











-
林木生长既是对光照和气温等环境条件的适应性表现[1],也是用材树种育种的基本目标性状[2]。林木为多年生植物,其生长过程有自身的节律,并受环境条件的影响[1,3]。目前,已在多类树种中报道了生长性状的遗传力和遗传相关等遗传参数的研究,如桉属(Eucalyptus L'Hérit.)[4-23]和松属(Pinus Linn.)[24-25]树种,但多数研究仅分析了单个年份或早期的生长性状。因此,需要加强林木多年生生长的遗传分析,这有助于加深对林木生长动态遗传基础的理解,也将为早期选择提供有效指导。
尾叶桉(E. urophylla S. T. Blake)和细叶桉(E. tereticornis Smith)均为桉属双蒴盖亚属(Symphyomyrtus (Schauer) Brooker)树种[26],前者天然分布于东帝汶和印度尼西亚的一些岛屿(7°23'~10° S),后者分布于巴布亚新几内亚的南部至澳大利亚的维多利亚州南部沿海的狭长区域(6°~38° S)[27-28]。两树种亲缘关系较近,其杂种常用于遗传参数的估算[9-10, 16, 20]。尾叶桉 × 细叶桉杂种具有较好的速生、高产、抗风和耐寒潜力[19, 29-30],已较广泛地用于华南地区桉树人工林的营建。
本研究对尾叶桉 × 细叶桉不完全析因交配产生的59个杂种组合13、16年生的树高、胸径和材积进行了狭义遗传力估算,并结合前期生长和材性[9]进行了遗传相关分析,以期进一步了解桉属树种生长动态的遗传规律,并为大径材良种培育的早期选择年龄的确定提供可靠依据。
-
参试材料为10株尾叶桉母本与10株细叶桉父本不完全析因交配产生的62个杂种组合,大田试验最终保存59个组合[9, 20];参试材料也包括10个母本的自由授粉家系。2003年7月在福建省漳州市长泰县岩溪林场美山工区(117°52′ E, 24°46′ N)营建试验林,试验地概况见文献[20]。试验设计为随机完全区组,4株单行小区,4次重复,株行距为2 m × 3 m。
-
生长性状包括树高(H)、胸径(D)和材积(V)。利用Vertex III激光超声波测高测距仪(瑞典Haglof公司)测量13、16年生的树高(分别缩写为H13和H16),利用胸径尺测量13、16年生的胸径(D13和D16)。前期生长包括0.5、1.5、2.5、4、7.5、10年生的树高(依次缩写为H0.5、H1.5、H2.5、H4、H7.5、H10)和1.5、2.5、4、7.5、10年生的胸径(依次为D1.5、D2.5、D4、D7.5、D10)及1.5、2.5、4、7.5、10、13、16年生的材积(依次为V1.5、V2.5、V4、V7.5、V10、V13、V16),按文献[16]的公式计算:
$ V=H \times D^{2}/30 \;000 $
10年生材性的测定见文献[9]。基于之前建立的预测模型[10],利用近红外光谱仪MPA (德国Bruker)测定10年生木材基本密度(BD10)、纤维素含量(CC10)、半纤维素含量(HC10)、木质素含量(LC10)和紫丁香基木质素/愈创木基木质素比例(S/G10)。
-
利用ASRmel程序包[31]在R软件上进行数据统计分析。对13、16年生树高、胸径和材积及杂种组合与自由授粉家系间的差异显著性通过t检验进行检测。方差分析和方差分量分析采用混合线性模型:
$ y=X b + Z u + e$ ,式中:y为单株性状值的向量,b为固定效应的向量(本分析中为重复的效应)、X为b的关联矩阵,u为随机效应的向量(包括母本、父本和母本 × 父本互作的效应)、Z为u的关联矩阵,e为随机误差的向量。采用程序包的似然比法进行随机效应的方差分量显著性检验。基于有关的方差分量计算单株狭义遗传力(
$ h_{s}^{2}$ )和显性遗传效应($ d_{s}^{2}$ ),公式如下:$ h_{s}^{2}=\frac{2(\sigma_{f}^{2} + \sigma_{m}^{2})}{\sigma_{f}^{2} + \sigma_{m}^{2} + \sigma_{f m}^{2} + \sigma_{e}^{2}} $
$ d_{s}^{2}=\frac{4 \sigma_{f m}^{2}}{\sigma_{f}^{2} + \sigma_{m}^{2} + \sigma_{f m}^{2} + \sigma_{e}^{2}} $
式中:
$ \sigma_{f}^{2}$ 、$ \sigma_{m}^{2}$ 、$ \sigma_{fm}^{2}$ 和$ \sigma_{e}^{2}$ 分别为母本效应、父本效应、母本 × 父本互作效应和随机误差效应的方差分量。标准误利用pin函数估算。生长性状不同林龄间、生长与材性性状间的表型相关系数和加性遗传相关系数估算公式如下:
$ r_{p}=\frac{\sigma_{p(x, y)}}{\sqrt{\sigma_{p x}^{2} \sigma_{p y}^{2}}} $
$ r_{g}=\frac{\sigma_{g(x, y)}}{\sqrt{\sigma_{g x}^{2} \sigma_{g y}^{2}}} $
式中:
$\sigma_{p(x, y)}$ 和$ \sigma_{g(x, y)}$ 分别为林龄x与y的生长性状或者性状x与y的表型和加性协方差,$ \sigma_{p x}^{2}$ 和$ \sigma_{p y}^{2}$ 分别为x和y的表型方差分量,$ \sigma_{g x}^{2}$ 和$ \sigma_{g y}^{2}$ 分别为x和y的加性遗传方差分量。相关显著性水平通过相关系数值与标准误(SE)的比较进行判定[32],即:≥1.45 × SE (P≤0.05)、≥2.326 × SE (P≤0.01)和 ≥3.09 × SE (P≤0.001)。基于主成分回归进行16年生(因变量Y)与早期生长(树高、胸径和材积)的回归分析,模型如下:
$ Y=b_{0} + b_{1} X_{1} + b_{2} X_{2} + b_{3} X_{3} + b_{4} X_{4} + b_{5} X_{5} + b_{6} X_{6} + f $
式中:
$ b_{0}$ 为常数,b1 ~ b6为回归系数,X1 ~ X6依次为1.5、2.5、4、7.5、10、13年生生长经主成分变换后的自变量矩阵,f为误差。 -
尾叶桉 × 细叶桉59个杂种组合13、16年生平均树高、胸径和材积均高于母本自由授粉家系(表1),表现出一定的杂种优势,尤其是13年生平均树高及13、16年生平均材积显著或极显著高于母本自由授粉家系(P≤0.05或0.001)。杂种组合的树高变异系数较小,但材积变异系数较大。
参照大径材(胸径≥28 cm)的标准[33],只16年生时1个杂种组合(UX-07 × T2508,11株)的平均胸径达28.34 cm,13、16年生时分别有来自21和28个杂种组合的29和47株达到大径材的规格(表1),这为进一步培育大径材良种(包括无性系)提供了备选材料。
表 1 尾叶桉 × 细叶桉杂种组合及母本自由授粉(OP)家系的生长及其变异系数(CV)
Table 1. Growth and coefficient of variation (CV) of Eucalyptus urophylla × E. tereticornis crosses and maternal open-pollinated (OP) families
生长性状
Growth trait杂种组合
Inter-specific cross自由授粉家系
OP family均值
Mean变异系数
CV/%平均胸径≥28 cm杂种组合数
The number of crosses≥28 cm in D13 or D16胸径≥28 cm单株数
The number of trees≥28 cm in D13 or D16均值
Mean变异系数
CV/%13年生 树高 H13/m 17.69* 26.8 16.60 28.5 胸径 D13/cm 15.89 43.5 0 29 14.58 41.2 材积 V13/m3 0.250*** 81.7 0.154 82.4 16年生 树高 H16/m 21.89 28.6 20.41 31.3 胸径 D16/cm 17.68 41.8 1 47 16.61 37.6 材积 V16/m3 0.377* 81.1 0.293 86.2 注:* P ≤ 0.05、*** P ≤ 0.001表示杂种组合与母本自由授粉家系的差异显著性水平分别达0.05、0.001。 Notes: * P ≤ 0.05, *** P ≤ 0.001 for the significant difference at 0.05 and 0.001, respectively, between inter-specific crosses and maternal OP families. -
表2表明:母本方差分量对13、16年生树高、胸径和材积均影响极显著(P≤0.001或0.01);父本方差分量只对13、16年生材积影响极显著(P≤0.01),对树高和胸径均影响不显著(尤其是树高的方差分量接近0);母本 × 父本互作的方差分量仅对13年生树高影响显著(P≤0.05)。这表明母本效应显著影响杂种组合的子代生长,而父本效应仅对材积性状影响显著,母本对子代生长的影响明显大于父本;同时,母本 × 父本互作效应仅对某些年龄的树高有显著影响,且随着年龄增长呈减弱趋势。
表 2 尾叶桉 × 细叶桉生长的母本(
${\color[RGB]{27,29,113}\sigma _f^2 }$ )、父本(${\color[RGB]{27,29,113} \sigma _m^2} $ )和母本 × 父本(${\color[RGB]{27,29,113} \sigma _{fm}^2 }$ )的方差分量及单株狭义遗传力(${\color[RGB]{27,29,113} h _s^2 }$ )和显性遗传效应(${\color[RGB]{27,29,113} d_s^2}$ )Table 2. Female (
${\color[RGB]{27,29,113} \sigma _f^2} $ ), male (${\color[RGB]{27,29,113} \sigma _m^2} $ ) and female × male (${\color[RGB]{27,29,113} \sigma _{fm}^2 } $ ) variance components as well as single-tree narrow sense heritability (${\color[RGB]{27,29,113} h _s^2} $ ) and dominance ratio of phenotypic variance (${\color[RGB]{27,29,113} d_s^2 }$ ) for growth of E. urophylla × E. tereticornis crosses生长性状
Growth trait方差分量 Variance component (±SE) 单株狭义遗传力
$h _s^2 $(±SE)显性遗传效应
$d _s^2 $(±SE)母本 $\sigma _f^2 $ 父本 $\sigma _m^2 $ 母本 × 父本 $\sigma _{fm}^2 $ 13年生 树高 H13 1.79 (±1.12)*** 1.84 (±0.12) × 10−6 0.85 (±0.60)* 0.17 (±0.10) 0.16 (±0.11) 胸径 D13 3.91 (±2.28)*** 0.52 (±0.63) 3.79 (±0.24) × 10−6 0.20 (±0.10) 3.37 (±0.18) × 10−7 材积 V13 2.91 (±1.78) × 10−3*** 1.17 (±0.93) × 10−3** 3.88 (±0.27) × 10−8 0.21 (±0.09) 3.91 (±0.20) × 10−5 16年生 树高 H16 3.17 (±2.03)*** 3.34 (±0.25) × 10−6 1.14 (±1.20) 0.17 (±0.10) 0.12 (±0.13) 胸径 D16 4.54 (±2.77)*** 1.09 (±1.10) 0.57 (±1.41) 0.22 (±0.11) 0.04 (±0.11) 材积 V16 4.96 (±3.72) × 10−3** 5.65 (±4.11) × 10−3** 2.58 (±3.41) × 10−3 0.23 (±0.11) 0.11 (±0.15) 注:SE,标准误;* P≤0.05,**P≤0.01,*** P≤0.001。下同。
Notes:SE,Standard error;* P ≤0.05,** P ≤0.01,*** P ≤0.001.The same below.13、16年生树高、胸径和材积的单株狭义遗传力(
$h^2_s $ )为0.17~0.23,均属中等偏低的水平,材积的$h_s^2 $ 稍高于树高和胸径,表明材积受相对较高的加性遗传控制。与母本 × 父本互作效应多不显著相对应,显性遗传效应($d^2_s $ )为3.37 × 10−7~0.16,均处于较低或极低的水平,表明显性效应对较晚年龄生长的影响普遍低于加性效应。这与早期生长的遗传效应有异,如0.5、1.5、2.5、4年生树高和胸径的$h_s^2 $ 为0.08~0.13,$d_s^2 $ 为0.11~017,加性效应均小于显性效应(除2.5年生胸径)[9],表明加性效应和显性效应在早晚年龄的相对重要性不同。 -
除1.5年生材积外,16年生树高、胸径和材积与之前年龄的相应生长性状的表型相关和加性遗传相关均达0.01或0.001的显著水平(图1),且加性遗传相关系数均大于对应的表型相关系数,这表明遗传相关是影响表型相关的重要因素。在2.5年生时加性遗传相关即较稳定,表明对16年生生长性状的早期选择可早至2.5年生时进行。
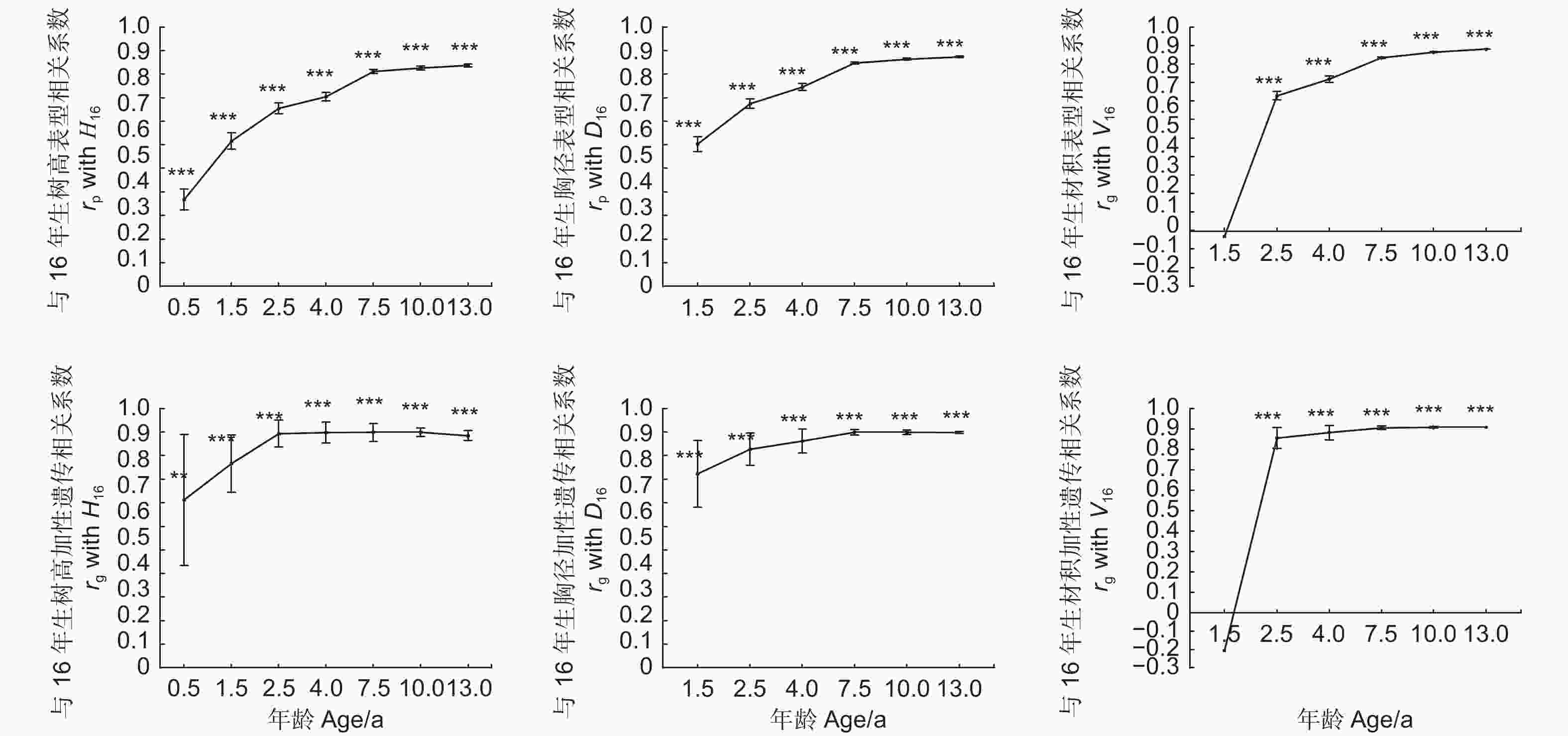
图 1 16年生树高(H16),胸径(D16)和材积(V16)与之前年龄相应生长的表型相关和加性遗传相关
Figure 1. Phenotypic and additive genetic correlations of 16-year-old height (H16), breast-high diameter (D16) and volume (V16) with earlier height, breast-high diameter and volume, respectively
13、16年生生长与10年生材性性状的表型相关系数和加性遗传相关系数见表3。无论是树高、胸径还是材积,13年生生长与材性的相关显著性与16年生类似(树高与木质素含量(LC10)的加性遗传相关除外)。13、16年生生长与10年生基本密度(BD10)均呈显著正相关(P≤0.01、0.05或0.001),而与LC10均呈显著负相关(P≤0.05或0.01,H16的rg除外),表明13、16年生时对生长的选择可间接提高木材基本密度,并降低木质素含量;13、16年生生长与10年生紫丁香基木质素/愈创木基木质素比例(S/G10)均呈显著的表型正相关(P≤0.01或0.001),胸径和材积与纤维素含量(CC10)均呈显著的表型负相关(P≤0.05或0.01),但加性遗传相关均不显著,3个生长性状与半纤维素含量(HC10)的表型相关和加性遗传相关均不显著。
表 3 13、16年生生长与10年生材性的表型相关(rp)和加性遗传相关(rg)
Table 3. Phenotypic (rp) and additive genetic (rg) correlations of 13- and 16-year-old growth with 10-year-old wood properties
生长性状
Growth trait相关系数
rp or rg材性性状 Wood property trait (±SE) BD10 CC10 HC10 LC10 S/G10 H13 rp 0.630 (±0.035)** −0.072 (±0.062) 0.007 (±0.059) −0.102 (±0.059)* 0.204 (±0.064)** rg 0.606 (±0.225)** 0.072 (±0.337) 0.108 (±0.416) −0.505 (±0.307)* 0.061 (±0.311) D13 rp 0.633 (±0.035)** −0.126(±0.062)* −0.001 (±0.056) −0.137 (±0.056)** 0.243 (±0.064)** rg 0.537 (±0.252)* 0.082 (±0.333) 0.043 (±0.435) −0.604 (±0.298)* 0.024 (±0.238) V13 rp 0.550 (±0.040)*** −0.162 (±0.061)** 0.015 (±0.053) −0.151 (±0.054)** 0.244 (±0.066) *** rg 0.472 (±0.261)* 0.035 (±0.345) −0.142 (±0.363) −0.508 (±0.306)* −0.147 (±0.315) H16 rp 0.618 (±0.038)** −0.005 (±0.067) 0.016 (±0.062) −0.097 (±0.061)* 0.208 (±0.054)** rg 0.493 (±0.257)* 0.001 (±0.346) −0.007 (±0.424) −0.427 (±0.334) −0.043 (±0.314) D16 rp 0.630 (±0.036)** −0.135 (±0.065)* −0.019 (±0.059) −0.144 (±0.058)** 0.222 (±0.067) ** rg 0.474 (±0.261)* 0.093 (±0.360) 0.012 (±0.414) −0.559 (±0.299)* −0.089 (±0.315) V16 rp 0.560 (±0.043)*** −0.131 (±0.068)* −0.030 (±0.056) −0.152 (±0.054) ** 0.238 (±0.071)*** rg 0.568 (±0.227)** −0.123 (±0.327) −0.152 (±0.325) −0.500 (±0.264)* −0.057 (±0.313) -
16年生与早期树高、胸径和材积回归分析的决定系数分别为0.842、0.942、0.960,第1主成分可分别解释83.0%、91.3%、93.7%的表型变异(表4),其中,2.5年生树高、胸径和材积可分别解释16年生时的73.9%、71.9%、66.2%的表型变异,表明2.5年时进行早期选择是可行的。
表 4 16年生与早期生长回归分析的第1主成分
Table 4. PC1 of principal components regression of 16-year-old growth with earlier growth traits
年龄
Age/a第1主成分 PC1/% 树高 H 胸径 D 材积 V 1.5 49.0 52.2 45.1 2.5 73.9 71.9 66.2 4 81.5 86.8 78.4 7.5 97.0 98.6 96.4 10 98.3 99.1 99.5 13 97.1 97.3 99.2 模型 Model 83.0 91.3 93.7 -
尾叶桉 × 细叶桉早期生长的杂种优势已有报道[20, 29-30]。本研究中,杂种组合13、16年生的平均生长均好于母本自由授粉家系,表明杂种优势持续存在,其中,1个杂种组合16年生的胸径达到了大径材的规格,后续可利用其亲本建立种子园,生产(母本的)优良种子用于营林实践,从而实现杂种优势的遗传增益。尾叶桉 × 细叶桉易于无性快繁[34],杂种组合的优良单株经无性系测试后亦可大规模地快速推广。
尾叶桉母本的效应显著影响杂种13、16年生生长,这与7.5年生[20, 30]和10年生[9]的研究结果一致,表明母本对尾叶桉 × 细叶桉整个生长期均存在显著的遗传效应。显著的母本效应也存在于尾叶桉 × 赤桉(E. camaldulensis Dehnh.)5年生[12]和8.3年生[22]生长性状中。相反,父本效应对13、16年生树高和胸径的影响不显著,这与10年生生长[9]的研究结果一致,但异于7.5年生生长[20, 30]的研究结果,表明父本效应可能随着年龄增长而趋于减弱。另一方面,父本效应对材积的影响显著。可见,杂交育种中需要重视杂交力高的优良母本和父本的选择。此外,母本 × 父本互作效应对13、16年生生长(除H13外)的影响均不显著,这与7.5年生[20, 30]和10年生树高、胸径[9]的研究结果相似,但与对1.5年生和2.5年生的显著影响[20]相反,表明母本 × 父本互作效应随年龄减弱,且程度可能比父本效应更大。当然,这并不能说明大径材良种培育中可以忽略母本 × 父本互作(特殊杂交力)的效应,因为,这些效应仍有一定程度的影响,尤其特定环境下或对生长之外的其他性状可能影响显著[9, 16]。
桉属树种中,对10年生以上性状(包括生长)遗传分析的报道不多,且材料均为纯种的自由授粉家系,如蓝桉(E. globulus Labill.)[5-6, 8, 15]、尾叶桉[7]和赤桉[23],因此,本研究基于尾叶桉 × 细叶桉杂种群体对此提供了有益的补充。树高、胸径和材积的遗传力一般较低,尾叶桉 × 细叶桉13、16年生生长的
$h_s^2 $ 值(0.17~0.23)与10年生及之前生长的相应估值[9]类似,均为桉属中报道的正常范围,如蓝桉在Latrobe试验点4、8、16年生胸径的狭义遗传力为0.15~0.29[5],在La Florida试验点3、11、17、23年生胸径的狭义遗传力为0.10~0.36[8];尾叶桉14年生树高和胸径的狭义遗传力分别为0.34和0.14[7],这也表明,桉属近缘杂种与纯种在生长性状上可能具有相似的遗传模式。早晚期生长的相关分析及主成分回归分析为尾叶桉 × 细叶桉早期选择提供了理论支持。类似地,蓝桉16年生与4年生胸径的加性遗传相关达0.95[5],23年生与3、11、17年生胸径的遗传相关分别为0.53、1.00、1.00[8],均表明3年生左右进行早期选择是可行的。采用主成分回归分析法估算了火炬松(P. taeda Linn.)[35]早期选择年龄,表明7年生胸径能够解释19年生胸径变异的88.13%。另一方面,尾叶桉 × 细叶桉晚期生长与木材基本密度和木质素含量的显著相关也为间接选择提供了可能。类似地,蓝桉16年生胸径与木质素含量呈显著的遗传负相关[6]、与基于Pilodyn检测的6年生木材密度呈显著正相关[5];但本研究中一些性状间的相关与之前的报道有异,如尾叶桉14年生树高和胸径与木材基本密度呈显著的遗传负相关[7],蓝桉16年生胸径与纤维素含量和S/G比例均呈显著的遗传相关[6],这种差异可能是由于不同树种、地点和年龄引起的[11, 16]。
-
尾叶桉 × 细叶桉13、16年生生长具有一定的杂种优势,具备大径材选育的潜力,这为下一步培育大径材良种(包括无性系)提供了备选材料。多年生生长性状持续受母本效应的显著影响,13、16年生时父本效应只对材积影响显著,杂交育种中需要重视优良母本和父本的选择。16年生与早期树高、胸径和材积遗传相关及回归分析均显示,在2.5年生时可解释16年生生长变异的66.2%~73.9%,表明尾叶桉 × 细叶桉大径材良种培育的早期选择可早至树龄2.5年时进行。晚期生长与10年生木材基本密度和木质素含量分别呈显著的正相关和负相关,对晚期生长的选择可间接提高木材基本密度,并降低木质素含量。
尾叶桉 × 细叶桉多年生生长及其与材性相关的遗传分析
Genetic Analysis on Multiple-year Growth Traits and Their Correlations with Wood Properties in Eucalyptus urophylla × E. tereticornis crosses
-
摘要:
目的 了解尾叶桉 × 细叶桉多年生生长的遗传规律,为大径材良种培育的早期选择年龄的确定提供可靠依据。 方法 对10株尾叶桉 × 10株细叶桉不完全析因交配产生的59个杂种组合13、16年生树高、胸径和材积采用混合线性模型估算母本、父本和母本 × 父本互作的方差分量,计算狭义遗传力,检测与早期生长及10年生材性性状间的表型相关和加性遗传相关,并基于遗传相关和主成分回归分析进行生长性状早期选择年龄的确定。 结果 尾细桉 × 细叶桉杂种组合13、16年生树高、胸径和材积均高于母本自由授粉家系,16年生时1个组合和47株单株的胸径达到≥28 cm的大径材标准;母本效应对2个年龄的树高、胸径和材积均显著,但父本效应仅对材积显著,母本 × 父本互作仅对13年生树高显著,表明母本对杂种生长的影响明显大于父本;2个年龄的树高、胸径和材积的狭义遗传力为0.17~0.23,均属中等偏低的水平;16年生树高、胸径和材积与之前年龄(0.5、1.5、2.5、4、7.5、10和13年生)的相应生长性状的表型相关和加性遗传相关均达0.01或0.001的显著水平(与1.5年生材积的相关除外),2.5年生开始加性遗传相关即较稳定。16年生与早期生长性状的回归分析显示:2.5年生能够分别解释16年生树高、胸径和材积变异的73.9%、71.9%和66.2%;13年生和16年生的生长与10年生木材基本密度均呈显著的正相关,而与木质素含量均呈显著的负相关(H16除外)。 结论 尾叶桉 × 细叶桉13、16年生生长具有一定的杂种优势,可用于选育大径材良种;生长性状持续受母本效应的显著影响,父本效应对晚期材积亦影响显著,需要重视优良杂交母本和父本的选择;早晚期生长显著相关,早期选择可在2.5年生时进行;晚期生长与10年生木材基本密度和木质素含量分别呈显著的正和负相关,可进行间接选择。 Abstract:Objective To understand the genetic mechanism underlying multiple-year growth in Eucalyptus urophylla × E. tereticornis crosses and provide reliable support for determining the early selection age in large-diameter-timber variety breeding. Method For 13- and 16-year-old tree height, breast-high diameter and volume of 59 crosses derived from an incomplete factorial mating between 10 E. urophylla females and 10 E. tereticornis males, a mixed linear model was used to estimate the variance components of female, male, and female × male interaction effects and calculate the narrow-sense heritability. Phenotypic and additive genetic correlations with earlier growth and 10-year-old wood properties were also analyzed. Genetic correlations and principal components regression (PCR) analysis were used to estimate early selection age for growth traits. Result Overall growth of crosses at ages 13 and 16 was better than the maternal open-pollinated families, and there were one cross and 47 individuals qualified for the large-diameter-timber criterion ≥28 cm. For all the three growth traits at both ages, the variance components of female, male, and female × male interaction were all significant, only significant for volume, and only significant for 13-year-old height, respectively. It indicated that the effect of female was larger than male. Narrow-sense heritability ranged from 0.17 to 0.23, being moderate to low levels of genetic control. Phenotypic and additive genetic correlations of 16-year-old height, diameter and volume with earlier (age of 0.5, 1.5, 2.5, 4, 7.5,10 and 13 years) counterparts were all positively significant except for 1.5-year-old volume, and the additive genetic correlations for 2.5 and higher years were notably stable. PCR analysis showed that 2.5-year-old growth could explain 73.9%, 71.9% and 66.2% of phenotypic variation in 16-year-old height, diameter and volume, respectively. Growth traits at both ages were significantly genetically correlated with 10-year-old wood basic density (positive) and lignin content (negative, except H16). Conclusion Certain degree of growth heterosis is present at ages 13 and 16 in E. urophylla × E. tereticornis crosses, indicating the potential of selection for large-diameter-timber varieties. Female effects on growth are consistently significant while male effect on volume is significant, demonstrating the necessity of selection of female and male parents in hybrid breeding. Both significant correlations between 16-year-old and earlier growth and around 70.0% of 16-year-old phenotypic variation explained by 2.5-year-old growth suggested the practical age of early selection at 2.5 years. Significant correlations of growth with 10-year-old wood basic density and lignin content imply the feasibility of indirect selection. -
Key words:
- E. urophylla × E. tereticornis
- / growth trait
- / heritability
- / genetic correlation
- / wood property
-
图 1 16年生树高(H16),胸径(D16)和材积(V16)与之前年龄相应生长的表型相关和加性遗传相关
Figure 1. Phenotypic and additive genetic correlations of 16-year-old height (H16), breast-high diameter (D16) and volume (V16) with earlier height, breast-high diameter and volume, respectively
表 1 尾叶桉 × 细叶桉杂种组合及母本自由授粉(OP)家系的生长及其变异系数(CV)
Table 1. Growth and coefficient of variation (CV) of Eucalyptus urophylla × E. tereticornis crosses and maternal open-pollinated (OP) families
生长性状
Growth trait杂种组合
Inter-specific cross自由授粉家系
OP family均值
Mean变异系数
CV/%平均胸径≥28 cm杂种组合数
The number of crosses≥28 cm in D13 or D16胸径≥28 cm单株数
The number of trees≥28 cm in D13 or D16均值
Mean变异系数
CV/%13年生 树高 H13/m 17.69* 26.8 16.60 28.5 胸径 D13/cm 15.89 43.5 0 29 14.58 41.2 材积 V13/m3 0.250*** 81.7 0.154 82.4 16年生 树高 H16/m 21.89 28.6 20.41 31.3 胸径 D16/cm 17.68 41.8 1 47 16.61 37.6 材积 V16/m3 0.377* 81.1 0.293 86.2 注:* P ≤ 0.05、*** P ≤ 0.001表示杂种组合与母本自由授粉家系的差异显著性水平分别达0.05、0.001。 Notes: * P ≤ 0.05, *** P ≤ 0.001 for the significant difference at 0.05 and 0.001, respectively, between inter-specific crosses and maternal OP families.  下载: 导出CSV
下载: 导出CSV
表 2 尾叶桉 × 细叶桉生长的母本(
${\color[RGB]{27,29,113}\sigma _f^2 }$ )、父本(${\color[RGB]{27,29,113} \sigma _m^2} $ )和母本 × 父本(${\color[RGB]{27,29,113} \sigma _{fm}^2 }$ )的方差分量及单株狭义遗传力(${\color[RGB]{27,29,113} h _s^2 }$ )和显性遗传效应(${\color[RGB]{27,29,113} d_s^2}$ )Table 2. Female (
${\color[RGB]{27,29,113} \sigma _f^2} $ ), male (${\color[RGB]{27,29,113} \sigma _m^2} $ ) and female × male (${\color[RGB]{27,29,113} \sigma _{fm}^2 } $ ) variance components as well as single-tree narrow sense heritability (${\color[RGB]{27,29,113} h _s^2} $ ) and dominance ratio of phenotypic variance (${\color[RGB]{27,29,113} d_s^2 }$ ) for growth of E. urophylla × E. tereticornis crosses生长性状
Growth trait方差分量 Variance component (±SE) 单株狭义遗传力
$h _s^2 $(±SE)显性遗传效应
$d _s^2 $(±SE)母本 $\sigma _f^2 $ 父本 $\sigma _m^2 $ 母本 × 父本 $\sigma _{fm}^2 $ 13年生 树高 H13 1.79 (±1.12)*** 1.84 (±0.12) × 10−6 0.85 (±0.60)* 0.17 (±0.10) 0.16 (±0.11) 胸径 D13 3.91 (±2.28)*** 0.52 (±0.63) 3.79 (±0.24) × 10−6 0.20 (±0.10) 3.37 (±0.18) × 10−7 材积 V13 2.91 (±1.78) × 10−3*** 1.17 (±0.93) × 10−3** 3.88 (±0.27) × 10−8 0.21 (±0.09) 3.91 (±0.20) × 10−5 16年生 树高 H16 3.17 (±2.03)*** 3.34 (±0.25) × 10−6 1.14 (±1.20) 0.17 (±0.10) 0.12 (±0.13) 胸径 D16 4.54 (±2.77)*** 1.09 (±1.10) 0.57 (±1.41) 0.22 (±0.11) 0.04 (±0.11) 材积 V16 4.96 (±3.72) × 10−3** 5.65 (±4.11) × 10−3** 2.58 (±3.41) × 10−3 0.23 (±0.11) 0.11 (±0.15) 注:SE,标准误;* P≤0.05,**P≤0.01,*** P≤0.001。下同。
Notes:SE,Standard error;* P ≤0.05,** P ≤0.01,*** P ≤0.001.The same below.
下载: 导出CSV
表 3 13、16年生生长与10年生材性的表型相关(rp)和加性遗传相关(rg)
Table 3. Phenotypic (rp) and additive genetic (rg) correlations of 13- and 16-year-old growth with 10-year-old wood properties
生长性状
Growth trait相关系数
rp or rg材性性状 Wood property trait (±SE) BD10 CC10 HC10 LC10 S/G10 H13 rp 0.630 (±0.035)** −0.072 (±0.062) 0.007 (±0.059) −0.102 (±0.059)* 0.204 (±0.064)** rg 0.606 (±0.225)** 0.072 (±0.337) 0.108 (±0.416) −0.505 (±0.307)* 0.061 (±0.311) D13 rp 0.633 (±0.035)** −0.126(±0.062)* −0.001 (±0.056) −0.137 (±0.056)** 0.243 (±0.064)** rg 0.537 (±0.252)* 0.082 (±0.333) 0.043 (±0.435) −0.604 (±0.298)* 0.024 (±0.238) V13 rp 0.550 (±0.040)*** −0.162 (±0.061)** 0.015 (±0.053) −0.151 (±0.054)** 0.244 (±0.066) *** rg 0.472 (±0.261)* 0.035 (±0.345) −0.142 (±0.363) −0.508 (±0.306)* −0.147 (±0.315) H16 rp 0.618 (±0.038)** −0.005 (±0.067) 0.016 (±0.062) −0.097 (±0.061)* 0.208 (±0.054)** rg 0.493 (±0.257)* 0.001 (±0.346) −0.007 (±0.424) −0.427 (±0.334) −0.043 (±0.314) D16 rp 0.630 (±0.036)** −0.135 (±0.065)* −0.019 (±0.059) −0.144 (±0.058)** 0.222 (±0.067) ** rg 0.474 (±0.261)* 0.093 (±0.360) 0.012 (±0.414) −0.559 (±0.299)* −0.089 (±0.315) V16 rp 0.560 (±0.043)*** −0.131 (±0.068)* −0.030 (±0.056) −0.152 (±0.054) ** 0.238 (±0.071)*** rg 0.568 (±0.227)** −0.123 (±0.327) −0.152 (±0.325) −0.500 (±0.264)* −0.057 (±0.313)
下载: 导出CSV
表 4 16年生与早期生长回归分析的第1主成分
Table 4. PC1 of principal components regression of 16-year-old growth with earlier growth traits
年龄
Age/a第1主成分 PC1/% 树高 H 胸径 D 材积 V 1.5 49.0 52.2 45.1 2.5 73.9 71.9 66.2 4 81.5 86.8 78.4 7.5 97.0 98.6 96.4 10 98.3 99.1 99.5 13 97.1 97.3 99.2 模型 Model 83.0 91.3 93.7
下载: 导出CSV
-
[1] Singh R K, Bhalerao R P, Eriksson M E. Growing in time: exploring the molecular mechanisms of tree growth[J]. Tree Physiology, 2021, 41(4): 657-678. doi: 10.1093/treephys/tpaa065 [2] 康向阳. 关于林木育种策略的思考[J]. 北京林业大学学报, 2019, 41(12):15-22. doi: 10.12171/j.1000-1522.20190412 [3] Grattapaglia D, Plomion C, Kirst M, et al. Genomics of growth traits in forest trees[J]. Current Opinion in Plant Biology, 2009, 12(2): 148-156. doi: 10.1016/j.pbi.2008.12.008 [4] Volker P W, Potts B M, Borralho N M G. Genetic parameters of intra- and inter-specific hybrids of Eucalyptus globulus and E. nitens[J]. Tree Genetics & Genomes, 2008, 4(3): 445-460. [5] Stackpole D J, Vaillancourt R E, de Aguigar M, et al. Age trends in genetic parameters for growth and wood density in Eucalyptus globulus[J]. Tree Genetics & Genomes, 2010, 6(2): 179-193. [6] Stackpole D J, Vaillancourt R E, Alves A, et al. Genetic variation in the chemical components of Eucalyptus globulus wood[J]. G3: Genes, Genomes, Genetics, 2011, 1(2): 151-159. [7] Hein P R G, Bouvet J-M, Mandrou E, et al. Age trends of microfibril angle inheritance and their genetic and environmental correlations with growth, density and chemical properties in Eucalyptus urophylla S. T. Blake wood[J]. Annals of Forest Science, 2012, 69(6): 681-691. doi: 10.1007/s13595-012-0186-3 [8] Salas M, Nieto V, Perafán L, et al. Genetic parameters and comparison between native and local landraces of Eucalyptus globulus Labill. ssp. globulus growing in the central highlands of Colombia[J]. Annals of Forest Science, 2014, 71(3): 405-414. doi: 10.1007/s13595-013-0342-4 [9] Chen S, Weng Q, Li F, et al. Genetic parameters for growth and wood chemical properties in Eucalyptus urophylla × E. tereticornis hybrids[J]. Annals of Forest Science, 2018, 75(1): 16. doi: 10.1007/s13595-018-0694-x [10] Yang H, Weng Q, Li F, et al. Genotypic variation and genotype-by-environment interactions in growth and wood properties in a cloned Eucalyptus urophylla × E. tereticornis family in southern China[J]. Forest Science, 2018, 64(3): 225-232. doi: 10.1093/forsci/fxx011 [11] MacDonald A C, Borralho N M G, Potts B M. Genetic variation for growth and wood density in Eucalyptus globulus ssp. globulus in Tasmania (Australia)[J]. Silvae Genetica, 1997, 46(4): 236-241. [12] Gan S, Li M, Li F, et al. Genetic analysis of growth and susceptibility to bacterial wilt (Ralstonia solanacearum) in Eucalyptus by interspecific factorial crossing[J]. Silvae Genetica, 2004, 53(5): 254-258. [13] 王建忠, 熊 涛, 张 磊, 等. 25年生大花序桉种源生长与形质性状的遗传变异及选择[J]. 林业科学研究, 2016, 29(5):705-713. doi: 10.3969/j.issn.1001-1498.2016.05.012 [14] Kien N D, Quang T H, Jansson G, et al. Cellulose content as a selection trait in breeding for kraft pulp yield in Eucalyptus urophylla[J]. Annals of Forest Science, 2009, 66(7): 711. doi: 10.1051/forest/2009064 [15] Stackpole D J, Vaillancourt R E, Downes G M, et al. Genetic control of kraft pulp yield in Eucalyptus globulus[J]. Canadian Journal of Forest Research, 2010, 40(5): 917-927. doi: 10.1139/X10-035 [16] He X, Li F, Li M, et al. Quantitative genetics of cold hardiness and growth in Eucalyptus as estimated from E. urophylla × E. tereticornis hybrids[J]. New Forests, 2012, 43(3): 383-394. doi: 10.1007/s11056-011-9287-3 [17] Callister A N, England N, Collins S. Genetic analysis of Eucalyptus globulus diameter, straightness, branch size, and forking in Western Australia[J]. Canadian Journal of Forest Research, 2011, 41(6): 1333-1343. doi: 10.1139/x11-036 [18] Madhibha T, Murepa R, Musokonyi C, et al. Genetic parameter estimates for interspecific Eucalyptus hybrids and implications for hybrid breeding strategy[J]. New Forests, 2013, 44(1): 63-84. doi: 10.1007/s11056-011-9302-8 [19] 彭仕尧, 徐建民, 李光友, 等. 尾细桉无性系在雷州半岛的生长与遗传分析[J]. 中南林业科技大学学报:自然科学版, 2013, 33(4):23-27. [20] Weng Q, He X, Li F, et al. Hybridizing ability and heterosis between Eucalyptus urophylla and E. tereticornis for growth and wood density over two environments[J]. Silvae Genetica, 2014, 63(1-2): 15-24. [21] Hung T D, Brawner J T, Roger M, et al. Estimates of genetic parameters for growth and wood properties in Eucalyptus pellita F. Muell. to support tree breeding in Vietnam[J]. Annals of Forest Science, 2015, 72(2): 205-217. doi: 10.1007/s13595-014-0426-9 [22] Zhu Y, Wu S, Xu J, et al. Genetic parameters for growth traits and stem-straightness in Eucalyptus urophylla × E. camaldulensis hybrids from a reciprocal mating design[J]. Euphytica, 2017, 213(7): 142. doi: 10.1007/s10681-017-1923-3 [23] Nezu I, Ishiguri F, Aiso H, et al. Repeatability of growth characteristics and wood properties for solid wood production from Eucalyptus camaldulensis half-sib families growing in Thailand[J]. Silave Genetica, 2020, 69(1): 36-43. doi: 10.2478/sg-2020-0006 [24] 董雷鸣, 张守攻, 孙晓梅. 日本落叶松全双列交配生长性状的遗传分析[J]. 林业科学研究, 2019, 32(4):11-18. [25] Belaber E C, Gauchat M E, Rodríguez G H, et al. Estimation of genetic parameters using spatial analysis of Pinus elliottii Engelm. var. ellittii second-generation progeny trials in Argentina[J]. New Forests, 2019, 50(4): 605-627. doi: 10.1007/s11056-018-9682-0 [26] Brooker M I H. A new classification of the genus Eucalyptus L'Hér. (Myrtaceae)[J]. Australian Systematic Botany, 2000, 13(1): 79-148. doi: 10.1071/SB98008 [27] FAO. Eucalypts for planting[M]. Rome: Food and Agriculture Organization of the United Nations, 1979. [28] Eldridge K, Davidson J, Harwood C, et al. Eucalypt domestication and breeding[M]. New York: Oxford University Press, 1993. [29] 何旭东, 李发根, 翁启杰, 等. 尾叶桉 × 细叶桉杂种生长和耐寒性的联合选择[J]. 中南林业科技大学学报:自然科学版, 2010, 30(8):68-71. [30] 陈升侃, 周长品, 翁启杰, 等. 尾叶桉 × 细叶桉木材密度与生长的联合选择[J]. 林业科学研究, 2018, 31(2):77-82. [31] Gilmour A R, Gogel B J, Cullis B R, et al. ASReml user guide release 3.0[M]. Hemel Hempstead: VSN International Ltd, 2009. [32] 林元震, 陈晓阳. R与ASRmel-R统计分析教程[M]. 北京: 中国林业出版社, 2014. [33] 陈少雄, 欧阳林男, 王军锋, 等. 中国桉树大径材培育与利用[M]. 北京: 中国林业出版社, 2020. [34] 刘奕清, 王大平. 尾细桉的组织培养和快速繁殖[J]. 西南农业大学学:自然科学版, 2005, 27(2):237-239. [35] 栾启福, 丁显印, 徐永勤, 等. 主成分回归分析在火炬松早期选择中的应用研究[J]. 安徽农业大学学报, 2019, 46(2):264-269. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3920
- HTML全文浏览量: 2315
- PDF下载量: 119
- 被引次数: 0


























