845 291.11 2.626 1.393 0.021 6 川芎内酯ligustilide C12H14O2 190.10 7.114 191.11 2.524 1.336 0.002 7 6-唾液乳糖6-sialyllactose C23H39NO19 633.21 4.907 634.22 2.021 1.015 0.019 8 阿齐沙坦azilsartan C25H20N4O5 438.14 1.307 439.14 1.931 0.949 0.012 4 tricin 4'-O-(beta- guaiacylglyceryl) ether C27H26O11 526.15 5.199 527.15 1.874 0.906 0.045 1 aurantio-obtusin C17H14O7 330.07 5.005 331.08 1.796 0.844 0.046 2 负离子化模式下 polyphyllin V C39H62O12 722.42 8.853 721.42 3.761 1.911 0.029 8 C-pentosyl-apigenin O-p- coumaroylhexoside C35H34O16 710.18 5.152 709.17 3.399 1.765 0.048 9 车前苷plantagoside C21H22O12 466.11 4.792 465.11 2.827 1.499 0.032 2 gracillin C45H72O17 884.48 8.575 883.47 2.448 1.291 0.017 5 2-aminoethylphosphonate C2H8NO3P 125.02 4.824 124.02 1.860 0.895 0.014 1 rauwolscine hydrochloride C21H27ClN2O3 390.17 5.431 389.16 1.586 0.665 0.035 2 phenprobamate C10H13NO2 179.09 5.521 178.09 1.549 0.632 0.019 7 bruceine A C26H34O11 522.21 5.815 521.20 1.448 0.534 0.000 2 1,2,3-cyclopropanetricarboxylic acid C6H6O6 174.02 1.909 173.01 1.394 0.479 0.027 6 2-aminoadipic acid C6H11NO4 161.07 4.771 160.06 1.386 0.471 0.045 3 -
差异代谢物的KEGG通路富集结果如表4所示。在正离子化模式下,有27种KEGG通路显著富集;在负离子化模式下,有11种KEGG通路显著富集。在这些通路中,差异代谢物较多的通路为代谢途径(9种差异代谢物)、嘌呤代谢途径(3种差异代谢物)以及ABC转运蛋白、氨基乙酰-tRNA生物合成通路、氨基酸合成通路、核黄素代谢通路、氨基糖和核苷糖代谢通路(各2种差异代谢物)。
表 4 硒肥叶面处理后最富集的KEGG通路
Table 4. The most enriched KEGG pathways after selenite treatment
MapID* MapTitle P value x y n N 正离子化模式 map00230 purine metabolism 0.047 8 3 9 14 139 map00100 steroid biosynthesis 0.100 7 1 1 14 139 map00290 valine, leucine and isoleucine biosynthesis 0.100 7 1 1 14 139 map00232 caffeine metabolism 0.191 9 1 2 14 139 map00261 monobactam biosynthesis 0.191 9 1 2 14 139 map00630 glyoxylate and dicarboxylate metabolism 0.191 9 1 2 14 139 map00904 diterpenoid biosynthesis 0.191 9 1 2 14 139 map00910 nitrogen metabolism 0.191 9 1 2 14 139 map00053 ascorbate and aldarate metabolism 0.274 5 1 3 14 139 map00780 biotin metabolism 0.274 5 1 3 14 139 map02010 ABC transporters 0.305 6 2 11 14 139 map00970 aminoacyl-tRNA biosynthesis 0.346 0 2 12 14 139 map00220 arginine biosynthesis 0.349 2 1 4 14 139 map00240 pyrimidine metabolism 0.349 2 1 4 14 139 map00250 alanine, aspartate and glutamate metabolism 0.349 2 1 4 14 139 map00310 lysine degradation 0.349 2 1 4 14 139 map00750 vitamin B6 metabolism 0.349 2 1 4 14 139 map00860 porphyrin and chlorophyll metabolism 0.349 2 1 4 14 139 map00260 glycine, serine and threonine metabolism 0.416 6 1 5 14 139 map00902 monoterpenoid biosynthesis 0.416 6 1 5 14 139 map00944 flavone and flavonol biosynthesis 0.416 6 1 5 14 139 map00360 phenylalanine metabolism 0.477 6 1 6 14 139 map00945 stilbenoid, diarylheptanoid and gingerol biosynthesis 0.477 6 1 6 14 139 map00960 tropane, piperidine and pyridine alkaloid biosynthesis 0.532 6 1 7 14 139 map00941 flavonoid biosynthesis 0.582 1 1 8 14 139 map01230 biosynthesis of amino acids 0.648 7 2 15 14 139 map01100 metabolic pathways 0.800 8 9 80 14 139 负离子化模式 map00740 riboflavin metabolism 0.013 6 2 2 9 73 map00520 amino sugar and nucleotide sugar metabolism 0.071 7 2 4 9 73 map00750 vitamin B6 metabolism 0.123 2 1 1 9 73 map00040 pentose and glucuronate interconversions 0.232 8 1 2 9 73 map00440 phosphonate and phosphinate metabolism 0.232 8 1 2 9 73 map00710 carbon fixation in photosynthetic organisms 0.330 1 1 3 9 73 map00030 pentose phosphate pathway 0.416 2 1 4 9 73 map00760 nicotinate and nicotinamide metabolism 0.416 2 1 4 9 73 map02010 ABC transporters 0.492 3 1 5 9 73 map01230 biosynthesis of amino acids 0.559 5 1 6 9 73 map01200 carbon metabolism 1 1 8 9 73 *注:(1) MapID:富集的KEGG Pathway的ID;(2) MapTitle:富集的KEGG Pathway名称;(3) P value:富集分析的P value;(4) x:与该通路相关的差异代谢物的数目;(5) y:与该通路相关的背景(所有)代谢物的数目;(6) n:KEGG注释的差异代谢物数目;(7) N:KEGG注释的背景(所有)代谢物的数目。
Note: (1) MapID: the ID of enriched KEGG pathway; (2) MapTitle: name of the enriched KEGG pathway; (3) p value: p value of enrichment analysis; (4) x: the number of differential metabolites related to KEGG pathway; (5) y: the number of all the secondary metabolites related to KEGG pathway; (6) n: the number of differential secondary metabolites annotated in KEGG; (7) N: the number of all the secondary metabolites annotated in KEGG. -
香榧果实作为国内正在兴起的休闲食品,日益为人们所接收,其消费量日益扩大。香榧果实的硒生物强化,可以适当满足人们对硒的摄入量,进而增强人们的身体健康。硒肥叶面喷施是一个很好的途径,在诸如葡萄[34]、芒果[35]、黄梨[36]等上使用,促进了这些水果的硒的生物强化。然而,这些报道只对硒叶面肥对果树生长、果实品种等进行了研究,没有对硒肥施用对果树叶片和果实的代谢组的影响进行探讨。本研究实验结果表明,香榧叶面施加亚硒酸钠上调了许多次生代谢物(表3)。例如,川芎内酯在叶片中增加2.524倍(与对照相比,表3)。川芎内酯对人体和动物的生理具有重要影响。如在脑缺血再灌注,川芎内酯可以通过激活依赖于PINK1/Parkin的中型噬菌体而减少海马趾神经元的伤害[37];在低氧口腔癌细胞中,通过激活内质网胁迫信号而诱导依赖于c-Myc的细胞凋亡[38]。该次生代谢物还可以通过TLR4通路而促进与癌症相关的纤维组织母细胞的凋亡[39]。在经过硒肥处理后,其叶片6-唾液乳糖含量增加2.021倍(表3)。有研究结果表明,人乳中的6-唾液乳糖通过抑制toll样的受体(TLR4)而抑制正在坏死的小肠结肠炎的发展[40],在血管平滑肌细胞中,它还可以对血管紧张素II引起的繁殖、迁移以及成骨性转换具有抑制作用[41]。在叶面硒肥应用后,阿齐沙坦含量增加1.931倍(表3,与对照相比)。阿齐沙坦是一种血管紧张素受体的阻断剂[42],可用于治疗高血压[43-44]。此外,该次生代谢物还可以用于治疗高血糖引起的血脑障壁的渗透压力过大[45]。在施用硒叶面肥后,香榧叶片中车前子苷含量增加2.827倍(表3,与对照相比)。该次生代谢物首次从车前(Plantago asiatica L.)种子中分离鉴定,为一种新奇的甘露糖苷酶抑制剂,具有抑制免疫反应的作用[46]。该次生代谢物近来用于治疗猴痘[47]。由此可见,对香榧叶片喷施亚硒酸钠叶面肥,可以增加很多对人体有益的次生代谢物。如果这些次生代谢物能在香榧果实中累积,无疑可以进入人体而增强人体的各种机能。因此,进一步的实验需要对这些次生代谢物在香榧果实中累积进行分析。然而,正如前面所述,硒进入生物体后要么以离子形式存在,要么以氨基酸和蛋白质形成存在,其含量可能很低,因此,本研究没有检测到含硒的次生代谢物。
KEGG富集通路分析表明,在亚硒酸钠叶面喷施后,香榧叶片中的代谢途径也发生很大的变化,其中包括黄酮类化合物的生物合成、氨基酸的生物合成以及碳固定途径(表4)。碳固定途径的富集,无疑表明了亚硒酸钠叶面喷施后可增强香榧叶片的光合作用,进而促进光合产物的累积而促进其生长。黄酮类化合物对人体具有非常重要的作用,如充当抗氧化剂[48]、抗癌[49]、消炎[50]。此外,香榧叶片喷施亚硒酸钠增加了其氨基酸的生物合成,然而均没有涉及到含硒氨基酸的合成通路富集(表4),这可能是质谱图库中没有相应的硒氨基酸合成通路。因此,要检测硒肥使用之后植物叶片中含硒氨基酸的生物合成途径的变化,应该在质谱图对比库中增添含硒氨基酸的图谱。
-
本研究结果表明,对香榧进行叶面喷施硒肥可以显著改变其代谢物的生物合成,其中一些代谢物对人体健康非常有益,因此香榧叶面喷施硒肥可以促进其叶片中有益于人体健康的代谢物的累积。
叶面喷施硒肥对香榧幼苗叶片代谢组的影响
Effects of Foliar Spray of Selenite on Metabolome in Leaves of Torreya grandis Seedlings
-
摘要:
目的 分析亚硒酸钠叶面喷施后对香榧(Torreya grandis)幼苗叶片代谢组的影响,探究亚硒酸钠喷施后香榧叶片次生代谢物的累积情况及其对果实品质的影响。 方法 以苗龄2 年的香榧幼苗为供试材料,叶面喷施亚硒酸钠(浓度为100 μg·L−1)2次,间隔15 d。第二次喷施后15 d,采取1年生叶片进行代谢组分析,以色谱-质谱研究香榧幼苗叶片代谢物差异。 结果 在正离子模式下,处理组和对照组之间有909种差异代谢物,其中69种显著变化(27种显著上调,42种显著下调)。在负离子模式下,出现433种差异代谢物,其中43种显著变化(11种显著上调,32种显著下调)。对人体有益的一些次生代谢物显著增加,如车前苷(2.827倍)、川芎内酯(2.524倍)、6-唾液乳糖(2.021倍)、阿齐沙坦(1.931倍)。 结论 香榧叶片喷施亚硒酸钠有利于促进对人体有益的次生代谢物的生物合成及其在叶片中的累积。 Abstract:Objective To investigate the effects of foliar spray of selenite on metabolome in the leaves of Torrey grandis seedlings and changes in levels of secondary metabolites in the leaves. Method Sodium selenite of 100 μg·L−1 was sprayed to the leaves of 2-year-old seedlings of T. grandis at an interval of 15 days. 15 days after the second spraying, one-year-old leaves were harvested for metabolome analysis. Result The results showed that there were 909 differential metabolites between treatment group and control under positive ionization mode, with 69 metabolites of significant changes (27 metabolites up-regulated and 42 metabolites down-regulated). Under negative ionization mode, 433 differential metabolite occurred between treatment group and control, with 43 differential metabolites of significant changes (11 metabolites up-regulated and 32 metabolites down-regulated). Some metabolites beneficial to human health increased significantly in the treated leaves of T. grandis seedlings, such as plantagoside (increasing 2.827 times as control), ligustilide (increasing 2.524 times) , 6-sialyllactose (increasing 2.021 times), and azilsartan (increasing 1.931 times). Conclusion The results indicate that selenite foliar application on the leaves of T. grandis can increase biosynthesis of beneficial metabolites and accumulation in their leaves. -
Key words:
- Torreya grandis
- / selenium fertilizer
- / metabolome
- / biofortification
-
图 1 总样本PCA分析。A-正离子模式;B-负离子模式。 CK_SE为对照,PLUS_SE为硒处理。QC为质控
Figure 1. PCA analysis of total samples. A-Positive ionization mode; B- negative ionization mode. CK_SE stands for control, PLUS_SE for selenium treatment, QC for quality control
图 2 差异代谢物筛选的PCA分析。A-正离子模式;B-负离子模式。 CK_SE为对照,PLUS_SE为硒处理。QC为质控
Figure 2. PCA used for selection of differential metabolites. A-positive ionization mode; B- negative ionization mode. CK_SE stands for control, PLUS_SE for selenium treatment, QC for quality control
图 4 差异代谢物火山图。 A-正离子化模式;B-负离子化模式
Figure 4. Volcano maps of differential metabolites. A-positive ionization mode; B-negative ionization mode.
表 1 色谱梯度洗脱程序
Table 1. Process of chromatographic gradient elution
时间 Time/min A/% B/% 0.0 98 2 1.5 98 2 3.0 15 85 10.0 0 100 10.1 98 2 11.0 98 2 12.0 98 2  下载: 导出CSV
下载: 导出CSV
表 2 差异代谢物筛选结果
Table 2. Selective results of differential metabolites
样品比对
Compared samples代谢物总数
Num. of total ident.差异显著的代谢物总数
Num. of total Sig.显著上调代谢物总数
Num. of Sig.up显著下调代谢物总数
Num. of Sig.down正离子化模式
+ Se vs. CK909 69 27 42 负离子化模式
+ Se vs. CK433 43 11 32 注:差异代谢物筛选时,设定阈值为VIP > 1.0,FC > 1.2 Note: Threshold values were set as VIP > 1.0 and FC > 1.2 when differential metabolites were selected.
下载: 导出CSV
表 3 硒肥叶面处理后正负离子化模式下上调最多10种差异代谢物
Table 3. Top 10 differential metabolites significantly up-regulated under positive and negative ionization modes in leaves of Torreya grandis seedlings after selenium foliar application
差异代谢物名称
Names of metabolites分子式
Formula分子量
Molecular weight滞留时间
RT/min质荷比
m/z倍数FC log2FC P value 正离子化模式下 N'-[6-(tert-butyl)thieno[3,2-d]pyrimidin-4-yl]-
4-methylbenzohydrazideC18H20N4OS 362.12 4.44 363.13 7.009 2.809 0.041 8 1,4-dihydroxy-1,4-dimethyl-7-(propan-2-ylidene)-
decahydroazulen-6-oneC15H24O3 234.16 7.041 235.17 3.179 1.669 0.029 7 butanedioic acid C28H36O10 532.23 6.167 533.24 2.718 1.442 0.037 6 HQH C17H24N8O5 442.18 7.086 443.18 2.712 1.439 0.000 9 (2,6-dimethylmorpholino)
(1-methyl-5-nitro-1H-pyrazol-4-yl)methanoneC11H16N4O4 290.10 1.845 291.11 2.626 1.393 0.021 6 川芎内酯ligustilide C12H14O2 190.10 7.114 191.11 2.524 1.336 0.002 7 6-唾液乳糖6-sialyllactose C23H39NO19 633.21 4.907 634.22 2.021 1.015 0.019 8 阿齐沙坦azilsartan C25H20N4O5 438.14 1.307 439.14 1.931 0.949 0.012 4 tricin 4'-O-(beta- guaiacylglyceryl) ether C27H26O11 526.15 5.199 527.15 1.874 0.906 0.045 1 aurantio-obtusin C17H14O7 330.07 5.005 331.08 1.796 0.844 0.046 2 负离子化模式下 polyphyllin V C39H62O12 722.42 8.853 721.42 3.761 1.911 0.029 8 C-pentosyl-apigenin O-p- coumaroylhexoside C35H34O16 710.18 5.152 709.17 3.399 1.765 0.048 9 车前苷plantagoside C21H22O12 466.11 4.792 465.11 2.827 1.499 0.032 2 gracillin C45H72O17 884.48 8.575 883.47 2.448 1.291 0.017 5 2-aminoethylphosphonate C2H8NO3P 125.02 4.824 124.02 1.860 0.895 0.014 1 rauwolscine hydrochloride C21H27ClN2O3 390.17 5.431 389.16 1.586 0.665 0.035 2 phenprobamate C10H13NO2 179.09 5.521 178.09 1.549 0.632 0.019 7 bruceine A C26H34O11 522.21 5.815 521.20 1.448 0.534 0.000 2 1,2,3-cyclopropanetricarboxylic acid C6H6O6 174.02 1.909 173.01 1.394 0.479 0.027 6 2-aminoadipic acid C6H11NO4 161.07 4.771 160.06 1.386 0.471 0.045 3
下载: 导出CSV
表 4 硒肥叶面处理后最富集的KEGG通路
Table 4. The most enriched KEGG pathways after selenite treatment
MapID* MapTitle P value x y n N 正离子化模式 map00230 purine metabolism 0.047 8 3 9 14 139 map00100 steroid biosynthesis 0.100 7 1 1 14 139 map00290 valine, leucine and isoleucine biosynthesis 0.100 7 1 1 14 139 map00232 caffeine metabolism 0.191 9 1 2 14 139 map00261 monobactam biosynthesis 0.191 9 1 2 14 139 map00630 glyoxylate and dicarboxylate metabolism 0.191 9 1 2 14 139 map00904 diterpenoid biosynthesis 0.191 9 1 2 14 139 map00910 nitrogen metabolism 0.191 9 1 2 14 139 map00053 ascorbate and aldarate metabolism 0.274 5 1 3 14 139 map00780 biotin metabolism 0.274 5 1 3 14 139 map02010 ABC transporters 0.305 6 2 11 14 139 map00970 aminoacyl-tRNA biosynthesis 0.346 0 2 12 14 139 map00220 arginine biosynthesis 0.349 2 1 4 14 139 map00240 pyrimidine metabolism 0.349 2 1 4 14 139 map00250 alanine, aspartate and glutamate metabolism 0.349 2 1 4 14 139 map00310 lysine degradation 0.349 2 1 4 14 139 map00750 vitamin B6 metabolism 0.349 2 1 4 14 139 map00860 porphyrin and chlorophyll metabolism 0.349 2 1 4 14 139 map00260 glycine, serine and threonine metabolism 0.416 6 1 5 14 139 map00902 monoterpenoid biosynthesis 0.416 6 1 5 14 139 map00944 flavone and flavonol biosynthesis 0.416 6 1 5 14 139 map00360 phenylalanine metabolism 0.477 6 1 6 14 139 map00945 stilbenoid, diarylheptanoid and gingerol biosynthesis 0.477 6 1 6 14 139 map00960 tropane, piperidine and pyridine alkaloid biosynthesis 0.532 6 1 7 14 139 map00941 flavonoid biosynthesis 0.582 1 1 8 14 139 map01230 biosynthesis of amino acids 0.648 7 2 15 14 139 map01100 metabolic pathways 0.800 8 9 80 14 139 负离子化模式 map00740 riboflavin metabolism 0.013 6 2 2 9 73 map00520 amino sugar and nucleotide sugar metabolism 0.071 7 2 4 9 73 map00750 vitamin B6 metabolism 0.123 2 1 1 9 73 map00040 pentose and glucuronate interconversions 0.232 8 1 2 9 73 map00440 phosphonate and phosphinate metabolism 0.232 8 1 2 9 73 map00710 carbon fixation in photosynthetic organisms 0.330 1 1 3 9 73 map00030 pentose phosphate pathway 0.416 2 1 4 9 73 map00760 nicotinate and nicotinamide metabolism 0.416 2 1 4 9 73 map02010 ABC transporters 0.492 3 1 5 9 73 map01230 biosynthesis of amino acids 0.559 5 1 6 9 73 map01200 carbon metabolism 1 1 8 9 73 *注:(1) MapID:富集的KEGG Pathway的ID;(2) MapTitle:富集的KEGG Pathway名称;(3) P value:富集分析的P value;(4) x:与该通路相关的差异代谢物的数目;(5) y:与该通路相关的背景(所有)代谢物的数目;(6) n:KEGG注释的差异代谢物数目;(7) N:KEGG注释的背景(所有)代谢物的数目。
Note: (1) MapID: the ID of enriched KEGG pathway; (2) MapTitle: name of the enriched KEGG pathway; (3) p value: p value of enrichment analysis; (4) x: the number of differential metabolites related to KEGG pathway; (5) y: the number of all the secondary metabolites related to KEGG pathway; (6) n: the number of differential secondary metabolites annotated in KEGG; (7) N: the number of all the secondary metabolites annotated in KEGG.
下载: 导出CSV
-
[1] Minich W B. Selenium metabolism and biosynthesis of selenoproteins in the human body[J]. Biochemistry (Mosc), 2022, 87(Suppl 1): S168-S177. [2] Gladyshev V N, Arnér E S, Berry M J, et al. Selenoprotein gene nomenclature[J]. J Biol Chem, 2016, 291(46): 24036-24040. doi: 10.1074/jbc.M116.756155 [3] Elhodaky M, Diamond A M. Selenium-binding protein 1 in human health and disease[J]. Int J Mol Sci, 2018, 19(11): 3437-3450. doi: 10.3390/ijms19113437 [4] Lei C, Niu X, Ma X, et al. Is selenium deficiency really the cause of Keshan disease?[J]. Environ Geochem Health, 2011, 33(2): 183-188. doi: 10.1007/s10653-010-9331-9 [5] Zhou H, Wang T, Li Q, et al. Prevention of Keshan disease by selenium supplementation: a systematic review and meta-analysis[J]. Biol Trace Elem Res, 2018, 186(1): 98-105. doi: 10.1007/s12011-018-1302-5 [6] Yao Y, Pei F, Kang P. Selenium, iodine, and the relation with Kashin-Beck disease[J]. Nutrition, 2011, 27(11-12): 1095-1100. doi: 10.1016/j.nut.2011.03.002 [7] Flores-Mateo G, Navas-Acien A, Pastor-Barriuso R, et al. Selenium and coronary heart disease: a meta-analysis[J]. Am J Clin Nutr, 2006, 84(4): 762-773. doi: 10.1093/ajcn/84.4.762 [8] Cui S, Luo Y, Li Y, et al. Selenium alleviates heart remodeling through Sirt1/AKT/GSK-3β pathway[J]. Int Immunopharmacol, 2022, 111: 109158. doi: 10.1016/j.intimp.2022.109158 [9] Yuan S, Mason A M, Carter P, et al. Selenium and cancer risk: Wide-angled Mendelian randomization analysis[J]. Int J Cancer, 2022, 150(7): 1134-1140. doi: 10.1002/ijc.33902 [10] Rua R M, Nogales F, Carreras O, et al. Selenium, selenoproteins and cancer of the thyroid[J]. J Trace Elem Med Biol, 2023, 76: 127115. doi: 10.1016/j.jtemb.2022.127115 [11] Gorini F, Sabatino L, Pingitore A, et al. Selenium: an element of life essential for thyroid function[J]. Molecules, 2021, 26(23): 7084. doi: 10.3390/molecules26237084 [12] Wang F, Li C, Li S, et al. Selenium and thyroid diseases[J]. Front Endocrinol (Lausanne), 2023, 14: 1133000. doi: 10.3389/fendo.2023.1133000 [13] Alcolea V, Pérez-Silanes S. Selenium as an interesting option for the treatment of Chagas disease: A review[J]. Eur J Med Chem, 2020, 206: 112673. doi: 10.1016/j.ejmech.2020.112673 [14] Martín-Escolano R, Etxebeste-Mitxeltorena M, Martín-Escolano J, et al. Selenium derivatives as promising therapy for Chagas disease: in vitro and in vivo studies[J]. ACS Infect Dis, 2021, 7(6): 1727-1738. doi: 10.1021/acsinfecdis.1c00048 [15] Hossain A, Skalicky M, Brestic M, et al. Selenium biofortification: roles, mechanisms, responses and prospects[J]. Molecules, 2021, 26(4): 881. doi: 10.3390/molecules26040881 [16] Ye Y, Qu J, Pu Y, et al. Selenium biofortification of crop food by beneficial microorganisms[J]. J Fungi (Basel), 2020, 6(2): 59. doi: 10.3390/jof6020059 [17] Yang D, Hu C, Wang X, et al. Microbes: a potential tool for selenium biofortification[J]. Metallomics, 2021, 13(10): mfab054. doi: 10.1093/mtomcs/mfab054 [18] Duborská E, Šebesta M, Matulová M, et al. Current strategies for selenium and iodine biofortification in crop plants[J]. Nutrients, 2022, 14(22): 4717. doi: 10.3390/nu14224717 [19] Zhang L, Chu C. Selenium uptake, transport, metabolism, reutilization, and biofortification in rice[J]. Rice (N Y), 2022, 15(1): 30. doi: 10.1186/s12284-022-00572-6 [20] Silva M A, de Sousa G F, Corguinha A P B, et al. Selenium biofortification of soybean genotypes in a tropical soil via Se-enriched phosphate fertilizers[J]. Front Plant Sci, 2022, 13: 988140. doi: 10.3389/fpls.2022.988140 [21] Liu Y, Huang S, Jiang Z, et al. Selenium biofortification modulates plant growth, microelement and heavy metal concentrations, selenium uptake, and accumulation in black-grained wheat[J]. Front Plant Sci, 2021, 12: 748523. doi: 10.3389/fpls.2021.748523 [22] Lei H, Zhou M, Li B, et al. Humic acid chelated selenium is suitable for wheat biofortification[J]. J Sci Food Agric, 2023, 103(10): 4887-4898. doi: 10.1002/jsfa.12564 [23] Groth S, Budke C, Neugart S, et al. Influence of a selenium biofortification on antioxidant properties and phenolic compounds of apples (Malus domestica)[J]. Antioxidants (Basel), 2020, 9(2): 187. doi: 10.3390/antiox9020187 [24] Hu T, Hui G, Li H, et al. Selenium biofortification in Hericium erinaceus (Lion's Mane mushroom) and its in vitro bioaccessibility[J]. Food Chem, 2020, 331: 127287. doi: 10.1016/j.foodchem.2020.127287 [25] Xu M, Zhu S, Wang L, et al. Influence of Selenium Biofortification on the Growth and Bioactive Metabolites of Ganoderma lucidum[J]. Foods, 2021, 10(8): 1860. doi: 10.3390/foods10081860 [26] Golob A, Novak T, Maršić N K, et al. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes) and increases their contents in tubers[J]. Plant Physiol Biochem, 2020, 150: 234-243. doi: 10.1016/j.plaphy.2020.02.044 [27] Francini A, Quattrini E, Giuffrida F, et al. Biofortification of baby leafy vegetables using nutrient solution containing selenium[J]. J Sci Food Agric, 2023, 103(11): 5472-5480. doi: 10.1002/jsfa.12622 [28] Manfio D, Rodrigues M N F, Savi G D, et al. Brazil nut (Bertholettia excels H. B. K. ) selenium distribution and physical chemical characteristics of shell, brown skin and edible part from two Amazon regions[J]. Asian J Agri Rural Dev, 2012, 2(2): 287-293. [29] 陈红星, 陈素贞, 张 迟, 等. 香榧新品种‘玉山果榧’的选育[J]. 中国果树, 2021(8):79-80,109. [30] 胡文翠, 张 迟, 厉 锋, 等. 香榧良种‘朱岩榧’的选育[J]. 果树学报, 2021, 38(3):455-458. [31] 任少华, 龙成昌, 王 陈, 等. 香榧及其优质高产栽培技术探讨[J]. 现代园艺, 2021, 44(19):72-74. doi: 10.3969/j.issn.1006-4958.2021.19.028 [32] 李爱华, 许秀环, 李金柱, 等. 施加硒肥条件下的印度梨形孢接种对香榧幼苗根系发育、气体交换及其叶片硒累积的影响[J]. 安徽农业大学学报, 2022, 49(5):735-740. [33] Want E J, Masson P, Michopoulos F, et al. Global metabolic profiling of animal and human tissues via UPLC-MS[J]. Nature Protocols, 2012, 8(1): 17-32. [34] 阙小峰, 余 雁, 方志成, 等. 葡萄叶面肥硒特性及可溶态硒分布研究[J]. 湖北农业科学, 2022, 61(7):31-34. [35] 陈锦平, 农梦玲, 何景云, 等. 不同硒肥处理对杧果果实硒含量及品种的影响[J]. 热带农业科学, 2021, 41(6):1-5. [36] 王永刚. 氨基酸硒叶面肥在晚秋黄梨上的应用效果[J]. 河北果树, 2017, 168(4):3-5. [37] Mao Z G, Tian L Y, Liu J, et al. Ligustilide ameliorates hippocampal neuronal injury after cerebral ischemia reperfusion through activating PINK1/Parkin-dependent mitophagy[J]. Phytomedicine, 2022, 101: 154111. doi: 10.1016/j.phymed.2022.154111 [38] Hsu R J, Peng K Y, Hsu W L, et al. Z-ligustilide induces c-Myc-dependent apoptosis via activation of ER-stress signaling in hypoxic oral cancer Cells[J]. Front Oncol, 2022, 12: 824043. doi: 10.3389/fonc.2022.824043 [39] Ma J, Mei J, Lu J, et al. Ligustilide promotes apoptosis of cancer-associated fibroblasts via the TLR4 pathways[J]. Food Chem Toxicol, 2020, 135: 110991. doi: 10.1016/j.fct.2019.110991 [40] Sodhi C P, Wipf P, Yamaguchi Y, et al. The human milk oligosaccharides 2'-fucosyllactose and 6'-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling[J]. Pediatr Res, 2021, 89(1): 91-101. doi: 10.1038/s41390-020-0852-3 [41] Nguyen T L L, Jin Y, Kim L, et al. Inhibitory effects of 6'-sialyllactose on angiotensin II-induced proliferation, migration, and osteogenic switching in vascular smooth muscle cells[J]. Arch Pharm Res, 2022, 45(9): 658-670. doi: 10.1007/s12272-022-01404-3 [42] Zaiken K, Cheng J W. Azilsartan medoxomil: a new Angiotensin receptor blocker[J]. Clin Ther, 2011, 33(11): 1577-1589. doi: 10.1016/j.clinthera.2011.10.007 [43] Baker W L, White W B. Azilsartan medoxomil: a new angiotensin II receptor antagonist for treatment of hypertension[J]. Ann Pharmacother, 2011, 45(12): 1506-1515. doi: 10.1345/aph.1Q468 [44] Pradhan A, Tiwari A, Sethi R. Azilsartan: Current Evidence and Perspectives in Management of Hypertension[J]. Int J Hypertens, 2019, 2019: 1824621. [45] Han J, Tang H, Yao L, et al. Azilsartan protects against hyperglycemia-induced hyperpermeability of the blood-brain barrier[J]. Bioengineered, 2021, 12(1): 3621-3633. doi: 10.1080/21655979.2021.1948950 [46] Yamada H, Nagai T, Takemoto N, et al. Plantagoside, a novel alpha-mannosidase inhibitor isolated from the seeds of Plantago asiatica, suppresses immune response[J]. Biochem Biophys Res Commun, 1989, 165(3): 1292-1298. doi: 10.1016/0006-291X(89)92743-5 [47] Bajrai L H, Alharbi A S, El-Day M M, et al. Identification of Antiviral Compounds against Monkeypox Virus Profilin-like Protein A42R from Plantago lanceolata[J]. Molecules, 2022, 27(22): 7718. doi: 10.3390/molecules27227718 [48] Rodríguez-Arce E, Saldías M. Antioxidant properties of flavonoid metal complexes and their potential inclusion in the development of novel strategies for the treatment against neurodegenerative diseases[J]. Biomed Pharmacother, 2021, 143: 112236. doi: 10.1016/j.biopha.2021.112236 [49] Gul S, Maqbool M F, Zheng D, et al. Alpinetin: a Dietary Flavonoid with Diverse Anticancer Effects[J]. Appl Biochem Biotechnol, 2022, 194(9): 4220-4243. doi: 10.1007/s12010-022-03960-2 [50] Serafini M, Peluso I, Raguzzini A. Flavonoids as anti-inflammatory agents[J]. Proc Nutr Soc, 2010, 69(3): 273-278. doi: 10.1017/S002966511000162X -

 点击查看大图
点击查看大图
计量
- 文章访问数: 890
- HTML全文浏览量: 370
- PDF下载量: 16
- 被引次数: 0
