




-
磷素(P)是仅次于氮素(N)的第二大限制植物生长的大量营养元素[1]。P在陆地生态系统中的循环模式与N不同,土壤中的P主要是通过矿物岩石风化所得,且随着时间的推移逐渐被风化耗尽[2]。第四纪冰川作用使大部分温带和北部地区的新鲜基岩暴露,而在热带地区则并未发生[3]。生物地球化学理论表明,受到高度风化的热带森林主要受到P的限制[1,3]。研究表明,由于N和P的耦合关系,随着氮的不断输入导致土壤酸化效应增强,加快了土壤中铁铝氧化物对无机磷的吸附固定,从而使土壤磷的有效性降低[4]。在热带和亚热带地区高N沉降促进了P的循环,加剧了P对植物生长的限制[5]。
植物在外界环境发生改变时,会通过改变自身功能性状以反映自身对环境变化的响应。因此,植物功能性状在研究物种生活史策略上具有重要意义[6]。叶片和细根是植物地上和地下进行物质分配和交换的重要场所[7]。近年来,关于土壤磷素胁迫与植物叶片、细根功能性状的相关研究已有不少。Rao等[8]指出高磷胁迫降低了植物性状网络连通性和植物功能潜力,进而抑制植物生长。对湿地松(Pinus elliottii Engelmann)的施磷研究表明磷添加能促进叶片生长代谢活动和叶面积增加,同时加快叶干物质含量向其他器官运输,使叶片干物质含量减少[9]。Duan等[10]通过欧洲油菜(Brassica napus Linnaeus)不同程度的施磷实验,得出磷在生长介质中的分布决定了根系统构架。林强[11]通过研究施磷对芒萁(Dicranopteris dichotoma(Houttuyn)Nakaike)功能性状的影响中得出,磷添加显著提高了芒萁的叶面积、根组织密度,但对比叶面积、叶组织密度、比根长无显著影响。因此,通过研究植物功能性状对磷添加处理下的变化规律能够很好的了解植物对外界磷素添加的响应。
杉木(Cunninghamia lanceolata(Lamb.) Hook.)是我国南方主要的造林树种之一[12]。第九次全国森林资源清查数据显示,杉木人工林的面积为990.20万 hm2,蓄积量为75 545.01万m3,分别占全国人工乔木林总面积和总蓄积量的17.33%和22.30%[13]。纯林经营是目前大多数杉木人工林所采用的模式,纯林连栽使杉木人工林生产力降低、林地地力衰退以及生物多样性降低等问题日益突出[14]。速生期阶段(7~11年生)是杉木生长的重要时期[15-16]。本论文以10年生杉木人工林为对象,通过设置6个不同梯度的土壤磷添加控制试验,探讨杉木叶片、细根典型功能性状对3年土壤磷添加的响应及其相关性,以期为提高杉木人工林的生产力提供科学依据。
-
试验地位于中国林业科学研究院亚热带林业实验中心长埠实验林场(27°33′ N,114°35′ E),地处江西省分宜县境内,为亚热带低山丘陵地区,平均海拔150 m左右;属亚热带湿润气候,年平均气温18.0 ℃,极端高温39.9 ℃,极端低温− 10 ℃,年平均降水量1 600 mm,全年无霜期270 d。土壤为花岗岩发育而成的黄红壤。长埠实验林场共有山林面积1 113.57 hm2,其中科研示范林206.8 hm2、经济林40 hm2;现有活立木蓄积量约7.1万m3,主要植被类型为杉木人工林、油茶(Camellia oleifera Abel.)林、毛竹(Phyllostachys edulis(Carriere) J. Houzeau)林以及针阔混交林等。
-
2018年6月,选择3块坡向为西坡或西北坡、坡度在25°~30°、直线距离至少60 m以上、株行距为2 m × 3 m的10年生杉木人工林。一块杉木人工林为一个区组,并设置6个10 m × 10 m的样方,每个样方之间间隔距离至少20 m。施肥前测定各样方不同土层深度(0~10、10~30、30~50 cm)土壤的基本特征指标,样方本底土壤调查结果(表1)表明各土壤特征指标无显著空间变异。每个区组随机分为P0(对照)、P1、P2、P3、P4、P5 6个处理,施磷量[17]分别为0、60、120、180、240、300 kg·hm−2·a−1。于2018年7月开始人工施磷,频率[18]为每2个月1 次。每次将P0、P1、P2、P3、P4、P5处理所需要添加的分析纯NaH2PO4·2H2O 0、0.39、0.78、1.17、1.56、1.95 kg分别溶解于20 L水里,用背带式电动喷雾器均匀喷洒到每个处理对应样方的土壤表面。
土壤指标
Soil indicators土层 Soil layer/cm 0~10 10~30 30~50 平均值 Mean P 平均值 Mean P 平均值 Mean P pH 4.27 ± 0.23 0.308 4.33 ± 0.10 0.368 4.29 ± 0.19 0.267 有机质OM/(g·kg−1) 41.64 ± 5.11 0.590 19.96 ± 7.54 0.103 14.23 ± 5.37 0.951 全氮TN/(g·kg−1) 1.69 ± 0.22 0.258 0.93 ± 0.32 0.236 0.67 ± 0.26 0.693 全磷TP/(g·kg−1) 0.28 ± 0.03 0.677 0.24 ± 0.03 0.252 0.23 ± 0.05 0.204 全钾TK/(g·kg−1) 26.66 ± 2.36 0.168 24.57 ± 3.07 0.309 24.09 ± 3.66 0.297 铵态氮NH4 + −N/(mg·kg−1) 6.47 ± 1.41 0.555 3.96 ± 1.57 0.178 3.13 ± 0.67 0.989 硝态氮NO3−−N/(mg·kg−1) 1.06 ± 0.21 0.147 0.79 ± 0.25 0.159 0.69 ± 0.29 0.662 有效磷AP/(mg·kg−1) 0.96 ± 0.24 0.187 0.43 ± 0.15 0.397 0.29 ± 0.13 0.476 注:OM:有机质;TN:全氮;TP:全磷;TK:全钾;NH4+-N:铵态氮;NO3−-N:硝态氮;AP:有效磷;n = 18。
Notes: OM: Organic matter; TN: Total nitrogen; TP: Total phosphorus; TK: Total potassium; NH4+-N: Ammonium nitrogen; NO3−-N: Nitrate nitrogen; AP: Available phosphorus; n = 18.Table 1. Basic characteristics of soil samples before P addition (mean ± SD)
-
2021年7月,基于对每个样方杉木的胸径调查数据选取3株标准木,用高枝剪采集冠层中部阳面相似高度枝条1~2根,在枝条上采取当年生健康且成熟的叶片;从标准木根部的3个方位(以树干为圆心的圆的三等分线上)进行细根采集,由于林内根系错综复杂,为确保细根取样准确,采用先找树木主根,挖取3~5条完整的根系,在根系中挑选出直径 ≤ 2 mm的活细根[19]。所采集的叶片、细根分别用于测定叶片、细根功能性状。每个样方3个重复,即每个处理共9个重复。
-
从每株标准木选择30片当年生健康完整且成熟的新鲜叶片,用精度为0.01 mm的电子游标卡尺在叶片的非叶脉前、中、末端分别测量叶片厚度,取算术平均值为该叶片的厚度(LT);用Epson Perfection V370 Photo扫描仪扫描每个叶片,并用ImageJ V1.52(National Institutes of Health,USA)计算叶面积(LA);测量完后将叶片样品浸泡在清水中至恒质量,然后用吸水纸吸去叶片表面多余的水分,利用电子天平(0.01 g精度)称量饱和鲜质量(LSFW),随后将样品放入烘箱中,在65 ℃下将叶片样品烘干至恒质量后称量干质量(LDW)。叶片功能性状的计算公式[20]:
式中:SLA为比叶面积;LTD为叶组织密度;LDMC为叶干物质含量。
-
用Epson Perfection V370 Photo扫描仪扫描直径 ≤ 2 mm的细根,并使用RhizoVision explorer v2.0.3(Noble Research Institute, LLC,USA)根系构型分析软件处理图像,获取细根的根长(RL)、细根表面积(RA)、细根体积(RV)等数据。扫描后的根系在 65 ℃下烘干至恒质量后称干质量(RDW)。细根功能性状的计算公式[19]如下:
式中:SRL为比根长;SRA为比根表面积;RTD为细根组织密度。
各个功能性状的可塑性指数(PI)参考Valladares等[21]的方法计算,公式如下:
式中:max(i)为功能性状在某处理下的最大值,min(i)为功能性状在某处理下的最小值。
变异系数(CV)的计算公式为:
式中:SD为功能性状在不同处理下的标准差,mean为功能性状在不同处理下的平均值。
-
采用Microsoft Excel 2016和SPSS 19.0软件(SPSS Inc.,Chicago,IL,USA)对数据进行统计分析,利用Origin 2019(OriginLab Corporation,Northampton,MA,USA)软件作图。土壤不同梯度P添加下杉木叶片和细根功能性状的差异采用SPSS 19.0软件的单因素方差分析(one way ANOVA)检验,并进行Duncan多重比较法进行不同处理之间差异性检验分析(P < 0.05),用Pearson相关分析检验土壤不同梯度P添加下各功能性状的相关性。
-
由图1A可知:随着施P量的增加,杉木叶面积(LA)、比叶面积(SLA)均先增大后减小,其中,P3处理的LA最大(1.17 cm2),P2处理的SLA最大,除P2处理的SLA外,P添加处理的LA 、SLA均与其它处理差异不显著。由图1B可知:随着施P量的增加,杉木叶组织密度(LDT)、叶干物质含量(LDMC)均呈先降低后升高的趋势。P2、P3、P4处理的LTD比P0分别显著降低了33.3%、25.7%、32.2%(P < 0.05),P1、P5处理的LTD与P0差异不显著;P2、P3处理的LDMC比P0分别显著降低了7.6%、10.4%(P < 0.05);而P1、P4、P5处理下的LDMC与P0差异不显著;P1、P2、P3、P4、P5处理间的LTD、LDMC均差异不显著(P > 0.05)。
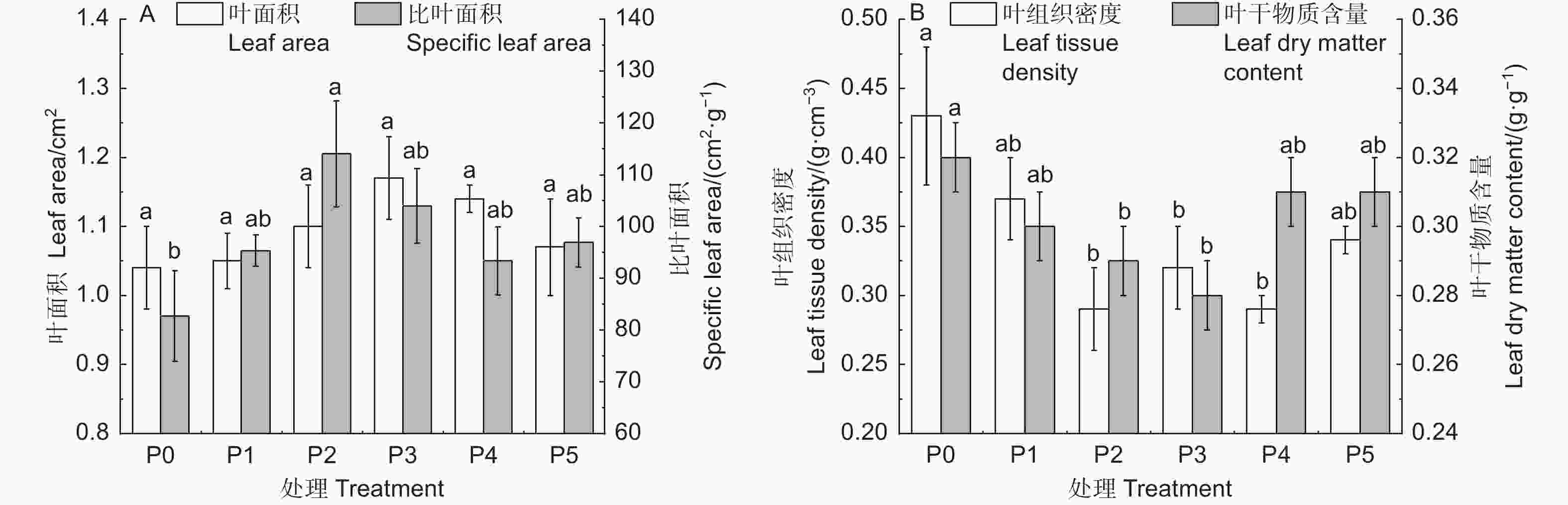
Figure 1. Response of leaf functional traits to soil different gradient phosphorus addition in Cunninghamia lanceolata (mean ± SE)
-
杉木比根长(SRL)随施P量的增加呈先增大后减小的趋势,但不同处理间差异不显著(图2A)。杉木根组织密度(RTD)随施P量的增加呈先降低后增大的趋势,不同梯度P添加处理的RTD与P0相比均差异不显著(P > 0.05);P2、P3处理的RTD显著低于P5处理(P < 0.05);P1、P4处理下的RTD与P2、P3、P5处理间均差异不显著(图2B)。杉木比根面积(SRA)随施P量的增加呈先增加后降低的趋势,不同P添加处理的SRA与P0均差异不显著(P > 0.05);P3处理的SRA最大,为183.95 ± 7.39 cm2·g−1,显著高于P4、P5处理的SRA(P < 0.05);P2处理的SRA显著高于P4处理的(P < 0.05); P1处理下的SRA与P2、P3、P4、P5处理的均差异不显著(图2C)。
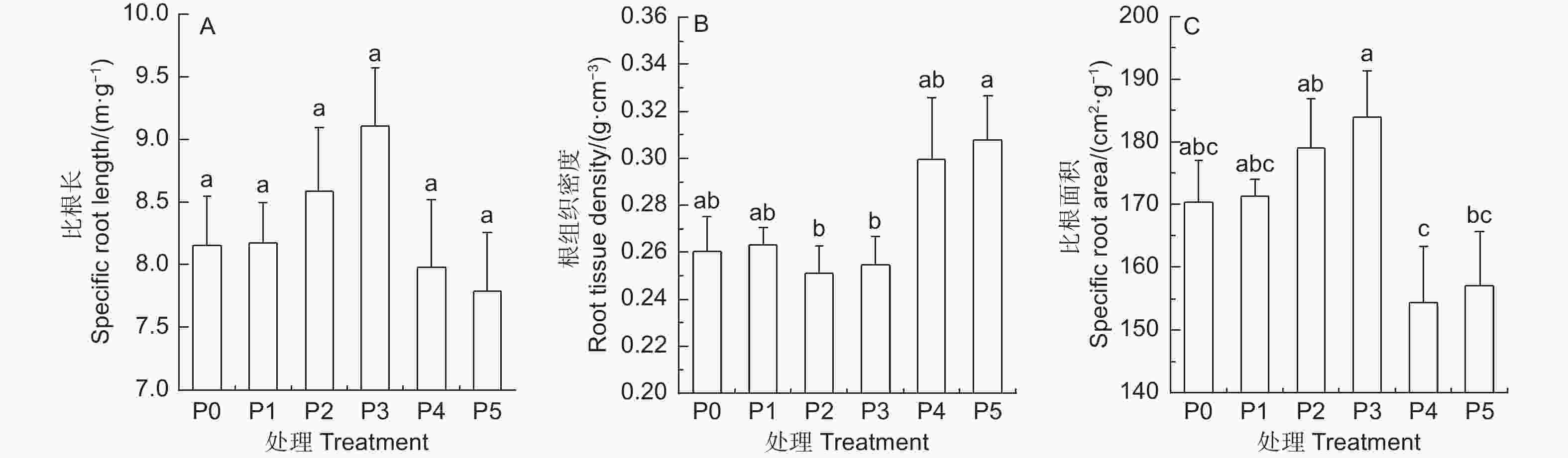
Figure 2. Response of root functional traits to soil different gradient phosphorus addition in Cunninghamia lanceolata (mean ± SE)
-
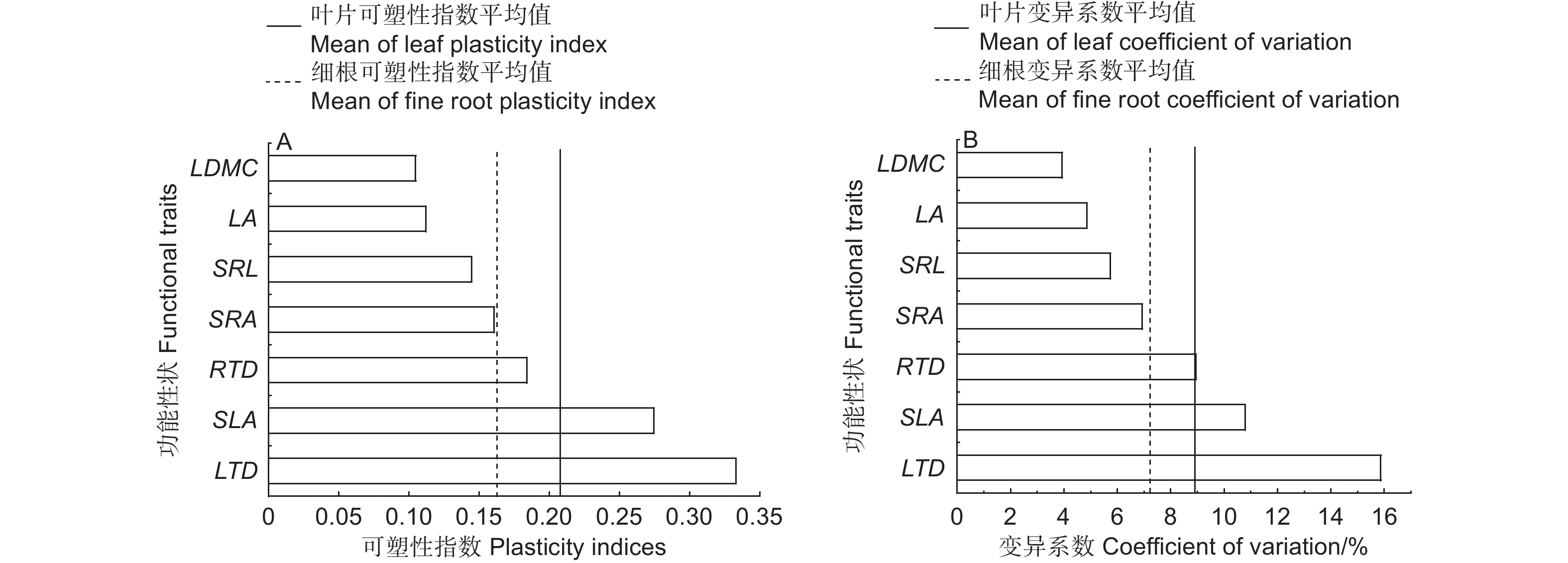
可塑性指数(PI)计算结果显示:叶片功能性状的可塑性指数平均为0.21,其中,LTD的PI最大,为0.33,其次依次为SLA、LA、LDMC,其PI值分别是0.27、0.11、0.10;细根功能性状的可塑性指数平均为0.16,其中,RTD的PI最大,为0.18,其次为SRA和SRL,PI值分别为0.16、0.14(图3A)。叶片和细根功能性状的变异系数(CV)大小排序与PI一致,叶片功能性状的变异系数平均为8.9%,LTD、SLA、LA、LDMC的CV分别为15.9%、10.8%、4.9%、3.9%;细根功能性状的变异系数平均为7.2%,其中,RTD的CV最大,为8.9%,其次为SRA和SRL,CV分别为6.9%、5.7%(图3B)。
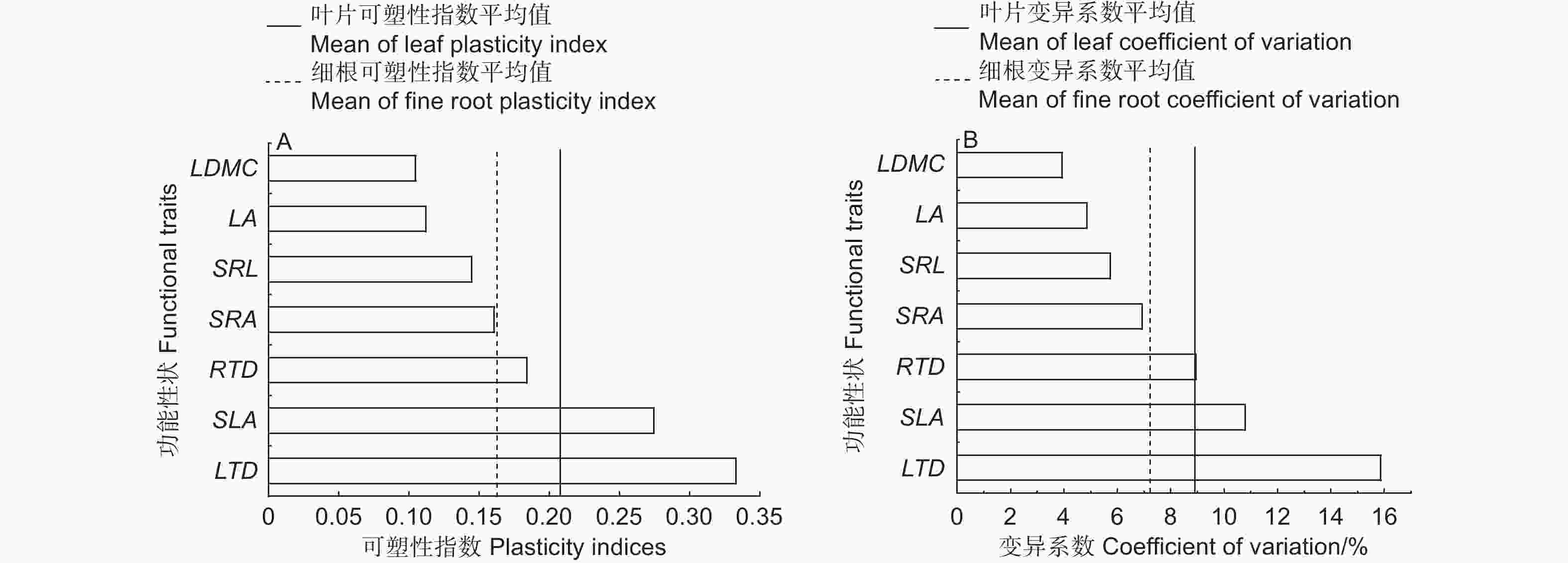
Figure 3. Plasticity index (PI) and coefficient of variation (CV) of functional traits in leaf and fine root of Cunninghamia lanceolata under soil different gradient phosphorus addition
-
由表2可知:杉木叶片、细根功能性状间的相关性在土壤不同梯度P添加下的响应不同。在P0处理下,LTD与LA、SLA呈显著负相关(P < 0.05);LA与SLA呈显著正相关(P < 0.05);SRA与SRL呈极显著正相关(P < 0.01),与RTD呈显著负相关(P < 0.05)。在P1处理下,除LDMC与LTD呈显著正相关(P < 0.05)外,其它功能性状间无显著相关性。在P2处理下,LTD与LA呈显著负相关(P < 0.05),与SLA呈极显著负相关(P < 0.01);SRA与RTD呈极显著负相关(P < 0.01)。在P3处理下,SLA与LA呈显著正相关(P < 0.05);RTD与SRL、SRA呈极显著负相关(P < 0.01);SRA与SRL呈极显著正相关(P < 0.01)。在P4处理下,SLA与LDMC呈极显著负相关(P < 0.01);SRA与SRL呈极显著正相关(P < 0.01),与RTD呈极显著负相关(P < 0.01);RTD与SRL呈显著负相关(P < 0.05);在P5处理下,SRA与SRL呈极显著正相关(P < 0.01),与RTD呈极显著负相关(P < 0.01)。
处理
Treatment功能性状
Functional traitLA LTD SLA LDMC SRL RTD P0 LTD −0.796* SLA 0.726* −0.792* LDMC −0.365 0.621 −0.072 SRL 0.397 −0.314 0.650 0.442 RTD −0.325 0.195 −0.339 −0.135 −0.263 SRA 0.450 −0.300 0.590 0.400 0.837** −0.734* P1 LTD 0.202 SLA 0.033 −0.578 LDMC −0.113 0.677* −0.530 SRL −0.432 −0.413 0.338 −0.323 RTD −0.595 0.034 −0.451 0.345 −0.204 SRA 0.166 −0.168 0.630 −0.087 −0.026 −0.168 P2 LTD −0.713* SLA 0.627 −0.881** LDMC −0.039 0.280 −0.488 SRL −0.616 0.587 −0.493 0.158 RTD 0.261 −0.560 0.405 −0.366 0.156 SRA −0.394 0.655 −0.592 0.330 −0.130 −0.874** P3 LTD −0.337 SLA 0.678* −0.604 LDMC −0.386 0.200 −0.536 SRL −0.401 −0.046 0.000 −0.022 RTD 0.368 −0.138 0.095 0.013 −0.913** SRA −0.396 0.011 −0.040 0.025 0.977** −0.974** P4 LTD −0.342 SLA 0.558 −0.299 LDMC −0.505 0.325 −0.855** SRL −0.042 −0.036 −0.257 0.118 RTD −0.125 0.234 0.297 −0.317 −0.717* SRA 0.020 −0.130 −0.332 0.214 0.945** −0.894** P5 LTD 0.220 SLA −0.363 0.110 LDMC 0.480 −0.169 −0.314 SRL −0.322 0.125 −0.113 0.176 RTD −0.033 0.255 −0.001 −0.230 −0.654 SRA −0.197 −0.043 −0.002 0.183 0.910** −0.902** 注:Pearson相关分析,双侧检验;** 表示P < 0.01,* 表示P < 0.05。
Notes: Pearson correlation analysis, two-tailed teat; ** means P < 0.01, * means P < 0.05.Table 2. Pearson correlation coefficients among functional traits
-
植物对生境资源的利用效率经常用比叶面积(SLA)进行衡量[22],SLA越大,表明植物叶片能够吸收更多的光能量,进而提高植物光合作用效率,使植物体内的资源流动速率加快[23],有利于提高植物的相对生长速率以促进植物对外界资源的获取。本研究中,在P0和P1处理下,杉木的生长主要受土壤P有效性的限制,较小的叶面积(LA)、比叶面积(SLA)能够减少植物蒸腾、降低植物的光合作用效率,使植物体内的资源流动速率变慢,从而降低了植物的相对生长速率,植物体内的养分大多数用来维持其基础生理活动。在P2处理下,杉木的SLA显著高于P0,因为在中磷水平处理下土壤养分逐渐改善,提高了土壤中可利用P的含量,促进了杉木对土壤P的高效利用,从而有利于杉木的生长发育,其LA、SLA的增大促使杉木针叶捕捉光资源能力提高,光合作用增强,这与贾婷等[9]对针叶树种湿地松的P添加研究结果一致;而在高P处理下,杉木的LA、SLA呈现出减小的趋势,说明杉木的生长受到了一定程度的胁迫。
叶干物质含量(LDMC)的大小反映了植物积累养分元素的能力[24]。本研究中,杉木的叶组织密度(LTD)、LDMC在P2、P3处理下显著低于P0,在P4、P5处理下,LTD和LDMC则呈增加趋势。在低磷情况下,较高的LDMC、LTD能够反映出植物把合成的干物质更多的用来构建叶片,进而通过增大叶肉密度和降低叶片含水量以增加叶片内的水分向叶片表面移动的距离或阻力[25],降低代谢速率,以维持叶片对资源的利用率和增加对营养物质的积累[26],满足杉木在土壤低P状况下的正常生长。在P2、P3处理下杉木代谢增强,LDMC低,表明植物降低了对叶片构建的投入比例,LTD较低,叶片内活动更强,植物通过吸收更多养分来投入到植物生长,增加了植物对养分的消耗,因此植物叶片对养分的储存能力降低,这与程雯等[27]的研究结果一致。刘旻霞等[28]提出,在养分充足的土壤环境下,植物光合能力较强,生长速率快,但植物本身对资源的捕捉利用能力会下降,这与本研究的结论一致;而在高P处理下,杉木的LTD、LDMC增加,进一步说明杉木在土壤高P条件下受到了胁迫。
-
比根长(SRL)和比根面积(SRA)是衡量细根对土壤中养分吸收效率的重要指标[29],对根的功能都起到关键作用,并对生态系统生产力有重要影响[30]。较低的比根组织密度(RTD)表明细根具有较高的水分和营养吸收效率[31]。本研究结果显示,在不同梯度P添加处理下,SRL无显著差异,说明杉木细根在环境变化中具有一定的内稳性;RTD呈先减小后增大的趋势,SRA则呈先增大后减小的趋势。低磷(P0、P1)水平下,较低的SRA和较高的RTD说明在低磷胁迫下杉木对土壤养分的吸收利用和运输受到抑制,这与于姣妲等[32]研究结论一致。当土壤磷素缺乏时,磷添加会促进植株养分吸收率和生长[33];当土壤P的供应充足,P不再是植物生长发育的限制条件时,过量的P添加使植物功能性状的潜力下降,进而抑制器官之间的有效资源运输,并且改变了植物体内其它元素的权衡比例,导致植物代谢紊乱,促使植物呼吸作用增强,大量糖分和能量被植物消耗,抑制植物生长[8]。本研究进一步验证了高P处理会抑制杉木生长的相关结论。
-
植物性状的可塑性表达在植物适应环境过程中发挥重要作用[34],在环境变化下植物不同部位性状的可塑性响应程度不同[35]。本研究中,杉木叶片和细根功能性状的可塑性指数(PI)范围分别为0.10~0.33、0.14~0.18,叶片功能性状的可塑性指数平均值高于细根功能性状的相应值,说明在土壤P添加下,杉木通过充分利用生境资源(尤其是对光资源的捕获与吸收)以加快植物的地上部分生长,而杉木地下部分可能有自身的生物学特性,在土壤P添加处理下,杉木细根获取营养物质的能力比叶片相对稳定,具有较强的环境适应能力。
遗传因素和外界环境的共同影响决定了植物的功能性状[36],而揭示植物对外部环境养分的响应和变化规律最方便的途径是通过对外部环境条件的改变来改变植物的功能性状。本研究中,土壤不同梯度P添加下,杉木的叶片和细根功能性状的变异系数(CV)范围为3.9%~15.9%,存在不同程度的变异,但CV均 < 20%,属于较弱变异[37],说明杉木叶片和细根功能性状指标存在一定的内稳性。何斌等[38]、贾婷等[9]研究马尾松(Pinus massoniana Lambert)人工林针叶功能性状、湿地松针叶功能性状等针叶树种中得到同样的结论。
-
植物对环境变化的响应不能只通过单一的性状描述,应充分考虑分析各个性状之间的耦合关系以揭示不同环境中植物对资源的分配和权衡策略[6]。本研究中,杉木LTD与LA、SLA在P0、P2处理下呈显著或极显著负相关,与LDMC在P1处理下呈显著正相关;SLA与LA在P0、P3处理下呈显著正相关,与LDMC在P4处理下呈极显著负相关,Jiang等[39]、Chang等[40]的研究中也得到过相似的结论。叶片功能性状在不同处理下的相关性规律说明在养分充足条件下,植物叶面积增大使植物接受光照的面积增大,进而增强植物对光的捕获能力,光合效率增加,加快了植物叶片的生理活动,降低叶肉密度,把更多资源投入到植物的生长中,进而降低了植物叶片对养分的积累,研究结果与贾婷等[9]研究结果一致。杉木SRL与SRA在P0、P3、P4、P5处理下呈极显著正相关,与RTD在P3、P4处理下呈显著或极显著负相关;RTD与SRA在P0、P2、P3、P4、P5处理下呈显著或极显著负相关,与Ning等[41]、王钊颖等[42]的研究结论相似。细根功能性状在不同P添加处理下的相关性规律说明当植物把养分更多的用于加速植物生长时,植物会增大细根长度和表面积来提高对土壤养分的高效率吸收,并降低细根组织密度来加快水分和营养的流动速率,这与李淑英[19]的研究结果一致。杉木叶片、细根功能性状间的相关性表明植物会通过各性状间的耦合关系来合理的对资源进行分配和权衡,以形成适应当前生境的形态指标组合。
-
在土壤不同梯度P添加处理下,杉木通过协调自身叶片和细根功能性状的耦合关系,以响应不同的环境。在土壤P缺乏和P过量条件下,杉木在自身结构性状方面通过减小LA、SLA、SRL、SRA,增加LTD、LDMC、RTD保证有效的利用资源进而适应逆境条件;当土壤的有效P达到一个充足的状态时,杉木通过增加LA、SLA以提高针叶捕捉光资源的能力;增加SRL、SRA来高效吸收土壤养分,使杉木在养分充足的条件下更好生长。因此,土壤施磷量在P2、P3处理下时有利于10年生杉木人工林的生长,为提高杉木人工林的生产力提供一定的科学参考。
Leaf and Fine Root Functional Traits Response of 10-Year-Old Cunninghamia lanceolate Plantations to Soil Phosphorus Addition
- Received Date: 2022-01-04
- Accepted Date: 2022-03-22
- Available Online: 2022-08-20
Abstract:

 DownLoad:
DownLoad: