



-
天然橡胶是一种重要的工业原料和战略物资,其主要来源于橡胶树(Hevea brasiliensis (Willd.ex A. Juss.) Muell.Arg.)。目前,橡胶树生产上所应用的种苗主要为芽接苗,即实生苗作为砧木嫁接优良品种的接穗所培育的种苗。因砧木来源于自然授粉的种子,遗传基础不一致,导致芽接苗植株间产量、生长势等存在显著差异;而橡胶树体胚苗是由体细胞胚直接发育而来的种苗,具有自主发育的主根、无需嫁接在砧木上的幼态化无性系(又称橡胶树自根幼态无性系),较目前生产上应用的种苗-芽接树,产量高10%~30%,生长快10%~20%,是天然橡胶产业新一代种植材料[1-4]。
自1978年王泽云等成功移植成活橡胶树花药体胚苗开始[5],法国、马来西亚、印度尼西亚、斯里兰卡、印度等国投入大量的人力、物力、财力对橡胶树的体胚苗生产进行了广泛的研究,取得了一定的进展,但体胚诱导率和植株再生频率较低,生产成本较高,无法满足生产需求,成为制约体胚苗工厂化生产的瓶颈[6-8]。橡胶树雄蕊具有数量大、便于采集、容易表面灭菌处理、易于诱导愈伤组织等优点,是研发橡胶树体胚发生体系应用最多的外植体。研究发现,橡胶树雄蕊诱导的愈伤组织和体胚主要来源于花药壁体细胞[9-13],其愈伤组织和体胚再生植株的倍性同母本一致。以花药诱导的体胚作为外植体,诱导次生体胚循环增殖,可实现一次花药体胚发生进一步诱导多次体胚发生,且次生体胚植株再生频率高达86%以上,成为体胚大量增殖的新途径[14-16]。但目前仍有许多品种的花药难以诱导体胚形成,即使能够诱导体胚的品种,也仅有少数品种的花药外植体体胚诱导率能达到50%[17-23],制约了这一技术的广泛应用。
植物体胚发生能力显著性的受遗传变异影响,存在细胞核因子遗传及核质互作遗传影响现象,且存在遗传背景与培养基互作影响现象,通过分析体胚发生时的转录组表明,体胚发生能力差异悬殊的基因型间存在显著的差异基因表达。在珍珠粟再生能力遗传研究中发现,胚性愈伤组织数量更大比例表现为核质互作遗传影响,而再生频率主要表现为细胞核遗传影响[24]。在板栗体胚发生研究中发现,遗传背景与培养基对体胚发生效率存在着显著的互作影响,当用美国板栗体胚发生标准流程时,中国板栗不能诱导形成胚性组织,在‘中国板栗与美国板栗杂交后代’中,杂交F1代、F1代与美国板栗回交BC1代也不能诱导形成胚性组织,而BC1代与美国板栗回交BC2代、BC2代与美国板栗回交形成BC3代的自交BC3F3代能够诱导形成胚性组织,推断杂交后代中中国板栗基因的占比可能存在一个体胚发生阈值,超过该阈值的杂交后代不能诱导形成胚性组织;但当用欧洲板栗体胚发生标准流程时,中国板栗能够诱导形成胚性组织,而美国板栗以及中国板栗与美国板栗的杂交后代F1、BC1、BC2、BC3F3均不能诱导形成胚性组织,系统发育研究表明栗属起源于亚洲,首先向西迁移至欧洲,然后迁移至北美,因此,作者推断中国板栗与欧洲板栗的亲缘关系更近,所以,用欧洲板栗体胚发生流程能够诱导中国板栗形成胚性组织[25]。通过对不同体胚发生能力棉花基因型的转录组进行对比分析,表明不同基因型的胚性愈伤组织是高度同质的,而不同基因型的非胚性愈伤组织是高度异质的[26]。通过分析棉花高效胚胎发生基因型和顽拗胚胎发生基因型的早期体胚发生转录组,表明高效胚胎发生基因型的差异表达基因在脂肪酸、色氨酸和丙酮酸代谢中显著富集,而顽拗胚胎发生基因型的差异表达基因则在 DNA 构象变化中显著富集[27]。
橡胶树品种遗传基础狭窄,为通过遗传背景分组优化不同基因型体胚发生体系提供了条件。本研究将重点探讨橡胶树体胚发生能力在不同基因型之间的差异,利用前期试验中在多个品种体胚诱导率都较高的两种培养基,对32个橡胶树品种花药外植体的体胚发生进行研究,以期明确橡胶树不同品种花药体胚发生能力与遗传背景的相关性,为通过基因型分组优化不同品种的体胚发生体系提供依据,为突破橡胶树优良品种体胚苗大量增殖的瓶颈提供基础。
-
以国家橡胶树种质资源圃(海南省儋州市宝岛新村)保存的32个橡胶树品种为材料(表1)。于春季3月份采集黄中带绿、未开放的雄花(花粉发育处于单核靠边期),用75%乙醇表面灭菌60 s后,立即用0.1%HgCl2处理10 min,最后用无菌水冲洗5次,每次3 min[14]。表面灭菌后的雄花用于后续愈伤组织诱导和体胚发生研究。
表 1 不同橡胶树品种花药愈伤组织、体胚、子叶形体胚诱导率
Table 1. Induction rate of anther callus, somatic embryo, cotyledon somatic embryo of different rubber tree cultivars
品种世代分级
Cultivar classification品种
Cultivar愈伤组织形成率/%
Callus induction
rate体胚形成率/%
Somatic embryo
induction rate子叶胚形成率/%
Cotyledon somatic
embryo induction rate体胚诱导能力分级
Somatic embryo
induction ability
classificationM培养基
M mediumS培养基
S mediumM培养基
M mediumS培养基
S mediumM培养基
M mediumS培养基
S medium初生代无性系
First--generation clonePR107 (82.2±16.8) (61.9±21.8) (4.2±3.3) i (3±3) g (0) e (0) c 难 Recalcitrant GT1 (75±8.1) (75.8±16.2) (0) i (0.3±0.6) g (0) e (0) c 难 Recalcitrant PB86 (66.5±11.7) (55.5±25.1) (26.8±13.8) efgh (19.8±9) defg (0) e (0) c 易 High Tjirl (59.2±15) (58.9±19.3) (0) i (0) g (0) e (0) c 难 Recalcitrant 海垦1 (73.7±2.1) (50.8±9.6) (31.8±3.7) cdef (7.9±2.1) g (4.1±1.5) de (0) c 易 High 天任31-45 (35.9±22.4) (28.9±18.6) (0) i (0) g (0) e (0) c 难 Recalcitrant 合口3-11 (68.3±31) (74.6±26.2) (10.7±3.8) hi (11±3) fg (0) e (0) c 可 Inducible 次生代无性系
Second-generation clone云研77-2 (40.9±10.9) (34.4±19.5) (0) i (0) g (0) e (0) c 难 Recalcitrant 云研77-4 (57.2±23.2) (46.7±19.6) (0) i (0) g (0) e (0) c 难 Recalcitrant 云研73-46 (71.8±13) (73.7±8.6) (6.7±1.4) i (0) g (0) e (0) c 难 Recalcitrant 保亭1-285 (68.9±9.8) (59.6±22.9) (0) i (0) g (0) e (0) c 难 Recalcitrant 93-114 (77.5±6.6) (66±13.5) (0) i (0) g (0) e (0) c 难 Recalcitrant RRIM513 (74.7±1.4) (76.3±21.1) (0) i (0) g (0) e (0) c 难 Recalcitrant IAN873 (74.2±18.5) (73.5±14.1) (0) i (0) g (0) e (0) c 难 Recalcitrant 大丰95 (55.5±27.2) (37.9±15.4) (13.5±4.1) ghi (30.9±34.3) cdef (1.7±2.9) de (6±10.3) b 易 High RRIM600 (77.8±19.2) (76.9±20.7) (13.2±13.8) ghi (11±10.1) fg (0) e (2.5±4.3) bc 可 Inducible 徐育141-2 (68.9±33.2) (61.3±43.7) (48.1±34.2) bc (47.2±39.1) bc (13.3±10.4) b (3.8±1.1) bc 易 High 文昌217 (87.6±6.9) (92.4±10.9) (17.6±1.2) fghi (16±2.6) defg (0) e (0) c 可 Inducible 三生代无性系
Third-generation clone湛试32713 (69.2±18.8) (56.7±10.3) (0.7±1.2) i (0) g (0) e (0) c 难 Recalcitrant 大丰117 (51.2±3.1) (43.3±8.4) (15.6±0.7) fghi (0) g (3.2±0.3) de (0) c 可 Inducible 热研217 (70±30) (49.3±33.4) (0) i (0) g (0) e (0) c 难 Recalcitrant 文昌11 (94.4±5.9) (89.8±10) (11.5±2.2) hi (6±1.7) g (0.6±1) e (0) c 可 Inducible 热研917 (86±5.2) (73.1±17) (47.6±19) bc (35.7±24.4) bcd (0.6±1) e (0) c 易 High 热研73397 (85.3±2.7) (79.1±6.7) (95±5) a (94±10.4) a (23.1±8.2) a (17.3±5.1) a 易 High 徐育3 (79±20.1) (74.8±22.3) (46.7±2.6) bcd (56±1.7) b (6.6±0.9) cd (5.6±1) b 易 High 保亭3412 (94.1±5.2) (94.3±6.1) (4.3±1.5) i (3.7±1.4) g (0) e (0) c 难 Recalcitrant 热研88-13 (90.1±7.9) (81.3±7) (83.5±11.7) a (77.4±19.9) a (11.5±6) bc (3.8±3.9) bc 易 High PB235 (46.9±12) (53.7±13.4) (35.5±0.8) cde (37.1±1.1) bcd (2.9±2.9) de (6±0.3) b 易 High 四生代无性系
Fourth-generation clone热垦628 (87.4±4.6) (80.8±20.1) (4.4±4.2) i (12.4±9.7) efg (0) e (0) c 可 Inducible 热垦525 (79.4±10.3) (67.2±11.1) (56.9±19) b (35±18.2) bcde (7.2±4.6) cd (6.1±5.4) b 易 High 热研879 (90.9±10.1) (78.6±16.6) (31±20.7) cdefg (32.5±15) cdef (3.7±6.4) de (0) c 易 High 热研918 (96.5±3.1) (79±22) (29.7±10.4) defg (36.1±17.7) bcd (7.5±5) cd (3.5±4.3) bc 易 High 注:括号内加减号前数字代表三次重复的均值,加减号后数字代表标准差。次生代无性系海垦2愈伤组织形成率达96.1%,体胚形成率 (45.0±3.5) %,属于易诱导品种,数据来自文献[5,17]。
Notes: In parentheses, the numbers before the plus and minus signs represent the mean of three repetitions, and the numbers after the plus and minus signs represent the standard deviation. The callus induction rate and the somatic embryo induction rate of the secondary-generation clone Haiken 2 were 96.1% and (45.0±3.5) %, respectively, which belonged to high frequency embryogenic cultivar, the data came from the literatures[5,17]. -
改良MS培养基用于愈伤组织诱导和体胚发育,其中,大量元素、微量元素、铁盐所需的药品和蔗糖购自广州化学试剂厂;有机营养、植物激素和生长调节剂采购于上海生物工程技术有限公司和Sigma-Aldrich公司;植物凝胶采购于Sigma-Aldrich公司。
花药愈伤组织诱导采用平底试管为培养容器。体胚分化阶段采用培养皿作为培养容器。
-
首先将雄蕊从雄花中剥出,然后接种到愈伤组织诱导培养基上(M培养基和S培养基),每个品种每处理接种数量超过120粒花药,重复3次,26 ℃~28 ℃暗室中培养40 d~60 d。形成愈伤组织后,将花药愈伤组织接种到体胚发生培养基上(E培养基),于24~26 ℃暗室中培养40~60 d。M培养基为改良MS添加4.6 μmol·L−1 ZT,4.1 μmol·L−1 Picloram,4.5 μmol·L−1 2,4-D。S培养基为改良MS添加2.3 μmol·L−1 KT,2.1 μmol·L−1 Picloram,2.3 μmol·L−1 2,4-D。E培养基为改良MS添加2.2 μmol·L−1 6-BA,14.0 μmol·L−1 KT,1.4 μmol·L−1 GA3,0.1 μmol·L−1 NAA,3.8 μmol·L−1 ABA[15]。以上培养基均添加204.5 mmol·L−1蔗糖,2.2 g·L−1 Phytagel,灭菌前调整pH至5.8。
-
愈伤组织形成率= 形成愈伤组织的雄蕊数/接种雄蕊总数 × 100%。
体细胞胚形成率= 形成体细胞胚的愈伤组织数/接种愈伤组织总数 × 100%。
子叶形体细胞胚形成率= 形成子叶形体细胞胚的愈伤组织数/接种愈伤组织总数 × 100%。
试验结果采用SPSS的Ducan法进行差异性分析。
-
体胚苗倍性鉴定用Sysmex Cy-Flow®Cube8流式细胞仪进行。鉴定方法参考仪器附送试剂使用方法(
http://www.sysmex-partec.com ),即取200 mg新鲜叶片置于塑料培养皿中,添加0.5 mL裂解缓冲液,用刀片快速切碎,30 μm孔径滤网过滤,添加1.6 mL DAPI染色体液,室温静置30 s后上机检测。每株体胚苗随机取1片叶片进行检测。流式数据使用机器自带软件作图。以已知二倍体和三倍体植株叶片混合检测作图为对照。 -
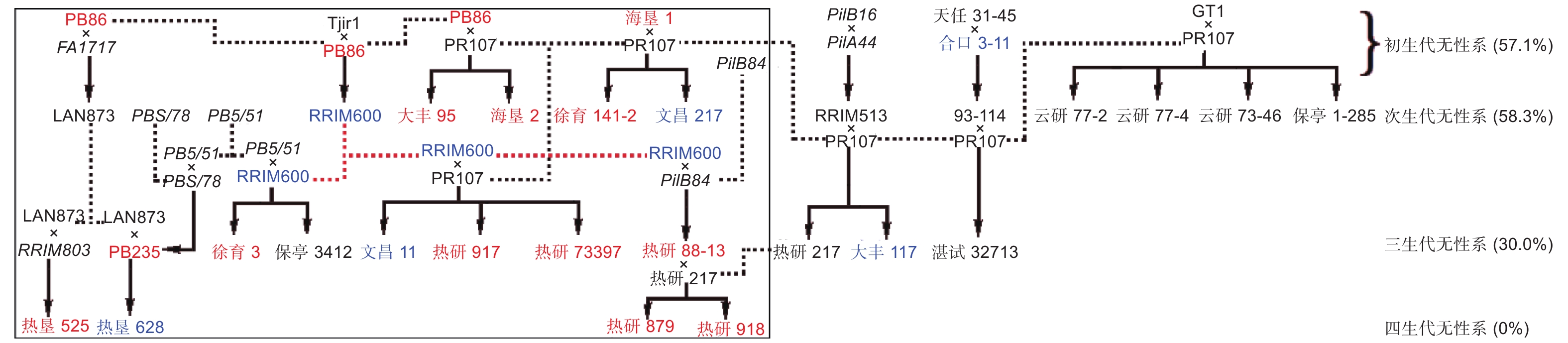
根据亲本来源,对国家橡胶树种质资源圃保存的32个橡胶树品种进行系谱分析,试验所用32个橡胶树品种分别属于初生代无性系、次生代无性系、三生代无性系和四生代无性系(图1)。PR107、GT1、PB86、Tjirl、海垦1、天任31-45、合口3-11这7个品种为初生代无性系,其余25个品种中的23个与以上7个初生代无性系中的1~3个存在亲缘关系,表明我国橡胶树栽培品种亲本来源狭窄。此外,图1系谱中,含初生代无性系遗传背景的后代中,以含PR107亲本的后代最多,共有14个,以PR107为父本的品种有GT1、93-114、RRIM513、PB86、RRIM600、海垦1共6个母本组。

图 1 橡胶树系谱和花药体胚发生能力差异分析
Figure 1. Pedigree analysis of rubber tree and analysis of differences in anther embryogenic ability
-
橡胶树花药愈伤组织、体细胞胚和体胚苗的诱导过程见图2。参试32个品种均能诱导出愈伤组织,多数品种在M培养基和S培养基上的愈伤组织诱导率接近,其中,31个品种的愈伤组织形成率均达50%以上(表1),表明所试愈伤组织诱导培养基在橡胶树花药愈伤组织诱导中具有广谱性。
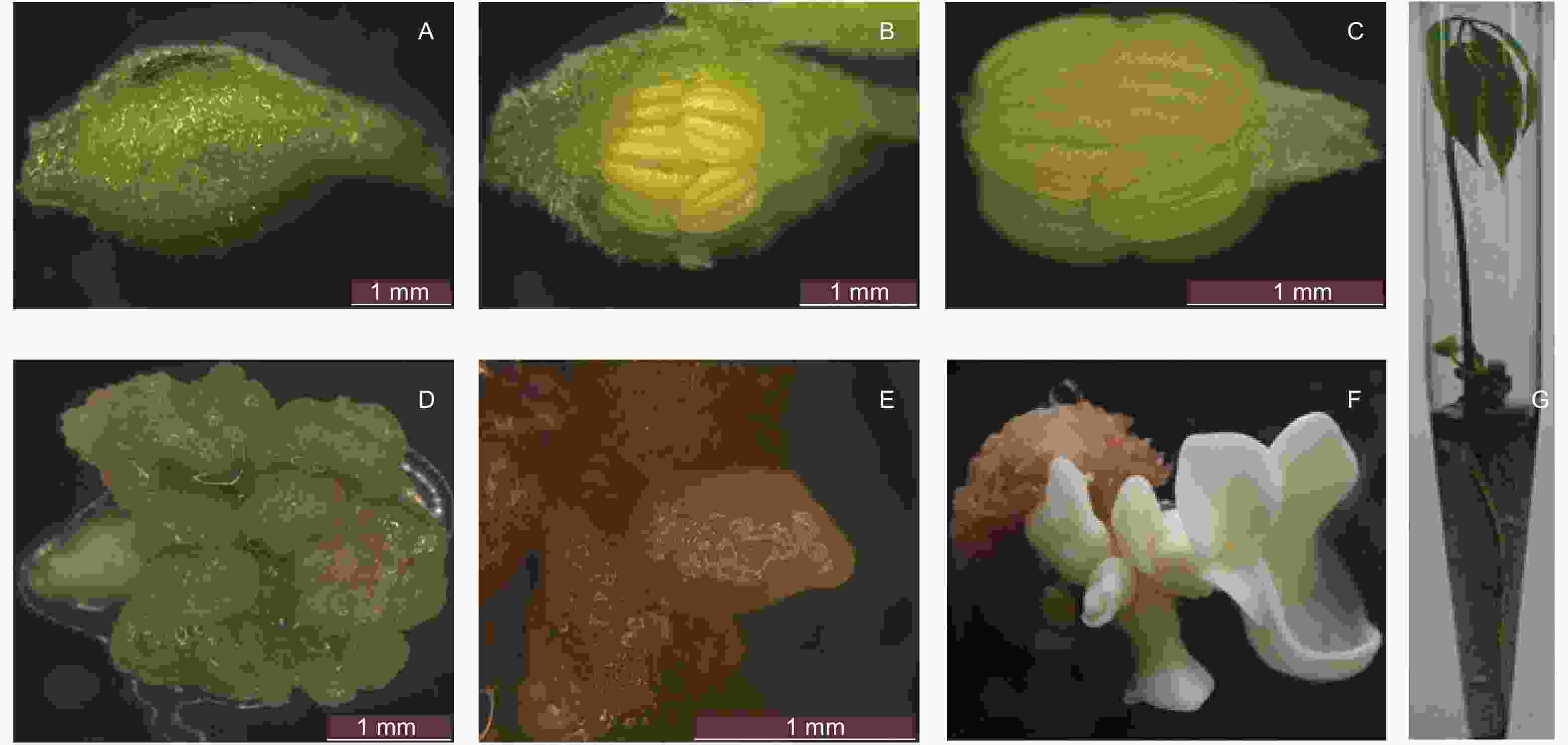
图 2 橡胶树花药体胚苗的诱导过程图
Figure 2. The Induction process of anther somatic embryo plantlet of rubber tree
愈伤组织转移到诱导体胚发生的E培养基后,来自M培养基和S培养基诱导的愈伤组织在大多数品种上体细胞胚诱导率较接近,只有个别品种存在显著性差异。23个品种能够诱导形成体细胞胚,且有14个品种能够诱导形成子叶形体细胞胚和体胚苗,其中,热研73397、热研88-13、热垦525和徐育3这4个品种的体细胞胚诱导率在50%以上,说明所试体胚发生培养基在橡胶树体胚发生中具有通用性,但仍有14个品种的体细胞胚诱导率在10%以下,且有9个品种未能诱导出体细胞胚(表1)。
-
根据品种体胚诱导能力,将体胚诱导率<10%的品种定义为难诱导品种,体胚诱导率在≥10%且≤20%的品种定义为可诱导品种,体胚诱导率>20%的品种定义为易诱导品种(图1)。7个初生代无性系中,GT1、PR107、天任31-45、Tjirl属于难诱导品种,合口3-11属于可诱导品种,PB86和海垦1属易诱导品种。12个次生代无性系中,云研77-2、云研77-4、云研73-46、保亭1-285、93-114、RRIM513、IAN873属于难诱导品种,RRIM600、文昌217属于可诱导品种,大丰95、海垦2[5,17]、徐育141-2属于易诱导品种。10个三生代无性系中,湛试32713、热研217、保亭3412属于难诱导品种,大丰117、文昌11属于可诱导品种,PB235、徐育3、热研917(热研7-20-59)、热研73397、热研88-13属于易诱导品种。4个四生代无性系中,热垦628属于可诱导品种,热垦525、热研879、热研918(热研8-333)属于易诱导品种。分析发现,已知体胚诱导率的初生代、次生代、三生代和四生代无性系中难诱导体胚发生品种占比分别为57.1%、58.3%、30.0%、0.0%。
亲本诱导能力决定其子代品种体胚诱导能力。2个亲本属于难诱导品种,其杂交后代多数为难诱导品种,如GT1 × PR107、93-114 × PR107、RRIM513 × PR107这3个杂交组合的后代有7个,6个为难诱导品种,只有1个为可诱导品种;1个亲本为难诱导品种,另1个为易或可诱导品种,其杂交后代多数属于易或可诱导品种,如PB86 × PR107、Tjir1 × PB86、海垦1 × PR107、天任31-45 × 合口3-11、RRIM600 × PR107、IAN873 × PB235、热研88-13 × 热研217这7个杂交组合的后代有12个,其中,7个品种属于易诱导品种,4个品种为可诱导品种,只有1个品种为难诱导品种。
具有相同亲本的橡胶树品种体细胞胚诱导率差异显著,且存在超亲现象。RRIM600 × PR107子代热研73397、热研917和文昌11三个品种中,热研73397的体胚诱导率和子叶形体胚诱导率分别为95.0±5%、23.1±8.2%,显著性高于热研917(47.6±19%、0.6±1%)、文昌11(11.5±2.2%、0.6±1%)和双亲;海垦1 × PR107子代徐育141-2和文昌217中,徐育141-2的体胚诱导率和子叶形体胚诱导率(48.1±34.2%、13.3±10.4%)显著性高于文昌217(17.6±1.2%、0)和双亲;RRIM513 × PR107子代热研217和大丰117中,热研217的体胚诱导率为零,而大丰117在M培养基上体胚诱导率达到15.6±0.7%,且显著性高于双亲;GT1 × PR107子代云研77-2、云研77-4、云研73-46和保亭1-285四个品种中,只有云研73-46在所试培养基上能够诱导得到体胚。
-
流式细胞仪倍性检测结果(图3和表2)表明:混合已知二倍体和三倍体植株叶片DNA相对含量在200和300位置均有波峰作为对照(图3A),二倍体的DNA相对含量为200(图3B),三倍体植株为300(图3C),流式细胞仪倍性检测可以准确鉴定出橡胶树体胚苗的倍性。
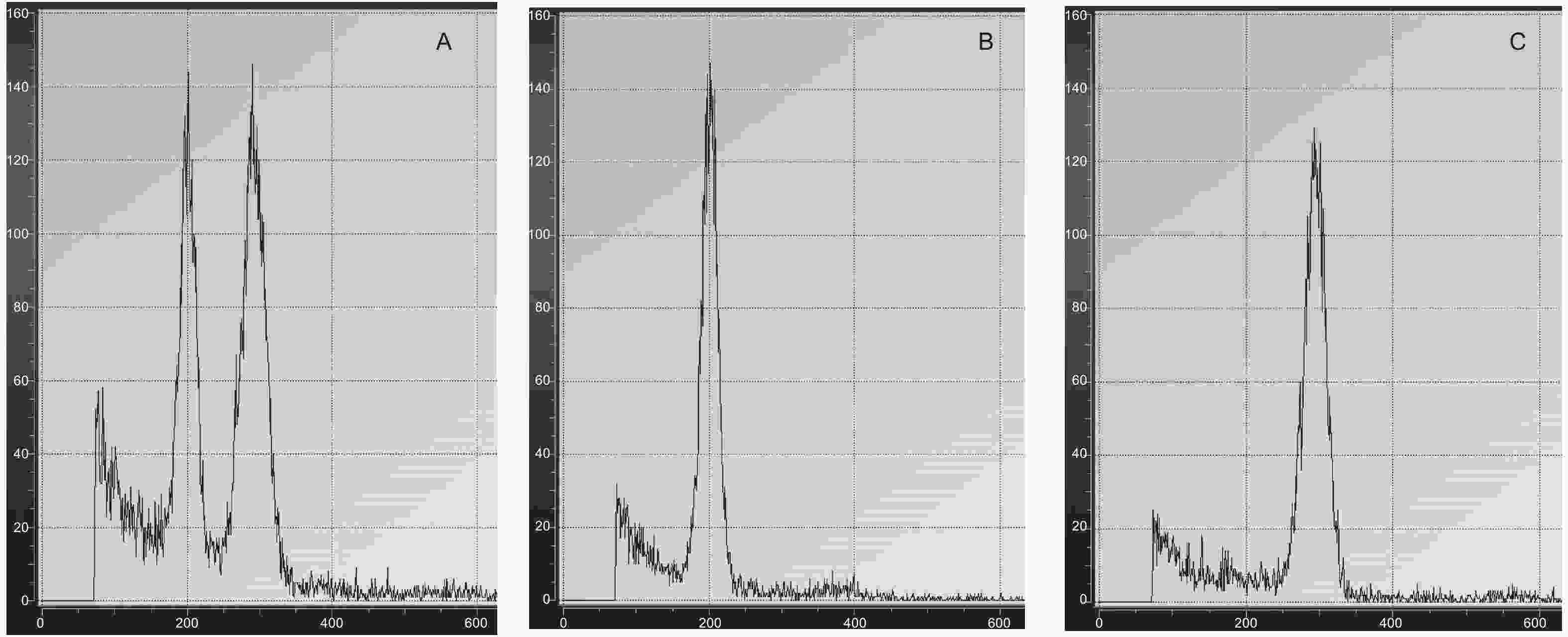
图 3 流式细胞倍性检测
Figure 3. Flow cytometric analysis
表 2 橡胶树不同基因型体胚苗倍性检测结果
Table 2. Ploidy test results of somatic embryo plants of different genotypes of rubber tree
品种
Cultivar母本倍性
Mother
plant
ploidy检测体胚苗株数
Number of somatic
embryo plants tested二倍体株数
Number of
diploid plants三倍体株数
Number of
triploid plants热垦525 二倍体 113 113 — 徐育141-2 二倍体 100 100 — 热研73397 二倍体 86 86 — 热研917 二倍体 50 50 — 热研88-13 二倍体 50 50 — 海垦2 二倍体 50 50 — PR107 二倍体 34 34 — 文昌217 二倍体 25 25 — 文昌11 二倍体 16 16 — 热研918 二倍体 14 14 — 云研73-46 三倍体 8 — 8 合口3-11 二倍体 3 3 — 海垦1 二倍体 1 1 — 云研774 三倍体 1 — 1 保亭1-285 二倍体 1 1 — 橡胶树不同基因型体胚苗倍性鉴定统计结果见表2,从15个品种共552株体胚苗倍性鉴定结果看,不同基因型体胚苗同其母本倍性一致,证明多基因型花药体胚苗来源于体细胞。
-
目前,未有深入开展橡胶树多基因型体胚发生能力研究的报道,在检索的少量研究报道中,橡胶树品种热研879花药体细胞胚诱导率为10%~14%,RRIM600为11%~49%,热垦525为51%左右,热研73397为21%左右,都属于易或可诱导体细胞胚品种;云研77-2、云研77-4、云研73-46、GT1和PR107花药体细胞胚诱导率都在10%以下,甚至为0,都属于难诱导体细胞胚品种,与本研究结果一致[17-23]。本研究的32个橡胶树品种中,有18个品种的体细胞胚诱导率在10%以上,有14个品种能够诱导得到子叶形体细胞胚,其中,热研73397的体细胞胚诱导率最高达95%,热研879的最高达32%以上,显著高于前人的研究结果。
-
Hodges等发现,在相同的培养条件下,玉米A188自交系的体细胞再生能力显著高于其他品系,以其作为亲本与无再生能力的自交系进行杂交,杂交后代具有再生能力;另外以两个再生能力差的自交系进行杂交,杂交后代再生能力也差,表现出不同基因型玉米的体细胞再生能力有较大差异且可遗传[31]。Parrott等发现,基因型对大豆体细胞胚发生能力有很大影响,有良好再生能力的品系在其系谱中都有一个或两个高再生能力的祖先亲本,且高再生能力基因型与低再生能力基因型的杂交F1代表现出中等再生能力,表明大豆体细胞再生能力可遗传[32]。Niskanen等发现,父本和母本的基因型对欧洲赤松体细胞胚发生都有影响,在胚性组织起始诱导阶段,母本的影响大于父本,在胚性组织长期继代增殖阶段,父本的影响相对母本增加[33]。在遗传变异对甜根子草胚性愈伤组织诱导和再生能力研究中发现,愈伤组织和胚性愈伤组织的诱导能力受遗传变异显著影响,而胚性愈伤组织的再生频率不受遗传变异影响,愈伤组织诱导能力与胚性愈伤组织诱导能力及再生频率密切相关,表明不同基因型的胚性愈伤组织诱导和再生频率能够基于愈伤组织诱导能力进行预测[34]。在橡胶树体细胞胚发生方面,前人开展了部分品种的花药体细胞胚发生研究,发现不同品种间的体细胞胚发生能力存在显著差异[17-23]。本研究通过对32个橡胶树品种的花药愈伤组织和体细胞胚诱导情况进行分析,发现橡胶树愈伤组织诱导能力不存在明显的基因型影响,而体细胞胚发生能力存在明显的基因型影响;对于橡胶树不同品种来说,子代的体细胞胚形成率与亲本体细胞胚发生能力相关,父本和母本均影响子代体细胞胚发生能力,至少含一个易于诱导或可诱导亲本,子代才易于或可诱导体胚,表明橡胶树体细胞胚发生能力可遗传。另外本研究发现,已知体胚诱导率的初生代、次生代、三生代和四生代无性系中难诱导体胚发生品种占比依次减少,推测随着易诱导亲本与难诱导亲本的杂交,后代中易诱导体胚发生品种有逐渐增多的趋势。
-
Gawel等将能产生体细胞胚的棉花品种与不能产生体细胞胚的品种进行杂交,分析了后代的体细胞胚发生能力,统计结果表明棉花体细胞胚发生是由多基因参与控制的[35]。叶兴国等选用13个小麦亲本基因型及其F1代进行花药培养,发现小麦花药培养力的遗传控制非常复杂,表现为多基因控制的数量性状遗传[36]。何平等用同一种培养基培养籼粳杂交F1代群体110个株系的花药,结果表明水稻花药培养力是由多基因控制的[37]。张晓玲等对来源于美国、墨西哥和中国的144份不同玉米自交系幼胚胚性愈伤组织的再生能力相关性状进行研究,结果表明玉米幼胚培养性状的遗传变异受多基因调控[38]。本研究发现,具有相同亲本的橡胶树品种体细胞胚诱导率差异显著,且存在超亲现象,表明橡胶树体细胞胚发生能力的遗传控制复杂,推测橡胶树体细胞胚发生也是由多基因参与控制的。
Hargreaves等改进了胚性细胞系的启动和早期增殖方法,在20个控制授粉的辐射松合子胚培养中得到成功应用,显著好于以前结果,表明有可能通过改进培养条件,打破遗传背景的影响[39]。李健英等经过连续再生驯化(SRA)的方法提高了棉花再生和转化效率,并在非胚性愈伤组织时期外施DNA甲基化抑制剂促进了体细胞胚发生[40]。王轲等通过在小麦中过表达再生基因TaWOX5,显著提高了小麦组织培养再生植株和转化效率,促进了质量较差愈伤组织的分化,基本克服了基因型依赖,而且未出现过表达WUS2和BBM等再生基因时出现的影响愈伤组织分化和根系生长或导致畸形植株的弊端,同时该基因在大麦、小黑麦、黑麦、玉米中的再生和转化作用也得到了验证[41]。本研究中多个橡胶树品种的体细胞胚诱导率显著高于前人的研究结果,推断得益于培养基改进。
-
在目前栽培的主要橡胶树品种中,含PB86、RRIM600、热研88-13和海垦1等亲本的后代基本为容易或可诱导体胚的品种类型;含GT1、天任31-45、93-114和RRIM513等亲本的后代体胚诱导率较低;含PR107亲本的后代体胚诱导难易均有,当其后代亲本中不含易或可诱导亲本遗传基础时,后代体胚诱导率低,如母本中含GT1、93-114和RRIM513等亲本的PR107后代体胚诱导率低,当其后代亲本中含有易或可诱导亲本时,后代体胚诱导率高,如母本中含PB86、RRIM600和海垦1等亲本的PR107后代基本为容易或可诱导体胚的品种类型。本研究结果为今后橡胶树体胚发生培养体系的优化指明了方向,可以按亲本进行分组,进一步优化体胚培养条件,如含PB86、RRIM600、热研88-13和海垦1等遗传基础的后代可在现有培养条件的基础上进一步优化,得到适宜培养条件,如含GT1、天任31-45、93-114和RRIM513等遗传基础的后代需在现有培养条件基础上进行大幅度的调整和优化,如从外植体选择与处理、基本培养基筛选、新药品探索、连续再生驯化、过表达再生基因等更多方向开展试验。同时本研究结果可以为橡胶树的传统杂交育种提供参考,以性状优良且体胚诱导率高的橡胶树品种为亲本配置杂交组合,获得高体胚诱导率的优良子代几率大,也为高产高抗无性系优质种苗的工厂化生产和转基因育种研究提供了基础。
-
本研究通过对32个橡胶树品种的遗传关系和体胚发生能力进行比较分析,发现体胚诱导率存在显著的基因型差异,且这种差异可遗传。依据体胚诱导率和遗传关系,将亲本的诱导率同子代的诱导率结合进行分析并分组,为通过基因型分组优化橡胶树多品种体胚发生提供了依据。结果表明,亲本PB86、RRIM600、热研88-13和海垦1及其后代可按本研究所采用的培养条件进行小幅度优化,得到更适合的培养条件;亲本GT1、天任31-45、93-114和RRIM513及其后代需要在现有培养条件基础上进行大幅度的优化,以明确其培养条件。本研究中多个橡胶树品种的体细胞胚诱导率显著高于前人的研究结果。另外,本研究通过对15个品种的体胚苗进行倍性鉴定,证明不同基因型花药体胚苗均来源于体细胞。
不同基因型橡胶树花药体胚发生能力的差异分析
Differential Analysis of Anther Embryogenesis between Different Genotypes of Hevea brasiliensis
-
摘要:
目的 探明不同基因型橡胶树的体胚发生能力差异,为优化橡胶树不同品种体胚发生体系提供理论基础。 方法 以国家橡胶树种质资源圃保存的32个橡胶树主栽品种及其亲本为材料,通过系谱分析明确品种间的遗传关系,并以花药作为外植体诱导愈伤组织并进一步诱导体胚发生,结合体胚诱导率和遗传关系分析不同基因型体胚发生能力的差异。 结果 对32个橡胶树品种进行系谱分析表明:PR107、GT1、PB86、Tjirl、海垦1、天任31-45、合口3-11这7个品种为初生代无性系,其余25个品种中的23个与以上7个初生代无性系中的1 ~ 3个存在亲缘关系,表明我国橡胶树主栽品种亲本来源狭窄。结合系谱和体胚发生结果分析表明:橡胶树品种的体胚发生能力与亲本来源密切相关,亲本体胚发生能力决定其子代品种体胚发生能力。父母本均为难诱导体胚发生品种时,其杂交后代多数难诱导体胚发生;父母本中含1个易诱导体胚发生品种,即使另1个亲本为难诱导体胚发生品种时,其杂交后代多数体胚发生效率高,但这些后代间体胚发生频率差异显著,且存在超亲现象,推测橡胶树体胚发生是由多基因参与控制的。GT1、天任31-45、93-114和RRIM513等为难诱导体胚发生品种,以其为亲本来源的杂交后代多数难以诱导体胚发生(如云研77-2、云研77-4、云研73-46、保亭1-285、湛试32713、热研217),而PB86、RRIM600、热研88-13和海垦1等为易于诱导体胚发生品种,以其为亲本之一的杂交后代多数体胚发生效率高(如大丰95、海垦2、热研917、热研73397、热研879、热研918、徐育3、徐育141-2)。 结论 本研究发现,橡胶树体胚发生能力存在明显的基因型差异,且这种差异可遗传。依据体胚诱导率和遗传关系,本文将橡胶树亲本的体胚形成率同子代的诱导率结合进行分析并分组,为利用基因型分组优化橡胶树栽培品种的体胚发生体系提供了依据。 Abstract:Objective Clarifying the effect of genotype on rubber tree somatic embryogenesis and combining with pedigree analysis to classify and group will provide a theoretical basis for accurately optimizing the somatic embryogenesis system of rubber trees. Method The genetic relationship between the 32 main rubber tree cultivars and their parents preserved in China National Rubber Tree Germplasm Repository was clarified through pedigree analysis. The two mediums with high somatic embryo induction rate in multiple cultivars in the previous experiment were used to induce callus and further induce somatic embryogenesis with anthers as explants. Finally, the induction rate of somatic embryos based on genetic relationship was analyzed. Result The pedigree analysis of 32 rubber tree cultivars showed that 7 cultivars were primary clones including PR107, GT1, PB86, Tjirl, Haiken1, Tianren31-45, and Hekou3-11. Twenty-three of the remaining 25 cultivars are genetically related to 1-3 of the above 7 cultivars. It is clear that the source of parents of rubber tree cultivars in China is narrow. Combining pedigree and somatic embryogenesis analysis showed that the somatic embryogenesis ability of rubber tree cultivars is closely related to the parental source, and the parental somatic embryogenesis ability determines the somatic embryogenesis ability of its offspring cultivars. When both parents are recalcitrant to somatic embryogenesis, most of their hybrid progeny are recalcitrant. When one of the parents has high somatic embryogenesis efficiency, even if the other parent is recalcitrant, most of its hybrid offspring have high somatic embryogenesis efficiency. However, the frequency of somatic embryogenesis varies significantly among these offspring, and there is a phenomenon of super-parental, which indicates that the rubber tree somatic embryogenesis is controlled by the participation of multiple genes. GT1, Tianren31-45, 93-114, RRIM513, and other parent-derived cultivars have low somatic embryo induction rate (e.g., Yunyan77-2, Yunyan77-4, Yunyan73-46, Baoting1-285, Zhanshi32713, and Reyan217), and PB86, RRIM600, Reyan88-13, and, Haiken1 and other parent-derived cultivars have high somatic embryo induction rate (e.g., Dafeng95, Haiken2, Reyan917, Reyan73397, Reyan879, Reyan918, Xuyu3, and Xuyu141-2). Conclusion There are obvious genotypic differences in rubber tree somatic embryogenesis, and this difference can be heritable. According to the somatic embryo induction efficiency and genetic relationship, analyzing and grouping 32 main rubber tree cultivars and their parents provide a way to optimize the somatic embryogenesis of rubber tree cultivars by genotype grouping. -
Key words:
- rubber tree
- / genotype
- / inheritance
- / parent
- / pedigree analysis
- / somatic embryogenesis
- / ploidy
-
图 1 橡胶树系谱和花药体胚发生能力差异分析
Figure 1. Pedigree analysis of rubber tree and analysis of differences in anther embryogenic ability
图 2 橡胶树花药体胚苗的诱导过程图
Figure 2. The Induction process of anther somatic embryo plantlet of rubber tree
表 1 不同橡胶树品种花药愈伤组织、体胚、子叶形体胚诱导率
Table 1. Induction rate of anther callus, somatic embryo, cotyledon somatic embryo of different rubber tree cultivars
品种世代分级
Cultivar classification品种
Cultivar愈伤组织形成率/%
Callus induction
rate体胚形成率/%
Somatic embryo
induction rate子叶胚形成率/%
Cotyledon somatic
embryo induction rate体胚诱导能力分级
Somatic embryo
induction ability
classificationM培养基
M mediumS培养基
S mediumM培养基
M mediumS培养基
S mediumM培养基
M mediumS培养基
S medium初生代无性系
First--generation clonePR107 (82.2±16.8) (61.9±21.8) (4.2±3.3) i (3±3) g (0) e (0) c 难 Recalcitrant GT1 (75±8.1) (75.8±16.2) (0) i (0.3±0.6) g (0) e (0) c 难 Recalcitrant PB86 (66.5±11.7) (55.5±25.1) (26.8±13.8) efgh (19.8±9) defg (0) e (0) c 易 High Tjirl (59.2±15) (58.9±19.3) (0) i (0) g (0) e (0) c 难 Recalcitrant 海垦1 (73.7±2.1) (50.8±9.6) (31.8±3.7) cdef (7.9±2.1) g (4.1±1.5) de (0) c 易 High 天任31-45 (35.9±22.4) (28.9±18.6) (0) i (0) g (0) e (0) c 难 Recalcitrant 合口3-11 (68.3±31) (74.6±26.2) (10.7±3.8) hi (11±3) fg (0) e (0) c 可 Inducible 次生代无性系
Second-generation clone云研77-2 (40.9±10.9) (34.4±19.5) (0) i (0) g (0) e (0) c 难 Recalcitrant 云研77-4 (57.2±23.2) (46.7±19.6) (0) i (0) g (0) e (0) c 难 Recalcitrant 云研73-46 (71.8±13) (73.7±8.6) (6.7±1.4) i (0) g (0) e (0) c 难 Recalcitrant 保亭1-285 (68.9±9.8) (59.6±22.9) (0) i (0) g (0) e (0) c 难 Recalcitrant 93-114 (77.5±6.6) (66±13.5) (0) i (0) g (0) e (0) c 难 Recalcitrant RRIM513 (74.7±1.4) (76.3±21.1) (0) i (0) g (0) e (0) c 难 Recalcitrant IAN873 (74.2±18.5) (73.5±14.1) (0) i (0) g (0) e (0) c 难 Recalcitrant 大丰95 (55.5±27.2) (37.9±15.4) (13.5±4.1) ghi (30.9±34.3) cdef (1.7±2.9) de (6±10.3) b 易 High RRIM600 (77.8±19.2) (76.9±20.7) (13.2±13.8) ghi (11±10.1) fg (0) e (2.5±4.3) bc 可 Inducible 徐育141-2 (68.9±33.2) (61.3±43.7) (48.1±34.2) bc (47.2±39.1) bc (13.3±10.4) b (3.8±1.1) bc 易 High 文昌217 (87.6±6.9) (92.4±10.9) (17.6±1.2) fghi (16±2.6) defg (0) e (0) c 可 Inducible 三生代无性系
Third-generation clone湛试32713 (69.2±18.8) (56.7±10.3) (0.7±1.2) i (0) g (0) e (0) c 难 Recalcitrant 大丰117 (51.2±3.1) (43.3±8.4) (15.6±0.7) fghi (0) g (3.2±0.3) de (0) c 可 Inducible 热研217 (70±30) (49.3±33.4) (0) i (0) g (0) e (0) c 难 Recalcitrant 文昌11 (94.4±5.9) (89.8±10) (11.5±2.2) hi (6±1.7) g (0.6±1) e (0) c 可 Inducible 热研917 (86±5.2) (73.1±17) (47.6±19) bc (35.7±24.4) bcd (0.6±1) e (0) c 易 High 热研73397 (85.3±2.7) (79.1±6.7) (95±5) a (94±10.4) a (23.1±8.2) a (17.3±5.1) a 易 High 徐育3 (79±20.1) (74.8±22.3) (46.7±2.6) bcd (56±1.7) b (6.6±0.9) cd (5.6±1) b 易 High 保亭3412 (94.1±5.2) (94.3±6.1) (4.3±1.5) i (3.7±1.4) g (0) e (0) c 难 Recalcitrant 热研88-13 (90.1±7.9) (81.3±7) (83.5±11.7) a (77.4±19.9) a (11.5±6) bc (3.8±3.9) bc 易 High PB235 (46.9±12) (53.7±13.4) (35.5±0.8) cde (37.1±1.1) bcd (2.9±2.9) de (6±0.3) b 易 High 四生代无性系
Fourth-generation clone热垦628 (87.4±4.6) (80.8±20.1) (4.4±4.2) i (12.4±9.7) efg (0) e (0) c 可 Inducible 热垦525 (79.4±10.3) (67.2±11.1) (56.9±19) b (35±18.2) bcde (7.2±4.6) cd (6.1±5.4) b 易 High 热研879 (90.9±10.1) (78.6±16.6) (31±20.7) cdefg (32.5±15) cdef (3.7±6.4) de (0) c 易 High 热研918 (96.5±3.1) (79±22) (29.7±10.4) defg (36.1±17.7) bcd (7.5±5) cd (3.5±4.3) bc 易 High 注:括号内加减号前数字代表三次重复的均值,加减号后数字代表标准差。次生代无性系海垦2愈伤组织形成率达96.1%,体胚形成率 (45.0±3.5) %,属于易诱导品种,数据来自文献[5,17]。
Notes: In parentheses, the numbers before the plus and minus signs represent the mean of three repetitions, and the numbers after the plus and minus signs represent the standard deviation. The callus induction rate and the somatic embryo induction rate of the secondary-generation clone Haiken 2 were 96.1% and (45.0±3.5) %, respectively, which belonged to high frequency embryogenic cultivar, the data came from the literatures[5,17]. 下载: 导出CSV
下载: 导出CSV
表 2 橡胶树不同基因型体胚苗倍性检测结果
Table 2. Ploidy test results of somatic embryo plants of different genotypes of rubber tree
品种
Cultivar母本倍性
Mother
plant
ploidy检测体胚苗株数
Number of somatic
embryo plants tested二倍体株数
Number of
diploid plants三倍体株数
Number of
triploid plants热垦525 二倍体 113 113 — 徐育141-2 二倍体 100 100 — 热研73397 二倍体 86 86 — 热研917 二倍体 50 50 — 热研88-13 二倍体 50 50 — 海垦2 二倍体 50 50 — PR107 二倍体 34 34 — 文昌217 二倍体 25 25 — 文昌11 二倍体 16 16 — 热研918 二倍体 14 14 — 云研73-46 三倍体 8 — 8 合口3-11 二倍体 3 3 — 海垦1 二倍体 1 1 — 云研774 三倍体 1 — 1 保亭1-285 二倍体 1 1 —
下载: 导出CSV
-
[1] 王泽云, 陈雄庭, 吴胡蝶. 橡胶树新型种植材料—体胚植株[J]. 热带农业科学, 2001, 6(94):11-15. [2] 陈雄庭, 王泽云, 吴胡蝶, 等. 橡胶树新种植材料—自根幼态无性系[J]. 热带作物学报, 2002, 23(1):19-23. doi: 10.3969/j.issn.1000-2561.2002.01.004 [3] Carron M P, Lardet L, Leconte A, et. al. Field trials network emphasizes the improvement of growth and yield through micropropagation in rubber tree (Hevea brasiliensis, Muëll.-Arg. )[J]. Acta Hort, 2009(812): 485-492. [4] Eline M, Stefaan W. Somatic embryogenesis as key technology for shaping the rubber tree of the future[J]. Frontiers in Plant Science, 2018, 9: 1-6. doi: 10.3389/fpls.2018.00001 [5] 王泽云, 曾宪松, 陈传琴, 等. 用离体花药诱导巴西橡胶植株的研究[J]. 热带作物学报, 1980, 1(1):16-26. [6] Carron M P, Etienne H, Michaux-Ferriere N, et al. Somatic embryogenesis in rubber tree (Hevea brasiliensis Müell. Arg. )[J]. Biotechnol Agric For, 1995, 30: 353-369. [7] Jayasree P K, Asokan M P, Sobha S, et al. Somatic embryogenesis and plant regeneration from immature stamens of Hevea brasiliensis (Muell. ) Arg[J]. Curr Sci, 1999, 76: 1242-1245. [8] Montoro P, Carron M P, Granet F, et al. Development of new varietal types based on rejuvenation by somatic embryogenesis and propagation by conventional budding or microcutting in Hevea brasiliensis[C]//D Geelen. 7th IS on In Vitro Culture and Horticultural Breeding. Argentina: Acta Hort, 2012, 961: 553-576. [9] 王泽云, 吴胡叶, 曾宪松, 等. 巴西橡胶花药胚的发生和花药植株起源的研究[J]. 热带作物学报, 1984, 5(1):9-13. [10] 王亚丽. 巴西橡胶树体外体细胞胚发育的细胞学和组织学研究[D]. 海南: 华南热带农业大学, 2004: 36-37. [11] 王天地, 华玉伟, 黄华孙. 橡胶树次生体胚植株染色体数目和DNA含量分析研究[J]. 热带农业科学, 2013, 33(5):33-37. doi: 10.3969/j.issn.1009-2196.2013.05.009 [12] Srichuay W, Kalawong S, Sirisom Y. Callus induction and somatic embryogenesis from anther cultures of Hevea brasiliensis (Muell. Arg. )[J]. Kasetsart J Nat Sci, 2014, 48: 364-375. [13] Wang T D, Huang T D, Huang H S, et al. Origin of secondary somatic embryos and genetic stability of the regenerated plants in Hevea brasiliensis[J]. Journal of Rubber Research, 2017, 20(2): 101-116. doi: 10.1007/BF03449145 [14] 华玉伟, 黄华孙, 黄天带, 等. 利用胚状体快速繁殖巴西橡胶树自根幼态无性系的方法[P]. 发明专利, 中国, 200710184892.5, 1-11. [15] Hua Y W, Huang T D, Huang H S. Micropropagation of self-rooting juvenile clones by secondary somatic embryogenesis in Hevea brasiliensis[J]. Plant Breeding, 2010, 129(2): 202-207. doi: 10.1111/j.1439-0523.2009.01663.x [16] 顾晓川, 徐正伟, 成 镜, 等. 植物凝胶和蔗糖对橡胶树体胚植株再生的影响[J]. 广西植物, 2018, 38(9):1164-1171. doi: 10.11931/guihaia.gxzw201712033 [17] 谭德冠. 巴西橡胶树体胚发生的改良及乳管分化研究[D]. 海口: 海南大学, 2011: 34. [18] 李 玲, 管 艳, 梁国平, 等. TDZ对橡胶树花药愈伤组织诱导和体细胞胚发生的影响[J]. 中国农学通报, 2014, 30(1):20-25. doi: 10.11924/j.issn.1000-6850.2013-2136 [19] 黄凤翔, 管 艳, 桂明春, 等. 正交实验法优选橡胶树花药愈伤组织诱导的研究[J]. 北方园艺, 2014(18):109-111. [20] 管 艳, 李 玲, 梁国平, 等. 不同品种橡胶树花药愈伤组织诱导、分化及植株再生的比较[J]. 中国农学通报, 2015, 31(4):40-44. doi: 10.11924/j.issn.1000-6850.2014-1970 [21] Nor Mayati C H. Effects of zeatin and kinetin on in vitro regeneration of Hevea brasiliensis RRIM 2025[J]. J Rubb Res, 2015, 18(3): 127-138. [22] 张源源, 王祥军, 高新生, 等. 不同品系橡胶树花药愈伤诱导和体胚发生研究[J]. 中国农学通报, 2018, 34(3):70-75. doi: 10.11924/j.issn.1000-6850.casb17070056 [23] 桂明春, 李 玲, 管 艳, 等. MS营养成分对橡胶树花药愈伤组织诱导效果的评价[J]. 植物生理学报, 2019, 55(8):1239-1246. [24] Satyavathi V V, Manga V, Rao M V S, et al. Genetic analysis of reciprocal differences in the inheritance of in vitro characters in pearl millet[J]. Genetics and molecular biology, 2016, 39: 54-61. doi: 10.1590/1678-4685-GMB-2014-0380 [25] Holtz C T, Tull A R, Merkle S A. Influence of species and hybrid status on induction of somatic embryogenesis in Castanea[J]. Canadian Journal of Forest Research, 2017, 47(3): 382-388. doi: 10.1139/cjfr-2016-0362 [26] Ruibin S U N, Ruiping T, Dan M A, et al. Comparative transcriptome study provides insights into acquisition of embryogenic ability in upland cotton during somatic embryogenesis[J]. Journal of Cotton Research, 2018, 1(1): 1-13. doi: 10.1186/s42397-018-0004-z [27] Fan Y, Yu X, Guo H, et al. Dynamic Transcriptome Analysis Reveals Uncharacterized Complex Regulatory Pathway Underlying Dose IBA-Induced Embryogenic Redifferentiation in Cotton[J]. International journal of molecular sciences, 2020, 21(2): 426. doi: 10.3390/ijms21020426 [28] 黄华孙. 中国橡胶树育种五十年[M]. 北京: 中国农业出版社, 2005: 174-190. [29] 赵建文. 我国育成橡胶树品种的遗传多样性分析及骨干亲本选择[D]. 海口: 海南大学, 2013: 15-18. [30] 高新生, 黄华孙, 李维国. 中国天然橡胶主要品种及系谱(1996-2020年)[M]. 北京: 中国农业出版社, 2021. [31] Hodges T K, Kamo K K, Imbrie C W, et al. Genotype specificity of somatic embryogenesis and regeneration in maize[J]. Bio/technology, 1986, 4(3): 219-223. [32] Parrott W A, Williams E G, Hildebrand D F, et al. Effect of genotype on somatic embryogenesis from immature cotyledons of soybean[J]. Plant Cell, Tissue and Organ Culture, 1989, 16(1): 15-21. doi: 10.1007/BF00044068 [33] Niskanen A M, Lu J, Seitz S, et al. Effect of parent genotype on somatic embryogenesis in Scots pine (Pinus sylvestris)[J]. Tree Physiology, 2004, 24(11): 1259-1265. doi: 10.1093/treephys/24.11.1259 [34] Li C, Li X, Lin X, et al. Genotypic variation in the response to embryogenic callus induction and regeneration in Saccharum spontaneum[J]. Plant Genetic Resources, 2021, 19(2): 153-158. doi: 10.1017/S1479262121000198 [35] Gawel N J, Robacker C D. Genetic control of somatic embryogenesis in cotton petiole callus cultures[J]. Euphytica, 1990, 49(3): 249-253. doi: 10.1007/BF00036296 [36] 叶兴国, 徐惠君, 徐琼芳, 等. 小麦花药培养力的基因型差异和配合力分析[J]. 中国农业科学, 1996, 30(6):49-54. [37] 何 平, 沈利爽, 陆朝福, 等. 水稻花药培养力的遗传分析及基因定位[J]. 遗传学报, 1998, 25(4):337-344. [38] 张晓玲, 龙 芸, 葛 飞, 等. 玉米幼胚胚性愈伤组织再生能力相关性状遗传研究[J]. 遗传, 2017, 39(2):143-155. [39] Hargreaves C L, Reeves C B, Gough K, et al. Overcoming the challenges of family and genotype representation and early cell line proliferation in somatic embryogenesis from control-pollinated seeds of Pinus radiata[J]. New Zealand Journal of Forestry Science (New Zealand Forest Research Institute Ltd (trading as Scion)), 2011, 41(7): 97-114. [40] Li J, Wang M, Li Y, et al. Multi-omics analyses reveal epigenomics basis for cotton somatic embryogenesis through successive regeneration acclimation process[J]. Plant biotechnology journal, 2019, 17(2): 435-450. doi: 10.1111/pbi.12988 [41] Wang K, Shi L, Liang X, et al. The gene T aWOX5 overcomes genotype dependency in wheat genetic transformation[J]. Nature Plants, 2022, 8: 110-117. -

 点击查看大图
点击查看大图
图(3) / 表(2)
计量
- 文章访问数: 4042
- HTML全文浏览量: 2001
- PDF下载量: 50
- 被引次数: 0
